
Abstract
Background
Many animals in coral reefs exhibit lunar cycles in their reproduction, showing synchronous gametogenesis and spawning at a particular moon phase. How these lunar reproductive cycles are endogenously regulated remains unknown, although changes in moonlight between the new moon and full moon are likely involved in this rhythmic event.
Results
This study evaluated the possible role of cryptochrome (cry1, cry2, cry3) in phase shifting and setting in the honeycomb grouper Epinephelus merra, which is a typical lunar spawner with full moon preference. qPCR analysis revealed that when fish were reared under alternating light-dark conditions, the transcript levels of cry1 and cry2, but not of cry3, in the diencephalon and pituitary gland showed daily variations. Weekly collection at midnight showed increases in the transcript levels of cry1 and cry2 in the diencephalon, but not the pituitary gland, from the first quarter moon through the last quarter moon. In comparison to the new moon, these transcript levels were significantly lower at all other sampling times. The artificial full moon conditions for 1 month resulted in increased cry transcript levels in both tissues at 2 (cry1) or 2 and 4 (cry2) weeks after the initiation of full moon conditions.
Conclusions
These results indicate the importance of transient changes in “brightness at night” in the response to moonlight for the phase shift and of “darkness at night” during the new moon for the phase set to the determined moon phase. We concluded that the moon phase-dependent oscillation of clock genes plays a role in lunar cycle-dependent behaviors in fish.
Similar content being viewed by others


Background
The lunar reproductive cycle is a rhythmic event observed in many animals, including corals and fish, that is repeated monthly during the spawning season. Physiological activity and behavioral responses related to lunar reproduction cycles are activated before and maximized during a particular moon phase [1]. Moon-derived cues observed from Earth are a potent entrainer for behavioral synchrony, although the extent to which such cues are perceived by animals and transduced into internal stimuli to drive lunar rhythmicity remains unclear. The moon phase when synchronous activity occurs differs among related species; for example, the little spinefoot Siganus spinus and white-spotted spinefoot S. canaliculatus synchronously release their gametes around the new moon [2, 3], while the goldlined spinefoot S. guttatus does so around the first quarter moon [4], and the streamlined spinefoot S. argenteus around the last quarter moon [3]. Therefore, it is likely that certain animals that exhibit lunar synchronization use transient changes in moonlight to adjust the timing of their synchronized behaviors, although adaptability to cues appears to show interspecific variation. This hypothesis is supported by experimental evidence that rearing the goldlined spinefoot for more than 2 weeks under conditions of constant new moon or full moon lights caused disruption of spawning at the predicted moon phase [5]. These previous findings imply that animals with lunar synchronization are capable of perceiving changes in moonlight using photoreceptors in the eyes or the brain, such as the pineal organ. This ability is indirectly assessed using moonlight-related fluctuations of melatonin in spinefoot; exposing fish to “brightness at night” lowered plasma levels of melatonin [5], and culturing of the pineal organ with moonlight suppressed the synthesis of melatonin [6].
The involvement of clock genes in the time-keeping system used for lunar synchronization has been noted in some animals [7,8,9,10]. A pioneer study of the reef-building coral Acropora millepora, which broadcast spawns after the full moon, revealed that the transcript levels of cryptochrome (cry2) were elevated around midnight during the full moon compared to that at the new moon, suggesting that this clock gene is upregulated by “brightness at night” and plays a role in entraining the internal clock of corals [7]. Similar findings have been obtained in fish such as the goldlined spinefoot, in which the transcript levels of cry1 and cry3 in the diencephalon peaked around the first quarter moon, which is the moon phase of synchronous spawning for this species [8], and cry3 and per1 (formally termed per4 [9]) in the brain of this fish oscillated even after nocturnal moonlight was interrupted [9]. In a recent study, when the moonlight stimulus is interrupted daily for one lunar cycle starting at the new moon, the expression patterns of clock genes were modified [10]. Based on these previous studies, we propose that an endogenous clock system involved in the lunar cycle exists in the brain and can be entrained to transient changes in moonlight [11]. We hypothesized that the mechanism underlying the phase setting process is triggered by the transition in moonlight levels from the new moon (dark night) through the full moon (bright night), as crys belongs to an ancient family of blue light-sensitive photoreceptors [12] and are upregulated after light exposure [13].
The honeycomb grouper Epinephelus merra (Epinephelinae) is a common species inhabiting the shallow reef fringe region [14]. This species is a lunar-synchronized spawner with full moon preference [14, 15] and is capable of perceiving moonlight [16]. Using this species, we tested the hypothesis stated above in this study. We cloned cryptochrome subtypes (cry1, cry2, and cry3) from the brain of this species and examined their daily and weekly expression profiles in diencephalon and pituitary tissues. We also compared cry transcript levels in these tissues of fish reared under artificial full moon and new moon conditions. In addition, the short-term effects of melatonin treatment on these transcript levels were evaluated.
Results
Three subtypes of honeycomb grouper crys (cry1 [LC521334], cry2 [LC521335], and cry3 [LC521336]) were partially cloned from the brain (Additional file 1: Fig. S1–3). The predicted amino acid sequences of these CRYs showed high identity with known CRY proteins (94.7–100.0% for CRY1, 81.4–100.0% for CRY2, and 84.9–99.5% for CRY3) (Additional file 1: Fig. S1–3). Phylogenic analysis indicated that CRY1, CRY2, and CRY3 clustered within the respective CRY sequence clades of teleosts (Fig. 1).

