
Abstract
Background
Different parts of the medicinal plant Tamarindus indica L. are full of phytochemicals that are able to reduce elevated blood pressure, blood sugar and lipids. These pharmacological effects are due to the presence of antioxidant type compounds in those parts of the plant. This study was aimed to explore the molecular mechanism of anti-obesity effects of ethanolic extract of T. indica L. leaves (TILE) through the evaluation of biochemical parameters and gene expression analysis in high-fat diet (HFD) consuming Wistar rats.
Methods
Male Wistar rats were supplied with a standard diet (SD), or HFD, or HFD with 100 mg/kg or 200 mg/kg or 400 mg/kg TILE for 8 weeks. The body weight, liver weight, fat weight, plasma lipids, and oxidative stress-related parameters were measured. The transcript levels of different adipogenesis related transcription factors, lipogenic enzymes, and lipolytic enzymes were also evaluated by quantitative real-time PCR.
Result
Phytochemical analysis demonstrated that TILE is enriched with a substantial level of polyphenols (287.20 ± 9.21 mg GAE/g extract) and flavonoids (107.52 ± 11.12 mg QE/g extract) which might be the reason of significant antioxidant and radical scavenging activities. Feeding of TILE (400 mg/kg/day) to HFD-fed rats increased activity of superoxide dismutase and catalase which is reflected as a significant reduction of oxidative stress markers like nitric oxide and malondialdehyde. TILE (400 mg/kg/day) feeding also down-regulated the mRNA levels of proadipogenic transcription factors including liver X receptor alpha (LXRα), peroxisome proliferator-activated receptor gamma (PPARγ), and sterol regulatory element-binding protein 1c (SREBP1c) in diet-induced obese rats. As a consequence of this, the mRNA level of lipogenic enzymes like acetyl-CoA carboxylase (ACC), fatty acid synthase (FAS), diacylglycerol acyltransferase (DGAT), and HMG-CoA reductase was down-regulated with a parallel up-regulation of the transcript level of lipolytic enzyme, hormone-sensitive lipase (HSL).
Conclusion
Observations from this study indicate that antioxidant-rich TILE can reduce HFD-induced body weight, fat weight and liver weight as well as blood lipids through down-regulating the gene expression of proadipogenic transcription factors and lipogenic enzymes with a concerted diminution of the gene expression of lipolytic enzyme, HSL.
Similar content being viewed by others

Introduction
Obesity, hypercholesterolemia and associated cardiovascular diseases contribute to a high percentage of morbidity and mortality in humans, making them major areas of public health interest [1]. While the synthesis of lipid-lowering drugs has been made possible in the last few decades through a plethora of scientific endeavors, in many cases, their adverse effects outweigh their benefits. For this reason, in last few decades, research interest has diverted towards the study of natural compounds such as medicinal plants, for the potential discovery of phytochemicals with the efficacy to lower lipid and glucose levels. Since the beginning of human civilization, various plant parts including flowers, fruits, leaves, roots, and barks are being used in virtually all cultures as a medicinal source due to their efficacy in treating various disorders [2]. For example, Tamarindus indica L., (family Fabaceae) is a common plant of tropical countries with medicinal, commercial and dietary importance. The brown sticky pulp of T. indica fruit is a very popular culinary additive in many recipes of India due to its sour and sweet taste which is attributed to the high concentration of tartaric acid and reducing sugars [3]. Historically, all across the world, almost every part of T. indica, but especially its fruits, leaves, seeds, and bark have been used as traditional medicine with several pharmacological applications [4]. Studies have shown that these parts of T. indica contain many compounds of therapeutic interest, including polyphenolic antioxidant molecules, caffeic acid and vitamin C [5]. For example, the ethanolic extract from the fruit’s pericarp reduced total and non-HDL cholesterol as well as triglyceride levels, while increased HDL-cholesterol levels in hamsters, as a result of its antioxidant activities [6]. The ethyl acetate and ethanol extracts of T. indica seed coat have been reported to possess antioxidant [7] and anti-inflammatory properties [8]. In a clinical trial, the powdered pulp of T. indica fruit was found to lower effectively the level of LDL-cholesterol and total cholesterol [9]. Moreover, in another study, the extract of T. indica seed was reported as effective in lowering blood sugar levels in hyperglycemic male rats [10].
In the pathophysiology of adipogenesis, both dietary and genetic factors play very important roles. Among the genetic factors, liver X receptor alpha (LXRα) plays a critical role by regulating the gene expression of two important transcription factors related to adipogenesis like peroxisome proliferators-activated receptor gamma (PPARγ), and sterol regulatory element-binding protein 1c (SREBP 1c) [11]. PPARγ and CCAAT enhancer-binding protein alpha (C/EBPα) controls the expression of down-stream adipocyte specific proteins and thus promotes the commitment of preadipocytes to lipid-laden matured adipocytes [12, 13]. On the other hand, SREBP1c controls the gene expression of vital lipogenic enzymes acetyl-CoA carboxylase (ACC), fatty acid synthase (FAS), and HMG-CoA reductase which are involved in the biosynthesis of de novo fatty acids, triacylglycerol (TG), and cholesterols [14,15,16]. Hence, any promising therapeutic intervention having antiobesity effect warrants the investigation of the gene expression of the abovementioned factors and enzymes to decode the molecular mechanism of action. Previous clinical and preclinical studies, through the investigation of biochemical parameters, have reported that different parts of T. indica L. have anti-oxidant, lipid-lowering [17], blood pressure and blood glucose-lowering properties [4]. However, there is a lack of information about the efficacy of T. indica L. leaf extract in regulating the expression of transcription factors that control adipocyte differentiation and enzymes which participate in lipid metabolism. Without exploring the gene expression of these abovementioned transcription factors and enzymes, the molecular mechanism of T. indica L. leaf extract would remain obscure. This study was designed to explore the effects of ethanolic extract of T. indica L. leaf on high-fat diet-fed obese Wistar rats by investigating biochemical parameters and gene expression pattern of transcription factors perform vital functions in the differentiation of adipocytes and enzymes involved in the lipid metabolism.
