
Abstract
Background
Understanding the role of agroecosystems as habitat for wildlife is crucial for long-term conservation planning, as different crop stratification and landscape elements can influence bird communities, which are also affected by seasonality. The goal of our study was to determine how agricultural landscapes varying in land cover characteristics affect bird richness and abundance. Bird surveys were conducted at 110 locations within agricultural landscapes in central Chile. The surveyed areas were characterized by land cover at two scales (50 and 500 m radii) through direct observation and photo-interpretation, during winter and spring seasons. Generalized Linear Mixed Models were used to evaluate the effects of different agricultural land covers on bird species and communities.
Results
Our results show that birds were more abundant during winter, in particular for insectivorous and granivorous birds, and that bird species richness was significantly increased due to cover provided by hedgerows at the plot scale.
Conclusions
We found that abundance of some bird species in agroecosystems in central Chile was higher in winter than in spring, and that overall bird richness was favored by structural diversity including non-crop structures such as hedgerows, which thus may be relevant for improving bird conservation management in temperate agroecosystems. Our results suggest that native vegetation proximity and area may affect seasonal changes in bird communities at larger scales, relationships which warrant further study.
Similar content being viewed by others


Background
Agriculture is one of the main threats to biodiversity conservation throughout the world [1,2,3]. Among the limited number of wild species able to persist in agroecosystems, birds are particularly important, for example in biological control of agricultural pests [4, 5] or using birds as bio-indicators of agricultural sustainability [6]. Bird presence in agroecosystems depends on the type of crop and its structural heterogeneity (e.g., a grassland with low vertical stratification vs trees with high vertical stratification), as well as the type of agricultural management and landscape composition [7,8,9]. For example, Verhulst et al. [10] found that intensive conventional management of vineyards (high input of external agrochemicals) significantly decreased bird richness, compared with an abandoned vineyard. A long-term study in Costa Rica showed that low intensity management in agroecosystems (e.g., polycultures with high structural diversity) can provide higher resilience and stability of the bird community than high intensity agricultural management [11]. Moorcroft et al. [12] found that the abundance of some granivorous species of conservation concern were associated with high amounts of weed seed and a larger proportion of bare soil. Bare ground in agroecosystems has been observed to be an important component of habitat for ground-foraging insectivorous birds [13]. Diversified farming systems that increase structural and temporal diversity of crops (polycultures, crop rotations, hedgerows) and native vegetation (riparian vegetation, forest fragments, and forest) within the farm can favor the presence of wildlife in agroecosystems [14], for example, by providing habitat for insect pollinators through non-crop hedgerows in California [14, 15] or in the UK, where crop rotation (annual crops), fallows, and non-crop hedgerows were found positive for birds [16].
Landscape heterogeneity and vegetation structure can enhance the richness and abundance of birds within agroecosystems by increasing the number of ecological niches [17, 18]. For example, bird richness was higher in Hungarian vineyards with greater diversity of landscape elements (e.g., shrubs) [10], and shaded coffee polycultures harbored more bird species than monoculture coffee farms [19]. Likewise, Mulwa et al. [20] reported that bird richness and abundance were higher in subsistence agriculture polycultures than in high intensity sugarcane monocultures. Vegetation diversity and structure influence bird presence at plot and landscape scales, with some forest birds preferring diverse environments instead of homogeneous ones [21]. Laiolo [22] found that vineyards were utilized by birds as habitat, but that this relationship depended more on the effects of the landscape matrix and season than on the vineyard itself. Luther et al. [23] found that bird abundance was related with plot and context variables, while bird richness was related solely with plot-level variables, such as shrub species richness.
Temperate agroecosystems follow seasonal abiotic variations that may influence wildlife communities due to the availability of food resources [24, 25]. In the Himalayas, Elsen et al. [26] demonstrated the importance of low intensity agriculture (small farms with diverse annual crops, orchards and small woodlots) for increasing abundance of forest birds during winter. Additionally, Guyot et al. [27] reported a significant seasonal influence on bird abundance in vineyards in Switzerland, with greater abundance during winter. The effects of intensive agricultural landscapes on bird communities in temperate regions of South America are poorly documented, yet understanding the role of agricultural lands as habitat for birds is crucial for long-term conservation planning in this region, particularly because the land surface area formally protected for conservation purposes in parks and reserves is low [28]. The aim of this study was to determine the role of agricultural landscapes as bird habitat in central Chile. We hypothesized a seasonal influence on bird community composition. Secondly, we expected that agricultural land cover with more vertical complexity would support more species than less stratified land cover, due to their similarity to native vegetation (scrubland) with highly vertical stratification. Thirdly, we expected higher bird abundance in low stratification crops due to their association with bird flocks. Finally, we expected that more specialized scrubland birds (e.g. ground-dwelling species, such as Rhinocryptids), would be strongly associated with the presence of native vegetation (high structural complexity).
Methods
Study area
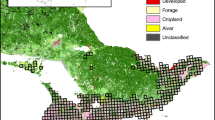
The study was conducted in agricultural landscapes south of the city of Santiago, Chile (Fig. 1), one of the most developed agricultural regions in the country. The landscape is composed of different annual crops, prairies, fallows, orchards, and vineyards. The region has a Mediterranean climate (mean precipitation: 419 mm; mean temperature: 13.6 °C) [29]. The native vegetation in the area is dominated by sclerophyllous woodlands (e.g., Quillaja saponaria) and scrublands (e.g., Acacia caven) [30].

