
Abstract
Marine bacterioplankton underpin the health and function of coral reefs and respond in a rapid and sensitive manner to environmental changes that affect reef ecosystem stability. Numerous meta-omics surveys over recent years have documented persistent associations of opportunistic seawater microbial taxa, and their associated functions, with metrics of environmental stress and poor reef health (e.g. elevated temperature, nutrient loads and macroalgae cover). Through positive feedback mechanisms, disturbance-triggered heterotrophic activity of seawater microbes is hypothesised to drive keystone benthic organisms towards the limit of their resilience and translate into shifts in biogeochemical cycles which influence marine food webs, ultimately affecting entire reef ecosystems. However, despite nearly two decades of work in this space, a major limitation to using seawater microbes in reef monitoring is a lack of a unified and focused approach that would move beyond the indicator discovery phase and towards the development of rapid microbial indicator assays for (near) real-time reef management and decision-making. By reviewing the current state of knowledge, we provide a comprehensive framework (defined as five phases of research and innovation) to catalyse a shift from fundamental to applied research, allowing us to move from descriptive to predictive reef monitoring, and from reactive to proactive reef management.
Similar content being viewed by others
Introduction
Coral reefs are some of the most biodiverse and productive aquatic environments on the planet, providing shelter, nutrition, and habitat for many marine species, and offering valuable ecosystem services to humans, including protection of coastal areas, tourism, and fisheries [1, 2]. Despite their ecological significance and economic value, coral reefs have suffered major declines in recent decades due to the synergistic effects of local chronic impacts and global climate change [3, 4] with recent estimates indicating that half the world’s coral cover has been lost since the 1950s [5]. To preserve coral reefs, an improved understanding is needed of the mechanisms involved in coral resilience to local and global environmental stressors.
Marine microorganisms account for ~ 65–90% of the marine biomass [6, 7] and therefore constitute the life support system of the biosphere, being central to planetary marine food webs and biogeochemical cycles, and responsible for approximately 50% of the world’s primary production [7,8,9,10,11,12,13,14]. Marine plankton also play a vital role in the stability and function of coral reefs by providing crucial ecosystem services. Heterotrophic microbes in reef seawater rapidly capture and recycle nutrients from the water column, for example, dissolving coral derived mucus before it sinks to the sediment [15]. By rapidly taking up nutrients from the water column, seawater microbes have a critical role in making these nutrients available to higher trophic levels [15,16,17,18]. This efficient recycling of nutrients ultimately allows corals to thrive in oligotrophic and nutrient-deplete environments, often referred to as ‘marine deserts’ [18].
Host-associated microbes also provide various functions to their metazoan hosts, including nutrition, removal of waste products (e.g. ammonia), protection from invading pathogens, and stimulation of developmental processes and morphogenesis [19,20,21,22,23,24]. However, environmental stressors such as eutrophication and elevated temperatures may shift host-associated microbial communities from mutualistic to pathogenic states once critical thresholds are reached [25,26,27]. The emergence of copiotrophic and potentially pathogenic microbes (e.g. Flavobacteriaceae, Cryomorphaceae, Rhodobacteraceae, Rhodospirillaceae, Vibrio) along with their associated functions (e.g. virulence factors and toxin production) has been associated with increases in coral diseases leading to tissue necrosis, and ultimately partial or whole colony mortality [26, 28, 29].
This sensitivity of reef microbes to environmental perturbations potentially allows microbes to be used as indicators of environmental change in the surrounding reef [30, 31]. Importantly, reef microorganisms may represent early warning indicators of environmental disturbances since microbial communities change in their composition and function before the development of visual signs of stress, such as coral disease, bleaching, and tissue necrosis [25, 30,31,32,33,34,35,36,37,38,39]. These traditional visual signs of reef disturbance often become evident only after prolonged periods and potentially once ecosystem tipping points are reached [40]. Current monitoring efforts are therefore often reactive, reporting the outcomes of impact with limited potential to mitigate future reef decline. Incorporating microbial processes within reef monitoring frameworks could represent a powerful way to observe early signs of stress, providing more time to implement reef management strategies and mitigate the impacts of environmental disturbances on reefs [30, 31].
A framework to implement microbial observations within reef monitoring programs was recently proposed [36]. Using indicator value analysis and machine learning approaches, seawater microbial communities (inferred from 16S rRNA gene amplicon sequencing data) were documented to provide accurate predictions of water temperature and eutrophication states of reefs. In contrast, macroalgae, coral and sponge microbiomes were predominantly structured by the host organism and less influenced by the environment [36]. Exposure of coral and sponge species to non-lethal stressors (temperature, acidification and salinity) in controlled experimental systems has similarly demonstrated that host factors strongly influence host-associated microbiomes [41,42,43,44,45], possibly limiting their use as early indicators of stress in reef monitoring. These trends were recently also documented at scale, using 16S rRNA gene amplicon sequencing data from the Tara Pacific Expedition [46]. Only 4–11% of variance in the coral microbiomes surveyed (Millepora, Porites and Pocillopora) was explained by physicochemical properties of the seawater, compared with ~ 30% variance in planktonic microbial communities explained by water chemistry [46]. Considering that seawater microbes provide accurate diagnostics of temperature and eutrophication states in the reef environment [36] and that seawater can be easily collected alongside in situ reef health surveys in a cost-effective and non-destructive manner, we assert there is realistic scope to incorporate microbial observations of seawater microbes alongside ongoing in situ reef health surveys (Fig. 1).

Overview of the diagnostic value of various coral reef microbiomes. The diagnostic value (indicated as stars) is based on the sum of advantages (+) and disadvantages (−) for key characteristics of optimal microbial indicators: (1) ease of sampling, (2) sensitivity towards environmental fluctuations, (3) uniformity of community assembly, (4) our ability to link microbiome shifts to host health. Based on these criteria, seawater microbial communities collectively have the highest diagnostic potential to be used as microbial indicators of reef health, followed by sediment-associated and host-associated microbial communities, respectively. Free-living microbial communities (seawater and sediment) can be easily collected, without interfering with ecosystem processes and/or the health of reef organisms, consistent with desirable characteristics for environmental monitoring programs. In contrast, the collection of host-associated microbiomes is labour intensive and potentially poses a certain risk for host health when collecting tissue, although collections of the host-biofilm are non-invasive for the host. Seawater also revealed the highest sensitivity to changes in the surrounding environment (e.g., temperature and eutrophication) due to uniform community assembly patterns of the seawater microbiome across replicates, while sediments were primarily influenced by site-specific patterns (e.g. grain size) and host-associated microbiomes predominantly showed a host-genotype modulation. While the diagnostic value is highest for most criteria in the seawater microbiome, it is challenging to link disturbance-induced shifts in marine bacterioplankton to host health. Given the importance of host-associated microbes to the health of reef holobionts, the establishment of microbial baselines for host-associated microbiomes and the search for host health microbial indicators are still warranted
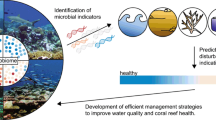
Apart from early detection of environmental changes (Fig. 2), seawater microbes are also important to predict reef functioning as environmental perturbations can destabilise reef bacterioplankton and alter their ecosystem services (Fig. 3), resulting in adverse implications on future reef dynamics via cascading effects and feedback loops [39, 47, 48]. For example, cumulative effects of nutrient eutrophication and elevated temperature can trigger heterotrophic microbial activity in seawater, resulting in harmful algae blooms and hypoxia at reef scales causing rapid coral mortality [48,49,50]. Heterotrophic seawater microbes were also proposed to be central to large-scale reef declines caused by chronic stressors such as elevated nutrients and overfishing via the DDAM (disease, dissolved organic carbon (DOC), algae and microbes) model [47, 51, 52]. The DDAM positive feedback loop begins with eutrophication and overfishing facilitating growth of fleshy macroalgae, which confers a competitive advantage to other macroalgae over coralline algae and calcifying corals by preventing settlement of coral larvae [51]. At the same time, ocean warming caused by climate change stimulates the release of dissolved organic carbon (DOC) by fleshy macroalgae, which results in the proliferation of copiotrophic and potentially pathogenic bacterial communities in seawater, a process referred to as microbialisation. Increased abundance and activity of opportunistic and potentially pathogenic microbes in the water column further fuels the DDAM positive feedback loop by causing additional coral decline through increased coral disease prevalence, which ultimately maintains algal competitive dominance [51]. This concept of microbialisation links changes in seawater reef microbes to reef health decline and is therefore important from a predictive monitoring perspective (Fig. 3).

