
Abstract
Background
There are many studies on disentangling the responses of autotrophic (AR) and heterotrophic (HR) respiration components of soil respiration (SR) to long-term drought, but few studies have focused on the mechanisms underlying its responses.
Methods
To explore the impact of prolonged drought on AR and HR, we conducted the 2-year measurements on soil CO2 effluxes in the 7th and 8th year of manipulated throughfall reduction (TFR) in a warm-temperate oak forest.
Results
Our results showed long-term TFR decreased HR, which was positively related to bacterial richness. More importantly, some bacterial taxa such as Novosphingobium and norank Acidimicrobiia, and fungal Leptobacillium were identified as major drivers of HR. In contrast, long-term TFR increased AR due to the increased fine root biomass and production. The increased AR accompanied by decreased HR appeared to counteract each other, and subsequently resulted in the unchanged SR under the TFR.
Conclusions
Our study shows that HR and AR respond in the opposite directions to long-term TFR. Soil microorganisms and fine roots account for the respective mechanisms underlying the divergent responses of HR and AR to long-term TFR. This highlights the contrasting responses of AR and HR to prolonged drought should be taken into account when predicting soil CO2 effluxes under future droughts.
Similar content being viewed by others


Background
Climate models predict widespread alterations in precipitation regimes, including longer, more intense droughts in the next decades (IPCC 2013). The increased drought has a considerable effect on terrestrial carbon (C) cycling (Tian et al. 2000; Batson et al. 2015; Vidon et al. 2016), particular to soil respiration (SR), which is the largest CO2 flux from terrestrial ecosystems back to the atmosphere (Janssens et al. 2001). Although multitudes of drought experiments have been conducted to explore the effect of drought on SR, the results of previous studies have been variable, including increase (Cleveland et al. 2010; Zhang et al. 2015), decrease (Schindlbacher et al. 2012; Selsted et al. 2012), and no change (Davidson et al. 2008; Lu et al. 2017). These inconsistent and often contradictory results constrain our understanding of feedbacks between soil C cycling and climate change.
Predicting the response of SR to drought is inherently difficult as SR is a combination of respiration associated with root activity (autotrophic respiration, AR) and soil organic matter (SOM) decomposition (heterotrophic respiration, HR) (Wang et al. 2014). Due to the difference in turnover times and control factors of plant and soil C pools, AR and HR often respond differently to drought (Borken et al. 2006; Huang et al. 2018; Sun et al. 2019). It has been shown that drought decreased SR by 19%, which was mainly ascribed to the reduced AR in a dry temperate forest (Hinko-Najera et al. 2015). Another study in a subtropical forest found that drought decreased both AR and HR, resulting in 17% reduction in SR (Zhou et al. 2020). However, a previous study in a warm-temperate oak forest suggested that drought increased SR by 26.7% at a small scale (e.g., 4 m × 4 m roof) throughfall reduction, which was mainly attributed to the increase in AR (Liu et al. 2016). All of these indicate roots and soil microbes have differential sensitivities to drought, and ultimately determine the direction of SR in response to drought in different ecosystems (Luo and Zhou 2006). Nevertheless, the mechanisms underlying different responses of AR and HR to drought are far from clear, which limits our comprehension of whether soil acts as a C sink or C source in the scenario of increased droughts.
Although these past studies provide important insights into the effects of drought on SR and its components, they have almost conducted for short-term drought periods, rather than long-term consecutive drought. The long-term experiments are crucial to revealing not only the transient responses of AR and HR to drought but also the adaptive response. A throughfall reduction experiment in a tropical rainforest found that drought increased SR during the first 3 years (Zhang et al. 2015), but did not change SR after 6 years of continuous drought because of the increased HR and decreased AR (Zhou et al. 2019). Thus, the drought responses of AR and HR may shift with the duration of drought, with consequences for SR (Metcalfe et al. 2007; da Costa et al. 2013).
To explore how prolonged drought affects SR and its components, we conducted 2-year field experiment with the consecutive manipulated throughfall reduction (TFR) in a warm-temperate oak forest. Our previous study reported that TFR barely affected SR and its components during the first 4 years (Lu et al. 2017). In the present study, we explored if TFR still had a slight effect on SR after 6-year consecutive TFR treatment, and if AR and HR responded differently to long-term TFR. Soil microbial community attributes (e.g., diversity, abundance) as well as fine root properties (e.g., biomass, production) were studied to reveal the mechanisms underlying the responses of AR and HR to long-term TFR.
