
Abstract
Acute kidney injury (AKI) is a common clinical symptom, which is mainly manifested by elevated serum creatinine and blood urea nitrogen levels. When AKI is not repaired in time, the patient is prone to develop chronic kidney disease (CKD). The kidney is composed of more than 30 different cells, and its structure is complex. It is extremely challenging to understand the lineage relationships and cell fate of these cells in the process of kidney injury and regeneration. Since the 20th century, lineage tracing technology has provided an important mean for studying organ development, tissue damage repair, and the differentiation and fate of single cells. However, traditional lineage tracing methods rely on sacrificing animals to make tissue slices and then take snapshots with conventional imaging tools to obtain interesting information. This method cannot achieve dynamic and continuous monitoring of cell actions on living animals. As a kind of intravital microscopy (IVM), two-photon microscopy (TPM) has successfully solved the above problems. Because TPM has the ability to penetrate deep tissues and can achieve imaging at the single cell level, lineage tracing technology with TPM is gradually becoming popular. In this review, we provided the key technical elements of lineage tracing, and how to use intravital imaging technology to visualize and quantify the fate of renal cells.
Similar content being viewed by others


Introduction
The kidney is an extremely complex heterogeneous organ, composed of an organized vascular system and well-defined epithelial components, which are highly coordinated to maintain normal kidney function [1,2,3]. It is a large challenge to understand the lineage relationship between these cells and the fate map. Furthermore, owing to the technological limitation, observing the behavior of cells in vivo is always out of reach, leaving the mechanism underlying renal tissue regeneration invisible. The use of genetically modified mice for lineage tracing offers a forceful instrument for tracking cell dynamics in vivo [4]. Previously, lineage tracing was mainly used in developmental biology, but now it is also applied to stem cell research as it can provide information about the number, location, and differentiation state of progenies of interest cells [5] and their behavior in the intact kidneys [6,7,8,9], leading to an increasing attention from researchers across a wide range of scientific disciplines.
Although lineage tracing techniques provide us with a lot of information about tissue development and regeneration, how to accurately interpret the results remains to be solved [10, 11]. Advances in tracking or tracing approaches from light microscopy to two-photon microscopy (TPM), combined with lineage tracing strategies, have allowed researchers to decipher the cellular behavior and fate of various biological processes in the kidney [12]. Owing to TPM can directly visualize the dynamic changes of tissue morphology at subcellular resolution in real time in live animals, TPM imaging becomes the preferred experimental technique for studying kidney physiology and diseases over time [13].
Here, we will describe genetic lineage tracing technologies and novel intravital imaging approaches, aiming to outline the existing technical strategies and to elaborate the application of these technologies in kidney development, injury, and regeneration as well as future development prospects. Specifically, we summarize the basic principles and advantages of TPM and the application examples in kidney live imaging technology.
Lineage tracing technology
Cell tracking is a relatively general concept, which refers to labeling and tracking cells. The exploration of cell tracing technology originated in the early 20th century. As early as 1905, Conklin et al. used the characteristics of the coloring difference of the early split bulbs of ascidian embryos to observe the splitting process of split bulbs [14]. Since then, researchers have tried to use water-soluble dyes, fat-soluble dyes, peroxidase, and fluorescent dyes, etc., to physically inject them into cells to track them. Although these technical methods solved some of the problems at the time, there are still many defects in the present view. First of all, the physical method cannot guarantee the accurate labeling of target cells. Secondly, the labeling dye will be gradually diluted until it disappears during the cell division process, and traditional labeling methods cannot effectively permanently label individual cells and their progeny cells. However, due to the backwardness of microscopy equipment at that time, researchers were unable to clearly and continuously track the entire process of cell division [12, 15].
With the maturity of genetic engineering technology, genetic lineage tracing using gene targeting technology has gradually developed. In the application of tracer markers, endogenous genetic markers are used instead of exogenous markers. This type of labeling occurs at the genetic level, enabling long-term labeling of cells and permanent tracing of all their progeny. Lineage tracing techniques require in vivo labeling of specific tissues and cell types at specific times, and site-specific recombinase (SSR) systems with gene targeting technology can meet this need. The SSR has two main members: the Cre (Cyclization Recombination Enzyme)/Loxp (locus of X over P1) recombinase system from Escherichia coli phage P1 [16, 17] and the FLP (Flippase)/FRT (Flippase recognition target) system from Saccharomyces cerevisiae [18]. The two systems are similar in principle and both cut and connect DNA at specific sites. However, the Cre/Loxp recombinase system has higher recombination efficiency than the FLP/FRT system.
Cre/Loxp recombinase system
The concept of genetic lineage tracing relies on a switch, which is usually a drug-regulated Cre recombinase, which activates the downstream reporter gene. Cre was found in bacteriophage P1 by Sternberg et al., which recognizes a specific 34 bp nucleotide sequence known as Loxp [19, 20]. Generally, Cre mouse line is hybridized with reporter mice carrying fluorescent proteins or enzymes in a ubiquitous promoter, such as the Rosa26 locus [21, 22]. The expression of the reporter genes is blocked by a strong transcriptional stop sequence flanked by two Loxp sites [22]. After the Cre recombinase is activated, the stop cassette is excised, turning the reporter ON in a way specific to the cell type. On condition that the deletion of the stop sequence is permanent, the reporter gene will be expressed in all descendants of that cell [6, 23,24,25].
Compared with conventional cell tracking methods, the Cre/Loxp system has a good targeting ability and can only occur in cells that express tissue-specific promoters, largely avoiding the problem of mislabeling. The modified genome and genetic characteristics are the biggest advantages of this system. During development, Cre-expressing cells and all their descendants will be permanently labeled.
