
Abstract
Shewanella algae strain C6G3, isolated from the 2 uppermost centimeters of muddy sediment of Arcachon Bay (SW Atlantic French coast, sampled in October 2007) has the capability to use a large panel of terminal electron acceptors under anaerobic condition, such as nitrate, nitrite and metal-oxide, and presents a great metabolic versatility. Here, we present the non-contiguous draft-genome sequence of Shewanella algae C6G3, which consists of a 4,879,425 bp. The chromosome contains 5792 predicted genes. In total, the genome consists of 24 rRNA genes, 86 tRNA genes and 5660 genes assigned as protein-coding genes.
Similar content being viewed by others

Introduction
The genus Shewanella comprises several Gram-negative species which are widely distributed in marine and freshwater environments. Shewanella algae (formerly classified as S. putrefaciens ) has been frequently isolated from marine water samples and spoiling fish [1–3]. They are capable of reducing trimethylamine N-oxide (TMAO) to trimethylamine and producing hydrogen sulfide, both of which are main components of the fishy odor present during low temperature storage. They were also isolated from human feces, skin and other clinical samples [4, 5]. The collected strains were heterogeneous with G + C values ranging from 43 % to 55 %. However, there were differences between environmental and clinical isolates. Most of the strains isolated from human clinical specimens and identified as S. putrefaciens showed beta-hemolysis on sheep blood agar whereas environmental strains were nonhemolytic [6, 7]. During a screening study of heterotrophic bacteria from the sediment of Arcachon Bay [8], a large set of isolates was obtained from different sampling sites and years (2). Among the 24 isolates, 15 strains belong to Shewanella genus and were able to reduce Mn(III/IV) and/or nitrate. The genus seems to play an important role in the turnover of organic matter coupled to anaerobic respiration electron acceptors, such as Fe(III), Mn(III/IV) and NO2/3. Here we report on further taxonomic and physiological studies on strain Shewanella algae strain C6G3 and present its main genomic features.
Organism information
Classification and features
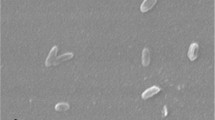
The genus Shewanella currently contains 62 species [9] including Shewanella algae ATCC 51192T, the first described [2]. Ribosomal gene of strain C6G3 exhibits 99 % similarity with available ribosomal gene of Shewanella algae (strains ATCC 51192T, ACDC [3], BrY [10] and FeRed [11]) and was affiliated to this specie (Fig. 1). Cells of strain C6G3 are straights rods (Fig. 2), Gram-negative, motile, free-living and non-sporulating. Different growth temperatures, pH and % NaCl were tested (Table 1). Optimal growth occurs at 30 °C, pH 8 and 10 % NaCl (w/v). For strain C6G3, ATCC 51192T and S. oneidensis MR-1T [12], the use of 95 carbon sources was tested with Biolog GN2 microplate™ (Microlog) (Additional file 1: Table S1). Strain C6G3 presents 32 positive results: 5/30 carbohydrates, 14/29 organic acids, 8/19 amino-acids. Similar results were obtained for strain ATCC 511392T (38/95). Among the two strains of S. algae , slightly different patterns of carbon source were noticed; however, profil of S oneidensis MR-1T was different (16/95). Some electron acceptors were also tested for strain C6G3 according to genome annotation and Shewanella algae literature [1] (Additional file 1: Table S1).