Potential of reef seawater microbes to inform on reef health status. Successful reef management interventions need to rely on acute and early identification of changes in the reef, before ecosystem ‘tipping points’ are reached (A). However, most reef monitoring programs are based on visual signs to assess ecosystem stress (e.g., coral disease, bleaching and community-level shifts), which become evident only after prolonged environmental disturbances (B). Due to their short generation times, seawater microbes respond rapidly to environmental changes, and it has therefore been well established that marine bacterioplankton allows accurate and early diagnostics of environmental fluctuations in the reef (C, middle). However, the predictive potential of the seawater microbiome has been largely unexplored and it remains unclear how environmental changes will alter microbial functioning of reef bacterioplankton, and how this may translate to reef ecosystem functioning via cascading effects and feedback loops (C, middle). Figure 2C was adjusted from Vanwonterghem and Webster [18] with permission from authors
Currently most of what we know about the potential for seawater microbes to predict reef health has been inferred from microbial taxonomy rather than microbial function [18]. Amplicon sequencing of the 16S rRNA gene (the universal taxonomic marker gene in bacteria and archaea) has identified opportunistic and potentially pathogenic microbes persistently associated with degraded reefs across independent meta-omics studies [26, 39, 52,53,54,55]. However, it is still unclear how seawater microbes can be applied as indicators of coral reef ecosystem health as (1) functional characterisation (i.e. survey of functional potential via metagenomic sequencing) of reef bacterioplankton is still largely lacking (but see Tara Pacific Expedition, [56,57,58, 65]), and because (2) frameworks still need to resolve the ecologically important functions that seawater microorganisms provide to the reef ecosystem. Meta-omics approaches (e.g., metagenomics, metatranscriptomics, metaproteomics and metabolomics) that survey specific functional genes extend beyond taxonomy and would allow scalable investigation of adaptive (i.e. community turnover) and acclimatory (e.g. physiological and gene expression changes) responses of microbes to their environment [59]. For example, undertaking meta-omic surveys to document microbial functional potential as part of reef microbial observations can elucidate how environmental change affects ecosystem services that seawater microbes provide to coral reefs, and to predict how this may translate to future reef dynamics (Fig. 3). However, when applying meta-omics for ecosystem monitoring, there are numerous strengths and weaknesses associated with the various technologies that need to be considered. These methodological considerations have been extensively reviewed previously [59,60,61,62,63,64] and are therefore not covered in detail here.
In this review, we demonstrate that there is realistic scope to extend reef monitoring efforts by including microbial meta-omic surveys that capture the metabolic and functional potential of free-living microbes in seawater. We discuss the current state of using seawater microbial indicators in reef monitoring programs in the context of a five-step framework (Fig. 4). We hope this review will accelerate the shift from fundamental towards applied research to develop rapid and cost-effective microbial-based assays for assessment of reef health, which would be invaluable in predictive reef monitoring and proactive management.
Seawater microbes are essential to predict ocean and reef health
Enormous amounts of molecular and environmental data have been collected in recent years on oceanic microbes, both through various global sampling efforts that collected snapshots of marine microbes in time and space [8, 10,11,12, 14, 56,57,58, 65,66,67] as well as various long-term microbial observatory stations [13]. These large microbial oceanography initiatives aim to predict how environmental change alters the distribution patterns as well as taxonomic and functional diversity of ocean plankton at global scales [7, 9, 14, 68, 69]. To successfully integrate these large and complex datasets, novel computational approaches such as multi-omics data integration [70], ecological niche modelling [71, 72], network analysis [72, 73], multivariate statistics and supervised learning needed to be applied, and these modelling and data integration efforts have already enabled marine scientists to transition from hindcasting to forecasting. For example, tropical marine biogeographical provinces are predicted to expand towards the poles due to climate change (at the expense of temperate and polar zones), followed by a compositional shift in marine plankton which is projected to decrease carbon export fluxes and affect nitrogen cycling [69]. Furthermore, marine viruses were identified as the best predictors of global ocean carbon flux in comparison with archaea, bacteria and eukaryotes [73]. Further integration of seawater microbial meta-omics data into models of Earth system functioning will be crucial to improve such models, as marine bacterioplankton are directly involved in the processes being modelled such as biogeochemical cycling, primary production, and carbon efflux under climate change scenarios [74].
Statistical learning models have also been applied to coral reefs to identify microbial indicators that inform ecosystem health, though at comparatively smaller scales [36, 39]. For example, random forest machine learning identified that seawater surface temperature in the Great Barrier Reef can be accurately predicted from reef bacterioplankton community structure [36], and a linear discriminant analysis (LDA) model was developed that accurately predicts reef categories (e.g. inshore, mid-shelf or offshore) in the GBR based on seawater microbial community profiles [39]. Importantly, vast amounts of meta-omic and multi-omic data streams have recently been collected on free-living and host-associated reef microbes in the Pacific Ocean (e.g. the Tara Pacific Expedition), which will allow further development of models to incorporate seawater microbes in predicting how climate change and human impact may affect reef functioning and health [56,57,58, 65, 75]. The Tara Pacific Expedition (2016–2018) has sampled seawater and coral for multi-omics sequencing in 32 island systems throughout the Pacific along with extensive environmental metadata, hence establishing spatial baselines of reef microbes in the Pacific Ocean [46, 56,57,58, 65, 75].
These large-scale meta- and multi-omics datasets will, for the first time, provide the necessary basis to assess how functions of reef seawater microbes (e.g. photosynthesis, nitrification, ammonia oxidation, sulfate reduction, methanogenesis, virulence etc.) shift with the environment at global scales. Such datasets will be crucial to extend beyond localised studies that have already identified seawater microbes indicating poor reef health, by establishing a robust baseline of microbial indicators that are shared across wide spatial and temporal scales. A recent literature review provided such a baseline, summarising how reef habitat degradation across regions in the Pacific Ocean and the Caribbean may alter microbe–DOM interactions, and the potential implications of shifts in microbial functioning contributing to further reef declines [76]. Further, analysis of large-scale meta-omics data of the seawater microbiome surrounding corals could provide insight about how local settings affect reef bacterioplankton, and how this may structure and affect dynamics of coral microbiomes [65, 75], improving our understanding of the role of seawater microbes in reef resilience and acclimatisation (Fig. 3). Considering the implication of free-living seawater microbes in feedback loops and cascading effects, developing regulatory guidelines is needed to protect seawater microbial functions that are ecologically relevant to the reef, which would be invaluable in reef monitoring if early detection of how specific microbial functions are disrupted could be used to predict and avoid additional coral mortality and reef declines [77].