Materials and methods
Study site
The research was conducted at the Baotianman Forest Ecosystem Research Station (111° 92′ E, 33° 49′ N), Henan Province, central China. The study area has a continental monsoon climate and has four distinctive seasons, with humid and hot summer, and dry and cold winter. The annual average air temperature is 15.1 °C, and the annual precipitation is 894 mm (1400 m a.s.l.) (Liu et al. 2016). The upland soil is dominated by Haplic luvisol and soil pH ranged from 4.4–5.1 (Luan et al. 2011; Lu et al. 2017). The soil has a sandy loam texture with 57%–62% sand, 11%–13% slit, and 27%–30% clay (Luan et al. 2011). The dominant deciduous broadleaf tree species include Quercus variabilis, Quercus aliena var. acuteserrata, and Fagus engleriana, and coniferous tree species include Pinus armandii, Pinus tabulaeformis, and Pinus massoniana.
Experimental design
In the spring of 2013, six plots (20 m × 20 m) were set up in a 60-year-old oak (Q. aliena var. acuteserrata) forest. Three plots with the ambient environment were designed as controls (“control”) and three plots were assigned to manipulated throughfall reduction (“TFR”). Detailed information for the TFR experiment refers to Lu et al. (2017). Briefly, about 160 shelter-panels (0.5 m × 3 m), covering 50% of the plot area, were installed in each TFR plot during the growing seasons (May–October) from 2013 to 2017. In the spring of 2018, we adjusted the magnitude of TFR from 50% to 70%. We inserted plastic barriers to a depth of 0.7 m around each TFR plot to inhibit the subsurface flow of water, and extended plastic flashing 5 cm above the ground to prevent overland flow. A buffer zone of 2.5 m width was set off along the inner edge of each plot and no measurements were made in the buffer zone. Litter that fell on the panels was collected weekly and distributed evenly throughout the plot to avoid variations in litter input on the ground.
Measurements of SR and its components
The HR was estimated using the trenching method as described by Lu et al. (2017). Briefly, five subplots (3 m × 3 m) were randomly assigned in each plot in March 2013 to measure HR. Trenches were dug about 1 m deep and placed plastic plates (5 mm thick) to inhibit root in-growth. In October of 2018, we dug the trenches again at the original position. To measure SR and HR, two PVC collars (19.6 cm inner diameter, 8 cm height) were installed 5 cm into the soil in each un-trenched and trenched subplot. We estimated AR as the difference between SR and HR.
The SR and HR were measured once a month during the growing seasons from 2019 to 2020 using a Li-8100 soil CO2 flux system (LI-COR Inc., Lincoln, NE, USA). In the meanwhile, soil temperature (ST) and soil moisture (SM) were manually measured by a portable temperature probe and soil moisture gauge (MPKit-BN, TZT Inc., Nantong, China) at three locations around each collar at 0–5 cm depth. Besides, in each plot, an EM50 data logger was installed with five 5TM soil temperature and moisture combined probes to continuously measure ST and SM at 30 min intervals. The precipitation data were obtained from the automatic weather station at Baotianman Forest Ecosystem Research Station, about 1 km away from the experimental plots.
Soil and fine root characteristics
Soil samples were collected in August 2019 and August 2020 within the same day of SR measurements to examine the effects of TFR on soil microbial communities and fine root characteristics. Two soil samples (0–10 cm) from each subplot were collected using soil augers (inner diameter 3.8 cm). The fresh soil samples sieved by 2-mm mesh were used for chemical and microbial analyses. Fine root biomass (< 2 mm) was oven-dried to a constant weight and then weighted. The soil organic carbon (SOC) and total nitrogen (N), fine root C and N content were determined using an elemental analyzer. The soil total phosphorus (P) and fine root P contents were measured using the alkaline fusion molybdenum-antimony colorimetry (Bao 2000). Fine root nonstructural carbohydrates (NSC): soluble sugar and starch were determined using the anthrone colorimetric method (Gao 2006). Soil microbial biomass C (MBC) and nitrogen (MBN) were analyzed by chloroform fumigation extraction method with conversion factors of 0.45 (Wu et al. 1990) and 0.54 (Vance et al. 1987), respectively. Soil β-glucosidase, polyphenolase, peroxidase, and amylase activities were assayed by the colorimetric method according to Guan (1986).