Location of bird survey plots in agricultural areas around Santiago, Chile. Maipo river is shown in the middle of the map, and Santiago city is shown in grey at the top of the map
Bird surveys
During the austral winter (June) and spring (October- November) of 2009 we recorded all the birds sighted and heard in 10-min, 50-m radius point counts at 110 survey locations (Fig. 1) [31, 32]. Birds were surveyed between 08:00 and 12:00 h by the same observer (A.M.S.), always under fair weather conditions. Surveys were conducted from lightly trafficked roads bordering agricultural areas. No counts were conducted in the presence of passing vehicles. In addition to counting all birds seen and heard, we recorded visibility as a covariate thought to affect the probability of detection. In order to assess visibility, we estimated the percentage of visibility from each of the survey points (average ± SD, 81.7% ± 18.5). The feeding behavior of all birds analyzed in the study was summarized from [33, 34] and scientific and common names were obtained from [35].
Land cover characteristics
We visually estimated land cover based on the percentage of crop or non-agricultural vegetation we observed at two spatial scales: plot (p) and landscape (L), using 50 and 500 m radii around the survey point, respectively. In addition to a visual assessment in the field, we estimated the area of each land use type through photo-interpretation of satellite images from Google Earth (Mountain View, California, USA), with a resolution of 0.43 m pixel size and processed with the ArcGIS 9.2 software (Redlands, California, USA) [36]. Table 1 shows the land cover classes used for both spatial scales.
Statistical analyses
In order to determine the relative influence of plot and landscape-scale variables and season on bird abundance, we used two methods: Model Averaging of Generalized Linear Mixed Models (GLMMs) [37,38,39] and Canonical Correspondence Analysis (CCA) [40, 41]. These techniques were chosen because GLMMs allow the selection of a species-specific subset of explanatory variables, while CCA allows all species to be modeled using the same set of environmental variables in order to obtain a graphic result for the community [42]. Model-averaging coefficients in GLMMs allows incorporating uncertainty from several models rather than relying on selecting a single best model, thus increasing the likelihood of a more robust prediction [39, 43]. All analyses were performed in R (version 3.3.0 [44]. To avoid problems involved in modelling rare species, we conservatively restricted all bird analyses to include those species and groups for which we had records in five or more plots per season [45,46,47,48]. We evaluated the effect of season by comparing community abundance, richness, and bird dietary groups using paired t-Student tests.
Prior to running our GLMMs and CCA models, we tested multicollinearity between variables by calculating Pearson’s correlations between all pairs to ensure that no variables were strongly correlated (|r| < 0.45) [37, 38, 49]. Due to the high correlation between low crops (p), low crops (L), orchards (p), and orchards (L) for all species, we chose to run models with only one of these four variables; the variable selected varied by species and was chosen according to the highest R squared in an intra-specific model comparison (see Table 1 and Additional file 1). Because the predictor variables for land cover were in different scales, we standardized all the variables to a mean of 0 and standard deviation of one beforehand to enable comparisons.
We then compared the Poisson and Negative Binomial distributions to assess the best model fit for each species. We used “odTest” function in the “pscl” package in R to test for overdispersion with a likelihood ratio (LR) test of the null hypothesis that the data followed a Poisson distribution [50]. Based on this analysis, we used the Negative Binomial family of GLMMs in all models except for the House wren (winter) and richness (both seasons), in which we used a Poisson distribution due to better fit. Finally, we fit GLMMs using the package “lme4” [51], with visibility as a random effect, and performed model averaging using the package “MuMIn” [52]. All models were ranked according to the QAICc, a quasi-likelihood version of AICc suitable for over-dispersed count data [43]. Variables included in the most parsimonious models with ΔQAICc <2 were identified by averaging their estimated coefficients and associated standard errors [43]. Coefficients and standard errors for the variables that had p-values <0.05 are presented in the Additional file 1. The adjusted-R2 value for each model was calculated as the average of the adjusted-R2 values of the most parsimonious models [38]. The performance of models was assessed by examining the spread of model residuals and the adjusted-R2 values.
To identify dominant relationships between each bird species and land cover, we used a linear combination of environmental variables that allowed for the species niches to be described in a canonical correspondence analysis (CCA) [40]. In order to make unbiased comparisons between the GLMM and CCA analyses, we used the same variables that were used in the GLMM analysis. However, in order to adequately visualize the relevance of each land cover variable at two scales (plot and landscape), we included all the variables analyzed in the study (see Table 1). CCA assumes that species’ responses along environmental gradients are unimodal and reduces the species data set to a few orthogonal gradients (i.e., CCA axes), which reflect the influence of the environmental variables included in the analysis [40, 53].
Results from CCA were visually represented on an ordination graph. We plotted weighted averages in order to find the relationships between birds and agricultural land cover and to allow better representation of the species abundance [40, 54, 55]. We conducted separate CCAs for data collected in winter and spring to explore whether there was seasonal variation in the relationship between species abundance and environmental variables. Finally, we applied a Monte Carlo test with 999 permutations to evaluate the significance of each CCA [56].