Taking advantage of the widely used Cre/Loxp system, the researchers designed a genetic strategy called “Brainbow” [26, 27]. This Brainbow transgenic mouse can randomly express a variety of fluorescent proteins (XFPs) under Cre-mediated recombination [27, 28]. The first generation of brainbow mice (Braindow-1) used Cre-mediated excision of fragments between Loxp sites to induce recombination events [27]. In Brainbow-2, Cre reverses the DNA fragments defined by the LoxP site and connects them in opposite directions to produce multiple recombination results [27, 28]. The R26R-Confetti mice were created to study the fate map of intestinal stem cells [9]. Clevers and colleagues combined the original brainbow2.1 cassette with the strong CAGG promoter and the LoxP site at Rosa26. After Cre recombination, this R26R-confetti heterozygous mouse showed random recombination of four different fluorescent proteins (nGFP, YFP, RFP, or mCFP) [9, 10]. The ‘Brainbow’, ‘Rainbow’, or ‘Confetti’ mouse models have become a powerful instrument for lineage tracing and has been developed for this purpose [22]. Under the control of a specific promoter, a multicolor reporter gene that randomly generates a large number of fluorescent protein co-expression has been confirmed to be a strategy for dynamic analysis of cell fate and tissue generation. Elena used it will induce this strategy, Bowman et al. used Rosa26-mT/mG mice and Axin2CreERT2 mice to demonstrate the important role of Wnt/β-catenin signaling pathway in neural stem cell function and homeostasis throughout the developmental process [29]. Lazzeri et al. tracked tubule cells in conditioned Pax8/Confetti mice and showed that despite the massive loss of renal tubule cells, renal function was restored after AKI. A small portion of Pax2+ tubular progenitor cells expand under the injury stress state and regenerate the necrotic tubules [30].
However, this feature also made it impossible to control the labeling time. To address the time specificity of labeling, Metzger et al. created a new mouse model based on Cre/Loxp [31]. This model takes advantage of the nature of estrogen receptors as nuclear receptors, thereby could regulate the time of Cre in the nucleus. Cre fused to a modified form of human estrogen receptor (CreERt) that does not bind estradiol but has affinity to tamoxifen. In the absence of tamoxifen, Cre cannot enter the nucleus due to estrogen receptors remaining in the cytoplasm, instead Cre enters the nucleus via estrogen receptors only under the condition of exogenous tamoxifen stimuli, accomplishing the recombinant modification of Loxp fragments [32,33,34]. Therefore, tamoxifen delivery can achieve the time specificity of the knockout event (Fig. 1a).

Lineage tracing of Lgr5+ or Sox9+ nephron stem/progenitor cells using the lacZ or GFP inducible genetic reporter system. a First, two transgenic mice were constructed, one was inserted with the Cre gene after the cell type or tissue-specific (TS) promoter, and the other was inserted with a reporter gene, such as GFP, after the promoter of ROSA26, and the expression of the reporter gene was stopped by the STOP sequence flanked by Loxp sites. When the mice of the two transgenic lines are crossed, the Cre enzyme will excise the STOP sequence and the reporter gene GFP will be successfully expressed in a specific cell or tissue [4]. By fusing the Cre gene to the tamoxifen-reactive hormone-binding region of the estrogen receptor (ER), a temporal restriction of expression can be achieved. In the absence of tamoxifen (TAM), Cre is inactive. After administration of tamoxifen, Cre is activated, transported from the cytoplasm to the nucleus, and mediates recombination between LoxP sites. As a result, the STOP codon is excised and the cells are permanently labeled with the reporter gene. The red arrows represent Loxp sites. b Schematic diagram of the nephron structure derived from Lgr5+ cells [35]. c Graphic abstract of the nephron structure derived from Sox9 positive cells [36]. Reproduced with permission from Ref. [35, 36]
Lineage tracing in kidney
The use of multicolor reporters can realize the labeling of multiple cell lineages in a specific tissue, so that different types of cells and their progeny can be easily distinguished and imaged by the difference of colors [37]. Recently, combined with time-controlled CreER lines and fluorescent reporter mice, lineage tracing studies have uncovered potential particulars and mechanisms in kidney development, homeostasis, and disease. Specifically, this technique is often used to study several developmental or injury-related renal progenitor cell populations and their lineages, including glomerular epithelial cells (e.g., Pax2 lineage [38]), proximal tubular cells (e.g., Sox9 [5, 36, 39] and SLC34a1 [40]), distal nephrons (e.g., Lgr5 [35]), and collecting ducts (e.g., p63 [41]). Below, we briefly introduce a few typical examples of kidney lineage tracing.
A good example is the research of Lgr5-positive stem cells in the kidney. Muscle, placenta, spinal cord, and other tissues express Lgr5. More specifically, in the intestine, kidney, stomach, ear, hair follicle, and liver, Lgr5 is considered a biomarker of adult stem cells [42]. In the year of 2012, Barker et al. used genetic lineage tracing technology to prove that during kidney development, Lgr5-positive cells are defined as progenitor cells in the kidney, which can develop into distal convoluted tubules and thick ascending limbs of Henle’s loop [35, 43]. Specifically, they crossed the Lgr5-EGFP-IRES-CreERT2 mouse lineage with Rosa26-Lacz reporter mice and gave tamoxifen at P1 to activate the Cre enzyme [35]. After Cre is activated, it enters the nucleus to initiate the expression of the lacZ reporter gene in cells expressing Lgr5 in the kidney [35]. Through lineage tracing, they observed that 1 week after the injection of tamoxifen, the progeny cells differentiated from lacZ-positive cells could proliferate rapidly to form a tubular structure [35]. Additionally, to explore the contribution of Lgr5-positive cells to the structure of the kidney during the development of the mouse kidney, they conducted a year-long induction [35]. Collectively, all these findings demonstrate that Lgr5, as the target of the Wnt pathway, is a biomarker gene of a nephron stem/progenitor population. It contributes to regenerate the thick ascending limb of Henle’s loop and distal convoluted tubules in the developing kidney [35] (Fig. 1b).
Another important application of genetic lineage tracing is to determine the epithelial hierarchy of the kidney during AKI and CKD. Sox9 is a transcription factor belonging to the Y box family of sex-determining regions and plays a pivotal role in the development of various tissues and organs including the kidney [44,45,46]. Studies have shown that mutations in Sox8 and Sox9 cause serious renal dysplasia in mice [36, 39]. In a recent study, researchers took advantage of Sox9-CreERT2 transgenic mice to confirm stem/progenitor cells in the kidney [36]. After kidney injury, Sox9-positive cells are activated and can repair damaged proximal renal tubules (Fig. 2c). In summary, the Sox9-expressing cells isolated from kidney tissue exhibited strong rapid expansion ability and progenitor-like ability in vitro. In vivo lineage tracing experiments showed that Sox9-positive cells were involved in epithelial regeneration after injury [36].