Phylogenetic analysis of CRY family including honeycomb grouper and other species. African malaria mosquito CRY1 and fruit fly CRY were used as the outgroup. One thousand bootstrap repetitions were performed, and values are shown at the in inner nodes. The scale bar is calibrated in substitutions per site. The following amino acid sequences were used for the analysis; African malaria mosquito Anopheles gambiae CRY1 (XP_321104.4); chicken Gallus gallus CRY1 (NP_989576.1), CRY2 (NP_989575.1), and CRY4 (NP_001034685.1); fruit fly Drosophila melanogaster CRY (NP_732407.1) and PHR (BAA12067.1); European seabass Dicentrarchus labrax CRY1 (AFP33464.1) and CRY2 (AFP33463.1); human Homo sapiens CRY1 (NP_004066.1) and CRY2 (NP_066940.2); Malabar grouper Epinephelus malabaricus CRY1 (BBJ06076.1), CRY2 (BBJ06077.1), and CRY3 (BBJ06078.1); medaka Oryzias latipes CRY2 (XP_004070204.1); mouse Mus musculus CRY1 (NP_031797.1) and CRY2 (NP_034093.1); takifugu Takifugu rubripes CRY2 (XP_011615916.1); African clawed frog Xenopus laevis CRY1 (NP_001081129.1), CRY2a (NP_001082139.1), and PHR (NP_001083936.1); western clawed frog X. tropicalis CRY4 (NP_001123706.1); zebrafish Danio rerio CRY1a (BAA96846.1), CRY1b (BAA96847.1), CRY2a (BAA96848.1), CRY3 (BAA96850.1), CRY4 (BAA96851.1), and PHR (BAA96852.1)
The transcript levels of crys in diencephalon and pituitary tissues were evaluated at 4 h intervals using qPCR (Fig. 2). Daily variations of cry1 and cry2 transcription were significant in the diencephalon (cry1, P < 0.01; cry2, P < 0.01) and pituitary gland (cry1, P < 0.01; cry2, P < 0.01), while that of cry3 did not show daily variations in the diencephalon (P = 0.58) or the pituitary gland (P = 0.06). The mRNA abundances of cry1 and cry2 increased in both tissues after the initiation of photophase. The cry1 acrophase was estimated to be at ZT06 ± 2.33 in the diencephalon and ZT06 ± 1.55 in the pituitary gland, while that of cry2 was ZT10 ± 2.00 in the diencephalon and ZT14 ± 1.30 in the pituitary gland.

Daily changes in the transcript levels of crys in the diencephalon and pituitary gland of honeycomb grouper. Fish (3 to 5 individuals per each sampling point) were reared under a programmable photoperiod (LD = 14:10) and tissues were taken from fish at 4 h intervals. The mRNA abundance of crys was measured using qPCR. The data were normalized by determining the amount of β-actin and ef1α. Each data is shown as means ± SEM. Different letters indicate statistically significant difference (ANOVA and Kruskal-Wallis Test*); cry1 (P < 0.01), cry2 (P < 0.01), and cry3 (P = 0.58) for the diencephalon; cry1 (P < 0.01*), cry2 (P < 0.01), and cry3 (P = 0.06) for the pituitary gland. The source data and statistical analysis data are included in Additional files 3 and 4, respectively
Using the new moon (dark night) as a reference, the transcript levels of crys in the diencephalon and pituitary tissues were evaluated at night during the first quarter moon (W01/W11), full moon (W02/W12), and last quarter moon (W03/W13) (Fig. 3). The time of sample collection was 0000 h based on the culmination time of the full moon period. As a result, significant increases in transcript levels of cry1 (P < 0.05 from W01 to W03 and P < 0.05 from W11 to W13), cry2 (P < 0.01 from W01 to W03 and P < 0.01 from W11 to W13), cry3 (P < 0.01 from W01 to W03 and P < 0.05 from W11 to W13) were observed in the diencephalon; cry1 (P < 0.01 from W01 to W03 and P < 0.05 from W11 to W13), cry2 (P < 0.01 from W01 to W03), and cry3 (P < 0.05 from W01 to W03) were also detected in the pituitary grand. When the transcript levels of crys during the new moon were compared with those of other moon phases, significantly lower expression of cry2 (P < 0.05) and cry3 (P < 0.05) were observed in the diencephalon around W01/W11 and W02, as well as of cry1 (P < 0.01), cry2 (P < 0.01), and cry3 (P < 0.001) in the pituitary gland throughout the other moon phases. Conversely, significantly higher expression was seen for cry1 (P < 0.05) and cry2 (P < 0.05) in the diencephalon around W13.

Weekly changes in the transcript levels of crys in the diencephalon and pituitary gland of honeycomb grouper. Fish (6 to 13 individuals per each sampling point) were collected at the new moon (NM), first quarter moon (W01 and W11), full moon (W02 and W12) and last quarter moon (W03 and W13) and tissue was taken at ZT18. The mRNA abundance of crys was measured using qPCR. The data were normalized by determining the amount of ef1α. Each point shows the fold change in the expression levels from the new moon. Each data is shown as means ± SEM. Moon phases are shown on each figure. Different letters indicate statistically significant difference one-way ANOVA; cry1 (P < 0.05 from W01 to W03 and P < 0.01 from W11 to W13), cry2 (P < 0.01 from W01 to W03 and P < 0.01 from W11 to W13), and cry3 (P < 0.01 from W01 to W03 and P < 0.05 from W11 to W13) for the diencephalon; cry1 (P < 0.01 from W01 to W03 and P < 0.05 from W11 to W13), cry2 (P < 0.01 from W01 to W03 and P = 0.11 from W11 to W13), and cry3 (P < 0.05 from W01 to W03 and P = 0.25 from W11 to W13) for the pituitary gland. Asterisks show significant difference (P < 0.05) from the new moon. The source data and statistical analysis data are included in Additional files 3 and 4, respectively
To examine whether moonlight exposure influences the transcript levels of cry genes in diencephalon and pituitary tissues, fish were reared for 1 month under artificial full moon or new moon conditions (Fig. 4). When transcript levels of crys in the diencephalon were evaluated at ZT18 (4 h after lights off) on W1 to W4, the mRNA abundances of cry1 and cry2, but not cry3, were significantly higher in the full moon exposure group at W2 than in the new moon group. Significant increases in cry1 and cry2 mRNA abundance were observed in the pituitary gland at W2 and W4, respectively. On the other hand, the transcript levels of cry3 in the pituitary gland at W3 were lower in the full moon group than the new moon group.