A modified ingrowth core method (Hertel and Leuschner 2002) was conducted to determine fine root production. Five stainless steel cubes (20 cm × 20 cm × 20 cm) with a 2-mm mesh were installed in each plot, and were refilled with rootless native soil in May 2019. The fine root in these in-growth blocks was collected at the end of October each year.
Soil bacterial and fungal community analyses
Soil microbial communities were assessed using amplicon-sequencing technology. In brief, microbial DNA from each soil sample was extracted using the E.Z.N.A.® soil DNA Kit (Omega Bio-tek, Norcross, GA, USA) following the manufacturer’s manual. The bacterial 16S rRNA and fungal ITS genes were amplified using the primers 515F/907R (Yusoff et al. 2013) and the primer ITS1F/ITS2R (Adams et al. 2013), respectively. The PCR and high-throughput sequencing were conducted by the Majorbio Company (Shanghai, China) using the Illumina MiSeq PE300 platform. Operational taxonomic units (OTUs) were classified at 97% similarity level using UPARSE (version 7.1), and chimeric sequences identified by UCHIME were discarded (Edgar 2013). The taxonomy of bacteria and fungi was assigned by RDP Classifier (Wang et al. 2007) against the Silva and Unite database, respectively, with a 70% confidence threshold. All samples were rarefied to the minimum sequence of the sample before the following analyses. We analyzed microbial composition and diversity on a platform (www.i-sanger.com) of Majorbio Company.
Data analysis
The statistical analyses were carried out using R and SPSS version 24.0 for Windows (SPSS, Chicago, Illinois, USA). We used linear mixed model to test the differences of SR, AR, HR, soil moisture and temperature between the control and TFR. TFR and month were set as fixed factors, and the plot was set as a random factor. In each year/month, we used one-way ANOVA to test the effects of TFR on SR, AR, HR, and microbial community attributes (e.g., diversity, abundance). We also used two-way ANOVA to test the effects of TFR and year on soil and root properties, microbial biomass, and enzymatic activities.
Regression modeling was used to investigate the relationships between SR, HR, or AR and ST, as well as SM. The temperature sensitivity (Q10 value) was estimated by the following function (Lloyd and Taylor 1994):
where, Q10 = e10β; R represents SR, HR, and AR; α and β are fitted parameters; T is the measured soil temperature.
Pearson correlation analyses were used to examine correlations between SR, HR, or AR and measured biotic and abiotic factors. Structure equation modeling (SEM) was conducted using AMOS 21.0 (SPSS Software, Chicago, Illinois, USA) to evaluate their relative importance of abiotic and biotic drivers in determining AR and HR (Wang et al. 2019). According to the results of Pearson correlation, the SEM only included dominant factors driving the changes in AR and HR. The SEM was fitted using the maximum likelihood estimation. The best model was selected using the lowest AIC value.
We used random forest analysis to identify the statistically significant bacterial and fungal predictors (genera) for HR (Trivedi et al. 2016). A total of 219 bacterial genera and 46 fungal genera were selected in the random forest modeling. After that, we used linear regressions to assess the relationships between the relative abundance of the selected predictive genera and HR.
Results
Soil physicochemical properties and soil enzymes
TFR decreased SM by 28% and 23% for un-trenched subplots and trenched subplots, respectively (P < 0.05), but did not change ST during the study periods (Fig. 1 and Table 1).

Effects of TFR on seasonal variability of soil temperature (ST; a and b) and soil moisture (SM; c and d) for the un-trenched and trenched subplots from 2019 to 2020. Error bars represent the mean standard error
TFR had no significant effects on MBC and MBN (Table S1), and had little effects on measured enzyme activities except for polyphenolase (P = 0.08) (Table S1).