Materials and methods
Chemicals and reagents
5,5′-dithio-bis-[2-nitrobenzoic acid] (DTNB), sulfanilamide, N-ethyl diaminedihydrochloride (NED), nitrobluetetrazolam (NBT), thiobarbituric acid (TBA), 2,2-Diphenyl-1-picrylhydrazyl (DPPH), trichloroacetic acid (TCA), gallic acid, quercetin, and trolox were purched from Sigma Aldrich (Massachusetts, USA). Kits for the assay of serum triglyceride, total cholesterol, and HDL cholesterol were of Plasmatec (Stafford, UK). GeneJET RNA Purification Kit, RevertAid First Strand cDNA Synthesis Kit and SYBR Green qPCR Master Mix were from Thermo-Fisher Scientific (Massachusetts, USA). Other chemicals used in different experiments were of high purity and of analytical grade.
Tamarindus indica L. leaf-extract preparation
Mature leaves of Tamarindus indica L. was collected from localities in Bangladesh in the mid of July. The plant species was confirmed by a botanical taxonomist of Bangladesh National Herbarium, Mirpur, Dhaka, Bangladesh (Accession No. 55785). The leaves were oven-dried at 40 °C and pulverized into fine particles using a laboratory mill. Each portion of 100 g powder was immersed in 500 ml ethanol for 7 days with continuous shaking and then filtered by Whatman filter paper (No. 41). The residue was extracted once again and filtered in the same way. All the filtrate was collected and condensed at 60 °C in a rotary evaporator and then dried under vacuum. In this way, 6.42 g dried extract was prepared from each of the 100 g dried powder of T. indica leaf. The collected extract was kept in the refrigerator at − 4 °C for further use.
Quantification of total phenolic and flavonoid content in T. indica L. leaf extract
In order to measure the phenolic and flavonoid content 50 mg crude extract was mixed with 5 ml of distilled water and the resulting mixture was further diluted with distilled water get a final concentration of 0.5 mg/ml. Folin-Ciocalteu method was employed to measure the extract’s phenolic content [18]. Briefly, 100 μL leaf extract, 500 μL of Folin-Ciocalteu reagent and 400 μL of 7.5% sodium bicarbonate were mixed according to the mentioned sequence and the resulting mixture was incubated at 40 °C for 20 min. Absorbance was taken at 755 nm using a spectrophotometer against a blank and total phenol content was calculated in mg/g using a standard curve of gallic acid. The flavonoid content was assayed by mixing 1 ml crude extract with 4 ml of deionized water and 0.3 ml of 5% NaNO2 solution. After 5 min, 3 ml of 2% AlCl3 solution was mixed which was followed by the addition of 2 ml of NaOH (1 M) solution. Then it was mixed properly and absorbance was recorded at 510 nm against a blank. The amount of total flavonoid was estimated as μg/g using quercetin as standard [19].
Assessment of antioxidant and radical scavenging capacity
The ferric reducing antioxidant power (FRAP) of the extract was carried out spectrophotometrically (at 593 nm), according to the method used by Pulido et al. [20]. The results are expressed as μmole Fe++/g dry extract. Radical scavenging property of T. indica L. leaf extract was quantified based on its ability to decolorize ABTS+ [2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)] cation. The absorbance was recorded at 734 nm and antioxidant activity was calculated using the equation obtained from trolox standard curve and was expressed as mmol Trolox/g of the dry extract [21].
The 2,2-diphenyl-1-1picrylhydrazyl (DPPH•) radical-scavenging activity of the extract was measured according to the method of Braca et al. [22]. Briefly, leaf extract of different concentrations were added to 0.004% (w/v) solution of DPPH. The absorbance values were measured at 517 nm after the mixtures were incubated for 30 min in a light protected container. The percentage inhibition of activity was determined using the equation-
Where, A0 is the absorbance of the control, and AE is the absorbance of the leaf extract or of the standards. The inhibition curve was prepared by plotting percent inhibition against concentration and median inhibitory concentration (IC50) values were calculated.
Experimental design, animal feeding and diet
Healthy 9–10 weeks old male Wistar rats (180 ± 10 g body weight) were collected from the Animal Care Unit, Department of Pharmaceutical Sciences, North South University, Bangladesh. They were housed in individual cages which were kept in a room of standard environmental conditions (temperature 23 ± 2 °C, and 12 h light/dark cycle) with uninterrupted access to food and water. Moreover, for ensuring their comfort and to reduce suffering all the protocols used in this study was approved by the Ethical Review Committee of the Department of Pharmaceutical Sciences, North South University, Bangladesh (ACE-0017-2018). Rats were organized into 5 groups randomly and each group consisted of 7 rats.
-
Group-1 (SD): Fed with standard laboratory diet (SD)
-
Group-2 (HFD): Fed with high-fat diet
-
Group-3 (HFD + TILE100): Fed with HFD and T. indica leaf extract (100 mg/kg/day)
-
Group-4 (HFD + TILE200): Fed with HFD and T. indica leaf extract (200 mg/kg/day)
-
Group-5 (HFD + TILE400): Fed with HFD and T. indica leaf extract (400 mg/kg/day)
The formula and preparation method of the standard laboratory diet (SD) and high-fat diet (HFD) were obtained from a previous report [23]. Food intake and change of body weight were calculated according to a method described earlier [24].
Collection of blood, adipose tissues, and liver
The rats were sacrificed after 8 week of treatment and blood was collected by from the heart. Collected blood was centrifuged (8000×g) for 15 min at 4 °C for separation of serum which was immediately stored at − 20 °C for further experiment. The liver was carefully cleared and adhering adipose tissue was separated and properly weighed. The visceral fat adhered to different internal organs were collected carefully, rinsed with phosphate-buffered saline, and then weights were measured. Weight of the liver and different adipose tissues were expressed in relation to per 100 g of body weight according to the method described earlier [25].
Measurement of plasma lipid-related parameters
The lipid parameters in the samples including total cholesterol (TC), triacylglycerol (TG), and high-density lipoprotein cholesterol (HDL-C) were measured by the diagnostic kits of Plasmatec Laboratories, (Allington, UK). The concentration of very-low-density lipoprotein cholesterol (VLDL-C) and low-density lipoprotein cholesterol (LDL-C) was calculated by Firedewald formula [26]:
Moreover, atherogenic index (AI) and coronary risk index (CRI) were calculated by the formula [27].