Effect of nocturnal light irradiance on the transcript levels of crys in the diencephalon and pituitary gland of honeycomb grouper. After acclimating under a programmable condition (LD = 14:10), fish (3 to 5 individuals per each sampling point) were exposed to artificial full moon and new moon condition and tissues were collected in one-week interval (W1 to W4) at ZT18. The mRNA abundance of crys was measured using qPCR. The data were normalized by determining the amount of β-actin and ef1α. Each point shows comparison of the fold change in the expression levels between the artificial new moon and full moon. Each data is shown as means ± SEM. Different letters indicate statistically significant difference (one-way ANOVA); cry1 (P = 0.10), cry2 (P < 0.05), and cry3 (P < 0.05) for the diencephalon; cry1 (P < 0.05), cry2 (P < 0.05), and cry3 (P = 0.23) for the pituitary gland. Asterisks show statistical difference (P < 0.05) from the new moon condition. The source data and statistical analysis data are included in Additional files 3 and 4, respectively
The effect of melatonin treatment on transcript levels of crys in diencephalon and pituitary tissues was examined in this study. Intraperitoneal injection of melatonin failed to alter the transcript levels of crys in either tissue (Fig. 5).

Effect of melatonin treatment on the transcript levels of crys in the diencephalon and pituitary gland of honeycomb grouper (n = 7). Melatonin (n = 7) and vehicle only (n = 7) were intraperitoneally injected to fish at ZT12 and tissues were collected at ZT19. The data were normalized by determining the amount of ef1α. Each data is shown as means ± SEM. P-value by Student’s t-test is indicated. The source data and statistical analysis data are included in Additional files 3 and 4, respectively
Discussion
Phylogenetic analysis of vertebrate CRY showed that CRY1 and CRY2 of the honeycomb grouper were classified into the teleost CRY1 and CRY2 clusters, in which zebrafish Danio rerio CRY1b and CRY2 cluster, respectively. Both of these proteins inhibited CLOCK:BMAL1-induced transcription [17, 18]. CRY3 of this species clustered with teleost CRY3. In addition to its circadian function, CRY3 may retain a non-circadian function, supported by the weak inhibitory effect of CLOCK:BMAL1-induced transcription in a zebrafish cell line [18].
This study revealed a day-high and night-low fluctuation of cry1 and cry2 transcription in the diencephalon and pituitary gland of the honeycomb grouper, when fish were reared under LD conditions (LD = 14:10). This result is in accordance with daily oscillations of cry1 and cry2 observed in the brain and pituitary gland of the European seabass Dicentrarchus labrax [19, 20] and the diencephalon of the goldlined spinefoot [10] and Malabar grouper Epinephelus malabaricus [21]. When the threespot wrasse Halichoeres trimaculatus was reared under constant dark conditions, the daily variation of cry1 in the brain was damped [22]. Therefore, some crys of teleost fish appear to be light responsive, as reported for crys of other vertebrates [23]. This possibility is supported by experiments showing that light exposure resulted in induction of cry1a and per2 in a zebrafish cell line and tissue [13, 24, 25]. This study also revealed that the expression of cry1 increased after lights on (acrophase around ZT6), while that of cry2 rose before lights off (acrophase around ZT10–14), suggesting that crys have different responses under LD. This is supported by reports that crys in the brain showed different expression patterns under LD [10, 19, 20]. In this regard, cry1 in the brain of the threespot wrasse oscillated under condition LL, but not DD, suggesting responsiveness to light stimuli [22]. In comparison, little daily variation of cry3 was observed in the diencephalon and pituitary gland of honeycomb grouper in this study, suggesting a function other than involvement in the circadian system. Alternatively, it is likely that cry3 in various nuclei oscillates differentially because nuclei with different functions are located in the diencephalon [26, 27]. Seasonal events may be related to the daily variation in cry3, as oscillations in cry1 and cry2 in relation to water temperature were marked in summer but not in winter in the European seabass [20]. However, seasonal effects may be eliminated in this study, as all tissue sampling was carried out during the spawning season (May to August), when water temperature is consistently high.
This study revealed that the mRNA abundances of three cry genes in the diencephalon and pituitary gland exhibit lunar cycle-aligned variations in samples collected weekly at 0000 h during sequential lunar phases. Interestingly, our results suggest at least two lunar cycle characteristics in cry transcript profiles in the brain. The first characteristic feature of lunar dependency is responsiveness in the diencephalon and pituitary gland to transient changes in moonlight; steady increases in mRNA abundances of three crys were observed clearly in the diencephalon and moderately in the pituitary gland from W01/W11 (first quarter moon) to W03/W13 (last quarter moon) (Fig. 3). This result shows that lunar cues such as the length and timing of “brightness at night” are possible inducers of crys in the brain. This possibility is supported by the present finding from fish reared under artificial full moon and new moon conditions (Fig. 4); the transcript levels of cry1 and cry2 in the diencephalon and of cry1 in the pituitary gland were upregulated at W2 (after 2 weeks of artificial moonlight exposure). A similar pattern of lunar phase-dependent expression of clock genes was reported in the goldlined spinefoot, in which the overall expression levels of cry1a and cry2 increased during the full moon and last quarter moon, respectively [10]. As other clock genes such as per2 and bmal2 are influenced by moonlight [10, 28], the circadian system in fish may successfully incorporate oscillations of the lunar cycle.