Reef microbial observation should extend beyond taxonomy and towards function, to move from descriptive to predictive reef monitoring. 16S rRNA amplicon-sequence data has clearly shown that opportunistic and potentially pathogenic microbes robustly correlate to degraded reefs characterised by poor water quality, increased macroalgae cover and coral disease/bleaching. However, amplicon-sequence data has a limited resolution to go beyond description of past or present changes in the reef, as the consequences of the enrichment of particular microbial indicator taxa on reef health often cannot be inferred from microbial taxonomy alone (left, shown in red). Microbial meta-omics data would allow prediction of how environmental changes will affect the services microbes provide to coral reefs (e.g., primary productivity, nutrient/biogeochemical cycling, and exposure to pathogens), and how the altered microbial activity may translate to reef ecosystem dynamics (right). This predictive monitoring is needed for successful reef management and decision making (bottom)
Experimental validation of microbes that predict poor reef health
Large meta- and multi-omic data streams such as Tara Pacific will be vital for identifying the indicator taxa and genes that predict poor reef health, however, this large data collection exercise is just one step in incorporating microbial related processes in reef monitoring frameworks (Fig. 4, phase 1). In many instances the microbial features selected by the models may be unrelated to degraded ecosystem health despite high correlations identified by the model [78]. As an example, an environmental overlap may exist between microbial indicators that correlate to coral bleaching (which occurs under accumulated thermal stress) and thermophilic bacteria which also proliferate at elevated temperatures, but do not impact reef health. Further, anaerobic bacteria and genes (which could indicate hypoxia in the reef) can still be present in the seawater days after a hypoxia-induced coral mortality event, and despite concentrations of dissolved oxygen having reverted to normal values [79]. Various additional factors may contribute to the noisy signal of microbial taxa-environment associations, including biotic interactions, limits to spatial dispersal and neutral demographic drift [80]. As there is a strong likelihood that only some microbial predictors of degraded reefs identified by the model will have a causal association with metrics of poor reef health, confirmatory experiments are needed to validate microbiome-environment associations [78] allowing interpretation of the suitability of the models and usefulness of the microbial indicator(s) (Fig. 4, phase 2), however this remains a distant goal. For example, while pathogenic microbes are hypothesised to play a role in coral disease, the causative microbial agents of coral disease have only been identified in a few cases, and often Koch postulates prove inconclusive [81,82,83,84,85]. The utility of seawater microbes as indicators of poor reef health should not be dismissed even when causal relationships are unknown, as there are promising microbes (most notably Flavobacteriaceae-affiliated taxa) that are persistently diagnostic of reef ecosystem degradation across independent omics studies [36, 39, 52,53,54,55, 86, 87]. Below we provide a shortlist of seawater taxa/genes that have been experimentally validated as indicators of poor reef health, and highlight they should be further studied to provide richer insights into their potential (causal) role in reef degradation.
Experimental validation of the DDAM loop and the microbialisation concept has confirmed bacterioplankton communities of coral- and algae-dominated reefs persist in laboratory conditions. A bottle experiment identified that rates of bacterioplankton growth and utilisation of DOC were elevated in algal exudate treatments compared to incubations with coral exudates and the control treatment, with macroalgae-derived sugars selecting for a less diverse community enriched in lineages of opportunistic Gammaproteobacteria including putative pathogens with known virulence factors (Pseudoalteromonadaceae and Vibrionaceae) [88]. Another bottle experiment identified that high DOC concentrations (as observed in macroalgae-enriched reefs) correlated to an enrichment of bacterial genera Alteromonas, Oceanicola, Erythrobacter, and Alcanivorax, which shifted in their metabolic capabilities from mutualistic towards pathogenic states via up-regulation of genes encoding for metalloproteases, siderophores, toxins, and antibiotic resistance factors [89]. Similar trends were identified by in situ mesocosm studies which placed benthic chambers over coral-, sand-, and macroalgae-dominated communities, to identify that macroalgae exudates facilitated a shift to a net heterotrophic system, with pelagic microbial communities displaying elevated consumption of macroalgae-released DOC as well as increased oxygen consumption [52]. Experimental validation of the DDAM loop clearly shows that the addition of macroalgae-derived nutrients (under laboratory conditions) causes microbial proliferation and a shift towards pathogenesis and carbon metabolism pathways that are less energetically efficient [52, 88, 89], as also observed in the field [47]. However, the cause-and-effect understanding of the DDAM mechanism still needs to be teased apart to understand if and how the increased abundance and activity of microbial copiotrophs and putative pathogens in seawater at macroalgae-enriched reefs directly contribute to coral disease and reef declines, before applying the concept of microbialisation in predictive reef monitoring and proactive management.
Hypoxia also represents a crucial mechanism in the DDAM feedback loop, as macroalgae-released DOC fuels heterotophic activity and respiration by seawater microbes, which can create localised hypoxic regions at the coral-macroalgae interaction zones [47, 76, 90,91,92]. Experimental studies show that the addition of antibiotics may eliminate hypoxia in coral–algal interfaces [90,91,92]. As hypoxia predominantly occurs at coral–algal interaction zones, reef water away from the benthos may not be enriched in anaerobic microbial taxa and genes (but see [93]). We therefore propose that monitoring for anaerobic microbes and functions is particularly relevant at the benthic and pelagic boundary layer, which represents a potential valuable environmental niche for reef monitoring to predict rapid declines in benthic organisms caused by hypoxia.
Another forecasting potential of seawater microbial monitoring is to predict coral disease outbreaks. The concept that animal disease outbreaks are driven by environmental change (e.g. climate warming) is well accepted across many terrestrial, freshwater and marine ecosystems [85, 94,95,96,97,98], and acquisition of microbial pathogens from the environment has been documented in some food- and waterborne diseases [37, 99]. For corals, there are many examples of putative microbial pathogens (isolated from diseased coral tissues) also being identified in the surrounding reef seawater, which often increase in their abundance and activity at elevated seawater temperatures [86, 100, 101]. The bacterial genus Vibrio sp. is particularly prominent as a potential causative agent of coral disease [84, 85]. A survey to elucidate Vibrio diversity in surrounding reef seawater using the well-curated pyrH (uridylate kinase) gene sequence identified that putative coral pathogens (i.e. V. coralliilyticus, V. neptunis, and V. owensii) were persistently present in the seawater of the Ishigaki coral reef system (Japan) across the entire 3-year survey period, with increased abundances correlated with elevated seawater temperatures for the majority of Vibrio species [86]. Another study identified a significant enrichment in Planctomycetota (lineages OM190 and CL500-3) and bacteria within genera Synechococcus and Vibrio during the marine heatwave on the GBR in April 2016, alongside an enrichment of Vibrio-derived virulence factors (i.e. metalloprotease genes vcpA, vcpB and vchA in V. coralliilyticus) [101]. Though as the authors highlight, a link between large-scale changes observed in the plankton-associated microbial community and reef ecosystem health could not be established based on their observations [101], which warrants for robust experimental validation to gain a cause-and-effect understanding between the presence of potentially pathogenic water-born microorganisms and coral disease outbreaks.