Soil respiration and its components
SR showed no significant difference between TFR and control in 2019 (2.48 ± 0.23 vs. 2.63 ± 0.19 μmol CO2∙m− 2∙s− 1) and in 2020 (2.92 ± 0.27 vs. 2.63 ± 0.24 μmol CO2∙m− 2∙s− 1) (P > 0.05; Fig. 2 and Table 1). TFR increased AR by 92% during the study periods (P = 0.056; Table 1), with respective interannual variation of 96% in 2019 and 88% in 2020 (P < 0.05; Fig. 2). TFR decreased HR by 23% during the study periods (P = 0.076; Table 1), 29% in 2019 (P < 0.05), and a slight decrease of 17% in 2020 (P = 0.09) (Fig. 2). TFR decreased relative contribution of HR to SR from 79% to 60% during the study periods (P < 0.05; Fig. 3).

Seasonal variation and average value of SR (a, d), HR (b, e) and AR (c, f) from 2019 to 2020. * indicates significant differences between the control and TFR at specific time (month or year) at 0.05 level

Seasonal variability in the relative contribution of AR and HR to SR under control (a) and TFR treatment (b)
Linking abiotic and biotic factors to soil CO2 efflux
SR, HR, and AR were significantly related to soil temperature under both treatments (Fig. 4). TFR did not change the temperature sensitivity (Q10 value) of SR, while decreased the Q10 value of HR from 2.69 to 2.10 and increased the Q10 value of AR from 3.42 to 3.97 (Fig. 4). Soil moisture was significantly correlated with SR and HR (P < 0.05) although at a lower coefficient of determination, but not AR (Fig. 4).

Relationships between soil temperature and SR (a), HR (b), and AR (c), and relationships between soil moisture and SR (d), HR (e) and AR (f) under control and TFR treatment. *P < 0.05, **P < 0.01, ***P < 0.001
TFR increased fine root production during the study periods (P < 0.05; Fig. 5), and was positively correlated with AR (P < 0.05) (Fig. 5). The SEM indicated that AR was directly controlled by fine root biomass, when considering other key soil properties (Fig. 7).

Fine root production (a) under the control and TFR treatment, and relationships between fine root production and the average of AR (b). *P < 0.05
TFR significantly decreased bacterial richness (P < 0.05), and was marginally correlated with HR (P = 0.07) (Fig. 6 and Table S2). The SEM also suggested that HR was directly driven by soil bacterial richness, when considering other key soil properties (Fig. 7). Our random forests modeling showed that many bacterial genera predictors of HR were distributed among the Proteobacteria, Acidobacteriota, Chloroflexi, and Actinobacteriota; fungal genera predictors of HR belong to the Ascomycota and Mucoromycota (Fig. S2). Some of these predictive genera were significantly related (linear regressions) to HR (Fig. 8). For example, TFR decreased the relative abundance of bacterial Novosphingobium, norank 11–24 and norank Vicinamibacterales (P < 0.05) (Table S4), and were positively related to HR (Fig. 8). TFR increased the abundance of bacterial 1959–1 and norank Acidimicrobiia (P < 0.05) (Table S4), and were negatively associated with HR (Fig. 8). In addition, TFR increased the relative abundance of fungal Leptobacillium (P < 0.05; Table S4), and was negatively related to HR (Fig. 8).

Effects of TFR on bacterial and fungal richness (Sobs index) (a), and relationships between bacterial richness and AR (b). *P < 0.05

Structure equation modeling (SEM) assessing the direct and indirect influences of driving factors on AR and HR. Blue and red arrows indicate negative and positive relationships, respectively. The numbers on arrows are standardized path coefficients, and arrow width is proportional to the strength of the relationship. *P < 0.05, **P < 0.01, ***P < 0.001

Linear regressions between the relative abundances of selected microbial genera and HR across different treatment
Discussion
Effects of TFR on HR, AR and SR
Consistent with those of other studies (Borken et al. 2006; Risk et al. 2012), we also found long-term TFR decreased HR. TFR significantly decreased bacterial richness, which may lead to the decreased HR due to the positive correlation between bacterial diversity and soil CO2 efflux (Delgado-Baquerizo et al. 2016; Liu et al. 2018). Generally, greater microbial species richness can allow for more metabolic activities, promoting the decomposition of organic matter (Naylor and Coleman-Derr 2018). Thus, the decreased HR after 6-year consecutive TFR could arise from the decreased bacterial diversity (Singh et al. 2010; Hutchins et al. 2019). Another study in this oak forest has shown that short-term variation in soil moisture had no significant impact on bacterial diversity (Wei et al. 2018), which may explain the unchanged HR under TFR treatment during the first 4 years (Lu et al. 2017). Since most of the studies on microbial response to drought are short term, long-term drought experiments are needed to further understand the mechanisms underlying microbial drought response over time.