The second characteristic feature of lunar dependency is that the mRNA abundance of crys during three moon phases (from first quarter moon to last quarter moon) was lower than that at the new moon. Differences in the transcript levels of crys were remarkable in the pituitary gland and moderate in the diencephalon. Therefore, it is possible that “darkness at night” at the new moon provides a stable cue that acts as a key entrainer of phase setting for lunar synchronization in this species. The importance of “darkness at night” for transcription of clock genes appears notable in the goldlined spinefoot, in which the overall expression level of cry3 in the diencephalon was higher during the new moon and interruption of moonlight resulted in upregulation of cry1b in the diencephalon and cry2 in the pituitary gland [10]. The previous finding that lunar cycle-dependent variation with an increase at the new moon occurred in per1, but not per2, in the diencephalon of the goldlined spinefoot may support the importance of “darkness at night” for phase setting [9]. This study may claim that superimposing “moonlight” and “its change with moon phase” to “darkness at night” plays a role in oscillating clock genes. Notably, the transcript levels of cry1 (and possibly cry2) increased at W4, corresponding to the new moon, when fish were reared under artificial full moon conditions. Regarding the molecular mechanism of the lunar cycle in fish, one must also consider other genes that change in the brain in response to changes in moonlight levels because our transcriptomic analysis of the diencephalon of the honeycomb grouper found differentially expressed genes (DEGs) between the new and full moon period (Fig. S4). In addition, the expressions of some genes (per1, per2b, clock1b, and bmal1) in the diencephalon of goldlined spinefoot showed moon phase-dependency [10].
The ability to perceive differences in moonlight is essential to lunar periodicity, since the luminosity of moonlight is markedly lower than that of sunlight. In this regard, some studies have demonstrated that melatonin is a good indicator of moonlight perception [5, 6, 29, 30]. This indoleamine shows diel variations (day-low and night-high) in many fish [31, 32]. In addition to daily fluctuations, exposing spinefoot fish to brightness at the full moon and new moon lowered the plasma level of melatonin [5, 30] and the transcript level of melatonin receptor (Mel1c) in the pineal gland [29]. Moreover, melatonin synthesis was suppressed when the pineal gland of the goldlined spinefoot was isolated and cultured under full moon and new moon light conditions [6]. Perception of moonlight is likely in the honeycomb grouper, as its ocular melatonin content is influenced by moonlight [16]. The retina and pineal organ are major candidates for moonlight perception and transduction. The involvement of other organs in perceiving moonlight may not be excluded because peripheral organs (heart and kidney) of the zebrafish can respond to the environmental light-dark cycle, to which their clocks entrain [33]. The saccus vasculosus is another candidate because, in the masu salmon Oncorhynchus masou, this fish-specific organ contains photoreceptors and plays an important part in seasonal reproduction [34].
Several lines of evidence show that melatonin has an impact on the transcript levels of clock genes in mammals [35, 36]. Melatonin appears to have a similar effect in fish, as supplementing feed with melatonin during the daytime altered the transcript levels of cry1 and cry2 in the pituitary gland of European seabass within 3 h [20]. Thus, melatonin plays a role in synchronizing the expression of clock genes in the brain [37]. This study showed that the transcript levels of crys in the diencephalon and pituitary gland did not change within 7 h of intraperitoneal injection of melatonin. The discrepancy between these two studies may be partially due to experimental design or timing (season), as difficulty of replication across seasons has been noted in European seabass [20]. Of note, the expression of gonadotropins (lhβ) in the pituitary and gonadotropin-releasing hormone (gnrh1) in the diencephalon of the honeycomb grouper showed lunar-related fluctuation and were suppressed by melatonin implantation [16], although it is not clear whether crys mediates the role of melatonin in relation to lunar reproduction.
Conclusions
In conclusion, this study revealed daily and monthly oscillations in the transcript levels of clock genes in the brain of honeycomb grouper, with moonlight playing a crucial role in the induction and phase setting of their transcription (Fig. 6). Apart from photic changes in sunlight, changes in moonlight at night can be described by several factors including length, timing, and luminosity. This study indicates that interactions among these factors drive lunar synchrony, forming a putative hourglass-type lunar timer as proposed previously [10]. Further studies are needed to clarify where the master lunar clock is located in the brain and how it is used for timekeeping of lunar-synchronized spawning.

Possible regulation of lunar clock in the honeycomb grouper. (1) “Darkness at night” at the new moon resets lunar clock, (2) transient change in nocturnal light from the first quarter moon adjusts lunar clock
Methods
Animals and experimental design
Honeycomb grouper (n = 155; total length, 189.6 ± 2.51 mm; body weight, 93.9 ± 3.45 g) were collected from coral reefs around Sesoko Island (26°38′46″N, 127°51′54″E) and Ou-Jima (26°07′46″N, 127°46′22″E) using a hook and line in November 2014 and from May to July of 2016, 2018, and 2019. During the acclimation period, the fish were fed commercial pellets (Himesakura, Higashimaru, Kagoshima, Japan) daily. All experiments were conducted in compliance with the guidelines of the Animal Care and Use Committee of the University of the Ryukyus and regulations for the care and use of laboratory animals in Japan.