Measurement of oxidative stress-related parameters and antioxidant enzymes’ activity
Lipid peroxidation was assessed by measuring the level of malondialdehyde based on its ability to interact with thiobarbituric acid which can be detected and quantified spectrophotometrically at 532 nm [28]. Nitric oxide (NO) level was measured by mixing the samples collected from blood and liver with Griess reagent which contains 1.5% sulfanilamide in 1 N HCl and 1% N-(1-)naphthylethylenediamine (NED) in H2O. The resulting pink colored mixture was evaluated by a spectrophotometer (at 548 nm) and the concentration of NO was calculated by a standard curve of sodium nitrite [29]. The activity of superoxide dismutase was measured according to the process developed by Kakkar et al. with some customizations [30]. Briefly, the reaction mixture contained 25 mM sodium phosphate buffer, 0.186 mM phenazine methosulphate and 0.3 mM nitroblue tetrazolium with 0.1 ml of sample collected from blood or liver. Then 0.7 mM reduced nicotinamide adenine dinucleotide (NADH) was added and the mixture was kept at 30 °C for 90 s. After that, it was mixed with 1.0 ml glacial acetic acid and 4 ml n-butanol and then the mixture was let stand for 10 min before centrifugation. Absorbance of the collected supernatant was taken at 560 nm against n-butanol blank. The activity of catalase was measured by mixing the plasma sample to 30 mM H2O2 and 50 mM phosphate buffer (pH 7.0) and by recording the absorbance at 230 nm according to a previously described method [31].
Gene expression of transcription factors and fat metabolizing proteins
Total mRNA, from the mesenteric adipose tissue sample, was extracted by GeneJET RNA Purification Kit (Thermo-Fisher Scientific, Massachusetts, USA) for checking the transcript level of adipocyte-specific marker genes. After measuring the concentration, 1 μg mRNA was used in reverse transcription reaction for synthesizing cDNA using RevertAid First Strand cDNA Synthesis Kit (Thermo-Fisher Scientific, USA) in a Thermal Cycler (Takara Bio Inc., Ohtsu, Japan). Using this cDNA as a template, the transcript level of adipogenesis related transcription factors and enzymes were quantified by real-time PCR using 2X SYBR Green qPCR Master Mix (Thermo Scientific, USA) and forward and reverse primers designed by Primer3 online software (Table 1). The PCR reaction was conducted in a CFX96 TouchTM Real-Time PCR Detection System (Bio-Rad Laboratories Inc., California, USA) and data were analyzed by CFX ManagerTM software (Bio-Rad, California, USA). The PCR was carried out at 95 °C for 1 min followed by 40 cycles for amplification: 95 °C (5 s) for denaturation, 60 °C (30 s) for annealing, and 72 °C (1 min) for synthesis which was followed by the final extension at 72 °C for 5 min. The transcript level of each of the target protein was relatively quantified by normalization to the transcript level of β-actin.
Histology of hepatic tissue
A suitable section of the liver from a representative rat of each group was collected, after measuring the weight the liver, and was kept in neutral buffered formalin (pH -7.4). For visualization and estimation of lipid accumulation within the hepatocytes, the liver tissue from a particular group of the rat was embedded on paraffin, cut as thin layer (5 μm) with a microtome, was stained with haematoxylin and eosin (H&E) according to standard histological process described earlier [32]. Photomicrograph was taken using a differential interference optical microscope (Model-ZEISS-AXIO, Carl Zeiss Microimaging, Jena Germany). Lipid area percentages were quantified from a different group of animals using ImageJ software (version 1.52a).
Statistical analysis
All phytochemical analyses were done in triplicate (n = 3), biochemical and gene expression analyses were done in sextuplicate (n = 6) and the data were expressed as mean ± SEM. For the determination of significant change among different groups, data were interpreted by one-way ANOVA and Newman-Kuels post hoc test by Graph Pad Prism.
Results
Quantification of polyphenols, flavonoids content and evaluation of relative antioxidant property of T. indica L. leaf extract
The antioxidant property of T. indica leaf extract (TILE) was close to that of butylated hydroxytoluene (BHT) which might be attributed to its higher level of polyphenols (287.20 ± 9.21 mg GAE/g extract) and flavonoids (107.52 ± 11.12 mg QE/g extract). But the antioxidant activity of TILE was significantly lower than that of quercetin and ascorbic acid as was revealed in DPPH and ABTS radical scavenging activities and ferric reducing activity (Table 2).
Food intake, body weight, and adipose tissue weight
The relationship between food intake, body weight gain, wet weight of liver and fat weight of different parts are represented in Table 3. The data suggests that HFD-induced increase of body weight, liver weight, and fat weight are not related to the amount of food consumed per day rather these are related to the composition of the food. Feeding of ethanolic extract of T. indica leaf (TILE) up to the level of 200 mg/kg everyday for 8 weeks was unable to reduce the HFD-induced body weight, fat weight and liver weight significantly. But TILE administration of 400 mg/kg was successful in reducing body weight, liver weight, mesenteric and peritoneal fat weights significantly (p < 0.05).
Assessment of plasma lipid profile
Feeding of HFD caused in a significant (p < 0.05) increase of TG, TC, LDL-C, VLDL-C, atherogenic index (AI) and coronary risk index (CRI) in comparison to standard laboratory diet (SD) fed group. On the other hand, the plasma level of HDL-C was reduced significantly (p < 0.05) due to the feeding of HFD. Oral administration of 100 mg/kg and 200 mg/kg ethanolic extract of T. indica leaf (TILE) was not able to change these parameters. But feeding of TILE at the higher dose (400 mg/kg) caused the significant reduction of HFD-induced increased levels of TG (32.2%), TC (25.6%), LDL-C (34.1%), VLDL-C (32.6%) levels in plasma and consequently reduced AI (46.3%) and CRI (39.6%). But feeding of TILE even at the highest dose (400 mg/kg) was failed to cause a significant change of HDL-C level (Table 4).
In vivo antioxidant effects of T. indica L. leaf extract
The level of malondialdehyde (MDA) increased 1.83 folds and the level of nitric oxide (NO) increased by 2.66-folds in the blood due to 8 weeks long feeding of a HFD comparison to the rats fed on standard laboratory diet (SD). Additionally, the HFD-mediated increase of MDA and NO levels were accompanied by a significant (p < 0.05) fall of superoxide dismutase (SOD) (56.2%) and catalase (49.7%) activities. Consumption of ethanolic extract of Tamarindus indica leaf (TILE) at low dose (100 mg/kg/day) was not able to reduce the level of MDA and NO markedly. But feeding of TILE at the highest dose (400 mg/kg/day) caused a significant (p < 0.05) reduction of MDA (39.7%) and NO (35.3%) levels in comparison to the HFD-fed rats (Fig. 1). Similarly, feeding of TILE at low level (100 mg/kg/day) was unable to increase the reduced SOD and catalase activities markedly. But consumption of TILE at the highest level (400 mg/kg/day) significantly increased the SOD (1.97 fold) and catalase (1.89 folds) activities in comparison to the HFD-fed rats. Moreover, catalase activity was also increased significantly due to feeding of TILE at intermediated level (200 mg/kg/day) (Fig. 1).