Results
We recorded 42 bird species in the study area, but only 27 species (Table 2) were analyzed according to our model selection criteria. Results from the t-test indicated that there was significantly greater abundance in winter (non-breeding) compared with the spring (breeding) season (P < 0.001, Fig. 2), while richness was only marginally greater in winter compared with spring (P = 0.0504, Fig. 2). Insectivores (t-test, P = 0.006) and native granivores (t-test, P = 0.002) had significantly lower abundance in spring, whereas exotic granivores and omnivores (t-test, P = 0.14, and P = 0.06, respectively) showed no significant difference in abundance between seasons (Fig. 3).

Comparison of abundance of individuals and species richness (± SE) between seasons (N = 111 plots). Different letters represent significant differences between seasons

Abundance (± SE) between diet groups between seasons (N = 111 plot stations). Different letters represent significant differences among each group. Number of species for winter and spring were, respectively, insectivores 6 and 5, granivores-native 7 and 6, granivores-exotic 3 and 3 and omnivores 6 and 7
Overall, individual species models indicated that during the winter, species were significantly positively affected with the proportion of shrub hedgerows at the plot scale followed by the cover of orchards at plot and/or landscape scale (Figs. 4 and 5). Other variables affecting the winter abundance of bird species were the percentage cover of low crops (p), the amount of urban development, and road cover. During the spring, low crops and orchards at both scales had a significant positive influence on the abundance of several species (low crops five species, orchards three species, Figs. 4 and 5). Road cover and urban development also positively influenced the abundance of two species and one negatively (Figs. 4 and 5). Native vegetation cover was positively related to the abundance of Fire eyed diucon in both seasons and Austral blackbird only in spring.

Relationship between species and land covers and scales during winter (left) and spring (right). Only significant results are showns

Significant positive and negative effects (GLMMs, p < 0.05) of land covers affecting birds at different scales in a winter and b spring
The first two CCA axes explained 62.6 and 59.9% of the canonical variance in winter and spring, respectively (Fig. 6). At the community level, CCA showed that hedgerows and native vegetation were associated with higher number of species in winter. In spring this pattern changed and orchards and low crops grouped the majority of the species. A few species were associated with the same agricultural land cover characteristics during both seasons, including the Southern lapwing and Chilean swallow (associated with low crops at both scales), and Rock dove (associated with urban development (L)). Groups of birds based on diet did not appear to be associated with any particular land cover type.

Plot of the first two axes of the canonical correspondence analysis (CCA) ordination between bird and agricultural land covers in central Chile in winter a and spring b. Acronyms appear in Table 2. Land covers at landscape scale are indicated with (L)
Discussion
Bird composition (abundance and species) were affected by agricultural land cover, and various species could persist within agroecosystems. Our study shows that agricultural environments provide habitat for a significant proportion (56%, see Additional file 1) of the avifauna of central Chile, when wetland, high elevation mountains, and coastal habitats are not considered [57]. Season strongly affected the relative abundance and diversity of birds in agricultural landscapes in the study area. As we hypothesized, this effect was stronger in winter, when shelter from cold weather conditions and food resource scarcity are issues. The seasonal effect that allowed higher diversity and abundance in winter is likely be related to more mild weather conditions (i.e., warmer temperature, lower precipitation) in our study area the central valley compared to nearby mountain habitats. The seasonal altitudinal movements of birds have previously described in Chile by Cody [58] and Kelt et al. [24]. Guyot et al. [27] found similar patterns in which bird richness and abundance were higher in winter within vineyards. Agricultural landscapes likely provide some alternative feeding resources during winter for birds that breed in higher elevations, including seeds and invertebrates [59, 60]. Guyot et al. [27] suggested that seasonality patterns in bird abundance were related with presence of winter flocks, a pattern also observed in this study for some species (e.g. Black-chinned siskin, Southern lapwing). Additionally, we also suggest that other altitudinal (Dark-faced ground-tyrant) and longitudinal (Green backed firecrown) migrations may play a role in increasing the abundance and richness in central Chile [61]. In addition, the increase in abundance of native birds during winter in agricultural areas could also be related with a reduction in the management operations within the agricultural fields (labors, use of agrochemicals, etc.) that also can favor the quality of agroecosystem as habitat during winter.
Land cover effect
Our models showed that non-crop land cover in agricultural lands play a key role in structuring bird communities in the agroecosystems of central Chile. Hedgerows at the plot scale increased the abundance of the majority of species analyzed in this study during winter (GLMMs, Fig. 5a; CCA Fig. 6a), and may contribute to species supplementary sources of food/shelter during winter time. Hedgerows represented a relative small proportion of the total study area (8%), but could be playing an important role as habitat, probably because of their structural similarity with the native scrub [62], that could increase resource availability (e.g. shelter, foraging areas) [17, 18]. In a study conducted at a similar scale, Pithon et al. [63] found that only Woodlarks (Lullula arborea) and Eurasian skylark (Alauda arvensis) were associated with orchards (vineyards), while the majority of the species that used agroecosystems were associated with semi-naturals habitat in French agroecosystems. Similar results have been reported in California agroecosystems, where the tree rows at the edges of alfalfa crops significantly increased the avian richness [5]. Other studies support the relevance of isolated trees [64] and hedgerows as factors that favor bird diversity in agricultural landscapes [65,66,67,68,69,70,71,72]. Here, our findings further highlight the relevance of shrub hedgerows for bird communities, documenting the benefits to biodiversity of retaining non-crop land cover characteristics in Chilean agroecosystems.