Fish (n = 2; average total length, 152.5 mm) used for molecular cloning were collected from coral reefs around Sesoko Island in November 2014. After anesthetization with 2-phenoxyethanol (Kanto Chemicals, Tokyo, Japan), total length and body weight were measured and the fish were decapitated. The brain was removed from the skull and immediately frozen with liquid nitrogen. The frozen tissues were stored at − 80 °C until use.
Daily expression profiles of crys in the diencephalon and pituitary tissues of honeycomb grouper were assessed in Experiment 1. Fish (n = 30; total length, 182.3 ± 4.83 mm; body weight, 78.2 ± 6.69 g) collected around Ou-Jima in July 2019 were housed in indoor tanks (150-L capacity, polypropylene square tank 160 cm × 100 cm × 20 cm) with running seawater and aeration under a programmable photoperiod (LD = 14:10, lights on 0600 h; lights off 2000 h) at Sesoko Station, Tropical Biosphere Research Center, University of the Ryukyus. Three fluorescent lamps (each 20 W) were placed above the tank and the light intensity at the water surface was 1020 lx. After acclimation for 1 month, fish were randomly taken from the tanks at zeitgeber time (ZT) 9, 13, 17, 21, 1 (25), and 5 (29), and anesthetized with 2-phenoxyethanol. After measuring the total length and body weight, the fish were sacrificed by decapitation. The brain was removed from the skull, immersed in RNA later (Sigma-Aldrich, St. Louis, MO, USA) and the diencephalon was separated. The diencephalon and pituitary tissues were placed in 1.5 mL tubes containing RNAiso Plus (Takara Bio, Kusatsu, Japan), immediately frozen in liquid nitrogen, and stored at − 80 °C until use. Tissue was collected at ZT17 and ZT21 under a dim red light (XP-G-R5, Cree, Durham, NC, USA).
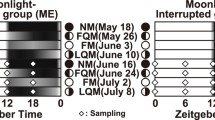
Weekly expression profiles of crys in the diencephalon and pituitary tissues of the honeycomb grouper were assessed in Experiment 2. Fish (n = 69; total length, 191.4 ± 3.18 mm; body weight, 97.6 ± 4.40 g) were collected around Ou-Jima during the first quarter moon (W01/W11), full moon (W02/W12), last quarter moon (W03/W13), and new moon of May to July 2016, and temporarily held in outdoor tanks (200-L capacity, polypropylene round tank height 75.5 cm × diameter 55.0 cm) at Sesoko Station with running seawater under natural sunlight and moonlight. After anesthetization with 2-phenoxyethanol, brains were collected at ZT18 under a dim red light and immersed in RNAlater. The diencephalon and pituitary tissues were collected and preserved as described above.
The effect of nocturnal moonlight on the transcript levels of crys in the brain was examined (Experiment 3). Fish (n = 40; total length, 200.9 ± 5.79 mm; body weight, 108.9 ± 8.31 g) were collected around Ou-Jima, housed in tanks (150-L capacity, polypropylene square tank 160 cm × 100 cm × 20 cm) at Sesoko Station with running seawater and fed daily at 1000 h with commercial pellets (Himesakura). After acclimation for 2 months under LD conditions (LD = 14:10) without moonlight signals, the fish were divided into two groups; fish in the control group (n = 20) were kept under LD conditions, while those in the moonlight group (n = 20) were exposed to a red fluorescent lamp (15 lx at the water surface, 2261 K, λp = 658 nm, S-3920, SUDO, Nagoya, Japan) during scotophase, based on a previous study [10]. After the fish were reared under the experimental conditions without additional moon signals, samples were collected on 3, 9, 17, and 25 July, 2019. Fish in both groups were removed from the tank at ZT18, anesthetized with 2-phenoxyethanol, and decapitated. The brain was removed from the skull and the diencephalon and pituitary gland were separated and immersed in RNAlater. Samples were collected out under a dim red light.
The effect of melatonin treatment on the expression of cry genes in the brain was examined during the spawning season (Experiment 4). Fish used in this experiment (n = 14; total length, 167.7 ± 7.55 mm; body weight, 67.1 ± 7.78 g) were collected from coral reefs around Ou-Jima in August 2018 and transported to Sesoko Station. The fish were acclimatized in an indoor tank (200-L capacity, polypropylene round tank height 75.5 cm × diameter 55.0 cm; programmable photoperiod, LD = 13:11, lights on 0600 h, lights off 1900 h) with running seawater and aeration for 1 week.
Melatonin was purchased from Sigma-Aldrich, dissolved in ethanol, and diluted to 1 mg/mL with saline. Fish were anesthetized with 2-phenoxyethanol at ZT12 in September 2018. Melatonin (500 ng/g) was intraperitoneally injected into fish of the treatment group (n = 7; MT+), while only vehicle (saline containing 1% ethanol) was injected into fish of the control group (n = 7; MT-). Fish of both groups were housed separately in aquariums (60-L capacity) with running seawater. At ZT19 (7 h after treatment), fish in both groups were decapitated following anesthetization with 2-phenoxyethanol, and subjected to sample collection as described above.