We subsequently identified major microbial taxa (genera) that predicted the changes in HR. We found the relative abundance of Novosphingobium, norank 11–24 and norank Vicinamibacterales belong to Gram-negative bacteria were positively associated with HR. In addition, the relative abundance of 1959–1 and norank Acidimicrobiia belong to Gram-positive bacteria were negatively related to HR. Studies have shown that soils with a high abundance of oligotrophs or low abundance of copiotrophs may have low CO2 emissions (Trivedi et al. 2013; Liu et al. 2018). It has also been shown that the Gram-negative bacteria have characteristics of copiotrophs, while Gram-positive bacteria contain characteristics of oligotrophs (Naylor and Coleman-Derr 2018). Thus, the TFR-induced changes in the relative abundance of these bacterial taxa were responsible for the decreased HR. In addition, we found the relative abundance of fungal Leptobacillium belong to Ascomycota were negatively correlated to HR, despite a weak correlation between overall fungal community diversity with HR. We know little about the mechanism of the association between Leptobacillium and C emission, and postulated the taxa are tolerant to drought and may have high C use efficiency (Liu et al. 2018). Our results suggest that there are phylotypes that can be used to consistently predict HR under prolonged drought conditions.
Previous studies have suggested that experimental drought often reduced AR due to the decreased fine root biomass or belowground C allocation among different ecosystems (Hinko-Najera et al. 2015; Huang et al. 2018; Zhou et al. 2019). However, we found long-term TFR increased AR, mainly because of higher fine root biomass and production under the TFR. According to the optimal partitioning theory (Bloom et al. 1985), plants should allocate more C to root growth from aboveground parts to reduce water limitation (Fuchslueger et al. 2014). However, many field throughfall exclusion experiments of forests have shown that fine root biomass did not always support this theory, including increase (Zhou et al. 2020), decrease (Moser et al. 2014), or little change during the first 4 years in this study (Lu et al. 2017), indicating that the responses of fine root to water deficit depends on intensity and duration of drought. In our system, we argued that mature trees increased belowground C allocation to adapt to the long-term drought, resulting in the higher AR.
Contrary to other studies (Sotta et al. 2007; Cleveland et al. 2010), we found long-term TFR had no significant effect on SR. Another study also showed that drought had no significant impact on soil CO2 flux but did not mention AR and HR in a tropical forest (Davidson et al. 2008). This may mask the contrasting responses of AR and HR to long-term drought due to the different sensitivities of fine roots and microbes to water deficit (Zhou et al. 2019). Here, the present result showed AR and HR had opposite responses after 6-year consecutive TFR, leading to the unchanged SR.
Seasonal variability of soil CO2 efflux
The pronounced seasonal variations of SR, AR, and HR were explained by soil temperature, which was in agreement with previous studies (Vincent et al. 2006; Liu et al. 2016). Besides, soil moisture can also partly explained the seasonal patterns of SR and HR. These indicated that soil temperature and soil moisture can solely control soil respiration through influencing decomposition rates and microbial activity (Barthel et al. 2011). However, we found a weak relationship between soil moisture and AR along with the seasonal changes. This did not mean that soil moisture was not important to roots, but it may be the inherent growth rhythm of roots and the utilization of deep soil water that masked the effect of soil moisture on AR. TFR increased the Q10 of AR while decreased it of HR. The changed Q10 value may reflect the shifts in the physiological status of plant roots and soil microbes (Zhang et al. 2014), which was potentially important for C-climate feedback models, and needs to be further evaluated.
Specifically, we found HR showed little difference between control and TFR in May and October 2020 (Fig. 2). A similar pattern was also found in AR in both 2019 and 2020 (Fig. 2). This may be attributed to the lower activities of both roots and soil microorganisms in May and October, and hence lower water requirement (Chapin et al. 2002). It is worth noting that the maximum AR in July 2019 and 2020 can determine the response of SR to drought, and thus the monitoring frequency of SR should be increased to accurately assess the response of SR to drought.