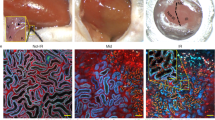
Cre-mediated dual fluorescent (mT/mG) reporter transgenic for intravital lineage tracing in the kidney. a After Cre-mediated recombinant modification, mT/mG mice were transformed from the expression of tdTomato to EGFP. In addition, both tdTomato and EGFP expressed by the mouse have cell membrane targeting properties. Therefore, the mouse can display the labeled cells in a fluorescent color different from the unlabeled cells under excitation light conditions, greatly improving the observation resolution. b Immunofluorescence image of healthy renal tubules in a double-fluorescent reporter transgenic mouse (Sox9-CreERT2; R26mTmG) treated with tamoxifen using TPM. In this mouse, intraperitoneal injection of tamoxifen effectively labeled Sox9+ cells and their progeny with membrane-located EGFP, while other cells were labeled with membrane-targeted tdTomato. The renal cells in the sham group were basically all tdTomato+, indicating that the majority of normal adult kidney cells were Sox9−. c After kidney injury, Sox9+ cells expanded in large numbers (showing EGFP fluorescence). Reproduced with permission from Ref. [5]
At present, researchers designed a double-fluorescent reporter mouse, which is the membrane-targeted tdTomato/membrane-targeted EGFP (mT/mG) [47]. In this reporter mouse model, Tomato Red protein is constitutively expressed in all cells in the mouse. The gene segment encoding mT is floxed by two Loxp sites; only cells that experience Cre-mediated recombination activity will express EGFP protein that situates downstream of the mT sequence, thus converting red fluorescence into green (Fig. 2a). In addition, both tdTomato and EGFP expressed in this mouse have cell membrane targeting properties; the labeled cells can be imaged under excitation light, distinguished from the unlabeled ones, responding to a higher resolution. In this way, researchers can verify whether the cells of interest express the reporter allele [47]. The mT/mG reporter system was introduced in the process of renal podocyte turnover and regeneration under three different conditions: aging, unilateral nephrectomy, and acute podocyte loss, respectively. To study the serious loss of glomerular podocytes, Wanner et al. designed a quadruple transgenic model in which mT/mG is located behind the Rosa26 promoter. After injection of doxycycline, it will induce a specific deletion of GFP, thereby marking the newborn podocytes in red [4, 48, 49].
Another mTmG application example is the kidney lineage tracing study from our laboratory [5]. We prepared a kind of mice induced by tamoxifen, named Sox9-CreERT2; by hybridizing CreERT2 mice driven by Sox9 promoter with R26mTmG mice, Sox9+ cells were specifically labeled. In these mice, intraperitoneal injection of tamoxifen can effectively label Sox9-positive cells and their progeny in green (GFP), while other cells are labeled in red (tdTomato) (Fig. 2b). We found that in normal mice, most of the cells showed tdTomato+; when AKI was induced, EGFP-labeled Sox9-positive cells expanded in large numbers, promoting the repair of renal tubular injury (Fig. 2c).
Researchers proposed that using Cre mice constructed with cell/tissue-specific promoters are able to analyze the contribution of potential intratubular stem/progenitor cells to tubular regeneration. SLC34a1 is a sodium-dependent inorganic phosphate transporter, which is only expressed in the differentiated proximal tubule. The researchers used it as a marker for tracking terminally differentiated proximal tubular cells. By integration between lineage tracing and the SLC34a1 promoter, Kusaba et al. concluded that progenitor cells did not contribute to the regeneration of tubular cells after injury, because the fluorescent protein markers were not diluted during the damage repair process [40]. This research depends on the use of cell markers to uniformly label cells and their progeny; however, it does not consider that the intrinsic progenitor cells of the kidney also express differentiated tubular cell markers to a certain extent. Of course, it has also been reported that tissue-specific progenitor cells express differentiation-related markers in several organs [50,51,52]. Benjamin D. Humphreys’ research team used lineage tracing technology to prove that the regeneration of surviving renal tubular epithelial cells is the main repair mechanism after adult mammalian kidney ischemia-reperfusion injury [53, 54]. As for renal fibrosis, their laboratory uses lineage tracing methods to show that myofibroblasts are not produced by tubular epithelial cells through epithelial to mesenchymal transition (EMT) but derived from endogenous pericytes and interstitial fibroblasts [55]. These studies illustrated well that lineage tracing can be used as a powerful means for exploring the mechanisms of kidney development, disease, and regeneration processes.
Intravital microscopy (IVM)
The structure of the kidney tissue is complex, with a highly branched epithelial morphology, and complicated three-dimensional (3D) imaging of thick tissue is required to analyze the entire fate map of the kidney. In traditional confocal microscopy, a selective filter only allows light of a specific wavelength to pass through, and a pinhole in front of the detector blocks any out-of-focus light, thereby generating an emission image corresponding to the excited sample point. However, confocal microscopy has limitations in photobleaching, excitation light scattering, emission light scattering, and imaging time [56,57,58].
Different from single photon excitation using continuous photon flux, the basic principle of two-photon excitation is that, at high photon densities, fluorescent molecules can absorb two long-wavelength photons simultaneously, and emit a short-wavelength photon after a short period of time, the so-called excited state lifetime [59]. The effect is equivalent to using a photon having a wavelength of half the long wavelength to excite the fluorescent molecule [58, 60, 61] (Fig. 3a). When two incident photons coincide spatially and temporally, the simultaneous absorption of a single photon results in a selective increase of photon energy, which then obtains the energy required to excite the fluorescent dye or protein of interest. In the case of two-photon excitation, mild infrared or near-infrared light, such as a 750-nm laser, can be used to obtain 450-nm fluorescence. Here, we also list some common fluorescent molecules that can be excited by two photons [57, 62,63,64,65] (Table 1). Compared with ordinary laser microscopy, TPM has the advantages of strong tissue penetrating power, high fluorescence signal-to-noise ratio, and small phototoxicity [66,67,68,69] (Table 2). It should be emphasized that the kidney is one of the most optically challenging organs for intravital microscopy imaging due to its strong autofluorescence, which is unfortunate for kidney research [70]. Compared with other tissues, such as neocortex, the depth of intravital TPM imaging can reach 1 mm, while the imaging depth of kidney tissue is about 200 μm, which is far superior to traditional microscopes [63, 71].