Effects of ethanolic extract of T. indica L. leaf (TILE) on (a) malondialdehyde (MDA), (b) nitric oxide (NO), (c) superoxide dismutase (SOD) activity, and (d) catalse activity in different group of Wistar rats fed with either of standard laboratory diet (SD), or high-fat diet (HFD), or HFD with TILE. TILE was given orally everyday throughout the study period either in low dose (HFD + TILE100), or intermediate dose (HFD + TILE200) or high dose (HFD + TILE400). Data was analyzed by one way ANOVA followed by Newman-Keuls’s post-hoc test and expressed as mean ± SEM (n = 6). Values with different superscript letters within the same row are significantly different at p < 0.05
T. indica leaf extract down-regulated the gene expression of adipogenesis promoting transcription factors
This part of our study was aimed to investigate the impact of T. indica leaf extract on the gene expression of these four important transcription factors like PPARγ, CCAAT/enhancer-binding protein α C/EBPα, LXRα, and SREBP1c which regulates numerous proteins, enzymes and factor that control the conversion of preadipocytes to matured adipocytes and fat metabolism. Our investigation revealed that feeding with a HFD significantly (P < 0.05) up-regulated the gene expression of LXRα (2.41 folds), PPARγ (1.98 folds), C/EBPα (2.11 folds), and SREBP1c (2.13 folds) in comparison to SD fed rats. Consumption of TILE at low (100 mg/kg) dose was unable to change the HFD-induced increased mRNA level. Feeding of TILE at moderate dose (200 mg/kg/day) together with HFD reduced the mRNA level of LXRα and SREBP1c significantly. Consumption of TILE at highest dose (400 mg/kg) every day for 8 weeks significantly (p < 0.05) lowered the transcript level of all the above mentioned factors except C/EBPα (Fig. 2).

Comparison of liver X receptor alpha (LXRα), peroxisome proliferator-activated receptor gamma (PPARγ), CCAAT enhancer binding protein alpha (C/EBPα), and sterol regulatory element binding protein 1c (SREBP1c) and gene expression in epididymal adipose tissue in different group of Wistar rats fed with either of standard laboratory diet (SD), or high-fat diet (HFD), or HFD with TILE. TILE was given orally everyday throughout the study period either in low dose (HFD + TILE100), or intermediate dose (HFD + TILE200) or high dose (HFD + TILE400). Data was analyzed by one way ANOVA followed by Newman-Keuls’s post-hoc test and expressed as mean ± SEM (n = 6). Values with different superscript letters within the same row are significantly different at p < 0.05
Effect of T. indica leaf extract on the gene expression of lipogenic and lipolytic enzymes
To get a deeper understanding the effect of T. indica extract on the transcript levels of lipogenic and lipolytic enzymes were also analyzed in quantitative real-time PCR after extraction of mRNA from different groups of test animals. This part of the study demonstrated that consumption of HFD resulted in augmented transcription of ACC, FAS, DGAT-2, and HMGCR by 2.32 folds, 1.91 folds 2.21 folds and 2.61 folds respectively, in comparison to rats fed on SD. Administration of T. indica leaf extract at low level (100 mg/kg) was unable to change the HFD-induced augmentation of this gene expression of lipogenic enzymes. However, consumption of TILE at highest dose significantly (p < 0.05) lowered of the transcript level of ACC (35.7%), FAS (36.4%), DGAT-2 (33.7%), and HMGCR (52.3%) compared to HFD group (Fig. 3). Moreover, feeding of TILE at intermediate dose (200 mg/kg) significantly (p < 0.05) reduced the gene expression of DGAT-2 and HMGCR (Fig. 3). On the other hand, in HFD consuming rats, the mRNA levels of lipolytic enzymes like hormone-sensitive lipase (HSL), adipose triglyceride lipase (ATGL) and monoacylglycerol lipase (MAGL) were reduced significantly (p < 0.05) in comparison to the rats which received SD. Treatment with TILE even at the highest dose (400 mg/kg) for 8 weeks was unable to change the transcript levels of ATGL and MAGL. But feeding of TILE, both at intermediate dose and high dose significantly (p < 0.05) increased HFD-inhibited level of HSL (Fig. 4).

Comparison of transcript level of acetyl-CoA carboxylase (ACC), fatty acid synthase (FAS), diacylglycerol acyltransferase 2 (DGAT-2), and HMG-CoA reductase (HMGCR) in epididymal adipose tissue in different group of Wistar rats fed with either of standard laboratory diet (SD), or high-fat diet (HFD), or HFD with TILE. TILE was given orally everyday throughout the study period either in low dose (HFD + TILE100), or intermediate dose (HFD + TILE200) or high dose (HFD + TILE400). Data was analyzed by one way ANOVA followed by Newman-Keuls’s post-hoc test and expressed as mean ± SEM (n = 6). Values with different superscript letters within the same row are significantly different at p < 0.05

Comparison of transcript level of adipose triglyceride lipase (ATGL), hormone sensitive lipase (HSL) and monoacyl glycerol lipase (MAGL) in epididymal adipose tissue in different group of Wistar rats fed with either of standard laboratory diet (SD), or high-fat diet (HFD), or HFD with TILE. TILE was given orally everyday throughout the study period either in low dose (HFD + TILE100), or intermediate dose (HFD + TILE200) or high dose (HFD + TILE400). Data was analyzed by one way ANOVA followed by Newman-Keuls’s post-hoc test and expressed as mean ± SEM (n = 6). Values with different superscript letters within the same row are significantly different at p < 0.05
Histological assessment of adipogenesis in hepatic tissue
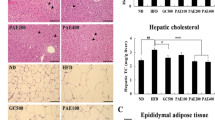
The anti-adipogenic effect of T. indica leaf extract (TILE) on HFD-induced fat accumulation is demonstrated by the photomicrographs of hepatic tissues from different group of rats (Fig. 5a-e). The liver tissue from the rats fed on standard laboratory diet showed very little amount of lipid accumulation which was increased significantly (p < 0.05) due to feeding of HFD. Rats consumed HFD with TILE in lower dose and intermediate dose showed an insignificant reduction of lipid accumulation in comparison to the HFD-fed group. However, feeding of TILE at higher dose together with HFD prevented the lipid deposition significantly (p < 0.05) (Fig. 5f).