Phylogenetic position of Shewanella algae C6G3 relative to the genus Shewanella and other strains of Shewanella algae. This Neighbor-joining tree is based on 1243 aligned characters of the 16S rRNA gene. The bootstrap percentages higher than 50 % are indicated at the node after 1000 resampled data sets. Branch length corresponds to sequence differences as indicated on the scale bar (substitutions per position). The proposed Shewanella species have been chosen from the List of Prokaryotic names with Standing in Nomenclature (type strain and sequence accession number) for their ability to use nitrate, nitrite and / or metal oxides. Species whose genome has been sequenced are marked with star (*). Escherichia coli [44] was used as out-group

Phase contrast micrograph of Shewanella algae C6G3. Bar scale: 2 μm
Chemotaxonomic data
The fatty acid analysis was performed on two strains of S. algae (C6G3, ATCC 51192T) and on S. oneidensis MR-1T (Additional file 1: Table S2). At the end of aerobic culture, fatty acids were extracted from cell pellet by alkaline hydrolysis and analyzed using chromatography-electron ionization mass spectrometry (GC-EIMS) following the protocol described in Zabeti et al., [13]. The overall fatty acid pattern of S. algae C6G3 is rather common for the genus Shewanella . The major ones were C16:1ω7 (35.2 %), C16:0 (34.6 %) (generally reported between 16–55 % and 5–31 %, respectively [14]) and 3OH-C12:0 (7.7 %). Interestingly, the C15:0 br is much lower in strain C6G3 (2.3 %) than in S. algae ATCC 51192T and S. oneidensis MR-1T (27.4 % and 20.4 %, respectively). S. algae C6G3 presents also a relatively higher percentage of short-chain fatty acids (shorter than C15, 16.0 %) than both S. oneidensis MR-1T and S. algae ATCC 51192T (8.5 %).
Genome sequencing information
Genome project history
S. algae C6G3 was isolated from intertidal marine sediment on the basis of its ability to use large range of electron acceptors particularly nitrate, nitrite and metal-oxides [8]. The genome of S. algae C6G3 is the second to be reported from that species, the other one being S. algae ACDC [15]. The genome project of S. algae C6G3 is deposited in the Genome On Line Database [16]. A summary of the project and information on compliance with MIGS version 2.0 [17] are shown (Table 2).
Growth conditions and DNA isolation
S. algae C6G3 was grown aerobically at 30 °C under stirring condition on artificial sea water [18] amended with lactate (3 g/L), yeast extract (1 g/L) and tryptone (5 g/L). DNA was extracted from cells collected in exponential growth phase using the protocol of Marteinsson et al. [19]. DNA concentration and purity were checking on biophotometer® (Eppendorf) before sequencing.
Genome sequencing and assembly
The genome sequencing of S. algae C6G3 was generated at the Molecular Research LP MR DNA Laboratory (USA). De novo whole-genome shotgun sequencing was performed using the Ion Torrent PGM (Life Technologies [20]) sequencing platform. This produced 1,444,981 reads with an average length of 200 bp for a total number of sequenced bases of 288,996,200 representing a sequencing depth of 50-fold. The assembly of S. algae C6G3 genome was generated at MR DNA Laboratory using the SeqMan NGen® software assembler (DNASTAR). The final assembly identified 43 contigs generating a genome size of 4,9 Mb.
Genome annotation
Genome annotation was performed on two platforms: on RAST [21] and on IMG/ER [22] (DOE Joint Genome Institute [23]). The tRNAscan-SE tool [24] (RAST and IMG/ER) was used to find tRNA genes, whereas ribosomal RNAs were detected using RMAmmer [25] (IMG/ER) and tool “search_for_rnas” (developed by Niels Larsen (available by the author), RAST). Open Reading Frames (ORFs) were predicted using GLIMMER2 [26] in RAST and using GeneMark (v.2.6.r) [27] and GenePRIMP [28] as a part of the DOE-JGI genome annotation pipeline. Gene prediction analyses and functional annotations were performed in RAST with a series of BLAST against FIG hands-curated subsystems [29]. They were also analyzed with FIGfams collection databases, and through comparative approaches with Integrated Microbial Genome – Expert Review platform (RPS-BLAST, BLAST, BLASTp, Hmmsearch (HMMER)) against non-redundant databases including COGs, Pfam [30], TIGR-fam [31], KEEG [32], IMG. Additional functional annotations were performed within the SEED framework (RAST) [33] and the IMG/ER (DOE-JGI) platform.
Genome properties
The assembly of non-contiguous finished draft genome consists of 43 contigs representing overall 4,879,425 pb. The DNA G + C content was 53.08 %. Using RAST and IMG/ER, 5770 and 5795 genes were respectively predicted. Among them 4149 and 5660 protein-coding genes and 108 and 135 RNAs were identified by RAST and IMG/ER, respectively. The properties and the statistics of the genome (IMG/ER data) are summarized in Table 3 and Fig. 3. Putative COG functions were assigned for 39.28 % of the protein-coding genes. The distribution of genes into COGs functional categories is presented in Table 4.