Native vegetation appears to play a strong role in structuring Chilean bird communities in agricultural lands, particularly in winter (Fig. 6a). Although native vegetation was not found to be a significant predictor in our models, this may be due to the fact that the amount of native vegetation in our study area was relatively small (2%). This likely resulted in some native scrub specialists being recorded very infrequently. Consequently, based on our criteria for inclusion in the analysis, we were unable to analyze these species. In fact, species such as the Chilean mockingbird (Mimus thenca), Moustached turca (Pteroptochos megapodius), Grey-hooded sierra finch (Phrygilus gayi), Variable hawk (Geranoaetus polyosoma), and Chilean flicker (Colaptes pitius), were mostly observed in the plots with some native vegetation cover. Thus, our results suggest that native vegetation increases the richness of the species in agricultural landscapes, and thus can be an especially important to recover to conserve or restore biodiversity in Chilean agricultural landscapes.
Considering the scarcity of native vegetation embedded in the studied agricultural landscapes and the relevance of hedgerows for supporting bird richness, the latter are clearly playing a key role at larger scales of analysis, opening opportunities for future research within Chilean agroecosystems. According to ODEPA [73], 83% of farms in central Chile are smaller than 10 ha, from which we argue that the proportion of hedgerows and live fences could be increased by planting native species along the metal wire fences. In the same way, Fahrig et al. [74] suggest that diversified small farms provide habitat heterogeneity favoring biodiversity, and avoid the impacts of biotic homogenization associated with large crop monocultures [75, 76].
In general, our results (Fig. 5) suggest that land covers at plot scale were more relevant in influencing bird abundance and richness during winter, while these relationships were less clear during spring. A similar study in French agroecosystems found that smaller scales (110 m to 330 m radii) better predicted bird presence although the pattern depended on the species [47]. In tropical ecosystems, Castaño-Villa et al. [77] found that vegetation measurements at fine scale (50 m2) influence significantly insectivore birds. Studies in other organisms (bees) showed that the importance of the native vegetation proportion as a predictor of bees abundance change according different landscape scales [78]. Based on our results, we suggest future research should evaluate the relevance of land cover elements at the plot scale as landscape features for improving connectivity for birds within the agricultural matrix.
Species associations
Most of the patterns observed for individual species were consistent with general expectations and previous observations (e.g. the high association between Southern lapwings and agricultural crops with low stratification [79]. Also, the fact that Austral thrushes were abundant within orchards during both seasons suggests that these omnivorous species can adapt to dynamic agricultural habitats, likely taking advantage of resources present during different seasons (e.g. fruit during spring, and invertebrates during winter) [80]. Some of these species may play a beneficial role as predators of insects, in particular soil insect larvae, potentially helping control agricultural pests within agroecosystems [4, 5, 81, 82].
Species associated with urban environments, such as the House sparrow and Eared dove, may also benefit from the presence of houses and other constructions, and not require native vegetation. Interestingly, exotic granivores was the only group that increased slightly during spring, in contrast to the native birds (Fig. 3). This should be considered for future studies due to the potential negative impacts of these exotic birds on agricultural yields during the growing season [83]. We expected to document a clear relationship between granivorous abundance and percentage cover of low stratified crops, but our results did not support our hypothesis. Interestingly, instead, some granivores such as Black-chinned siskin and Rufous-collared sparrow were mainly associated with highly stratified crops such as orchards, although it has been reported that both species can behave as facultative, changing their diet according to the availability of the resources, including insects [84,85,86]. This could be related with weed plants that were growing and seeding in the inter row of the orchards, a variable that was not measured in this study, but may be relevant for future research. This also may be related to previous research showing that open low stratification vegetation was not part of the original Chilean matorral [30, 87, 88]. Additionally, granivores could be positively related to orchards due to the protection that these areas offer from predation by birds of prey [60]. In contrast, other studies support a preference of granivorous for low stratification crops (e.g. fallow areas) [89] due to the presence of weeds as seed resources [90]. In our study, this relationship was observed with the exotic House sparrow in spring, opening the question of whether habitat preferences vary between exotic and native granivorous species.
Most of the insectivores analyzed in this study differ in their feeding behavior, which could account for the lack of a general pattern of habitat association for this guild. On the contrary, the lack of such a pattern for omnivores was expected. This suggests that other factors may be more relevant in determining its presence within agricultural landscapes, such as the soil tillage and subsequent exposition of soil invertebrates for Southern lapwing and Chimango caracara [91] or the presence of berries and other complementary food resources for Austral thrush [92].
Although our results showed an association between bird species distribution and agricultural land cover, the amount of unexplained variation (based on our CCA) suggests the influence of other important drivers not considered in this study. Similarly, analysis over larger landscape scales could be relevant for some species, depending on their home ranges, but predictions of which scales may be relevant were limited in our study due to the lack of literature on the bird communities and number of species involved. An additional limitation of our study was that all the bird counts were conducted from roads, potentially limiting the detectability of some species. However, we considered this factor in the field and we incorporated it as a random effect in our models (see methods). In spite of these limitations, we believe that our study presents relevant information to increase the limited knowledge of avifauna within Chilean agroecosystems.