Comparison of lipid accumulation in the liver of Wistar rats which was fed with either of standard laboratory diet (SD), or high-fat diet (HFD), or HFD with TILE. TILE was given orally everyday throughout the study period either in low dose (HFD + TILE100), or intermediate dose (HFD + TILE200) or high dose (HFD + TILE400). Photomicrographs (a-e) showing lipid droplets (LD) are representative one of the 3 experiments for a particular group. Additionally, percent lipid accumulation (f) was quantified by ImageJ software (version 1.52a). Data was analyzed by one way ANOVA followed by Newman-Keuls’s post-hoc test and expressed as mean ± SEM (n = 6). Values with different superscript letters within the same row are significantly different at p < 0.05
Discussion
Generally in physiological conditions, the level of pro-oxidants and antioxidants are equilibrated in a balanced condition which might be disturbed by overproduction of reactive oxygen species (ROS) and/or feeble antioxidant defense and may cause oxidative stress [33]. This imbalance can be a major risk factor of metabolic derangement which leading to the conditions like dyslipidemia, obesity, hypertension, insulin resistance and diabetes mellitus [34]. Studies have confirmed a strong connection between high-fat diet consumption and oxidative stress in both humans and in other animals [35]. For example, obesity induced by a high-fat diet has been linked to oxidative stress which causes overproduction of oxidized LDL-cholesterol that plays vital role in the pathogenesis of dyslipidemia, atherosclerosis and other cardiovascular diseases [36]. Therefore, we postulated that any plant extract having antioxidant properties could be a good option for the treatment of ailments related to oxidative stress. Our present investigation revealed that T. indica leaf extract has an antioxidant property which is equivalent to BHT but significantly lower than quercetin and ascorbic acid (Table 2). This was in agreement with the reports where phytochemical analysis confirmed that different parts of T. indica are full of antioxidant compounds like procyanidin B2, epicatechin, tartaric acid, limonene, longifoline, kaempferol, etc. [37]. More specifically, the leaves of T. indica contain some powerful antioxidant compounds including limonene, vitamin C, β-carotene, flavonoids, and different polyphenols [38]. In agreement with in vitro antioxidant and radical scavenging activity T. indica leaf extract was also shown to possess strong antioxidant property in vivo. This study demonstrated that T. indica leaf extract can reduce high-fat diet-induced elevated levels of nitric oxide and malondialdehyde which was accompanied by augmented activities of major antioxidant enzymes like superoxide dismutase and catalase (Fig. 1). This antioxidant power of T. indica leaf might be attributed to a higher level of total polyphenols and flavonoids found in the ethanolic extract (Table 2). Polyphenolic compounds can inhibit the activity of xanthine oxidase enzyme which is involved in the production of superoxide radicals and stimulation of antioxidant enzymes’ activity [39]. Our observations showed similarity with a previous report where an ethanolic extract of T. indica leaf suppressed the generation of ROS and lipid peroxidation while enhanced the antioxidant enzymes’ activity and thus reversed oxidative stress-induced damage in HepG2 cells [40].
The polyphenolic compounds can act as an antioxidant through direct interaction with both radical and non-radical type ROS as well as indirectly by modulating the gene expression of proteins and enzymes having antioxidant activity [41]. Therefore, our next objective was to evaluate the efficiency of polyphenolic compound-rich T. indica leaf extract to ameliorate dyslipidemia, and fat storage in HFD-induced obese Wistar rats through controlling the gene expression of adipocyte differentiation-related transcription factors, lipogenic and lipolytic enzymes. In the 8 weeks long feeding period, T. indica leaf extract was able to reduce HFD-induced increased body weight, liver weight, mesenteric and peritoneal adipose tissue weight (Table 3) with the simultaneous lowering of TG, TC, LDL-C and VLDL-C (Table 4). That increase of lipid in the blood triggered the increase of fat accumulation in the hepatic tissue as was revealed in the photomicrographs taken after hematoxylin and eosin staining (Fig. 5). Our findings were in agreement with another report where the extract of T. indica seeds was shown to have potent plasma lipid and glucose-lowering activities in streptozotocin-induced diabetic rats [10]. However, our observations were slightly different from another report where T. indica aqueous pulp extract considerably reduced serum cholesterol, body weight, and LDL-C but increased HDL-C and triglycerides in normal chow diet-fed healthy rats [42].
Obesity is a lipid metabolism disorder that involves the participation of diverse genetic and dietary factors. Over the last few years, considerable progress has been accomplished in the conceptualization of molecular mechanism of adipogenesis. Proper mapping of transcriptional regulation and identification of pro- and anti-adipogenic endogenous factors is therefore vital in decoding the molecular mechanism of any therapeutic intervention of obesity and associated health issues [43]. Among those factors, PPARγ and C/EBPα are the transcription factors that have very important functions in the conversion of preadipocytes to matured adipocytes [44]. These two factors up-regulate the expression of downstream adipocyte-specific factors, enzymes, and proteins. For this reason, these two are regarded as the master regulators of adipogenesis [45]. The gene expression of PPARγ is controlled by up-stream transcription factor LXRα which also controls the expression of another vital transcription factor SREBP1c [46]. Studies have reported that SREBP1c by interacting with specific sterol regulatory element sequences of DNA enhances the expression of enzymes like ACC, FAS and HMGCR and thus enhances lipogenesis [47]. Therefore, we hypothesized that the anti-adipogenic effect of antioxidant-rich T. indica leaf extract might have been mediated through controlling the gene expression of the above-mentioned transcription factors and enzymes. By considering the objective of delving deeper into the molecular mechanism, we explored the gene expression of those pre-adipocyte differentiation-related transcription factors and some important lipogenic and lipolytic enzymes which are directly or indirectly controlled by those factors. This part of our investigation revealed that administration of T. indica leaf extract reduced the gene expression of LXRα, and its down-stream PPARγ and SREBP1c significantly (P < 0.05), all of which were augmented due to feeding of HFD (Fig. 2). Similar pattern of changes have been reported in previous studies where researchers found that crude plant extracts reduced diet-induced augmented gene expression of PPARγ in rats which resulted in the reduced plasma level of TG and reduced body weight [48, 49].