Relative contribution of AR and HR to SR
Although partitioning AR and HR of SR and evaluating the responses of AR and HR to drought is vital to understanding whether and how drought facilitates soil C sequestration, there was limited research on this crucial issue in the warm-temperate forests of China (Luan et al. 2012; Liu et al. 2016; Lu et al. 2017). We used the trenching method to distinguish AR and HR, which was widely applied in forest ecosystems (Liu et al. 2016; Huang et al. 2018). This method may underestimate HR due to the elimination of roots and associated root exudation, which are respired by soil microbes and often lead to a priming effect resulting from SOC decomposition (Hanson et al. 2000). It may also overestimate HR due to the elevated soil moisture in the trenched subplots (Yan et al. 2010). Moreover, the AR in our study may be overestimated as the estimation of AR includes both root respiration and rhizosphere respiration (e.g., mycorrhizal respiration) (Hopkins et al. 2013).
Nevertheless, our study provides empirical evidence that long-term TFR decreased the relative contribution of HR to SR, indicating that the proportion of CO2 released from microbial SOM decomposition was lower than that from root activities under TFR treatment. As is known to all, HR is an important indicator of SOC decomposition and also plays a crucial role in the stability of SOC (Janssens et al. 2010). Therefore, the decreased relative contribution of HR to SR may lead to more soil C sink (Wang et al. 2019). On the other hand, soil C storage is also largely dependent on the C input (Riggs et al. 2015), thus the higher fine root biomass and production under TFR treatment may also facilitate soil C sequestration. However, recent studies have demonstrated that plant roots or belowground C allocation can drive the soil SOM decomposition (Moore et al. 2020; Street et al. 2020), indicating that the fine roots had a dual role in regulating soil C storage (Dijkstra et al. 2020). Our results suggest that how prolonged drought will ultimately influence SR, and therefore soil C storage, will depend not only on soil microorganisms but also on plant belowground C allocation.
Conclusions
In this warm-temperate oak forest, long-term TFR decreased HR and was positively associated with bacterial richness. More importantly, some bacterial taxa such as Novosphingobium and norank Acidimicrobiia, and fungal Leptobacillium were identified as the key drivers of HR. However, TFR significantly increased AR, which was attributed to the increased fine root biomass and production. The increase in AR offset the decrease in HR, resulting in unaltered SR under the TFR treatment. Our findings highlight the different response mechanisms of AR and HR to prolonged drought should be considered when predicting soil CO2 emissions under future droughts.
Availability of data and materials
The datasets used during the current study are available from the corresponding author on reasonable request.
Abbreviations
- SR:
-
Soil respiration
- AR:
-
Autotrophic respiration
- HR:
-
Heterotrophic respiration
- TFR:
-
Throughfall reduction
- SOM:
-
Soil organic matter
- SM:
-
Soil moisture
- ST:
-
Soil temperature
- SOC:
-
Soil organic carbon
- C:
-
Carbon
- N:
-
Nitrogen
- P:
-
Phosphorus
- NSC:
-
Nonstructural carbohydrates
- MBC:
-
Microbial biomass C
- MBN:
-
Microbial biomass nitrogen
- SEM:
-
Structure equation modeling
References
Adams RI, Miletto M, Taylor JW, Bruns TD (2013) Dispersal in microbes: fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J 7:1262–1273
Bao SD (2000) Soil agricultural chemistry analysis, 3rd edn. China Agriculture Press, Beijing
Barthel M, Hammerle A, Sturm P, Baur T, Gentsch L, Knohl A (2011) The diel imprint of leaf metabolism on the δ13 C signal of soil respiration under control and drought conditions. New Phytol 192:925–938
Batson J, Noe GB, Hupp CR, Krauss KW, Rybicki NB, Schenk ER (2015) Soil greenhouse gas emissions and carbon budgeting in a short-hydroperiod floodplain wetland. J Geophys Res-Biogeo 120:77–95