Principle and schematic setup of intravital two-photon microscopy. a Scheme for one-and two photon excitations. Unlike single photon excitation, which uses a continuous photon flux, two-photon effect is a phenomenon in which a molecule “almost” simultaneously absorbs two photons of the same or different frequencies, causing the molecule to transition from a low-level to a high-energy state, which is a nonlinear optical effect. b Schematic of the intravital two-photon system using AIW
The innovation of the abdominal imaging window (AIW) makes the imaging deepness of the TPM deeper and can perform kidney intravital imaging over multiple days. This window is composed of a reusable titanium ring, with a 1-mm groove on the side, and a glass slide attached to the top, which is sutured with a purse string to make it tightly fixed on the skin and abdominal wall [72]. The purse-string suture is located in the annular groove and does not directly expose the abdominal cavity, so there is no risk of infection [72,73,74,75] (Fig. 3b).
For kidney AIW, the specific surgical procedures are as follows: First, make a 1-cm-long dorsal abdominal incision above the left kidney. Use non-absorbable surgical sutures to connect the skin to the muscular layer of the abdominal wall by pure string suture. Then, we use a 10-μL pipette tip to apply cyanoacrylate glue to the inner surface of the AIW, where the coverslip is attached to the titanium ring. Use forceps to carefully place the AIW on the exposed kidney surface and hold for 5 min until the glue is dry to ensure proper implantation of the window. After AIW implantation, place the abdominal wall and muscles in the titanium ring groove, and carefully tighten both ends of the purse-string suture to fix the window in place. Tie a double knot at the end of the purse-string suture and then hide the knotted end of the suture in the titanium ring to ensure that the mouse will not lose the suture knot. Allow the mouse to recover on the heating pad for approximately 30 min, and then return it to the cage for postoperative monitoring [76,77,78]. Under normal circumstances, AIW can last about 2 weeks in mice. After recovery from surgery, AIW serves as a temporary window for observing the live kidney of mice, allowing the study of structural and functional changes during kidney injury and regeneration.
IVM technologies improve the efficiency of kidney lineage tracing studies
The sensitivity and accuracy of fluorescent reporters continue to increase, making genetic recombination very suitable for live imaging of living cells. Today, primary technical progress in the evolution of microscopy (e.g., two-photon microscopy) combined with genetically encoded fluorescent proteins of cell lineage enables intravital imaging technology to be used for monitoring biological events in real time in living animals [79,80,81]. Thus, at present, intravital microscopy (IVM) largely relies on the detection of fluorescence signals and it has helped to track the fate and function of single renal cells (Table 3).
Scientists have utilized the characteristics of firefly luciferase and Renilla luciferase to perform bioluminescence imaging (BLI) in the presence of specific substrates to develop a reporter system that allows noninvasive tracking of cells [90, 91]. For example, the Barasch group fused the Ngal gene, a marker of kidney damage, with the luciferase reporter gene to generate Ngal-Luc reporter mice. In this mouse model, when kidney damage occurs, the expression of the Ngal gene is induced, thereby activating luciferase expression, reproducing the endogenous information, and illuminating damage in vivo in real time [82].
The above strategy was different from the Cre/Loxp system. Although BLI has a small background and high signal-to-noise ratio, it cannot provide information on cell resolution [92]. This problem can be circumvented by using intravital microscopy (e.g., confocal TPM) combined with fluorescent reporter transgenic animal models. It is not surprising that this was mainly established in zebrafish and mouse models, which have the most advanced and comprehensive genetically modified strains [70].
Podocytes together with vascular endothelial cells and glomerular basement membrane constitute the glomerular blood filtration barrier. The special anatomical location of podocytes makes it difficult to study in vivo. The János Peti-Peterdi group used multiphoton microscopy (MPM) to track the lineage development of the same glomerulus over several consecutive days to visualize the movement of podocytes and parietal epithelial cells (PEC) in vivo. The transgenic mice they used were Podocin-GFP mice, Podocin-confetti mice, and PEPCK-GFP mice. Their results show that the glomerular environment and cell composition are highly dynamic rather than static. MPM intravital imaging combined with genetic lineage tracing is expected to promote the understanding of glomerular injury and regeneration mechanisms [83]. Similarly, the Benjamin D. Humphreys laboratory utilized a multicolor labeling method to track the changes in podocyte morphology after injury over time, which were performed by conventional fluorescence (optical) microscopy in fixed kidneys and ex vivo [84].
Intravital microscopy imaging can also observe the inside of a single cell of the intact living kidney, and quantitatively display intracellular variables and cell signals. Researchers combined genetic lineage tracing techniques with intravital imaging technologies to study Ca2+ signals in the nephron. Burford et al. conducted an experiment, they developed an imaging approach that used TPM to directly visualize podocyte Ca2+ dynamics within the intact kidneys of live mice expressing a fluorescent calcium indicator only in podocytes [85]. They designed a new podocin/Cre-GCaMP3/flox mouse model and combined with MPM, so that they can directly visualize Ca2+ dynamics in the critical but inaccessible renal cell type of podocytes in vivo in the intact kidney [85]. For the first time, they combined intravital TPM imaging with transgenic mice for research. This method can be a powerful tool for detecting changes in podocyte Ca2+ in glomerular function in physical or disease states. In Kornelia Szebenyi group, they used a calcium-sensitive fluorescent probe to provide the first detailed description of intracellular calcium signals in kidney tubules in vivo. They produced a transgenic rat stably expressing the GFP-calmodulin based genetically encoded calcium indicator (GCaMP2) predominantly in the kidney proximal tubule. They used in vitro confocal microscopy and in vivo TPM to detect basic calcium levels in proximal tubular epithelial cells, as well as changes in these levels caused by ligands and drugs [87].
In glomerular disease, podocyte damage leads to large changes in cell morphology. However, whether podocytes are static or actively moving cells in the body is still controversial. To analyze the motility of podocytes in vivo, the researchers utilized intravital TPM to image the kidneys of Confetti mice in vivo. They discovered that uninjured podocytes stay inactive, while transfer into a dynamic state after suffering from injury [86].
To better illuminate the potential role of interstitial cells in renal tubular regeneration, the laboratory of Ina Maria Schiessl recently used AIW to trace PDGFRβ-mTmG transgenic mice via intravital MPM. This research provided visual evidence that renal interstitial cells play a crucial role in renal tubular epithelial regeneration [76].