Among the lipogenic enzymes, ACC and FAS are directly modulated by SREBP1c and play the most vital roles in fatty acid biosynthesis [50]. On the other hand, DGAT catalyzes the biosynthesis of triglyceride [51]. Additionally, the expression of HMGCR enzyme that catalyzes the most important step in cholesterol biosynthesis was also explored. Our investigation revealed that feeding to T. indica leaf extract significantly suppressed the HFD-induced augmented mRNA levels of ACC, FAS, DGAT-2, and HMGCR (Fig. 3). Thus our findings were in agreement with another report where researchers using the Syrian hamster as model animal demonstrated that anti-obesity effect of T. indica fruit pulp might be attributed to the suppression of HMG-CoA reductase gene expression [17]. On the other hand, ATGL, HSL, and MAGL are the most important enzymes involved in TG catabolism in adipose and other tissues [52, 53]. Our study revealed that the high-fat diet-mediated suppression of HSL was normalized significantly (P < 0.05) due to the feeding of 400 mg/kg T. indica leaf extract daily for 8 weeks (Fig. 4).
Conclusion
This study demonstrated that oral administration of antioxidant-rich Tamarindus indica L. leaf extract (TILE) for 8 weeks significantly reduced the HFD-induced oxidative stress by increasing the activities of antioxidant enzymes. TILE also down-regulated the mRNA levels of adipogenesis controlling transcription factors and lipogenic enzymes with a parallel up-regulation of the transcript level of lipolytic enzyme, HSL. Consistently, TILE reduced the amount of TG, TC, LDL-C and VLDL-C in the serum with an orchestrated lowering of adipose tissue weight, liver weight and body weight.
Availability of data and materials
The corresponding author is entitled to provide the data upon reasonable request.
Abbreviations
- ACC:
-
actyl-CoA carboxylase
- ATGL:
-
adipose triglyceride lipase
- C/EBPα:
-
CCAAT-enhancer-binding protein α
- DGAT:
-
diacylglycerol acyltransferase
- FAS:
-
fatty acid synthase
- FRAP:
-
ferric reducing antioxidant power
- GPx:
-
glutathione peroxidase
- HDL-C:
-
high-density lipoprotein cholesterol
- HFD:
-
high-fat diet
- HMGCR:
-
3-hydroxy-3-methyl-glutaryl-coenzyme A reductase
- HSL:
-
hormone-sensitive lipase
- LDL-C:
-
low-density lipoprotein cholesterol
- LXRα:
-
liver X receptor-α
- MAGL:
-
monoacylglycerol lipase
- PPARγ:
-
peroxisome proliferator-activated receptor-γ
- RT-PCR:
-
reverse-transcription polymerase chain reaction
- SOD:
-
superoxide dismutase
- SREBP:
-
sterol regulatory element-binding protein
- TC:
-
total cholesterol
- TG:
-
triacylglycerol or triglyceride
- TILE:
-
ethanolic extract of Tamarindus indica L. leaf extract
- VLDL-C:
-
very-low-density lipoprotein cholesterol
References
Khan TJ, Ahmed YM, Zamzami MA, Mohamed SA, Khan I, Baothman OA, et al. Effect of atorvastatin on the gut microbiota of high fat diet-induced hypercholesterolemic rats. Sci Rep. 2018;8(1):662 https://doi.org/10.1038/s41598-017-19013-2.
Petrovska BB. Historical review of medicinal plants' usage. Pharmacogn Rev. 2012;6(11):1–5 https://doi.org/10.4103/0973-7847.95849.
Devarajan A, Mohanmarugaraja M. A comprehensive review on Rasam: a south Indian traditional functional food. Pharmacogn Rev. 2017;11(22):73 https://doi.org/10.4103/phrev.phrev_13_17.
Bhadoriya SS, Ganeshpurkar A, Narwaria J, Rai G, Jain AP. Tamarindus indica: extent of explored potential. Pharmacogn Rev. 2011;5(9):73 https://doi.org/10.4103/0973-7847.79102.
Sudjaroen Y, Haubner R, Würtele G, Hull W, Erben G, Spiegelhalder B, et al. Isolation and structure elucidation of phenolic antioxidants from tamarind (Tamarindus indica L.) seeds and pericarp. Food Chem Toxicol. 2005;43(11):1673–82 https://doi.org/10.1016/j.fct.2005.05.013.
Martinello F, Soares S, Franco JJ, Santos ACd, Sugohara A, Garcia SB, et al. Hypolipemic and antioxidant activities from Tamarindus indica L. pulp fruit extract in hypercholesterolemic hamsters. Food Chem Toxicol 2006;44(6):810–818. https://doi.org/10.1016/j.fct.2005.10.011.
Luengthanaphol S, Mongkholkhajornsilp D, Douglas S, Douglas PL. Pengsopa L-i, Pongamphai S. extraction of antioxidants from sweet Thai tamarind seed coat––preliminary experiments. J Food Eng. 2004;63(3):247–52 https://doi.org/10.1016/j.jfoodeng.2003.07.006.
Ushanandini S, Nagaraju S, Harish Kumar K, Vedavathi M, Machiah D, Kemparaju K, et al. The anti-snake venom properties of Tamarindus indica (leguminosae) seed extract. Phytother Res. 2006;20(10):851–8 https://doi.org/10.1002/ptr.1951.
Iftekhar A, Rayhan I, Quadir MA, Akhteruzzaman S, Hasnat A. Effect of Tamarindus indica fruits on blood pressure and lipid-profile in human model: an in vivo approach. Pak J Pharm Sci. 2006;19(2):125–9.
Maiti R, Jana D, Das U, Ghosh D. Antidiabetic effect of aqueous extract of seed of Tamarindus indica in streptozotocin-induced diabetic rats. J Ethnopharmacol. 2004;92(1):85–91 https://doi.org/10.1016/j.jep.2004.02.002.
Seo JB, Moon HM, Kim WS, Lee YS, Jeong HW, Yoo EJ, et al. Activated liver X receptors stimulate adipocyte differentiation through induction of peroxisome proliferator-activated receptor γ expression. Mol Cell Biol. 2004;24(8):3430–44 https://doi.org/10.1128/mcb.24.8.3430-3444.2004.