Interestingly, the signal of copiotrophic and potentially pathogenic microbes in seawater often persists even after the environmental disturbances have passed [79, 101], and this concept is known as the microbial ‘legacy effect’ [78]. Anaerobic microbes can persist in seawater for days after dissolved oxygen concentrations revert to normoxic values [79], and the microbial signal of potentially pathogenic microbes (e.g. Planctomycetota and Vibrio) and functions (e.g. Vibrio-derived metalloproteases) that were enriched during the marine heatwave on the GBR in April 2016 remained apparent until August 2016, months after the marine heatwave had dissipated [101]. This microbial ‘legacy’ effect may be important to explore from a monitoring perspective of free-living seawater microbes, as a shift in microbial functioning may cause additional coral decline even after the disturbance has passed. As an example, a number of studies have documented that coral disease outbreaks can exacerbate the impacts of bleaching events [102,103,104,105,106,107]. The cumulative stress corals face during thermal stress may make them susceptible to opportunistic microbial pathogens that persist in the seawater following marine heatwaves [101], further compromising their health and increasing mortality. Experimentally validating this ‘legacy effect’ may be crucial from a monitoring perspective to understand how long opportunistic microbes and functions persists after different environmental disturbances, and to predict if this may affect future reef dynamics.
Formulation of seawater microbial indices for reef monitoring
Once experimentally validated, a list of microbial taxa and/or functions can be compiled to formulate robust microbial indicators which associate to metrics of poor reef health across both field and laboratory studies (Fig. 4, phase 3). Such efforts have already been made to formulate microbial indices for reef monitoring based on free-living seawater microbes. Most notably, microbialisation scores (defined as the ratio of metabolic rates between bacterioplankton and reef fish) have been proposed within the DDAM model as a metric of human impact on coral reefs [47, 76, 93, 108, 109]. This concept of microbialisation (a shift in biomass production and metabolic rates from macro to micro-organisms) has been well documented in macroalgae-enriched reefs in field observations [32, 109], across local and regional scales [47], and also validated experimentally in laboratory bottle experiments [88, 89, 110] and in situ mesocosm studies [52]. Despite their potential, microbialisation scores have not been implemented into standard reef monitoring efforts to date, primarily since the scores represent a metric relevant to shifting coral-algal dynamics, which is not universally applicable to reefs under environmental pressures that still maintain high coral cover and/or high fish biomass.

The proposed five-step framework of research and innovation to move from descriptive to predictive reef microbial monitoring, and from reactive to proactive reef management. Functional meta-omics datasets are critical to discover microbial indicators of poor reef health in the field (Phase 1), however high costs (see ‘Assay price’) and long bioinformatics processing times (see ‘Timeframes’) of microbial meta-omics datasets suggest their limited utility for rapid decision-making in reef management. We highlight that this milestone has been largely achieved through various localised studies, though in the years to come, the integration of recently generated datasets obtained in large-scale surveys (most notably the Tara Pacific Expedition) will be crucial to understand the ubiquity of identified microbial indicators at global scales. Once microbial indicators of poor ecosystem health are identified based on functional meta-omics datasets, experimental validation (Phase 2) is needed to confirm the same patterns occur in laboratory conditions, as well as to identify the causality of microbiome-environment associations from the field, which we predict still remains a distant goal and will require years of research. Once experimentally validated, microbial indices can be formulated (Phase 3) and applied research can commence to develop rapid (within weeks, days or minutes, see ‘Timeframes’) and cost-effective (see ‘Assay price’) assays to quickly assess reef health in the field (Phase 4), which can be used in proactive reef management and rapid decision-making (Phase 5)
A recent meta-analysis of reef bacterioplankton identified several microbial indices of poor reef health across the Great Barrier Reef [39]. It was proposed that Prochlorococcaceae and Synechococcaceae families represent potential indicators of cross-shelf nutrient levels with an increasing Synechococcaceae:Prochlorococcaceae abundance ratio being a proxy for increased nutrient loads in reef waters [39]. In addition, Flavobacteriaceae-affiliated taxa were potentially diagnostic of reef ecosystem degradation, with an increasing abundance ratio of copiotrophic (e.g. OCS155, Flavobacteraceae, Cryomorphaceae and Rhodobacteraceae) taxa relative to oligotrophic taxa (e.g. Pelagibacteraceae (SAR11) and SAR86) as an index of eutrophication [39]. Finally, an increasing prevalence of opportunistic and potentially pathogenic taxa (Rhodospirillaceae, Rhodobacteraceae and Vibrionaceae) were also indicative of degraded inshore reef systems [39]. Another study proposed that increased Bacteroidota (prevalent in waters of inshore reefs in the GBR) relative to Alphaproteobacteria (more abundant in offshore GBR reefs) in reef surface waters could indicate enhanced macroalgae growth, elevated nutrient loads, and the start of microbial proliferation for inshore coral reefs on the GBR, which aligns with the concept of the DDAM loop [87]. Recently, a detailed overview has been provided on indicator microbes across different reef benthic habitats (i.e. nearshore and offshore, or coral- and macroalgae-dominated), highlighting the ubiquity of these patterns in the Great Barrier Reef, Caribbean, and Pacific Ocean regions (Table 1 of [76]). This baseline knowledge is relevant as it provides a list of putative indicator microbes (with their expected relative abundances in different habitats) as stable predictors of poor reef health at broad spatio-temporal scales [76], which can be used as a starting point to create applied assays for rapid reef health assessment in the field using our framework (Fig. 4).
Despite their potential, seawater microbes are still largely overlooked by reef health surveys and these seawater microbial indices are yet to be validated as useful monitoring assays in the field (Fig. 4, phase 4). Such applied research should be pursued as the seawater microbiome possesses numerous additional characteristics that align with criteria of good indicators [30], in addition to its utility to infer and predict environmental fluctuations [36, 46]. Seawater sampling and processing is simple, non-destructive and can be performed with minimal training required, which facilitates large-scale sampling alongside ongoing in situ coral health surveys that already collect metrics on water chemistry and benthic cover in the reef. Furthermore, seawater collection and processing protocols are largely standardised due to global plankton sampling expeditions such as Tara Ocean and Tara Pacific [7, 11, 14, 56,57,58, 65, 75] which also ensures comparability of data streams from different studies. Lastly, while monitoring whole-community dynamics is preferred compared to focusing on a subset of indicators, it is simply not feasible for macro-organisms in highly biodiverse ecosystems such as coral reefs. However, whole-community monitoring can be done when working with seawater microbial communities, and tolerance thresholds to environmental disturbances can be determined for individual microbial species from seawater. Such information could be utilised to construct cumulative species and functional sensitivity distributions, allowing to quantify the proportional impact of environmental stress on the entire microbial communities [77].
Conclusions: a framework for incorporating microbial indicators into coral reef management
This review discusses the current state of using seawater microbes as indicators in predictive reef monitoring in the context of a five-step framework (Fig. 4), building on frameworks proposed by [64] and [111]. Many small-scale field studies combined with the emerging global studies (e.g. Tara Oceans) have identified candidate microbial taxa and genes that predict poor ecosystem and reef health [25, 31,32,33,34, 36, 39], hence the barrier does not lie in the indicator discovery phase (Fig. 4, phase 1) but largely in subsequent phases of experimental validation (Fig. 4, phase 2), formulation of microbial indices (Fig. 4, phase 3), applied research to generate microbial assays that can be used in the field (Fig. 4, phase 4), and implementation of microbial indicators in reef management and decision-making processes (Fig. 4, phase 5). Phase 1 (indicator discovery) is an ongoing process with large-scale multi-omics data streams, many that are yet to be published (e.g. Tara Pacific), fundamental to identify novel indicators and generate spatio-temporally coherent baselines of seawater microbial predictors that associate with metrics of poor reef health. Phases 2 (Experimental validation) and 3 (Formulation of seawater microbial indices to predict poor reef health) are currently a work in progress (Fig. 4) with a few studies experimentally validating microbial indicators (Fig. 4, phase 2), although primarily in the context of the DDAM loop, and we still lack conclusive experimental evidence that water-born microbial pathogens can indeed cause coral disease. Further, some indices based on seawater microbes (most notably the microbialisation scores) have been formulated to assess reef health (Fig. 4, phase 3), but these indices are still not used in standard reef monitoring.