Another outstanding example of the application of intravital imaging technology via AIW to enhance the efficiency of kidney lineage tracing comes from our research group. We utilized the AIW to perform intravital imaging of Sox9-positive cells under TPM. Using the high resolution of AIW, we performed TPM intravital imaging with a deepness of 200 μm and observed the kidney structure. We performed serial8 real-time imaging on days 1, 3, 7, and 14 after ischemia-reperfusion injury to continuously observe the proliferation of Sox9-positive cells. By combining kidney Sox9 lineage tracing with two-photon intravital imaging technology, we can intuitively observe the kidney injury and regeneration in real time in vivo, greatly improving the understanding of the results of lineage tracing [5].
In summary, the application of genetic lineage tracing techniques in combination with modern intravital imaging will enable long-term lineage tracing studies of kidney development, homeostasis, and disease. However, so far, there are not many studies on the combined application of intravital imaging technology and genetic lineage tracing strategy to the kidney. This may be affected by factors such as the imaging depth of the intravital microscopy, the complex structure of the kidney, and the respiratory rate.
Conclusion
Lineage tracing is an overwhelming strategy that is widely used to interpret the pathophysiological mechanisms of kidney injury and regeneration and now becomes accessible to nearly all scientists. With the appearance of a large number of novel Cre driver mouse lines, fate tracking of any cell type in the kidney can be achieved. This forceful technology also has limitations, such as the difficulty of interpreting complicated results. Despite all these deficiencies, lineage tracing techniques provide more intuitive and scientific research means for exploring the origin of cells in normal development, disease occurrence, and injury repair processes. Recently, intravital imaging with advanced optical equipment such as TPM has provided a forceful tool for the tracing of live cell lineages. It has been demonstrated that using intravital microscopy can accurately quantify the complex dynamic cellular processes in the kidneys and assess their real-time response in pathological states. With the continuous improvement of laser, optics, and microscopy technologies, the development of new TPM imaging modalities and their combination with transgenic animal models will further promote their use in the study of kidney disease or may be used for clinical diagnosis.
Availability of data and materials
Not applicable.
Abbreviations
- AKI:
-
Acute kidney injury
- BFP:
-
Blue fluorescent protein
- BLI:
-
Bioluminescence imaging
- CFP:
-
Cyan fluorescent protein
- CKD:
-
Chronic kidney disease
- Cre:
-
Cyclization Recombination Enzyme
- GFP:
-
Green fluorescent protein
- IVM:
-
Intravital microscopy
- MPM:
-
Multi-photon microscopy
- mTmG:
-
Membrane-targeted tdTomato/membrane-targeted EGFP
- PEC:
-
Parietal epithelial cells
- RFP:
-
Red fluorescent protein
- TPM:
-
Two-photon microscopy
- YFP:
-
Yellow fluorescent protein
References
Hato T, Winfree S, Dagher PC. Intravital imaging of the kidney. Methods. 2017;128:33–9. https://doi.org/10.1016/j.ymeth.2017.03.024.
Dunn KW, Sandoval RM, Molitoris BA. Intravital imaging of the kidney using multiparameter multiphoton microscopy. Nephron Exp Nephrol. 2003;94(1):e7–11. https://doi.org/10.1159/000070813.
Liu Y, Cui J, Wang H, Hezam K, Zhao X, Huang H, et al. Enhanced therapeutic effects of MSC-derived extracellular vesicles with an injectable collagen matrix for experimental acute kidney injury treatment. Stem Cell Res Ther. 2020;11(1):161. https://doi.org/10.1186/s13287-020-01668-w.
Romagnani P, Rinkevich Y, Dekel B. The use of lineage tracing to study kidney injury and regeneration. Nat Rev Nephrol. 2015;11(7):420–31. https://doi.org/10.1038/nrneph.2015.67.
Zhang K, Chen S, Sun H, Wang L, Li H, Zhao J, et al. In vivo two-photon microscopy reveals the contribution of Sox9+ cell to kidney regeneration in a mouse model with extracellular vesicle treatment. J Biol Chem. 2020;295(34):12203–13.
Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449(7165):1003–7. https://doi.org/10.1038/nature06196.
Chaible LM, Corat MA, Abdelhay E, Dagli ML. Genetically modified animals for use in research and biotechnology. Genet Mol Res. 2010;9(3):1469–82. https://doi.org/10.4238/vol9-3gmr867.
Rios AC, Fu NY, Lindeman GJ, Visvader JE. In situ identification of bipotent stem cells in the mammary gland. Nature. 2014;506(7488):322–7.
Snippert HJ, van der Flier LG, Sato T, van Es JH, van den Born M, Kroon-Veenboer C, et al. Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell. 2010;143(1):134–44. https://doi.org/10.1016/j.cell.2010.09.016.
Kretzschmar K, Watt FM. Lineage tracing. Cell. 2012;148(1-2):33–45. https://doi.org/10.1016/j.cell.2012.01.002.
Lee HW, Gangadaran P, Kalimuthu S, Ahn BC. Advances in molecular imaging strategies for in vivo tracking of immune cells. Biomed Res Int. 2016;2016:1946585.
Fink J, Andersson-Rolf A, Koo B-K. Adult stem cell lineage tracing and deep tissue imaging. BMB Reports. 2015;48(12):655–67. https://doi.org/10.5483/BMBRep.2015.48.12.249.
Peti-Peterdi J, Kidokoro K, Riquier-Brison A. Intravital imaging in the kidney. Curr Opin Nephrol Hypertens. 2016;25(3):168–73. https://doi.org/10.1097/MNH.0000000000000219.
Kaverina NV, Eng DG, Freedman BS, Kutz JN, Chozinski TJ, Vaughan JC, et al. Dual lineage tracing shows that glomerular parietal epithelial cells can transdifferentiate toward the adult podocyte fate. Kidney Int. 2019;96(3):597–611. https://doi.org/10.1016/j.kint.2019.03.014.
Pittet MJ, Weissleder R. Intravital imaging. Cell. 2011;147(5):983–91. https://doi.org/10.1016/j.cell.2011.11.004.
Chang-Panesso M, Kadyrov FF, Lalli M, Wu H, Ikeda S, Kefaloyianni E, et al. FOXM1 drives proximal tubule proliferation during repair from acute ischemic kidney injury. J Clin Invest. 2019;129(12):5501–17.
Häcker I, Schetelig MF. Molecular tools to create new strains for mosquito sexing and vector control. Parasit Vectors. 2018;11(Suppl 2):645. https://doi.org/10.1186/s13071-018-3209-6.
Theodosiou NA, Xu T. Use of FLP/FRT system to study Drosophila development. Methods. 1998;14(4):355–65. https://doi.org/10.1006/meth.1998.0591.