Graphical map of 6 contigs containing ORF involved in nitrate, nitrite and metal oxides utilization. Nitrate reduction (contig #203 and #65), dissimilative reduction of nitrite into ammonium, (contig #82 and #35) and metal reduction (contig #105 and #166). From bottom to the top: genes on forward strand (color by COG), genes on reverse strand (color by COG), operon/gene cited (pointed red), RNA genes (tRNAs green, rRNAs red, other RNAs black), GC content, GC skew
Insights into the genome sequence
Genome of S. algae C6G3 encodes genes for complete glycolysis and tricarboxylic acid (TCA) cycle. A focus has been made on the enzymes involved in the reductive respiratory reactions of the N-cycle (NAP, NRFA) and in extracellular electron transfer through the outer-membrane (Metal Transfer Reducing such as Fe(III) and Mn(III/IV)) on the basis of protein system described in S. oneidensis MR-1T.
Nitrate reduction
Nitrate respiration involves two distinct enzyme systems: the NapAB localized in the periplasm and the membrane-bound nitrate reductase NarGHI enzyme localized on the cytoplasmic face of the cytoplasmic membrane. The sole nitrate reductase of S. algae C6G3 is NapAB (Fig. 4). As most of the Shewanella species, S. algae C6G3 genome encodes the two NAP isoforms, each comprising three catalytic subunits: NapA where nitrate reduction takes place, a di-haem cytochrome NapB and a maturation chaperone NapD. The two isoforms present different membrane-intrinsic subunits [34] named NAP-α (NapEDABC) and NAP-β (NapDAGHB). NAP-β (NapDAGBH) possesses NapGH, an iron–sulfur cluster ferredoxins instead of NapC. The functional differences between these systems may be explained by differential regulation in the composition of the available quinol pool. S. oneidensis MR-1T encodes only Nap-β isoform in which NapC is lacking (Fig. 4). This membrane-anchored tetrahem c-Cyt mediates electron transport from the quinol pool to NapB. The function of NapC in NAP-β in S. oneidensis MR-1T may be met by CymA, an homologue of periplasmic tetrahem c-Cyt of the NapC/NirT family, which is also found in S. algae C6G3 genome.

NAP complex functioning. The proposed electron-transfer pathway of periplasmic nitrate reductase and membrane bound electron donors of S. algae C6G3 (NAP-α-β) (a) and S. oneidensis MR-1T (NAP-β) (b). Colored proteins are annotated from KEGG and putatively functional
Dissimilative reduction of nitrite into ammonium
Nitrite can be reduced to ammonium (NH4) by a periplasmic nitrite reduction system (NRF) [35]. As NAP systems, there are two types of NRF: NrfABCD and NrfAH types. NrfA is the terminal reductase while NrfBCD/NrfH are responsible for electron transfer from menaquinol pool to NrfA. Shewanella strains are known for encoding NrfABCD system only. Genome annotations of S. algae C6G3 and S. oneidensis MR-1T identify nrfA in both cases but nrfBCD were found in S. algae C6G3 only (Fig. 5). As previously described for NAP system, S. oneidensis MR-1T genome lacks genes encoding for the specific compounds that deliver electrons to the terminal reductase (nrfBCD). Indeed, nrfB is lacking and nrfCD are present but proposed to be pseudogene because of truncation [36]. Gao et al., [37] suggested that the tetraheme c-Cyt CymA, a cytoplasmic membrane electron transport protein, is likely to be the functional replacement of both NapC and NrfBCD/NrfH allowing to NrfAH-like system to be efficient in S. oneidensis MR-1T.

NRF complex functioning. The proposed electron-transfer pathway of dissimilatory nitrite reduction to ammonium of S. algae C6G3 (a) and S. oneidensis MR-1T (b). Colored proteins are annotated from KEGG and putatively functional. Streaked genes are annotated but probably not functional
Metal oxide reduction
S. algae C6G3 can utilize extracellular mineral metal oxides of Fe(III) and Mn(III/IV) as respiratory electron acceptors (unpublished data). Inspection of its genome confirmed the presence of genes involved in pathway (i.e. metal-reducing or MTR pathway) for transferring electrons from the inner membrane through the periplasm and across the outer membrane where metal oxides are reduced [38]. In S. algae C6G3 genome, genes that encode MtrCBA and OmcA are located in the same region, which also includes mtrD (an mtrA homologue), mtrE (an mtrB homologue) and mtrF (an mtrC homologue) (Fig. 6).