Validated microbial-based molecular assays for rapid screening of seawater microbial indicators to predict reef decline are yet to be crafted (e.g. screening for anaerobic microbes to predict hypoxia-induced coral mortality events), hence phase 4 (applied research to craft microbial diagnostic and predictive tools) still needs to be developed. Such rapid and cost-effective assays based on seawater microbes (PCR, magnetic beads, and proteomic/colorimetric assays) have been successfully applied for environmental management (Fig. 4, phase 5), although generally inform on single stressors or have a narrow focus on risks to human health and well-being. Some examples include the presence/increased abundance of coliforms in public swimming waters indicating faecal pollution [112, 113], enrichment of antibiotic resistance genes indicating human impact [114], enrichment of hydrocarbon-degrading taxa and genes tracking oil spills [115] and anaerobic genes from sulphur-oxidising bacteria as indicators to trace the spread of oxygen minimum zones in the ocean [116]. Instead of assaying the reef environment for individual microbial indicators, it is potentially more productive to compile a list of target microbial taxa and functions that associate to poor reef health. For example, potentially pathogenic microbes increase in abundance and/or activity at elevated temperature and nutrient concentrations [85, 86, 101, 117]. Therefore screening for these taxa is a necessity during the summer period, particularly before, during and after bleaching events when coral health becomes compromised.
To move towards proactive reef management, improved communication between researchers and practitioners is needed to determine whether microbial indicators are desired in reef monitoring, as well as a cost/benefit analysis to identify which putative markers should be prioritised in applied research to develop targeted microbial-based assays. By reviewing current knowledge gaps, we highlight that seawater microbes should not be overlooked in reef monitoring efforts as marine plankton is an essential proxy of reef health, and we hope this review will catalyse further research towards predictive reef microbial monitoring and proactive management, which can be achieved if objectives are aligned between scientists, managers, and funding bodies.
Availability of data and materials
Not applicable.
Abbreviations
- IMOS:
-
Australia’s Integrated Marine Observing System
- GBR:
-
Great Barrier Reef
- DDAM:
-
Disease, dissolved organic carbon, algae, microbes
- DOC:
-
Dissolved organic carbon
- LDA:
-
Linear discriminant analysis
- rRNA:
-
Ribosomal ribonucleic acid
- DOM:
-
Dissolved organic matter
References
Moberg F, Folke C. Ecological goods and services of coral reef ecosystems. Ecol Econ. 1999;29:215–33.
Costanza R, Groot R, Sutton P, Van der Ploeg S, Anderson S, Farber S, et al. Changes in the global value of ecosystem services. Glob Environ Chang. 2014;26:152–8.
De’ath G, Fabricius KE, Sweatman H, Puotinen M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc Natl Acad Sci. 2012;109:17995–9.
Hoegh-Guldberg O, Poloczanska ES, Skirving W, Dove S. Coral Reef Ecosystems under climate change and Ocean acidification. Front Mar Sci. 2017;4:158.
Eddy TD, Lam VWY, Reygondeau G, Cisneros-Montemayor AM, Greer K, Palomares MLD, et al. Global decline in capacity of coral reefs to provide ecosystem services. One Earth. 2021;4:1278–85.
Sogin ML, Morrison HG, Huber JA, Welch DM, Huse SM, Neal PR, et al. Microbial diversity in the deep sea and the underexplored “rare biosphere.” Proc Natl Acad Sci. 2006;103:12115–20.
Abreu A, Bourgois E, Gristwood A, Troublé R, Acinas SG, Bork P, et al. Priorities for ocean microbiome research. Nat Microbiol. 2022;7:937–47.
Rusch DB, Halpern AL, Sutton G, Heidelberg KB, Williamson S, Yooseph S, et al. The sorcerer II Global Ocean sampling expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol. 2007;5: e77.
Karsenti E, Acinas SG, Bork P, Bowler C, De Vargas C, Raes J, et al. A Holistic approach to marine eco-systems biology. PLoS Biol. 2011;9: e1001177.
Duarte C. Seafaring in the 21St Century: The Malaspina 2010 Circumnavigation Expedition. Limnol Oceanography Bull. 2015;24.
Bork P, Bowler C, de Vargas C, Gorsky G, Karsenti E, Wincker P. Tara Oceans studies plankton at planetary scale. Introduction Sci. 2015;348:873.
Sunagawa S, Coelho LP, Chaffron S, Kultima JR, Labadie K, Salazar G, et al. Ocean plankton. Structure and function of the global ocean microbiome. Science. 2015;348:1261359.
Buttigieg PL, Fadeev E, Bienhold C, Hehemann L, Offre P, Boetius A. Marine microbes in 4D-using time series observation to assess the dynamics of the ocean microbiome and its links to ocean health. Curr Opin Microbiol. 2018;43:169–85.
Sunagawa S, Acinas SG, Bork P, Bowler C, Acinas SG, Tara Oceans Coordinators, et al. Tara Oceans: towards global ocean ecosystems biology. Nat Rev Microbiol. 2020;18:428–45.
Wild C, Huettel M, Klueter A, Kremb SG, Rasheed MYM, Jørgensen BB. Coral mucus functions as an energy carrier and particle trap in the reef ecosystem. Nature. 2004;428:66–70.
Furnas M, Mitchell A, Skuza M, Brodie J. In the other 90%: phytoplankton responses to enhanced nutrient availability in the Great Barrier Reef Lagoon. Mar Pollut Bull. 2005;51:253–65.
Garren M, Azam F. New directions in coral reef microbial ecology. Environ Microbiol. 2012;14:833–44.
Vanwonterghem I, Webster NS. Coral reef microorganisms in a changing climate. iScience. 2020;23:100972.
Bourne DG, Morrow KM, Webster NS. Insights into the coral microbiome: underpinning the health and resilience of reef ecosystems. Annu Rev Microbiol. 2016;70:317–40.
Pita L, Rix L, Slaby BM, Franke A, Hentschel U. The sponge holobiont in a changing ocean: from microbes to ecosystems. Microbiome. 2018;6:46.
Engelberts JP, Robbins SJ, de Goeij JM, Aranda M, Bell SC, Webster NS. Characterization of a sponge microbiome using an integrative genome-centric approach. ISME J. 2020;14:1100–10.
Marangon E, Laffy PW, Bourne DG, Webster NS. Microbiome-mediated mechanisms contributing to the environmental tolerance of reef invertebrate species. Mar Biol. 2021;168:89.
Robbins SJ, Song W, Engelberts JP, Glasl B, Slaby BM, Boyd J, et al. A genomic view of the microbiome of coral reef demosponges. ISME J. 2021;15:1641–54.
Moeller FU, Herbold CW, Schintlmeister A, Mooshammer M, Motti C, Glasl B, et al. Taurine as a key intermediate for host-symbiont interaction in the tropical sponge Ianthella basta. ISME J. 2023;17:1208–23.