Nagy A. Cre recombinase: the universal reagent for genome tailoring. Genesis. 2000;26(2):99–109. https://doi.org/10.1002/(SICI)1526-968X(200002)26:2<99::AID-GENE1>3.0.CO;2-B.
Song DL, Chalepakis G, Gruss P, Joyner AL. Two Pax-binding sites are required for early embryonic brain expression of an Engrailed-2 transgene. Development. 1996;122(2):627–35. https://doi.org/10.1242/dev.122.2.627.
Soriano P. Generalized lacZ expression with the ROSA26 Cre reporter strain. Nat Genet. 1999;21(1):70–1. https://doi.org/10.1038/5007.
Hsu YC. Theory and practice of lineage tracing. Stem Cells. 2015;33(11):3197–204. https://doi.org/10.1002/stem.2123.
Ito M, Liu Y, Yang Z, Nguyen J, Liang F, Morris RJ, et al. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat Med. 2005;11(12):1351–4. https://doi.org/10.1038/nm1328.
Tata PR, Mou H, Pardo-Saganta A, Zhao R, Prabhu M, Law BM, et al. Dedifferentiation of committed epithelial cells into stem cells in vivo. Nature. 2013;503(7475):218–23. https://doi.org/10.1038/nature12777.
Jensen KB, Collins CA, Nascimento E, Tan DW, Frye M, Itami S, et al. Lrig1 expression defines a distinct multipotent stem cell population in mammalian epidermis. Cell Stem Cell. 2009;4(5):427–39. https://doi.org/10.1016/j.stem.2009.04.014.
Zong H, Espinosa JS, Su HH, Muzumdar MD, Luo L. Mosaic analysis with double markers in mice. Cell. 2005;121(3):479–92. https://doi.org/10.1016/j.cell.2005.02.012.
Livet J, Weissman TA, Kang H, Draft RW, Lu J, Bennis RA, et al. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature. 2007;450(7166):56–62. https://doi.org/10.1038/nature06293.
de Roo JJ, Vloemans SA, Vrolijk H, de Haas EF, Staal FJ. Development of an in vivo model to study clonal lineage relationships in hematopoietic cells using Brainbow2.1/Confetti mice. Future Sci. 2019;5(10):FSO427.
Bowman AN, van Amerongen R, Palmer TD, Nusse R. Lineage tracing with Axin2 reveals distinct developmental and adult populations of Wnt/beta-catenin-responsive neural stem cells. Proc Natl Acad Sci U S A. 2013;110(18):7324–9. https://doi.org/10.1073/pnas.1305411110.
Lazzeri E, Angelotti ML, Peired A, Conte C, Marschner JA, Maggi L, et al. Endocycle-related tubular cell hypertrophy and progenitor proliferation recover renal function after acute kidney injury. Nat Commun. 2018;9(1):1344. https://doi.org/10.1038/s41467-018-03753-4.
Metzger D, Chambon P. Site- and time-specific gene targeting in the mouse. Methods. 2001;24(1):71–80. https://doi.org/10.1006/meth.2001.1159.
Littlewood TD, Hancock DC, Danielian PS, Parker MG, Evan GI. A modified oestrogen receptor ligand-binding domain as an improved switch for the regulation of heterologous proteins. Nucleic Acids Res. 1995;23(10):1686–90.
Hayashi S, McMahon AP. Efficient recombination in diverse tissues by a tamoxifen-inducible form of Cre: a tool for temporally regulated gene activation/inactivation in the mouse. Dev Biol. 2002;244(2):305–18. https://doi.org/10.1006/dbio.2002.0597.
Feil R, Wagner J, Metzger D, Chambon P. Regulation of Cre recombinase activity by mutated estrogen receptor ligand-binding domains. Biochem Biophys Res Commun. 1997;237(3):752–7. https://doi.org/10.1006/bbrc.1997.7124.
Barker N, Rookmaaker MB, Kujala P, Ng A, Leushacke M, Snippert H, et al. Lgr5(+ve) stem/progenitor cells contribute to nephron formation during kidney development. Cell Rep. 2012;2(3):540–52. https://doi.org/10.1016/j.celrep.2012.08.018.
Kang HM, Huang S, Reidy K, Han SH, Chinga F, Susztak K. Sox9-positive progenitor cells play a key role in renal tubule epithelial regeneration in mice. Cell Rep. 2016;14(4):861–71. https://doi.org/10.1016/j.celrep.2015.12.071.
Rinkevich Y, Lindau P, Ueno H, Longaker MT, Weissman IL. Germ-layer and lineage-restricted stem/progenitors regenerate the mouse digit tip. Nature. 2011;476(7361):409–13. https://doi.org/10.1038/nature10346.
Lasagni L, Angelotti ML, Ronconi E, Lombardi D, Nardi S, Peired A, et al. Podocyte regeneration driven by renal progenitors determines glomerular disease remission and can be pharmacologically enhanced. Stem Cell Rep. 2015;5(2):248–63. https://doi.org/10.1016/j.stemcr.2015.07.003.
Kumar S, Liu J, Pang P, Krautzberger AM, Reginensi A, Akiyama H, et al. Sox9 activation highlights a cellular pathway of renal repair in the acutely injured mammalian kidney. Cell Rep. 2015;12(8):1325–38. https://doi.org/10.1016/j.celrep.2015.07.034.
Kusaba T, Lalli M, Kramann R, Kobayashi A, Humphreys BD. Differentiated kidney epithelial cells repair injured proximal tubule. Proc Natl Acad Sci U S A. 2014;111(4):1527–32. https://doi.org/10.1073/pnas.1310653110.
El-Dahr SS, Li Y, Liu J, Gutierrez E, Hering-Smith KS, Signoretti S, et al. p63+ ureteric bud tip cells are progenitors of intercalated cells. JCI Insight. 2017;2(9):e89996.
Hsu SY, Liang SG, Hsueh AJ. Characterization of two LGR genes homologous to gonadotropin and thyrotropin receptors with extracellular leucine-rich repeats and a G protein-coupled, seven-transmembrane region. Mol Endocrinol. 1998;12(12):1830–45. https://doi.org/10.1210/mend.12.12.0211.
Humphreys BD, DiRocco DP. Lineage-tracing methods and the kidney. Kidney Int. 2014;86(3):481–8. https://doi.org/10.1038/ki.2013.368.