MTR pathway functioning. The proposed MTR extracellular electron-transfer pathway of S. algae C6G3 (a) and S. oneidensis MR-1T (b). Colored proteins are annotated from KEGG and putatively functional
Table 5 shows the BLASTP for amino acid sequences of MTR pathway in S. algae C6G3 versus those in metal-reducing S. oneidensis MR-1T .
The numbers of genes found in the MTR clusters of the analyzed Shewanella strains varies from four, such as omcA1-mtrC-mtrA-mtrB in Shewanella frigidimarina [11], to nine, such as mtrD-mtrE-mtrF-omcA1-undB-omcA1-mtrC-mtrA-mtrB in Shewanella halifaxensis [39, 40]. CymA identified as the entry point for electrons into the MTR pathway [41] is not located in the MTR gene cluster in S. algae C6G3 as described in S. oneidensis MR-1T. Furthermore, S. algae C6G3 has an additional protein encoding for decahem c-Cyt of the OmcA/MtrC family. The role of this cytochrome is not defined. On RAST platform, this ORF has been annotated as mtrH in S. algae C6G3 and S. halifaxensis HAW-EB4T.
Conclusion
The Shewanella genus comprises a diverse group of facultative anaerobes. Their ability to couple the oxidation of various carbon sources to the reduction of a broad range of terminal electron acceptors imparts a respiratory flexibility that allows colonization of varied and changeable marine and freshwater environments [39, 42, 43]. The occurrence of the two different NAP operons, NRF, and that of CymA in S. algae C6G3 accords with the renowned anaerobic respiratory flexibility of Shewanella . S. algae C6G3 is also capable of using solid Fe(III) and Mn(III/IV) as terminal electron acceptors. Reduction of these particulates occurs at the cell surface and is catalyzed by multihaem cytochromes whose properties are beginning to emerge.
Abbreviations
- TMAO:
-
Trimethylamine N-oxide
References
Caccavo F, Blakemore RP, Lovley DR. A Hydrogen-Oxidizing, Fe(III)-Reducing Microorganism from the Great Bay Estuary. New-Hampshire Appl Environ Microb. 1992;58(10):3211–6.
Simidu U, Kitatsukamoto K, Yasumoto T, Yotsu M. Taxonomy of 4 Marine Bacterial Strains That Produce Tetrodotoxin. Int J Syst Bacteriol. 1990;40(4):331–6.
Clark IC, Melnyk RA, Engelbrektson A, Coates JD. Structure and Evolution of Chlorate Reduction Composite Transposons. Mbio. 2013;4(4):e00379-13. doi:10.1128.
Beleneva IA, Magarlamov TY, Eliseikina MG, Zhukova NV. Biochemical and pathogenic properties of the natural isolate of Shewanella algae from Peter the Great Bay, Sea of Japan. J Invertebr Pathol. 2009;102(3):250–5. doi:10.1016/j.jip.2009.09.001.
Gram L, Bundvad A, Melchiorsen J, Johansen C, Vogel BF. Occurrence of Shewanella algae in Danish coastal water and effects of water temperature and culture conditions on its survival. Appl Environ Microb. 1999;65(9):3896–900.
Nozue H, Hayashi T, Hashimoto Y, Ezaki T, Hamasaki K, Ohwada K, et al. Isolation and Characterization of Shewanella-Alga from Human Clinical Specimens and Emendation of the Description of S-Alga Simidu Et-Al, 1990, 335. Int J Syst Bacteriol. 1992;42(4):628–34.
Baumann P, Gauthier MJ, Baumann L. Genus Alteromonas. New York: Springer; 1984.
Javanaud C, Michotey V, Guasco S, Garcia N, Anschutz P, Canton M et al. Anaerobic ammonium oxidation mediated by Mn-oxides: from sediment to strain level. Res Microbiol. 2011;162(9):848–57. doi:10.1016/j.resmic.2011.01.011.