Between 2009 and 2015, the total area of low crops in the study region decreased from 26,763 to 18,404 ha, while the area of orchards increased from 75,239 to 77,303 ha [93]. Agricultural management practices (e.g. organic vs conventional) and levels of intensification are known to influence wildlife [14, 94]. Our results suggest that this land use change could affect the bird communities present in these landscapes, and this should be considered in land use planning within agroecosystems in the region. Our results in Chile support the findings of previous literature from elsewhere that changes in farming systems, via diversification of landscape structures, could increase the use of these agricultural environments by birds. Managing agroecosystems for bird habitat conservation at the plot scale, by using native plants in hedgerows, for example, should be included within a broader management frame in order to benefit biodiversity at the landscape scale while also favoring connectivity [95, 96].
Conclusions
Agricultural landscapes provided habitat for an important number of native birds. The relevance of agroecosystems for birds changed by season, with increased abundance of birds in winter. Hedgerows, as non-crop structures, can play a key role as habitat within these agroecosystems, especially during winter. These results provide novel information that could be used for land use management and planning in the central Chile biodiversity hotspot.
Abbreviations
- CCA:
-
Canonical Correspondence Analysis
- GLMMs:
-
Generalized Linear Mixed Models
- ODEPA:
-
Oficina de Estudios y Políticas Agrarias (Chile), Chilean Department of Agricultural Studies and Policy
- SD:
-
Standard deviation
References
Balmford A, Green R, Phalan B. What conservationists need to know about farming. Proc Biol Sci. 2012;279:2714–24.
Krebs JR, Wilson JD, Bradbury RB, Siriwardena GM. The second silent spring? Nature. 1999;400:611–2.
Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, et al. Global consequences of land use. Science. 2005;309:570–4.
Jedlicka J, Greenberg R, Letourneau DK. Avian conservation practices strengthen ecosystem services in California vineyards. PLoS One. 2011;6:e27347.
Kross SM, Kelsey TR, Mccoll CJ, Townsend JM. Field-scale habitat complexity enhances avian conservation and avian-mediated pest-control services in an intensive agricultural crop. Agric Ecosyst Environ. 2016;225:140–9.
Mas AH, Dietsch TV. Linking shade coffee certification to biodiversity conservation: butterflies and birds in Chiapas, Mexico. Ecol Appl. 2004;14:642–54.
Taft OW, Haig SM. Landscape context mediates influence of local food abundance on wetland use by wintering shorebirds in an agricultural valley. Biol Conserv. 2006;128:298–307.
Bruggisser OT, Schmidt-Entling MH, Bacher S. Effects of vineyard management on biodiversity at three trophic levels. Biol Conserv. 2010;143:1521–8.
Wretenberg J, Part T, Berg A. Changes in local species richness of farmland birds in relation to land-use changes and landscape structure. Biol Conserv. 2010;143:375–81.
Verhulst J, Báldi A, Kleijn D. Relationship between land-use intensity and species richness and abundance of birds in Hungary. Agric Ecosyst Environ. 2004;104:465–73.
Karp DS, Ziv G, Zook J, Ehrlich PR, Daily GC. Resilience and stability in bird guilds across tropical countryside. Proc Natl Acad Sci. 2011;108:21134–9.
Moorcroft D, Moorcroft D, Whittingham MJ, Whittingham MJ, Bradbury RB, Bradbury RB, et al. The selection of stubble fields by wintering granivorous passerine birds reflects vegetation cover and food abundance. J Appl Ecol. 2002;39:535–47.
Schaub M, Martinez N, Tagmann-Ioset A, Weisshaupt N, Maurer ML, Reichlin TS, et al. Patches of bare ground as a staple commodity for declining ground-foraging insectivorous farmland birds. PLoS One. 2010;5:e13115.
Kremen C, Iles A, Bacon C. Diversified farming systems : an agroecological, systems-based alternative to modern industrial agriculture. Ecol Soc. 2012;17:44.
M’Gonigle LK, Ponisio LC, Cutler K, Kremen C. Habitat restoration promotes pollinator persistence and colonization in intensively managed agriculture. Ecol Appl. 2015;25:1557–65.
Henderson IG, Ravenscroft N, Smith G, Holloway S. Effects of crop diversification and low pesticide inputs on bird populations on arable land. Agric Ecosyst Environ. 2009;129:149–56.
Heikinnen RK, Luoto M, Virkkala R, Rainio K. Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural – forest mosaic. J Appl Ecol. 2004;41:824–35.
De La Montaña E, Rey-Benayas JM, Carrascal LM. Response of bird communities to silvicultural thinning of Mediterranean maquis. J Appl Ecol. 2006;43:651–9.
Perfecto I, Vandermeer JH, Bautista GL, Nuñez GI, Greenberg R, Bichier P, et al. Greater predation in shaded coffee farms : the role of resident neotropical birds. Ecology. 2004;85:2677–81.
Mulwa RK, Böhning-Gaese K, Schleuning M. High bird species diversity in structurally heterogeneous farmland in western Kenya. Biotropica. 2012;44:801–9.
Haila Y, Nicholls AO, Hanski IK, Raivio S. Stochasticity in bird habitat selection : year-to-year changes in territory locations in boreal forest bird assemblage. Oikos. 1996;76:536–52.
Laiolo P. Spatial and seasonal patterns of bird communities in Italian agroecosystems. Conserv Biol. 2005;19:1547–56.