Shao X, Wang M, Wei X, Deng S, Fu N, Peng Q, et al. Peroxisome proliferator-activated receptor-γ: master regulator of adipogenesis and obesity. Curr Stem Cell Res Ther. 2016;11(3):282–9 https://doi.org/10.2174/1574888x10666150528144905.
Rosen ED, Hsu C-H, Wang X, Sakai S, Freeman MW, Gonzalez FJ, et al. C/EBPα induces adipogenesis through PPARγ: a unified pathway. Genes Dev. 2002;16(1):22–6 https://doi.org/10.1101/gad.948702.
Tang X, Li J, Xiang W, Cui Y, Xie B, Wang X, et al. Metformin increases hepatic leptin receptor and decreases steatosis in mice. J Endocrinol. 2016;230(2):227–37 https://doi.org/10.1530/joe-16-0142.
Xu X, So J-S, Park J-G, Lee A-H, editors. Transcriptional control of hepatic lipid metabolism by SREBP and ChREBP. Seminars in liver disease; 2013: Thieme Medical Publishers. https://doi.org/10.1055/s-0033-1358523.
Ferre P, Foufelle F. SREBP-1c transcription factor and lipid homeostasis: clinical perspective. Horm Res Paediatr. 2007;68(2):72–82 https://doi.org/10.1159/000100426.
Lim CY, Junit SM, Abdulla MA, Aziz AA. In vivo biochemical and gene expression analyses of the antioxidant activities and hypocholesterolaemic properties of Tamarindus indica fruit pulp extract. PLoS One. 2013;8(7):e70058 https://doi.org/10.1371/journal.pone.0070058.
Noreen H, Semmar N, Farman M, McCullagh JS. Measurement of total phenolic content and antioxidant activity of aerial parts of medicinal plant Coronopus didymus. Asian Pac J Trop Med. 2017;10(8):792–801 https://doi.org/10.1016/j.apjtm.2017.07.024.
Baba SA, Malik SA. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J Taibah Univ Sci. 2015;9(4):449–54 https://doi.org/10.1016/j.jtusci.2014.11.001.
Pulido R, Bravo L, Saura-Calixto F. Antioxidant activity of dietary polyphenols as determined by a modified ferric reducing/antioxidant power assay. J Agric Food Chem. 2000;48(8):3396–402 https://doi.org/10.1021/jf9913458.
Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice-Evans C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol Med. 1999;26(9–10):1231–7 https://doi.org/10.1016/s0891-5849(98)00315-3.
Braca A, & Tommasi N, Di BL, Pizza C, & Politi M, Morelli, I. Antioxidant principles from Bauhinia terapotensis. J Nat Prod. 2001; 64. 892–5.
Choi H-K, Won E-K, Jang YP, Choung S-Y. Antiobesity effect of Codonopsis lanceolata in high-calorie/high-fat-diet-induced obese rats. Evid Based Complement Alternat Med. 2013;2013 https://doi.org/10.1155/2013/210297.
Akase T, Shimada T, Terabayashi S, Ikeya Y, Sanada H, Aburada M. Antiobesity effects of Kaempferia parviflora in spontaneously obese type II diabetic mice. J Nat Med. 2011;65(1):73–80 https://doi.org/10.1007/s11418-010-0461-2.
Kim S-J, Jung JY, Kim HW, Park T. Anti-obesity effects of Juniperus chinensis extract are associated with increased AMP-activated protein kinase expression and phosphorylation in the visceral adipose tissue of rats. Biol Pharm Bull. 2008;31(7):1415–21 https://doi.org/10.1248/bpb.31.1415.
Martin SS, Blaha MJ, Elshazly MB, Toth PP, Kwiterovich PO, Blumenthal RS, et al. Comparison of a novel method vs the Friedewald equation for estimating low-density lipoprotein cholesterol levels from the standard lipid profile. JAMA. 2013;310(19):2061–8 https://doi.org/10.1001/jama.2013.280532.
Kazemi T, Hajihosseini M, Moossavi M, Hemmati M, Ziaee M. Cardiovascular risk factors and Atherogenic indices in an Iranian population: Birjand east of Iran. Clin Med Insights Cardiol. 2018;12:1179546818759286 https://doi.org/10.1177/1179546818759286.
Zeb A, Ullah F. A simple spectrophotometric method for the determination of thiobarbituric acid reactive substances in fried fast foods. J Anal Methods Chem. 2016;2016 https://doi.org/10.1155/2016/9412767.
Ghasemi A, Hedayati M, Biabani H. Protein precipitation methods evaluated for determination of serum nitric oxide end products by the Griess assay. JMSR. 2007;2(15):29–32.
Kakkar P, Das B, Viswanathan P. A modified spectrophotometric assay of superoxide dismutase. 1984. Indian J Biochem Biophys. 1984;21(2):130–2.
Hadwan MH. Simple spectrophotometric assay for measuring catalase activity in biological tissues. BMC Biochem. 2018;19(1):7 https://doi.org/10.1186/s12858-018-0097-5.
Noratto GD, Lage NN, Chew BP, Mertens-Talcott SU, Talcott ST, Pedrosa ML. Non-anthocyanin phenolics in cherry (Prunus avium L.) modulate IL-6, liver lipids and expression of PPARδ and LXRs in obese diabetic (db/db) mice. Food Chem. 2018;266:405–14 https://doi.org/10.1016/j.foodchem.2018.06.020.
Birben E, Sahiner UM, Sackesen C, Erzurum S, Kalayci O. Oxidative stress and antioxidant defense. World Allergy Organ J. 2012;5(1):9 https://doi.org/10.1097/wox.0b013e3182439613.
Tangvarasittichai S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J Diabetes. 2015;6(3):456 https://doi.org/10.4239/wjd.v6.i3.456.
Duan Y, Zeng L, Zheng C, Song B, Li F, Kong X, et al. Inflammatory links between high fat diets and diseases. Front Immunol. 2018;13(9):2649 https://doi.org/10.3389/fimmu.2018.02649.
Mashima R, Witting PK, Stocker R. Oxidants and antioxidants in atherosclerosis. Curr Opin Lipidol. 2001;12:411–8 https://doi.org/10.1097/00041433-200108000-00007.