Dawyndt P, Verheyde B, De Baets B, De Vos P, Van Brabant B, Gillis W et al. http://www.straininfo.net/taxa/1507. straininfonet. 2014.
Nevin KP, Lovley DR. Mechanisms for Fe(III) oxide reduction in sedimentary environments. Geomicrobiol J. 2002;19(2):141–59. doi:10.1080/01490450252864253.
Bowman JP, McCammon SA, Nichols DS, Skerratt JH, Rea SM, Nichols PD, et al. Shewanella gelidimarina sp. nov. and Shewanella frigidimarina sp. nov., novel Antarctic species with the ability to produce eicosapentaenoic acid (20:5 omega 3) and grow anaerobically by dissimilatory Fe(III) reduction. Int J Syst Bacteriol. 1997;47(4):1040–7.
Venkateswaran K, Moser DP, Dollhopf ME, Lies DP, Saffarini DA, MacGregor BJ, et al. Polyphasic taxonomy of the genus Shewanella and description of Shewanella oneidensis sp. nov. Int J Syst Bacteriol. 1999;49:705–24.
Zabeti N, Bonin P, Volkman JK, Guasco S, Rontani JF. Fatty acid composition of bacterial strains associated with living cells of the haptophyte Emiliania huxleyi. Org Geochem. 2010;41(7):627–36. doi:10.1016/j.orggeochem.2010.04.009.
Brenner DJ, Krieg NR, Staley JT, Garrity GM, Boone DR, De Vos P et al. Family I. Altermonadaceae. Genus XIII. Shewanella. Bergey's Manual of Systematic Bacteriology. Volume 2: The Proteobacteria, Part B: The Gammaproteobacteria. New York Springer; 2005.
Clark IC, Melnyk RA, Engelbrektson A, Coates JD. Structure and Evolution of Chlorate Reduction Composite Transposons. Mbio. 2013;4(4). doi:ARTN e00379-13. doi:10.1128/mBio.00379-13.
Pagani I, Liolios K, Jansson J, Chen IMA, Smirnova T, Nosrat B et al. The Genomes OnLine Database (GOLD) v.4: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res. 2012;40(D1):D571-D9. doi:10.1093/Nar/Gkr1100.
Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P et al. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol. 2008;26(5):541–7. doi:10.1038/Nbt1360.
Baumann P, Baumann L, Mandel M. Taxonomy of marine bacteria: the genus Beneckea. J Bacteriol. 1971;107(1):268–94.
Marteinsson VT, Watrin L, Prieur D, Caprais JC, Raguenes G, Erauso G. Phenotypic Characterization, DNA Similarities, and Protein Profiles of 20 Sulfur-Metabolizing Hyperthermophilic Anaerobic Archaea Isolated from Hydrothermal Vents in the Southwestern Pacific-Ocean. Int J Syst Bacteriol. 1995;45(4):623–32.
Rothberg JM, Hinz W, Rearick TM, Schultz J, Mileski W, Davey M et al. An integrated semiconductor device enabling non-optical genome sequencing. Nature. 2011;475(7356):348–52. doi:10.1038/Nature10242.
Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA et al. The RAST server: Rapid annotations using subsystems technology. Bmc Genomics. 2008;9. doi:Artn 75. doi:10.1186/1471-2164-9-75.
Markowitz VM, Chen IMA, Palaniappan K, Chu K, Szeto E, Pillay M et al. IMG 4 version of the integrated microbial genomes comparative analysis system. Nucleic Acids Res. 2014;42(D1):D560-D7. doi:10.1093/Nar/Gkt963.
Mavromatis K, Ivanova NN, Chen IMA, Szeto E, Markowitz VM, Kyrpides NC. The DOE-JGI Standard Operating Procedure for the Annotations of Microbial Genomes. Stand Genomic Sci. 2009;1(1):63–7. doi:10.4056/Sigs.632.
Lowe TM, Eddy SR. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997;25(5):955–64. doi:10.1093/Nar/25.5.955.
Lagesen K, Hallin P, Rodland EA, Staerfeldt HH, Rognes T, Ussery DW. RNAmmer: consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007;35(9):3100–8. doi:10.1093/Nar/Gkm160.