Luther D, Hilty J, Weiss J, Cornwall C, Wipf M, Ballard G. Assessing the impact of local habitat variables and landscape context on riparian birds in agricultural, urbanized, and native landscapes. Biodivers Conserv. 2008;17:1923–35.
Kelt DA, Engilis AJ, Monárdez J, Walsh R, Meserve PL, Gutiérrez JR. Seasonal and multiannual patterns in avian assemblage structure and composition in northern Chilean thorn-scrub. Condor. 2012;114:30–43.
Gutiérrez JR, Meserve PL, Kelt DA, Engilis AJ, Andrea Previtali M, Bryan Milstead W, et al. Long-term research in Bosque fray Jorge national park: twenty years studying the role of biotic and abiotic factors in a Chilean semiarid scrubland. Rev Chil Hist Nat. 2010;83:69–98.
Elsen PR, Kalyanaraman R, Ramesh K, Wilcove DS. The importance of agricultural lands for Himalayan birds in winter. Conserv Biol. 2017;31:416–26.
Guyot C, Arlettaz R, Korner P, Jacot A, Vickery J, Arlettaz R, et al. Temporal and spatial scales matter: circannual habitat selection by bird communities in vineyards. PLoS One. 2017;12:e0170176.
Cox RL, Underwood EC. The importance of conserving biodiversity outside of protected areas in mediterranean ecosystems. PLoS One. 2011;6:e14508.
Santibáñez F, Uribe J. Atlas agroclimático de Chile: regiones VI y VII. Santiago: Universidad de Chile, Fac. de Cs. Agrarias y Forestales; 1993.
La GR. Vegetación natural de Chile: clasificación y distribución geográfica. Santiago: Editorial Universitaria; 1994.
Ralph C, Droege S, Sauer J. Managing and monitoring birds using point counts: standards and applications. USDA For Serv Gen Tech Rep PSW-GTR-149. 1995. PSW-GTR-14:161–8.
Merenlender AM, Reed SE, Heise KL. Exurban development influences woodland bird composition. Landsc Urban Plan. 2009;92:255–63.
Estades CF. Bird-habitat relationships in a vegetational gradient in the Andes of central Chile. Condor. 1997;99:719–27.
Martinez D, Gonzalez G. Las Aves de Chile: Nueva guía de campo. Santiago: Ediciones del Naturalista; 2004.
Remsen JVJ, Areta JI, Cadena CD, Claramunt S, Jaramillo A, Pacheco JF, et al. A classification of the bird species of South America. American Ornithologists’ Union. 2016. http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm. Accessed 16 Dec 2016.
ESRI. ArcGis desktop 9.2 Environmental systems research institute. Redlands, California. 2006. http://www.esri.com/.
Bowen ME, McAlpine CA, House APN, Smith GC. Agricultural landscape modification increases the abundance of an important food resource: mistletoes, birds and brigalow. Biol Conserv. 2009;142:122–33.
Kang W, Minor ES, Park C, Lee D. Effects of habitat structure, human disturbance, and habitat connectivity on urban forest bird communities. Urban Ecosyst. 2015;18:857–70.
Zuur AF, Ieno EN, Walker N, Saveliev AA, Smith GM. Mixed effects models and extensions in ecology with R. Statistics. New York: Springer; 2009.
Liang J, Hua S, Zeng G, Yuan Y, Lai X, Li X, et al. Application of weight method based on canonical correspondence analysis for assessment of anatidae habitat suitability: a case study in east dongting lake, middle china. Ecol Eng. 2015;77:119–26.
Azpiroz AB, Blake JG. Associations of grassland birds with vegetation structure in the northern Campos of Uruguay. Condor. 2016;118:12–23.
Guisan A, Weiss SB, Weiss AD. GLM versus CCA spatial modeling of plant species distribution. Plant Ecol. 1999;143:107–22.
Burnham KKP, Anderson DRD. Model selection and multimodel inference. New York: Springer; 2004.
R Development Core Team. R: a language and environment for statistical computing. R Found Stat Comput. 2016;1(3.3):409.
Stockwell DR, Peterson AT. Effects of sample size on accuracy of species distribution models. Ecol Modell. 2002;148:1–13.
Buckingham DL, Peach WJ, Fox DS. Effects of agricultural management on the use of lowland grassland by foraging birds. Agric Ecosyst Environ. 2006;112:21–40.
Pelosi C, Bonthoux S, Castellarini F, Goulard M, Ladet S, Balent G. Is there an optimum scale for predicting bird species’ distribution in agricultural landscapes? J Environ Manage. 2014;136:54–61.
Berg Å, Wretenberg J, Żmihorski M, Hiron M, Pärt T. Linking occurrence and changes in local abundance of farmland bird species to landscape composition and land-use changes. Agric Ecosyst Environ. 2015;204:1–7.
Rodríguez-Estrella R. Land use changes affect distributional patterns of desert birds in the Baja California peninsula, Mexico. Divers Distrib. 2007;13:877–89.
Jackman S. pscl: Classes and Methods for R Developed in the Political Science Computational Laboratory, Stanford University. 2015. Department of Political Science, Stanford University. Stanford, California. R package version 1.4.9. http://pscl.stanford.edu/. Accessed 16 Jan 2016.
Bates D, Maechler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67:1–48.
Barton K. MuMIn: multi-model inference. 2016. http://cran.r-project.org/web/packages/MuMIn/index.html.