Bourne D, Iida Y, Uthicke S, Smith-Keune C. Changes in coral-associated microbial communities during a bleaching event. ISME J. 2008;2:350–63.
Littman RA, Willis BL, Bourne DG. Metagenomic analysis of the coral holobiont during a natural bleaching event on the Great Barrier Reef. Environ Microbiol Rep. 2011;3(6):651–60.
McDevitt-Irwin JM, Baum JK, Garren M, Vega Thurber RL. Responses of coral-associated bacterial communities to local and global stressors. Front Mar Sci. 2017;4:262.
Fan L, Liu M, Simister R, Webster NS, Thomas T. Marine microbial symbiosis heats up: the phylogenetic and functional response of a sponge holobiont to thermal stress. ISME J. 2013;7:991–1002.
van Oppen MJH, Blackall LL. Coral microbiome dynamics, functions and design in a changing world. Nat Rev Microbiol. 2019;17:557–67.
Glasl B, Webster NS, Bourne DG. Microbial indicators as a diagnostic tool for assessing water quality and climate stress in coral reef ecosystems. Mar Biol. 2017;164:91.
Roitman S, Joseph Pollock F, Medina M. Coral microbiomes as bioindicators of reef health. In: Oleksiak MF, Rajora OP, editors. Population genomics: marine organisms. Cham: Springer International Publishing; 2018. p. 39–57. https://doi.org/10.1007/13836_2018_29.
Dinsdale EA, Pantos O, Smriga S, Edwards RA, Angly F, Wegley L, et al. Microbial ecology of four coral atolls in the Northern Line Islands. PLoS ONE. 2008;3: e1584.
Faust K, Lahti L, Gonze D, de Vos WM, Raes J. Metagenomics meets time series analysis: unraveling microbial community dynamics. Curr Opin Microbiol. 2015;25:56–66.
Peixoto RS, Rosado PM, de Leite DC, Rosado AS, Bourne DG. Beneficial microorganisms for corals (BMC): proposed mechanisms for coral health and resilience. Front Microbiol. 2017. https://doi.org/10.3389/fmicb.2017.00341.
Glasl B, Bourne D, Frade P, Webster N. Establishing microbial baselines to identify indicators of coral reef health. Microbiol Aust. 2018;39:42–6.
Glasl B, Bourne DG, Frade PR, Thomas T, Schaffelke B, Webster NS. Microbial indicators of environmental perturbations in coral reef ecosystems. Microbiome. 2019;7:94.
Semenza JC, Trinanes J, Lohr W, Sudre B, Löfdahl M, Martinez-Urtaza J, et al. Environmental suitability of Vibrio infections in a warming climate: an early warning system. Environ Health Perspect. 2017;125: 107004.
Leite DCA, Salles JF, Calderon EN, Castro CB, Bianchini A, Marques JA, et al. Coral bacterial-core abundance and network complexity as proxies for anthropogenic pollution. Front Microbiol. 2018;9:833.
Frade PR, Glasl B, Matthews SA, Mellin C, Serrão EA, Wolfe K, et al. Spatial patterns of microbial communities across surface waters of the great barrier reef. Commun Biol. 2020;3:442.
Hughes TP, Baird AH, Bellwood DR, Card M, Connolly SR, Folke C, et al. Climate change, human impacts, and the resilience of coral reefs. Science. 2003;301:929–33.
Glasl B, Smith CE, Bourne DG, Webster NS. Disentangling the effect of host-genotype and environment on the microbiome of the coral Acropora tenuis. PeerJ. 2019;7: e6377.
Glasl B, Smith CE, Bourne DG, Webster NS. Exploring the diversity-stability paradigm using sponge microbial communities. Sci Rep. 2018;8:8425.
Simister R, Taylor MW, Tsai P, Fan L, Bruxner TJ, Crowe ML, et al. Thermal stress responses in the bacterial biosphere of the Great Barrier Reef sponge, Rhopaloeides odorabile. Environ Microbiol. 2012;14:3232–46.
Luter HM, Gibb K, Webster NS. Eutrophication has no short-term effect on the Cymbastela stipitata holobiont. Front Microbiol. 2014. https://doi.org/10.3389/fmicb.2014.00216.
Pineda M-C, Strehlow B, Sternel M, Duckworth A, Jones R, Webster NS. Effects of suspended sediments on the sponge holobiont with implications for dredging management. Sci Rep. 2017;7:4925.
Galand PE, Ruscheweyh H-J, Salazar G, Hochart C, Henry N, Hume BCC, et al. Diversity of the Pacific Ocean coral reef microbiome. Nat Commun. 2023;14:3039.
Haas AF, Fairoz MFM, Kelly LW, Nelson CE, Dinsdale EA, Edwards RA, et al. Global microbialization of coral reefs. Nat Microbiol. 2016;1:16042.
Raj KD, Mathews G, Obura DO, Laju RL, Bharath MS, Kumar PD, et al. Low oxygen levels caused by Noctiluca scintillans bloom kills corals in Gulf of Mannar. India Sci Rep. 2020;10:22133.
Bush T, Diao M, Allen RJ, Sinnige R, Muyzer G, Huisman J. Oxic-anoxic regime shifts mediated by feedbacks between biogeochemical processes and microbial community dynamics. Nat Commun. 2017;8:789.
Johnson MD, Scott JJ, Leray M, Lucey N, Bravo LMR, Wied WL, et al. Rapid ecosystem-scale consequences of acute deoxygenation on a Caribbean coral reef. Nat Commun. 2021;12:4522.
Barott KL, Rohwer FL. Unseen players shape benthic competition on coral reefs. Trends Microbiol. 2012;20:621–8.
Haas AF, Nelson CE, Rohwer F, Wegley-Kelly L, Quistad SD, Carlson CA, et al. Influence of coral and algal exudates on microbially mediated reef metabolism. PeerJ. 2013;1: e108.
Weber L, González-Díaz P, Armenteros M, Ferrer VM, Bretos F, Bartels E, et al. Microbial signatures of protected and impacted Northern Caribbean reefs: changes from Cuba to the Florida keys. Environ Microbiol. 2020;22:499–519.
Wambua S, Gourlé H, de Villiers EP, Karlsson-Lindsjö O, Wambiji N, Macdonald A, et al. Cross-sectional variations in structure and function of coral reef microbiome with local anthropogenic impacts on the Kenyan coast of the Indian Ocean. Front Microbiol. 2021. https://doi.org/10.3389/fmicb.2021.673128.
Angly FE, Heath C, Morgan TC, Tonin H, Rich V, Schaffelke B, et al. Marine microbial communities of the Great Barrier Reef lagoon are influenced by riverine floodwaters and seasonal weather events. PeerJ. 2016;4: e1511.
Gorsky G, Bourdin G, Lombard F, Pedrotti ML, Audrain S, Bin N, et al. Expanding Tara oceans protocols for underway, ecosystemic sampling of the ocean-atmosphere interface during Tara Pacific expedition (2016–2018). Front Mar Sci. 2019. https://doi.org/10.3389/fmars.2019.00750.
Planes S, Allemand D, Agostini S, Banaigs B, Boissin E, Boss E, et al. The Tara Pacific expedition—a pan-ecosystemic approach of the “-omics” complexity of coral reef holobionts across the Pacific Ocean. PLoS Biol. 2019;17: e3000483.