Delcher AL, Harmon D, Kasif S, White O, Salzberg SL. Improved microbial gene identification with GLIMMER. Nucleic Acids Res. 1999;27(23):4636–41. doi:10.1093/nar/27.23.4636.
Besemer J, Lomsadze A, Borodovsky M. GeneMarkS: a self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001;29(12):2607–18. doi:10.1093/nar/29.12.2607.
Pati A, Ivanova NN, Mikhailova N, Ovchinnikova G, Hooper SD, Lykidis A et al. GenePRIMP: a gene prediction improvement pipeline for prokaryotic genomes. Nat Methods. 2010;7(6):455-U62. doi:10.1038/Nmeth.1457.
Overbeek R, Begley T, Butler RM, Choudhuri JV, Chuang HY, Cohoon M et al. The subsystems approach to genome annotation and its use in the project to annotate 1000 genomes. Nucleic Acids Res. 2005;33(17):5691–702. doi:10.1093/Nar/Gki866.
Finn RD, Bateman A, Clements J, Coggill P, Eberhardt RY, Eddy SR et al. Pfam: the protein families database. Nucleic Acids Res. 2014;42(D1):D222-D30. doi:10.1093/Nar/Gkt1223.
Haft DH, Selengut JD, Richter RA, Harkins D, Basu MK, Beck E. TIGRFAMs and Genome Properties in 2013. Nucleic Acids Res. 2013;41(D1):D387-D95. doi:10.1093/Nar/Gks1234.
Kanehisa M, Araki M, Goto S, Hattori M, Hirakawa M, Itoh M et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008;36:D480-D4. doi:10.1093/Nar/Gkm882.
The SEED framework for comparative genomics. In: http://www.theseed.org. 2014.
Simpson PJL RD, Codd R. The periplasmic nitrate reductase in Shewanella: the resolution, distribution and functional implications of two NAP isoforms, NapEDABC and NapDAGHB. Microbiology. 2010;156:302–12. doi:I 10.1099.
Simon J. Enzymology and bioenergetics of respiratory nitrite ammonification. Fems Microbiol Rev. 2002;26(3):285–309. doi: S0168-6445(02)00111-0. doi:10.1111/J.1574-6976.2002.Tb00616.X.
Romine MF, Carlson TS, Norbeck AD, Mccue LA, Lipton MS. Identification of mobile elements and pseudogenes in the Shewanella oneidensis MR-1 genome. Appl Environ Microb. 2008;74(10):3257–65. doi:10.1128/Aem.02720-07.
Gao HC, Yang ZK, Barua S, Reed SB, Romine MF, Nealson KH et al. Reduction of nitrate in Shewanella oneidensis depends on atypical NAP and NRF systems with NapB as a preferred electron transport protein from CymA to NapA. Isme J. 2009;3(8):966–76. doi:10.1038/ismej.2009.40.
Shi L, Rosso KM, Clarke TA, Richardson DJ, Zachara JM, Fredrickson JK. Molecular Underpinnings of Fe(III) Oxide Reduction by Shewanella Oneidensis MR-1. Front Microbiol. 2012;3:50. doi:10.3389/fmicb.2012.00050.
Fredrickson JK, Romine MF, Beliaev AS, Auchtung JM, Driscoll ME, Gardner TS et al. Towards environmental systems biology of Shewanella. Nat Rev Microbiol. 2008;6(8):592–603. doi:10.5301/RU.2013.11500.
Zhao JS, Manno D, Leggiadro C, O'Neill D, Hawari J. Shewanella halifaxensis sp nov., a novel obligately respiratory and denitrifying psychrophile. Int J Syst Evol Micr. 2006;56:205–12. doi:10.1099/ijs.0.63829-0.
Myers CR, Myers JM. Cloning and sequence of cymA a gene encoding a tetraheme cytochrome c required for reduction of iron(III), fumarate, and nitrate by Shewanella putrefaciens MR-1. J Bacteriol. 1997;179(4):1143–52.
Hau HH, Gralnick JA. Ecology and biotechnology of the genus Shewanella. Annu Rev Microbiol. 2007;61:237–58. doi:10.1146/annurev.micro.61.080706.093257.