Brandolin PG, Blendinger PG. Effect of habitat and landscape structure on waterbird abundance in wetlands of central Argentina. Wetl Ecol Manag. 2016;24:93–105.
Oksanen J. Design decisions and implementation details in vegan. 2016;:1–11. https://cran.r-project.org/web/packages/vegan/vignettes/decision-vegan.pdf. Accessed 9 Jul 2016.
Graffelman J, Tuft R. Site scores and conditional biplots in canonical correspondence analysis. Environmetrics. 2004;15:67–80.
Palmer MW. Putting things in even better order: the advantages of canonical correspondence analysis. Ecology. 1993;74:2215–30.
Estades C, Vukasovic MA. Estado de conservación de las Aves de la región de O’Higgins. In: Serey I, Ricci M, Smith-Ramírez C, editors. Libro rojo de la región de O’Higgins. Rancagua: Corporación Nacional Forestal – Universidad de Chile; 2007. p. 53–8.
Cody ML. Chilean bird distribution. Ecology. 1970;51:455–64.
González-Acuña D, Riquelme-Salasar P, Cruzatt-Molina J, López-Sepúlveda P, Moreno-Salas L, Figueroa-Rojas R. Dieta de la codorniz californiana (callipepla californica) en áreas agrícolas del Centro sur de Chile. Rev Cient FCV-LUZ. 2013;23:312–7.
Figueroa R, Corales S. Seasonal diet of the aplomado falcon (falco femoralis) in an agricultural area of araucania, southern Chile. J Raptor Res. 2005;39:135–7.
Jaksic FM, Lazo I. Response of a bird assemblage in semiarid Chile to the 1997-1998 El Niño. Wilson Bull. 1999;111:527–35.
Lazo I, Anabalon JJ. Dinamica reproductiva de un conjunto de Aves Passeriformes de la sabana de espinos de Chile central. Ornitol Neotrop. 1992;3:57–64.
Pithon JA, Beaujouan V, Daniel H, Pain G, Vallet J. Are vineyards important habitats for birds at local or landscape scales? Basic Appl Ecol. 2016;17:240–51.
Martin EA, Viano M, Ratsimisetra L, Laloë F, Carrière SM. Maintenance of bird functional diversity in a traditional agroecosystem of Madagascar. Agric Ecosyst Environ. 2012;149:1–9.
Parish T, Lakhani KH, Sparks TH. Between bird population modelling the relationship and other field margin variables and hedgerow I. Species richness of winter, summer and attributes. Birds breeding. J Appl Ecol. 1994;31:764–75.
Hinsley SA, Bellamy PE. The influence of hedge structure, management and landscape context on the value of hedgerows to birds: a review. J Environ Manage. 2000;60:33–49.
Padoa-Schioppa E, Baietto M, Massa R, Bottoni L. Bird communities as bioindicators: the focal species concept in agricultural landscapes. Ecol Indic. 2006;6:83–93.
Sierro A, Arlettaz R, Naef-Daenzer B, Strebel S, Zbinden N. Habitat use and foraging ecology of the nightjar in the Swiss alps: towards a conservation scheme. Biol Conserv. 1994;2001(98):325–31.
Whittingham MJ, Bradbury RB, Wilson JD, Morris AJ, Perkins AJ, Siriwardena GM. Foraging patterns, nestling survival and territory distribution on lowland farmland. Bird Study. 2001;48:257–70.
Whittingham MJ, Krebs JR, Swetnam RD, Thewlis RM, Wilson JD, Freckleton RP. Habitat associations of British breeding farmland birds. Bird Study. 2009;56:43–52.
Hiron M, Berg Å, Eggers S, Josefsson J, Pärt T. Bird diversity relates to agri-environment schemes at local and landscape level in intensive farmland. Agric Ecosyst Environ. 2013;176:9–16.
Berg Å. Breeding birds in short-rotation coppices on farmland in central Sweden—the importance of Salix height and adjacent habitats. Agric Ecosyst Environ. 2002;90:265–76.
ODEPA. Agricultura chilena 2014:una perspectiva a mediano plazo. 2005; 242. http://www.odepa.cl/articulo/agricultura-chilena-2014una-perspectiva-de-mediano-plazo/. Accessed 16 Sept 2014.
Fahrig L, Girard J, Duro D, Pasher J, Smith A, Javorek S, et al. Farmlands with smaller crop fields have higher within-field biodiversity. Agric Ecosyst Environ. 2015;200:219–34.
Clavel J, Julliard R, Devictor V. Worldwide decline of specialist species: toward a global functional homogenization? Front Ecol Environ. 2011;9:222–8.
Karp DS, Rominger AJ, Zook J, Ranganathan J, Ehrlich PR, Daily GC. Intensive agriculture erodes β-diversity at large scales. Ecol Lett. 2012;15:963–70.
Castaño-Villa GJ, Ramos-Valencia SA, Fontúrbel FE. Fine-scale habitat structure complexity determines insectivorous bird diversity in a tropical forest. Acta Oecol. 2014;61:19–23.
Steffan-Dewenter I, Nzenberg UM, Rger CB, Thies C, Tscharntke T. Scale-dependent effects of landscape context on three pollinator guilds. Ecology. 2002;83:1421–32.
Azpiroz A, Blake J. Associations of grassland birds with vegetation structure in the northern Campos of Uruguay associations of grassland birds with vegetation structure in the northern Campos of Uruguay. Condor. 2016;118:12–23.