Lombard F, Bourdin G, Pesant S, Agostini S, Baudena A, Boissin E, et al. Open science resources from the Tara Pacific expedition across coral reef and surface ocean ecosystems. Sci Data. 2023;10:324.
Salazar G, Sunagawa S. Marine microbial diversity. Curr Biol. 2017;27:R489–94.
Lombard F, Boss E, Waite AM, Vogt M, Uitz J, Stemmann L, et al. Globally consistent quantitative observations of planktonic ecosystems. Front Mar Sci. 2019;6:196.
Cordier T, Alonso-Sáez L, Apothéloz-Perret-Gentil L, Aylagas E, Bohan DA, Bouchez A, et al. Ecosystems monitoring powered by environmental genomics: a review of current strategies with an implementation roadmap. Mol Ecol. 2021;30:2937–58.
Beale DJ, Jones OAH, Bose U, Broadbent JA, Walsh TK, van de Kamp J, et al. Omics-based ecosurveillance for the assessment of ecosystem function, health, and resilience. Emerg Top Life Sci. 2022;6:185–99.
Djemiel C, Dequiedt S, Karimi B, Cottin A, Horrigue W, Bailly A, et al. Potential of meta-omics to provide modern microbial indicators for monitoring soil quality and securing food production. Front Microbiol. 2022. https://doi.org/10.3389/fmicb.2022.889788.
Parkinson JE, Baker AC, Baums IB, Davies SW, Grottoli AG, Kitchen SA, et al. Molecular tools for coral reef restoration: beyond biomarker discovery. Conserv Lett. 2020;13: e12687.
Belser C, Poulain J, Labadie K, Gavory F, Alberti A, Guy J, et al. Integrative omics framework for characterization of coral reef ecosystems from the Tara Pacific expedition. Sci Data. 2023;10:326.
Tully BJ, Graham ED, Heidelberg JF. The reconstruction of 2,631 draft metagenome-assembled genomes from the global oceans. Sci Data. 2018;5: 170203.
Pachiadaki MG, Brown JM, Brown J, Bezuidt O, Berube PM, Biller SJ, et al. Charting the complexity of the marine microbiome through single-cell genomics. Cell. 2019;179:1623-1635.e11.
Faure E, Ayata S-D, Bittner L. Towards omics-based predictions of planktonic functional composition from environmental data. Nat Commun. 2021;12:4361.
Frémont P, Gehlen M, Vrac M, Leconte J, Delmont T, Wincker P, et al. Restructuring of plankton genomic biogeography in the surface ocean under climate change. Nat Clim Chang. 2022;12:1–9.
Rohart F, Gautier B, Singh A, Lê Cao K-A. mixOmics: an R package for ‘omics feature selection and multiple data integration. PLoS Comput Biol. 2017;13: e1005752.
Alneberg J, Bennke C, Beier S, Bunse C, Quince C, Ininbergs K, et al. Ecosystem-wide metagenomic binning enables prediction of ecological niches from genomes. Commun Biol. 2020;3:1–10.
Chaffron S, Delage E, Budinich M, Vintache D, Henry N, Nef C, et al. Environmental vulnerability of the global ocean epipelagic plankton community interactome. Sci Adv. 2021;7:eabg1921.
Guidi L, Chaffron S, Bittner L, Eveillard D, Larhlimi A, Roux S, et al. Plankton networks driving carbon export in the oligotrophic ocean. Nature. 2016;532:465–70.
Tagliabue A. ‘Oceans are hugely complex’: modelling marine microbes is key to climate forecasts. Nature. 2023;623:250–2.
Planes S, Allemand D. Insights and achievements from the Tara Pacific expedition. Nat Commun. 2023;14:3131.
Nelson CE, Wegley Kelly L, Haas AF. Microbial interactions with dissolved organic matter are central to coral reef ecosystem function and resilience. Annu Rev Mar Sci. 2023;15:431–60.
Webster NS, Wagner M, Negri AP. Microbial conservation in the Anthropocene. Environ Microbiol. 2018;20:1925–8.
Correa-Garcia S, Constant P, Yergeau E. The forecasting power of the microbiome. Trends in Microbiology [Internet]. 2022 [cited 2023 Mar 7]; Available from: https://www.sciencedirect.com/science/article/pii/S0966842X22003213
Doyle SM, Self MJ, Hayes J, Shamberger KE, Correa AM, Davies SW, et al. Microbial community dynamics provide evidence for hypoxia during a coral reef mortality event. Appl Environ Microbiol. 2022;88:e00347-e422.
Louca S, Polz MF, Mazel F, Albright MBN, Huber JA, O’Connor MI, et al. Function and functional redundancy in microbial systems. Nat Ecol Evol. 2018;2:936–43.
Alker AP, Smith GW, Kim K. Characterization of Aspergillus sydowii (Thom et Church), a fungal pathogen of Caribbean sea fan corals. Hydrobiologia. 2001;460:105–11.
Bourne DG, Garren M, Work TM, Rosenberg E, Smith GW, Harvell CD. Microbial disease and the coral holobiont. Trends Microbiol. 2009;17:554–62.
Sutherland KP, Shaban S, Joyner JL, Porter JW, Lipp EK. Human pathogen shown to cause disease in the threatened Eklhorn Coral Acropora palmata. PLoS ONE. 2011;6: e23468.
Bourne DG, Smith HA, Page CA. Diseases of scleractinian corals. In: Rowley AF, Coates CJ, Whitten MW, editors. Invertebrate pathology. Oxford: Oxford University Press; 2022. https://doi.org/10.1093/oso/9780198853756.003.0004.
Munn CB. The role of vibrios in diseases of corals. Microbiol Spectr. 2015;3:3–4.
Amin AKMR, Feng G, Al-saari N, Meirelles PM, Yamazaki Y, Mino S, et al. The first temporal and spatial assessment of Vibrio diversity of the surrounding seawater of coral reefs in Ishigaki, Japan. Front Microbiol. 2016. https://doi.org/10.3389/fmicb.2016.01185.
Glasl B, Robbins S, Frade PR, Marangon E, Laffy PW, Bourne DG, et al. Comparative genome-centric analysis reveals seasonal variation in the function of coral reef microbiomes. ISME J. 2020;14:1435–50.
Nelson CE, Goldberg SJ, Wegley Kelly L, Haas AF, Smith JE, Rohwer F, et al. Coral and macroalgal exudates vary in neutral sugar composition and differentially enrich reef bacterioplankton lineages. ISME J. 2013;7:962–79.
Cárdenas A, Neave MJ, Haroon MF, Pogoreutz C, Rädecker N, Wild C, et al. Excess labile carbon promotes the expression of virulence factors in coral reef bacterioplankton. ISME J. 2018;12:59–76.
Smith JE, Shaw M, Edwards RA, Obura D, Pantos O, Sala E, et al. Indirect effects of algae on coral: algae-mediated, microbe-induced coral mortality. Ecol Lett. 2006;9:835–45.
Brown A, Carpenter R. Water-flow mediated oxygen dynamics within massive Porites-algal turf interactions. Mar Ecol Prog Ser. 2013;490:1–10.
Jorissen H, Skinner C, Osinga R, de Beer D, Nugues MM. Evidence for water-mediated mechanisms in coral–algal interactions. Proc Biol Sci. 2016;283:20161137.
Walsh K, Haggerty JM, Doane MP, Hansen JJ, Morris MM, Moreira APB, et al. Aura-biomes are present in the water layer above coral reef benthic macro-organisms. PeerJ. 2017;5: e3666.