Marritt SJ, Lowe TG, Bye J, McMillan DGG, Shi L, Fredrickson J et al. A functional description of CymA, an electron-transfer hub supporting anaerobic respiratory flexibility in Shewanella. Biochem J. 2012;444:465–74. doi:10.1042/Bj20120197.
Castellani A, Chalmers AJ. Genus Escherichia Castellani and Chalmers. 3rd ed. Williams: Wood and Co.; 1919.
Woese CR, Kandler O, Wheelis ML. Towards a Natural System of Organisms - Proposal for the Domains Archaea, Bacteria, and Eucarya. P Natl Acad Sci USA. 1990;87(12):4576–9. doi:10.1073/pnas.87.12.4576.
Baumann L, Baumann P, Mandel M, Allen RD. Taxonomy of aerobic marine eubacteria. J Bacteriol. 1972;110:402–29.
Skerman VBD, Mcgowan V, Sneath PHA. Approved lists of bacterial names. Int J Syst Bacteriol. 1980;30(1):225–420.
Garrity GM, Bell JA, Phylum LT, XIV. Proteobacteria phyl. nov. Bergey’s Manual of Systematic Bacteriology. New York: Springer; 2005.
Garrity GM, Bell JA, Class LT, III. Gammaproteobacteria class. nov. Bergey’s Manual of Systematic Bacteriology. New York: Springer; 2005.
Editor L. Validation list no. 106. Validation of publication of new names and new combinations previously effectively published outside the IJSEM. Int J Syst Evol Micr. 2005;55:2235–8.
Bowman JP, McMeekin TA. Order X. Alteromonadales ord. nov. New York: Springer; 2005.
Ivanova EP, Flavier S, Christen R. Phylogenetic relationships among marine Alteromonas-like proteobacteria: emended description of the family Alteromonadaceae and proposal of Pseudoalteromonadaceae fam. nov., Colwelliaceae fam. nov., Shewanellaceae fam. nov., Montellaceae fam. nov., Ferrimonadaceae fam. nov., Idiomarinaceae fam. nov and Psychromonadaceae fam. nov. Int J Syst Evol Micr. 2004;54:1773–88. doi:10.1099/ijs.0.02997-0.
Macdonell MT, Colwell RR. Phylogeny of the Vibrionaceae, and Recommendation for 2 New Genera, Listonella and Shewanella. Syst Appl Microbiol. 1985;6(2):171–82.
Editor L. Validation list no. 20. Validation of publication of new names and new combinations previously effectively published outside the IJSB. Int J Syst Evol Micr. 1986;36:354–6.
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene Ontology: tool for the unification of biology. Nat Genet. 2000;25(1):25–9.
Acknowledgements
We are grateful to Dr. Scot E. Dowd (Molecular Research LP MR DNA Laboratory) for sequencing, assembly and annotation on RAST of our genome. We are grateful to the U.S. Department of Energy Joint Genome Institute for annotation on IMG/ER platform of our genome. And, we are also grateful to Jean-François Rontani (Researcher at Mediterranean Institute of Oceanography (M. I. O.), Marseille, France) and Rémi Amiraux (phD at M. I. O.) for extraction and analysis of fatty acid. This work has been support by ANR IZOFLUX (ANR-10-BLAN-0612) and PROTIDAL (ANR-06-BLAN-0040) grants.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interest.
Authors’ contributions
AA carried out the molecular genetic studies, the sequence alignment and participated in write manuscript. VM and PB conceived the study, and participated in its design and coordination and drafted the manuscript. All authors read and approved the final manuscript.
Additional files
Additional file 1:
Table S1. Presentation of positives carbon sources (Biolog GN2 microplateTM) & electron acceptors for S. algae strain C6G3 and ATCC 51192T and S. oneidensis MR-1T (differences are distinct in bold type). Table S2. Main fatty acids composition (90.6 %) of S. algae C6G3 and percentage of this fatty acids in S. algae ATCC 51192T (73.6 % of total pattern) and S. oneidensis MR-1T (92.5 % of total pattern).
Additional file 2:
Associated MIGS Record.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Aigle, A., Michotey, V. & Bonin, P. Draft-genome sequence of Shewanella algae strain C6G3. Stand in Genomic Sci 10, 43 (2015). https://doi.org/10.1186/s40793-015-0022-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40793-015-0022-0