Molecular procedures
Total RNA was extracted from diencephalon and pituitary tissues using RNAiso Plus (Takara Bio), according to the manufacturer’s protocol, and reverse-transcribed using the Prime Script RT reagent kit with gDNA Eraser (Takara Bio) to obtain single stranded cDNA. Degenerate primers for the partial cloning of crys from the honeycomb grouper were designed using the cry sequences of the zebrafish (AB042248.1, AB042249.1, and AB042252.1), threespot wrasse (HQ893881.1), medaka Oryzias latipes (XM_004070156), Atlantic salmon Salmo salar (XM_014125353.1), goldlined spinefoot (AB643455.1 and AB643456.1), and Japanese pufferfish Takifugu rubripes (XM_011617614.1) obtained from GenBank using Primer3Plus (ver. 2.4.2) (Table 1). The cDNA fragments encoding crys were amplified using polymerase chain reaction (PCR) performed in a volume of 12.5 μL containing 50% GoTaq Green Master Mix (Promega, Madison, WI, USA), 0.3 μM each of forward and reverse primers, and 4% template cDNA at 95 °C for 3 min; 40 cycles at 95 °C for 45 s, 60 °C for 45 s, and 72 °C for 1–2 min; and finally 72 °C for 5 min. The PCR products were electrophoresed in a 2% agarose gel containing ethidium bromide. Bands of the predicted size were purified using the Wizard SV Gel and PCR Clean-Up System (Promega) and subcloned into pGEM-T Easy Vector (Promega). The inserted fragment was sequenced using a 3730xl DNA Analyzer (Applied Biosystems, Waltham, MA, USA). Its putative amino acid sequence was deduced using ORF Finder (NCBI; http://www.ncbi.nlm.nih.gov/projects/gorf/) and the Pfam database (https://pfam.xfam.org/). BLAST (https://blast.ncbi.nlm.nih.gov/blast.cgi) was used to identify the isolated sequence. Multiple alignment and phylogenetic analysis were performed using MEGA-X [38] with the partial sequences of crys. The neighbor-joining method was used to generate a phylogenetic tree with bootstrap confidence values based on 1000 replicates [39, 40].
Primers for measuring cry1, cry2, cry3, β-actin, and elongation factor 1 alpha (ef1α) in the diencephalon and pituitary tissues of the honeycomb grouper were designed using Primer3Plus (ver. 2.4.2) (Table 1). Efficiency (E) and correlation coefficient (R2) values obtained for each assay through primer checking are summarized in Additional file 2: Table S1. After RNA quality was determined through agarose electrophoresis, samples with distinct ribosomal bands were selected for further analysis using quantitative PCR (qPCR), comprising 95 °C for 2 min and 40 cycles of 95 °C for 15 s and 60 °C for 1 min, followed by melting curve analysis. The mRNA abundances of these genes in the diencephalon and pituitary tissues were determined using the CFX96 Real Time System (Bio-Rad, Hercules, CA, USA) and TB Green Premix Ex Taq (Takara Bio). Expression levels were normalized using the ΔΔCt method with β-actin or ef1α. All qPCR experiments were performed in duplicate.
Statistical analysis
All data are shown as the mean ± standard error of the mean. The source data are included in Additional file 3. Data were analyzed using one-way analysis of variance followed by Tukey’s multiple comparison test for parametric data and the Kruskal–Wallis test combined with the Bonferroni post-hoc test or Steel–Dwass post-hoc test for non-parametric data. Acro (http://www.circadian.org/softwar.html) was used to detect the acrophase in the daily changes in crys, the data for which are shown as the mean ± 95% confidence interval. Dunnett’s multiple comparison tests against the new moon for the other moon phases were used in Experiment 2. Student’s t-test was used to compare data between the moonlight control and treatment groups in Experiment 3 and between MT- and MT+ in Experiment 4. Statistical significance was considered to be P < 0.05. All statistical analyses were performed using R ver. 3.4.2 and the analysis data are included in Additional file 4.
Availability of data and materials
Protein and RNA sequences of crys had been registered. The accession numbers (protein: BBZ90056-BBZ90058, mRNA: LC521334-LC521336) were assigned. The data obtained by qPCR in this study are included in the Additional files 3 and 4. All raw data required to evaluate the present study are deposited in University of the Ryukyus and can be acquired from the corresponding author.
Abbreviations
- β-actin:
-
Beta actin
- Bmal2:
-
Brain and muscle ARNT-like protein-2
- bp:
-
Base pair
- Ct:
-
Cycle threshold
- Cry:
-
Cryptochrome
- cDNA:
-
Complementary deoxyribonucleic acid
- DNA:
-
Deoxyribonucleic acid
- Ef1α:
-
Elongation factor alpha 1
- JSPS:
-
Japan Society for the Promotion of Science
- K:
-
Kelvin
- LD:
-
Light and dark
- λp:
-
Peak wavelength
- lx:
-
Lux
- mRNA:
-
Messenger ribonucleic acid
- MT-:
-
Melatonin not-injected group
- MT+:
-
Melatonin injected group
- ng:
-
Nanograms
- Per2:
-
Period2
- qPCR:
-
Real-time quantitative polymerase chain reaction
- ZT:
-
Zeitgeber time
References
Johannes RE. Reproductive strategies of coastal marine fishes in the tropics. Environ Biol Fish. 1978;3:65–84.
Hoque MM, Takemura A, Matsuyama M, Matsuura S, Takano K. Lunar spawning in Siganus canaliculatus. J Fish Biol. 1999;55:1213–22.
Park Y-J, Takemura A, Lee Y-D. Annual and lunar-synchronized ovarian activity in two rabbitfish species in the Chuuk lagoon. Micronesia Fish Sci. 2006;72:166–72.
Rahman MS, Takemura A, Takano K. Correlation between plasma steroid hormones and vitellogenin profiles and lunar periodicity in the female golden rabbitfish, Siganus guttatus (Bloch). Comp Biochem Physiol Part B Biochem Mol Biol. 2000;127:113–22.