Wagner T, Wirth J, Meyer J, Zabel B, Held M, Zimmer J, et al. Autosomal sex reversal and campomelic dysplasia are caused by mutations in and around the SRY-related gene SOX9. Cell. 1994;79(6):1111–20. https://doi.org/10.1016/0092-8674(94)90041-8.
Foster JW, Dominguez-Steglich MA, Guioli S, Kwok C, Weller PA, Stevanovic M, et al. Campomelic dysplasia and autosomal sex reversal caused by mutations in an SRY-related gene. Nature. 1994;372(6506):525–30. https://doi.org/10.1038/372525a0.
Reginensi A, Clarkson M, Neirijnck Y, Lu B, Ohyama T, Groves AK, et al. SOX9 controls epithelial branching by activating RET effector genes during kidney development. Hum Mol Genet. 2011;20(6):1143–53.
Muzumdar MD, Tasic B, Miyamichi K, Li L, Luo L. A global double-fluorescent Cre reporter mouse. Genesis. 2007;45(9):593–605.
Wanner N, Hartleben B, Herbach N, Goedel M, Stickel N, Zeiser R, et al. Unraveling the role of podocyte turnover in glomerular aging and injury. J Am Soc Nephrol. 2014;25(4):707–16. https://doi.org/10.1681/ASN.2013050452.
Dekel B, Zangi L, Shezen E, Reich-Zeliger S, Eventov-Friedman S, Katchman H, et al. Isolation and characterization of nontubular sca-1+lin- multipotent stem/progenitor cells from adult mouse kidney. J Am Soc Nephrol. 2006;17(12):3300–14. https://doi.org/10.1681/ASN.2005020195.
Mascre G, Dekoninck S, Drogat B, Youssef KK, Brohee S, Sotiropoulou PA, et al. Distinct contribution of stem and progenitor cells to epidermal maintenance. Nature. 2012;489(7415):257–62. https://doi.org/10.1038/nature11393.
Pina C, Fugazza C, Tipping AJ, Brown J, Soneji S, Teles J, et al. Inferring rules of lineage commitment in haematopoiesis. Nat Cell Biol. 2012;14(3):287–94. https://doi.org/10.1038/ncb2442.
Awatramani R, Soriano P, Rodriguez C, Mai JJ, Dymecki SM. Cryptic boundaries in roof plate and choroid plexus identified by intersectional gene activation. Nat Genet. 2003;35(1):70–5. https://doi.org/10.1038/ng1228.
Humphreys BD, Valerius MT, Kobayashi A, Mugford JW, Soeung S, Duffield JS, et al. Intrinsic epithelial cells repair the kidney after injury. Cell Stem Cell. 2008;2(3):284–91. https://doi.org/10.1016/j.stem.2008.01.014.
Swanhart LM, Cosentino CC, Diep CQ, Davidson AJ, de Caestecker M, Hukriede NA. Zebrafish kidney development: basic science to translational research. Birth Defects Res C Embryo Today. 2011;93(2):141–56. https://doi.org/10.1002/bdrc.20209.
Molitoris BA, Sandoval RM. Intravital multiphoton microscopy of dynamic renal processes. Am J Physiol Renal Physiol. 2005;288(6):F1084–9. https://doi.org/10.1152/ajprenal.00473.2004.
Wang E, Babbey CM, Dunn KW. Performance comparison between the high-speed Yokogawa spinning disc confocal system and single-point scanning confocal systems. J Microsc. 2005;218(Pt 2):148–59. https://doi.org/10.1111/j.1365-2818.2005.01473.x.
Small DM, Sanchez WY, Gobe GC. Intravital multiphoton imaging of the kidney: tubular structure and metabolism. Methods Mol Biol. 2016;1397:155–72. https://doi.org/10.1007/978-1-4939-3353-2_12.
Hall AM, Schuh CD, Haenni D. New frontiers in intravital microscopy of the kidney. Curr Opin Nephrol Hypertens. 2017;26(3):172–8. https://doi.org/10.1097/MNH.0000000000000313.
Santos MA, Goertz DE, Hynynen K. Focused ultrasound hyperthermia mediated drug delivery using thermosensitive liposomes and visualized with in vivo two-photon microscopy. Theranostics. 2017;7(10):2718–31. https://doi.org/10.7150/thno.19662.
Denk W, Strickler JH, Webb WW. Two-photon laser scanning fluorescence microscopy. Science. 1990;248(4951):73–6. https://doi.org/10.1126/science.2321027.
Masedunskas A, Milberg O, Porat-Shliom N, Sramkova M, Wigand T, Amornphimoltham P, et al. Intravital microscopy: a practical guide on imaging intracellular structures in live animals. Bioarchitecture. 2012;2(5):143–57. https://doi.org/10.4161/bioa.21758.
Diaspro A, Chirico G, Collini M. Two-photon fluorescence excitation and related techniques in biological microscopy. Q Rev Biophys. 2005;38(2):97–166. https://doi.org/10.1017/S0033583505004129.
Nimmerjahn A, Kirchhoff F, Kerr JN, Helmchen F. Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat Methods. 2004;1(1):31–7. https://doi.org/10.1038/nmeth706.
Svoboda K, Denk W, Kleinfeld D, Tank DW. In vivo dendritic calcium dynamics in neocortical pyramidal neurons. Nature. 1997;385(6612):161–5.
Gibson EA, Masihzadeh O, Lei TC, Ammar DA, Kahook MY. Multiphoton microscopy for ophthalmic imaging. J Ophthalmol. 2011;2011:870879.
Bewersdorf J, Pick R, Hell SW. Multifocal multiphoton microscopy. Opt Lett. 1998;23(9):655–7. https://doi.org/10.1364/OL.23.000655.
Fan Y, Sun Y, Chang W, Zhang X, Tang J, Zhang L, et al. Bioluminescence imaging and two-photon microscopy guided laser ablation of GBM decreases tumor burden. Theranostics. 2018;8(15):4072–85. https://doi.org/10.7150/thno.25357.
Grüneboom A, Kling L, Christiansen S, Mill L, Maier A, Engelke K, et al. Next-generation imaging of the skeletal system and its blood supply. Nat Rev Rheumatol. 2019;15(9):533–49. https://doi.org/10.1038/s41584-019-0274-y.