Harvell CD, Mitchell CE, Ward JR, Altizer S, Dobson AP, Ostfeld RS, et al. Climate warming and disease risks for terrestrial and marine biota. Science. 2002;296:2158–62.
Harvell D, Jordán-Dahlgren E, Merkel S, Rosenberg E, Raymundo L, Smith G, et al. Coral disease, environmental drivers, and the balance between coral and microbial associates. Oceanography (Washington DC). 2007;20:58–81.
Altizer S, Ostfeld RS, Johnson PTJ, Kutz S, Harvell CD. Climate change and infectious diseases: from evidence to a predictive framework. Science. 2013;341:514–9.
Maynard J, van Hooidonk R, Harvell CD, Eakin CM, Liu G, Willis BL, et al. Improving marine disease surveillance through sea temperature monitoring, outlooks and projections. Philos Trans R Soc B Biol Sci. 2016;371:20150208.
Cavicchioli R, Ripple WJ, Timmis KN, Azam F, Bakken LR, Baylis M, et al. Scientists’ warning to humanity: microorganisms and climate change. Nat Rev Microbiol. 2019;17:569–86.
Semenza JC, Herbst S, Rechenburg A, Suk JE, Höser C, Schreiber C, et al. Climate change impact assessment of food- and waterborne diseases. Crit Rev Environ Sci Technol. 2012;42:857–90.
Tout J, Siboni N, Messer LF, Garren M, Stocker R, Webster NS, et al. Increased seawater temperature increases the abundance and alters the structure of natural Vibrio populations associated with the coral Pocillopora damicornis. Front Microbiol. 2015. https://doi.org/10.3389/fmicb.2015.00432.
Doni L, Oliveri C, Lasa A, Di Cesare A, Petrin S, Martinez-Urtaza J, et al. Large-scale impact of the 2016 Marine Heatwave on the plankton-associated microbial communities of the Great Barrier Reef (Australia). Mar Pollut Bull. 2023;188: 114685.
Selig ER, Drew Harvell C, Bruno JF, Willis BL, Page CA, Casey KS, et al. Analyzing the relationship between ocean temperature anomalies and coral disease outbreaks at broad spatial scales. In: Phinney JT, Hoegh‐Guldberg O, Kleypas J, Skirving W, Strong A, editors. Coastal and estuarine studies [Internet]. Washington, D. C.: American Geophysical Union; 2006 [cited 2023 Mar 7]. p. 111–28. Available from: http://www.agu.org/books/ce/v061/61CE07/61CE07.shtml
Brandt ME, McManus JW. Disease incidence is related to bleaching extent in reef-building corals. Ecology. 2009;90:2859–67.
Miller J, Muller E, Rogers C, Waara R, Atkinson A, Whelan KRT, et al. Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs. 2009;28:925–37.
Randall CJ, van Woesik R. Contemporary white-band disease in Caribbean corals driven by climate change. Nat Clim Change. 2015;5:375–9.
Precht WF, Gintert BE, Robbart ML, Fura R, van Woesik R. Unprecedented disease-related coral mortality in Southeastern Florida. Sci Rep. 2016;6:31374.
Brodnicke OB, Bourne DG, Heron SF, Pears RJ, Stella JS, Smith HA, et al. Unravelling the links between heat stress, bleaching and disease: fate of tabular corals following a combined disease and bleaching event. Coral Reefs. 2019;38:591–603.
Jackson JBC, Kirby MX, Berger WH, Bjorndal KA, Botsford LW, Bourque BJ, et al. Historical overfishing and the recent collapse of coastal ecosystems. Science. 2001;293:629–37.
McDole T, Nulton J, Barott KL, Felts B, Hand C, Hatay M, et al. Assessing Coral Reefs on a Pacific-Wide Scale Using the Microbialization Score. PLoS ONE. 2012;7: e43233.
Silveira CB, Luque A, Roach TN, Villela H, Barno A, Green K, et al. Biophysical and physiological processes causing oxygen loss from coral reefs. Elife. 2019;8: e49114.
Webster N, Gorsuch H. Monitoring additional values within the Reef 2050 Integrated Monitoring and Reporting Program: final report of the microbes expert group [Internet]. Great Barrier Reef Marine Park Authority. 2020. Available from: https://elibrary.gbrmpa.gov.au/jspui/handle/11017/3594
Bushon RN, Brady AM, Likirdopulos CA, Cireddu JV. Rapid detection of Escherichia coli and enterococci in recreational water using an immunomagnetic separation/adenosine triphosphate technique. J Appl Microbiol. 2009;106:432–41.
Rosario K, Symonds EM, Sinigalliano C, Stewart J, Breitbart M. Pepper Mild Mottle Virus as an indicator of fecal pollution. Appl Environ Microbiol. 2009;75:7261–7.
Li B, Yang Y, Ma L, Ju F, Guo F, Tiedje JM, et al. Metagenomic and network analysis reveal wide distribution and co-occurrence of environmental antibiotic resistance genes. ISME J. 2015;9:2490–502.
Laroche O, Wood SA, Tremblay LA, Ellis JI, Lear G, Pochon X. A cross-taxa study using environmental DNA/RNA metabarcoding to measure biological impacts of offshore oil and gas drilling and production operations. Mar Pollut Bull. 2018;127:97–107.
Walsh DA, Zaikova E, Howes CG, Song YC, Wright JJ, Tringe SG, et al. Metagenome of a versatile chemolithoautotroph from expanding oceanic dead zones. Science. 2009;326:578–82.
Kimes NE, Grim CJ, Johnson WR, Hasan NA, Tall BD, Kothary MH, et al. Temperature regulation of virulence factors in the pathogen Vibrio coralliilyticus. ISME J. 2012;6:835–46.
Acknowledgements
This work took place at the Australian Institute of Marine Science (AIMS) headquarters at Cape Ferguson, and we wish to acknowledge the Wulgurukaba and Bindal peoples as the Traditional Owners of that land. This research was also undertaken at the JCU Townsville Bebegu Yumba campus, and the authors acknowledge that the Australian Aboriginal and Torres Strait Islander peoples are the original inhabitants and traditional custodians of this continent and have unique cultural and spiritual relationships to the land and waters. We pay our respects to their Elders past, present, and emerging.
Funding
This study forms part of the Australia’s Integrated Marine Observing System (IMOS) Great Barrier Reef Microbial Genomic Database sub-facility, funded by the Queensland Research Infrastructure Co-investment Fund (RICF) by the Department of Environment and Science, Queensland. IMOS is enabled by the National Collaborative Research Infrastructure Strategy (NCRIS). It is operated by a consortium of institutions as an unincorporated joint venture, with the University of Tasmania as Lead Agent. This study was also funded by an AIMS@JCU PhD Scholarship to M.T. The funders had no role in performing the literature review, preparation of the manuscript, or decision to publish.
Author information
Authors and Affiliations
Contributions
M.T performed the literature search. M.T, D.G.B and P.W.L conceived the review topic and scoped the sections. M.T, D.G.B and P.W.L wrote the manuscript, and S.R, Y.K.Y, P.R.F, B.G, and N.S.W reviewed and edited the manuscript, and provided input on the full manuscript. M.T and B.G created the figures.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Terzin, M., Laffy, P.W., Robbins, S. et al. The road forward to incorporate seawater microbes in predictive reef monitoring. Environmental Microbiome 19, 5 (2024). https://doi.org/10.1186/s40793-023-00543-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40793-023-00543-4