Orellana JI, Smith-Ramírez C, Rau JR, Sade S, Gantz A, Valdivia CE. Phenological synchrony between the austral thrush turdus falcklandii (Passeriformes : turdidae) and its food resources within forests and prairies in southern Chile. Rev Chil Hist Nat. 2014;87:1–8.
Gavier-Pizarro GI, Calamari NC, Thompson JJ, Canavelli SB, Solari LM, Decarre J, et al. Expansion and intensification of row crop agriculture in the pampas and espinal of Argentina can reduce ecosystem service provision by changing avian density. Agric Ecosyst Environ. 2012;154:44–55.
Barbaro L, Brockerhoff EG, Giffard B, van Halder I. Edge and area effects on avian assemblages and insectivory in fragmented native forests. Landsc Ecol. 2012;27:1451–63.
Somers CM, Morris RD. Birds and wine grapes: foraging activity causes small-scale damage patterns in single vineyards. J Appl Ecol. 2002;39:511–23.
Archuby D, Marti L, Montalti D, Soave G, Camperi A, Arambarri A, et al. Alimentación del cabecitanegra austral (carduelis barbata) Durante el otoño. Hornero. 2007;22:65–8.
Lopez-Calleja MV. Dieta de zonotrichia capensis (emberizidae) y diuca diuca (fringillidae): efecto de la variación estacional de los recursos tróficos y la riqueza de Aves granívoras en Chile central. Rev Chil Hist Nat. 1995;68:321–31. Wiens 1984.
Simeone A, Valencia JC, Schlatter R, Lanfranco D, Ide S. Depredación de Aves sobre larvas de rhyacionia buoliana (Schiff.) (Lepidoptera: tortricidae) en plantaciones jóvenes de pinus radiata D.Don en el sur de Chile. Bosque. 1997;18:67–75.
Fuentes ER, Hoffmann AJ, Poiani A, Alliende MC. patterns in the Chilean matorral. Oecologia. 1986;68:358–66.
Armesto JJ, Arroyo MTK, Hinojosa LF. The mediterranean environment of central Chile. In: Veblen TT, Young KR, Orme AR, editors. The physical geography os south America. New York: Oxford University Press; 2007. p. 184–99.
Henderson IG, Cooper J, Fuller RJ, Vickery J. The relative abundance of birds on set-aside and neighbouring fields in summer. J Appl Ecol. 2000;37:335–47.
Marshall EJP, Brown VK, Boatman ND, Lutman PJW, Squire GR, Ward LK. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003;43:77–89.
Cunningham HM, Chaney K, Bradbury RB, Wilcox A. Non-inversion tillage and farmland birds: a review with special reference to the UK and Europe. Ibis. 2004;146:192–202.
Salvande M, Figueroa JA, Armesto JJ. Quantity component of the effectiveness of seed dispersal by birds in the temperate rainforest of chiloé, Chile. Bosque (Valdivia). 2011;32:39–45.
ODEPA. Estadisticas productivas. 2016. http://www.odepa.gob.cl/estadisticas/productivas/. Accessed 1 Jan 2016.
Tuck SL, Winqvist C, Mota F, Ahnström J, Turnbull LA, Bengtsson J, et al. Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J Appl Ecol. 2014;51:746–55.
Tscharntke T, Milder JC, Schroth G, Clough Y, Declerck F, Waldron A, et al. Conserving biodiversity through certification of tropical agroforestry crops at local and landscape scales. Conserv Lett. 2015;8:14–23.
Kremen C, M’Gonigle LK. Small-scale restoration in intensive agricultural landscapes supports more specialized and less mobile pollinator species. J Appl Ecol. 2015;52:602–10.
Acknowledgements
We thank A. Merenlender, P. Elsen, and L. Renwick, and two anonymous reviewers for comments that greatly improved this manuscript.
Funding
Funds were provided by the Chilean Commission for Scientific and Technological Research (CONICYT) to A.M.S. through a scholarship (#22090103). None of the funders had any influence on the content of the submitted or published manuscript. None of the funders require approval of the final manuscript to be published.
Availability of data and materials
The dataset supporting the conclusions of this article is available in the Figshare repository, DOI 10.6084/m9.figshare.4980542, https://figshare.com/s/ac4406ac5153355d1c8c.
Authors’ contributions
A.M.-S., J.P.-Q. and C.F.E. conceived the idea, designed methods, and experiment. A.M.-S. collected the data and conducted the research, and analyzed the data. A.M.-S. wrote the first draft of the manuscript; and all authors contributed substantially to revisions.
Competing interests
The authors declare that they have no competing interests
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional file
Additional file 1: Table S1.
Correlation matrix between land covers at different scales. S.2. Results of the GLMMs by species and Adjusted R-squared. Model variable and coefficient estimates (± standard error). Significant values are represented by ***(p < 0.001), **(p < 0.01), *(p < 0.05). S.3. Bird species recorded in this study. Names according Remsen et al. [35]. (DOCX 30 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Muñoz-Sáez, A., Perez-Quezada, J.F. & Estades, C.F. Agricultural landscapes as habitat for birds in central Chile. Rev. Chil. de Hist. Nat. 90, 3 (2017). https://doi.org/10.1186/s40693-017-0067-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40693-017-0067-0