Schuh CD, Haenni D, Craigie E, Ziegler U, Weber B, Devuyst O, et al. Long wavelength multiphoton excitation is advantageous for intravital kidney imaging. Kidney Int. 2016;89(3):712–9. https://doi.org/10.1038/ki.2015.323.
Peti-Peterdi J, Kidokoro K, Riquier-Brison A. Novel in vivo techniques to visualize kidney anatomy and function. Kidney Int. 2015;88(1):44–51. https://doi.org/10.1038/ki.2015.65.
Helmchen F, Denk W. Deep tissue two-photon microscopy. Nat Methods. 2005;2(12):932–40. https://doi.org/10.1038/nmeth818.
Ritsma L, Steller EJ, Beerling E, Loomans CJ, Zomer A, Gerlach C, et al. Intravital microscopy through an abdominal imaging window reveals a pre-micrometastasis stage during liver metastasis. Sci Transl Med. 2012;4(158):158ra45.
Zhang C, Feng W, Zhao Y, Yu T, Li P, Xu T, et al. A large, switchable optical clearing skull window for cerebrovascular imaging. Theranostics. 2018;8(10):2696–708.
Huang S, Rompolas P. Two-photon microscopy for intracutaneous imaging of stem cell activity in mice. Exp Dermatol. 2017;26(5):379–83. https://doi.org/10.1111/exd.13221.
Revell DZ, Yoder BK. Intravital visualization of the primary cilium, tubule flow, and innate immune cells in the kidney utilizing an abdominal window imaging approach. Methods Cell Biol. 2019;154:67–83. https://doi.org/10.1016/bs.mcb.2019.04.012.
Schiessl IM, Grill A, Fremter K, Steppan D, Hellmuth MK, Castrop H. Renal interstitial platelet-derived growth factor receptor-beta cells support proximal tubular regeneration. J Am Soc Nephrol. 2018;29(5):1383–96. https://doi.org/10.1681/ASN.2017101069.
Shroff UN, Schiessl IM, Gyarmati G, Riquier-Brison A, Peti-Peterdi J. Novel fluorescence techniques to quantitate renal cell biology. Methods Cell Biol. 2019;154:85–107. https://doi.org/10.1016/bs.mcb.2019.04.013.
Schiessl IM, Fremter K, Burford JL, Castrop H, Peti-Peterdi J. Long-term cell fate tracking of individual renal cells using serial intravital microscopy. Methods Mol Biol. 2020;2150:25–44.
Lichtman JW, Conchello JA. Fluorescence microscopy. Nat Methods. 2005;2(12):910–9. https://doi.org/10.1038/nmeth817.
Schroeder T. Imaging stem-cell-driven regeneration in mammals. Nature. 2008;453(7193):345–51. https://doi.org/10.1038/nature07043.
Kokkaliaris KD, Loeffler D, Schroeder T. Advances in tracking hematopoiesis at the single-cell level. Curr Opin Hematol. 2012;19(4):243–9. https://doi.org/10.1097/MOH.0b013e32835421de.
Paragas N, Qiu A, Zhang Q, Samstein B, Deng SX, Schmidt-Ott KM, et al. The Ngal reporter mouse detects the response of the kidney to injury in real time. Nat Med. 2011;17(2):216–22. https://doi.org/10.1038/nm.2290.
Hackl MJ, Burford JL, Villanueva K, Lam L, Suszták K, Schermer B, et al. Tracking the fate of glomerular epithelial cells in vivo using serial multiphoton imaging in new mouse models with fluorescent lineage tags. Nat Med. 2013;19(12):1661–6. https://doi.org/10.1038/nm.3405.
Grgic I, Brooks CR, Hofmeister AF, Bijol V, Bonventre JV, Humphreys BD. Imaging of podocyte foot processes by fluorescence microscopy. J Am Soc Nephrol. 2012;23(5):785–91. https://doi.org/10.1681/ASN.2011100988.
Burford JL, Villanueva K, Lam L, Riquier-Brison A, Hackl MJ, Pippin J, et al. Intravital imaging of podocyte calcium in glomerular injury and disease. J Clin Invest. 2014;124(5):2050–8. https://doi.org/10.1172/JCI71702.
Brahler S, Yu H, Suleiman H, Krishnan GM, Saunders BT, Kopp JB, et al. Intravital and kidney slice imaging of podocyte membrane dynamics. J Am Soc Nephrol. 2016;27(11):3285–90. https://doi.org/10.1681/ASN.2015121303.
Szebenyi K, Furedi A, Kolacsek O, Csohany R, Prokai A, Kis-Petik K, et al. Visualization of calcium dynamics in kidney proximal tubules. J Am Soc Nephrol. 2015;26(11):2731–40. https://doi.org/10.1681/ASN.2014070705.
Hickmann L, Steglich A, Gerlach M, Al-Mekhlafi M, Sradnick J, Lachmann P, et al. Persistent and inducible neogenesis repopulates progenitor renin lineage cells in the kidney. Kidney Int. 2017;92(6):1419–32. https://doi.org/10.1016/j.kint.2017.04.014.
Kaverina NV, Kadoya H, Eng DG, Rusiniak ME, Sequeira-Lopez ML, Gomez RA, et al. Tracking the stochastic fate of cells of the renin lineage after podocyte depletion using multicolor reporters and intravital imaging. PLoS One. 2017;12(3):e0173891. https://doi.org/10.1371/journal.pone.0173891.
Kocher B, Piwnica-Worms D. Illuminating cancer systems with genetically engineered mouse models and coupled luciferase reporters in vivo. Cancer Discov. 2013;3(6):616–29. https://doi.org/10.1158/2159-8290.CD-12-0503.
Greer LF 3rd, Szalay AA. Imaging of light emission from the expression of luciferases in living cells and organisms: a review. Luminescence. 2002;17(1):43–74. https://doi.org/10.1002/bio.676.
Sato A, Klaunberg B, Tolwani R. In vivo bioluminescence imaging. Comp Med. 2004;54(6):631–4.
Acknowledgements
Not applicable.
Funding
This research was partially supported by the National Key R&D Program of China (2017YFA0103200) and National Natural Science Foundation of China (U2004126).
Author information
Authors and Affiliations
Contributions
YL conceived and wrote the manuscript. ZL revised and approved the final manuscript. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, Y., Li, Z. Intravital microscopy imaging of kidney injury and regeneration. Ren Replace Ther 7, 23 (2021). https://doi.org/10.1186/s41100-021-00342-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41100-021-00342-y