Bhadoriya S, Mishra V, Raut S, Ganeshpurkar A, JAIN S. Anti-inflammatory and antinociceptive activities of a hydroethanolic extract of Tamarindus indica leaves. Sci Pharm. 2012;80(3):685–700 https://doi.org/10.3797/scipharm.1110-09.
Escalona-Arranz JC, Péres-Roses R, Urdaneta-Laffita I, Camacho-Pozo MI, Rodríguez-Amado J, Licea-Jiménez I. Antimicrobial activity of extracts from Tamarindus indica L. leaves. Pharmacogn Mag. 2010;6(23):242–7 https://doi.org/10.4103/0973-1296.66944.
Rodrigo R, Miranda A, Vergara L. Modulation of endogenous antioxidant system by wine polyphenols in human disease. Clin Chim Acta. 2011;412(5–6):410–24 https://doi.org/10.1016/j.cca.2010.11.034.
Razali N, Aziz AA, Lim CY, Junit SM. Investigation into the effects of antioxidant-rich extract of Tamarindus indica leaf on antioxidant enzyme activities, oxidative stress and gene expression profiles in HepG2 cells. PeerJ. 2015;3:e1292 https://doi.org/10.7717/peerj.1292.
Stoclet J-C, Chataigneau T, Ndiaye M, Oak M-H, El Bedoui J, Chataigneau M, et al. Vascular protection by dietary polyphenols. Eur J Pharmacol. 2004;500(1–3):299–313 https://doi.org/10.1016/j.ejphar.2004.07.034.
Jindal V, Dhingra D, Sharma S, Parle M, Harna RK. Hypolipidemic and weight reducing activity of the ethanolic extract of Tamarindus indica fruit pulp in cafeteria diet-and sulpiride-induced obese rats. J Pharmacol Pharmacother. 2011;2(2):80–4 https://doi.org/10.4103/0976-500x.81896.
Lefterova MI, Lazar MA. New developments in adipogenesis. Trends Endocrinol Metab. 2009;20(3):107–14 https://doi.org/10.1016/j.tem.2008.11.005.
Lefterova MI, Zhang Y, Steger DJ, Schupp M, Schug J, Cristancho A, et al. PPARγ and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008;22(21):2941–52 https://doi.org/10.1101/gad.1709008.
Madsen MS, Siersbæk R, Boergesen M, Nielsen R, Mandrup S. Peroxisome proliferator-activated receptor γ and C/EBPα synergistically activate key metabolic adipocyte genes by assisted loading. Mol Cell Biol. 2014;34(6):939–54 https://doi.org/10.1128/mcb.01344-13.
Seo JB, Moon HM, Noh MJ, Lee YS, Jeong HW, Yoo EJ, et al. Adipocyte determination-and differentiation-dependent factor 1/sterol regulatory element-binding protein 1c regulates mouse adiponectin expression. J Biol Chem. 2004;279(21):22108–17 https://doi.org/10.1074/jbc.m400238200.
Kidani Y, Bensinger SJ. Liver X receptor and peroxisome proliferator-activated receptor as integrators of lipid homeostasis and immunity. Immunol Rev. 2012;249(1):72–83 https://doi.org/10.1111/j.1600-065x.2012.01153.x.
Lestari SR, Djati MS, Rudijanto A, Fatchiyah F. PPARγ expression by rambutan peel extract in obesity rat model-induced high-calorie diet. Asian Pac J Trop Biomed. 2015;5(10):852–7 https://doi.org/10.1016/j.apjtb.2015.01.030.
Kim JH, Kim OK, Yoon HG, et al. Anti-obesity effect of extract from fermented Curcuma longa L. through regulation of adipogenesis and lipolysis pathway in high-fat diet-induced obese rats. Food Nutr Res. 2016;60(1):30428 https://doi.org/10.3402/fnr.v60.30428.
Menendez JA, Vazquez-Martin A, Ortega FJ, Fernandez-Real JM. Fatty acid synthase: association with insulin resistance, type 2 diabetes, and cancer. Clin Chem. 2009;55(3):425–38 https://doi.org/10.1373/clinchem.2008.115352.
Yen C-LE, Stone SJ, Koliwad S, Harris C, Farese RV. Thematic review series: glycerolipids. DGAT enzymes and triacylglycerol biosynthesis. J Lipid Res. 2008;49(11):2283–301 https://doi.org/10.1194/jlr.r800018-jlr200.
Schweiger M, Schreiber R, Haemmerle G, Lass A, Fledelius C, Jacobsen P, et al. Adipose triglyceride lipase and hormone-sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J Biol Chem. 2006;281(52):40236–41 https://doi.org/10.1074/jbc.m608048200.
Morak M, Schmidinger H, Riesenhuber G, Rechberger GN, Kollroser M, Haemmerle G, et al. Adipose triglyceride lipase (ATGL) and hormone-sensitive lipase (HSL) deficiencies affect expression of lipolytic activities in mouse adipose tissues. Mol Cell Proteomics. 2012;11(12):1777–89 https://doi.org/10.1074/mcp.m111.015743.
Acknowledgments
The authors are grateful to the chairman, Department of Pharmaceutical Sciences, North South University Dhaka-1229, Bangladesh and to the director, North South University Genome Research Institute, Dhaka-1229, for allowing us to use the laboratory instruments.
Funding
This work was funded by North South University Research Grant Cycle 2018–2019 awarded to Dr. Ferdous Khan (CTRG code: NSU-RP-18-013). The authors are also grateful to the Department of Pharmaceutical Sciences, North South University and North South University Genome Research Institute for their helpful gestures to conduct the research.
Author information
Authors and Affiliations
Contributions
SAK and MIB performed biochemical assays; NS and MHS prepared the extract and performed antioxidant assay; AR and MMH extracted RNA and did the quantitative real-time PCR; MAA and FK designed the study and prepared the manuscript. All authors checked and approved the final version of this manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The study protocol was approved by Ethical Review Committee of the Department of Pharmaceutical Sciences, North South University, Bangladesh (Ref No.: ACE-0017-2018).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kuddus, S.A., Bhuiyan, M.I., Subhan, N. et al. Antioxidant-rich Tamarindus indica L. leaf extract reduced high-fat diet-induced obesity in rat through modulation of gene expression. Clin Phytosci 6, 68 (2020). https://doi.org/10.1186/s40816-020-00213-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40816-020-00213-9