Takemura A, Susilo ES, Rahman MDS, Morita M. Perception and possible utilization of moonlight intensity for reproductive activities in a lunar-synchronized spawner, the golden rabbitfish. J Exp Zool. 2004;301:844–51.
Takemura A, Ueda S, Hiyakawa N, Nikaido Y. A direct influence of moonlight intensity on changes in melatonin production by cultured pineal glands of the golden rabbitfish, Siganus guttatus. J Pineal Res. 2006;40:236–41.
Levy O, Appelbaum L, Leggat W, Gothlif Y, Hayward DC, Miller DJ, Hoegh-Guldberg O. Light-responsive cryptochromes from a simple multicellular animal, the coral Acropora millepora. Science. 2007;318:467–70.
Fukushiro M, Takeuchi T, Takeuchi Y, Hur S-P, Sugama N, Takemura A, Kubo Y, Okano K, Okano T. Lunar phase-dependent expression of cryptochrome and a photoperiodic mechanism for lunar phase-recognition in a reef fish, goldlined spinefoot. PLoS One. 2011;6:e28643.
Toda R, Okano K, Takeuchi Y, Yamauchi C, Fukushiro M, Takemura A, Okano T. Hypothalamic expression and moonlight-independent changes of Cry3 and Per4 implicate their roles in lunar clock oscillators of the lunar-responsive goldlined spinefoot. PLoS One. 2014;9:e109119.
Takeuchi Y, Kabutomori R, Yamauchi C, Miyagi H, Takemura A. Moonlight controls lunar-phase-dependency and regular oscillation of clock gene expressions in a goldlined spinefoot. Sci Rep. 2018;8:1–12.
Takemura A, Rahman MS, Park Y-J. External and internal controls of lunar-related reproductive rhythms in fishes. J Fish Biol. 2010;76:7–26.
Cashmore AR. Cryptochromes: blue light receptors for plants and animals. Science. 1999;284:760–5.
Tamai TK, Young LC, Whitmore D. Light signaling to the zebrafish circadian clock by Cryptochrome 1a. Proc Natl Acad Sci U S A. 2007;104:14712–7.
Lee Y-D, Park S-H, Takemura A, Takano K. Histological observations of seasonal reproductive and lunar-related spawning cycles in the female honeycomb grouper Epinephelus merra in Okinawan waters. Fish Sci. 2002;68:872–7.
Soyano K, Masumoto T, Tanaka H, Takushima M, Nakamura M. Lunar-related spawning in honeycomb grouper, Epinephelus merra. Fish Physiol Biochem. 2003;28:447–8.
Fukunaga K, Yamashina F, Ohta N, Mizuno H, Takeuchi Y, Yamauchi C, Takemura A. Involvement of melatonin in transducing moon-related signals into the reproductive network of the female honeycomb grouper Epinephelus merra. Gen Comp Endocrinol. 2019;282:113211.
Kobayashi Y, Ishikawa T, Hirayama J, Daiyasu H, Kanai S, Toh H, Fukuda I, Tsujimura T, Terada N, Kamei Y, Yuba S, Iwai S, Todo T. Molecular analysis of zebrafish photolyase/cryptochrome family: two types of cryptochromes present in zebrafish. Genes Cells. 2000;5:725–38.
Ishikawa T, Hirayama J, Kobayashi Y, Todo T. Zebrafish CRY represses transcription mediated by CLOCK-BMAL heterodimer without inhibiting its binding to DNA. Genes Cells. 2002;7:1073–86.
del Pozo A, Vera LM, Sánchez JA, Sánchez-Vázquez FJ. Molecular cloning, tissue distribution and daily expression of cry1 and cry2 clock genes in European seabass (Dicentrarchus labrax). Comp Biochem Physiol A. 2012;163:364–71.
Herrero MJ, Lepesant JMJ. Daily and seasonal expression of clock genes in the pituitary of the European sea bass (Dicentrarchus labrax). Gen Comp Endocrinol. 2014;208:30–8.
Yamashina F, Takeuchi Y, Fukunaga K, Udagawa S, Tan ES, Byun J-H, Yamauchi C, Takemura A. Daily expression of a clock gene in the brain and pituitary of the Malabar grouper (Epinephelus malabaricus). Gen Comp Endocrinol. 2019;280:9–14.
Hur S-P, Takeuchi Y, Itoh H, Uchimura M, Takahashi K, Kang H-C, Lee Y-D, Kim S-J, Takemura A. Fish sleeping under sandy bottom: interplay of melatonin and clock genes. Gen Comp Endocrinol. 2012;177:37–45.
Sancar A. Regulation of the mammalian circadian clock by cryptochrome. J Biol Chem. 2004;279:34079–82.
Cermakian N, Pando MP, Thompson CL, Pinchak AB, Selby CP, Gutierrez L, Wells DE, Cahill GM, Sancar A, Sassone-Corsi P. Light induction of a vertebrate clock gene involves signaling through blue-light receptors and MAP kinases. Curr Biol. 2002;12:844–8.
Moore HA, Whitmore D. Circadian rhythmicity and light sensitivity of the zebrafish brain. PLoS One. 2014;9:e86176.
Yasuo S, Watanabe M, Okabayashi N, Ebihara S, Yoshimura T. Circadian clock genes and photoperiodism: comprehensive analysis of clock gene expression in the mediobasal hypothalamus, the suprachiasmatic nucleus, and the pineal gland of Japanese quail under various light schedules. Endocrinology. 2003;144:3742–8.
Ikegami K, Katou Y, Higashi K, Yoshimura T. Localization of circadian clock protein BMAL1 in the photoperiodic signal transduction machinery in Japanese quail. J Comp Neurol. 2009;517:397–404.
Sugama N, Park J-G, Park Y-J, Takeuchi Y, Kim S-J, Takemura A. Moonlight affects nocturnal Period2 transcript levels in the pineal gland of the reef fish Siganus guttatus. J Pineal Res. 2008;45:133–41.
Park Y-J, Park J-G, Takeuchi Y, Hur S-P, Lee Y-D, Kim S-J, Takemura A. Influence of moonlight on mRNA expression patterns of melatonin receptor subtypes in the pineal organ of a tropical fish. Mar Genomics. 2014;14:67–70.
Rahman MS, Kim B-H, Takemura A, Park C-B, Lee Y-D. Effects of moonlight exposure on plasma melatonin rhythms in the seagrass rabbitfish, Siganus canaliculatus. J Biol Rhythms. 2004;19:325–34.
Bromage N, Porter M, Randall C. The environmental regulation of maturation in farmed finfish with special reference to the role of photoperiod and melatonin. Aquaculture. 2001;197:63–98.
Falcón J, Migaud H, Muñoz-Cueto JA, Carrillo M. Current knowledge on the melatonin system in teleost fish. Gen Comp Endocrinol. 2010;165:469–82.
Whitmore D, Foulkes NS, Sassone-Corsi P. Light acts directly on organs and cells in culture to set the vertebrate circadian clock. Nature. 2000;404:87–91.
Nakane Y, Ikegami K, Iigo M, Ono H, Takeda K, Takahashi D, Uesaka M, Kimijima M, Hashimoto R, Arai N, Suga T, Kosuge K, Abe T, Maeda R, Senga T, Amiya N, Azuma T, Amano M, Abe H, Yamamoto N, Yoshimura T. The saccus vasculosus of fish is a sensor of seasonal changes in day length. Nat Commun. 2013;4:2108.
Dardente H, Menet JS, Poirel VJ, Streicher D, Gauer F, Vivien-Roels B, Klosen P, Pévet P, Masson-Pévet M. Melatonin induces Cry1 expression in the pars tuberalis of the rat. Mol Brain Res. 2003;114:101–6.
Hazlerigg DG, Andersson H, Johnston JD, Lincoln G. Molecular characterization of the long-day response in the soay sheep, a seasonal mammal. Curr Biol. 2004;14:334–9.
Von Gall C, Weaver DR, Moek J, Jilg A, Stehle JH, Korf HW. Melatonin plays a crucial role in the regulation of rhythmic clock gene expression in the mouse pars tuberalis. Ann N Y Acad Sci. 2005;1040:508–11.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35:1547–9.
Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783.
Naruya S, Masatoshi N. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–25.
Acknowledgements
We gratefully acknowledge to staff of Sesoko Station, Tropical Biosphere Research Center, University of the Ryukyus, Okinawa, Japan, for use of facilities and assistance of maintaining of animals used in this study.
Funding
KF was supported by a Grant-in-Aid for JSPS Research Fellow to KF (KAKENHI Grant Number JP18J20958). This funding contributed to support costs for collecting fish from the nature, housing of fish, and molecular experiments such as molecular cloning and real time qPCR assay.
Author information
Authors and Affiliations
Contributions
KF designed and carried out the entire experiments and analyzed all the data obtained in the present study. FY and YT contributed to prepare samples and accomplish experiments. CY contributed to store fish used in this study. They participated in preparing the manuscript. AT was a collaborator and supervisor in analyzing the data and writing the manuscript. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All experiments in the present study were conducted in compliance with the guidelines of the Animal Care and Use Committee of the University of the Ryukyus and regulations for the care and use of laboratory animals in Japan and approved by the relevant facilities at Sesoko Station, University of the Ryukyus (#22973–2018 and #24657–2019). Collection of specimens used in the present study was conducted in accordance with local legislation.
Consent for publication
Not applicable.
Competing interests
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Fig. S1.
Nucleotide (top line) and deduced amino acid (bottom line) sequences (A), and multiple alignment of deduced amino acid sequence (B) of honeycomb grouper cry1. The number of nucleotide and amino acid are indicated on the right. Candidate region of FAD binding domain (PF03441) are underlined with single line. The asterisks indicate the fully conserved residues between proteins. Fig. S2. Nucleotide (top line) and deduced amino acid (bottom line) sequences (A), and multiple alignment of deduced amino acid sequence (B) of honeycomb grouper cry2. The number of nucleotide and amino acid are indicated on the right. Candidate region of FAD binding domain (PF03441) are underlined with single line. The asterisks indicate the fully conserved residues between proteins. Fig. S3. Nucleotide (top line) and deduced amino acid (bottom line) sequences (A), and multiple alignment of deduced amino acid sequence (B) of honeycomb grouper cry3. The number of nucleotide and amino acid are indicated on the right. Candidate region of FAD binding domain (PF03441) are underlined with single line and DNA photolyase domain (PF00875) is underlined with dotted line. The asterisks indicate the fully conserved residues between proteins. Fig. S4. RNA-seq analysis in the hypothalamus of honeycomb grouper. Differentially expressed genes (DEGs) between the new and full moon period (A). Red and blue lines show upregulated and downregulated DEGs, respectively. Gene ontology (GO) annotation is indicated beside on the plots summarized (B). Venn diagram of the number of GO annotation in the upregulated and downregulated DEGs (C).
Additional file 2: Table S1.
Summary of the primer check analysis in each assay of real-time PCR.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Fukunaga, K., Yamashina, F., Takeuchi, Y. et al. Moonlight is a key entrainer of lunar clock in the brain of the tropical grouper with full moon preference. BMC Zool 5, 11 (2020). https://doi.org/10.1186/s40850-020-00060-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40850-020-00060-8