Bloom AJ, Chapin FSIII, Mooney HA (1985) Resource limitation in plants-an economic analogy. Annu Rev Ecol Syst 16:363–392
Borken W, Savage K, Davidson EA, Trumbore SE (2006) Effects of experimental drought on soil respiration and radio carbon efflux from a temperate forest soil. Glob Chang Biol 12:177–193
Chapin FS, Matson PA, Mooney HA (2002) Principles of terrestrial ecosystem ecology. Springer, New York
Cleveland CC, Wieder WR, Reed SC, Townsend AR (2010) Experimental drought in a tropical rain forest increases soil carbon dioxide losses to the atmosphere. Ecology 91:2313–2323
da Costa ACL, Metcalfe DB, Doughty CE, de Oliveira AAR, Neto GFC, da Costa MC, Silva JA Jr, Aragão LEOC, Almeida S, Galbraith DR, Rowland LM, Meir P, Malhi Y (2013) Ecosystem respiration and net primary productivity after 8–10 years of experimental through-fall reduction in an eastern Amazon forest. Plant Ecol Divers 7:7–24
Davidson EA, Nepstad DC, Ishida FY, Brando PM (2008) Effects of an experimental drought and recovery on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest. Glob Chang Biol 14:2582–2590
Delgado-Baquerizo M, Giaramida L, Reich PB, Khachane AN, Hamonts K, Edwards C, Lawton LA, Singh BK (2016) Lack of functional redundancy in the relationship between microbial diversity and ecosystem functioning. J Ecol 104:936–946
Dijkstra FA, Zhu B, Cheng WX (2020) Root effects on soil organic carbon: a double-edged sword. New Phytol. https://doi.org/10.1111/nph.17082
Edgar RC (2013) UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods 10:996–998
Fuchslueger L, Bahn M, Fritz K, Hasibeder R, Richter A (2014) Experimental drought reduces the transfer of recently fixed plant carbon to soil microbes and alters the bacterial community composition in a mountain meadow. New Phytol 201:916–927
Gao JF (2006) Instruction for plant physiology experiments. Higher Education Press, Beijing
Guan SY (1986) Soil enzyme and its research methods. Agricultural Press, Beijing
Hanson PJ, Edwards NT, Garten CT, Andrews JA (2000) Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry 48:115–146
Hertel D, Leuschner C (2002) A comparison of four different fine root production estimates with ecosystem carbon balance data in a Fagus–Quercus mixed forest. Plant Soil 239:237–251
Hinko-Najera N, Fest B, Livesley SJ, Arndt S (2015) Reduced throughfall decreases autotrophic respiration: but not heterotrophic respiration in a dry temperate broadleaved evergreen forest. Agric For Meteorol 200:66–77
Hopkins F, Gonzalez-Meler M, Flower C, Lynch D, Czimczik C, Tang J, Subke J (2013) Ecosystem-level controls on root-rhizosphere respiration. New Phytol 199:339–351
Huang S, Ye G, Lin J, Chen K, Xu X, Ruan H, Tan F, Chen HYH (2018) Autotrophic and heterotrophic soil respiration responds asymmetrically to drought in a subtropical forest in the Southeast China. Soil Biol Biochem 123:242–249
Hutchins DA, Jansson JK, Remais JV, Rich VI, Singh BK, Trivedi P (2019) Climate change microbiology – problems and perspectives. Nat Rev Microbiol 17:391–396
IPCC (2013) Working group I contribution to the IPCC fifth assessment report. Climate change 2013: the physical science basis summary for policymakers, Cambridge, UK/New York, NY, USA.
Janssens IA, Dieleman W, Luyssaert S, Subke JA, Reichstein M, Ceulemans R, Ciais P, Dolman AJ, Grace J, Matteucci G, Papale D, Piao SL, Schulze ED, Tang J, Law BE (2010) Reduction of forest soil respiration in response to nitrogen deposition. Nat Geosci 3:315–322
Janssens IA, Lankreijer H, Matteucci G, Kowalski AS, Buchmann N, Epron D, Pilegaard K, Kutsch W, Longdoz B, Grünwald T, Montagnani L, Dore S, Rebmann C, Moors EJ, Grelle A, RannikÜ MK, Oltchev S, Clement R, Guðmundsson J, Minerbi S, Berbigier P, Ibrom A, Moncrieff J, Aubinet M, Bernhofer C, Jensen NO, Vesala T, Granier A, Schulze E-D, Lindroth A, Dolman AJ, Jarvis PG, Ceulemans R, Valentini R (2001) Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Glob Chang Biol 7(3):269–278
Liu YC, Liu SR, Wan SQ, Wang JX, Luan JW, Wang H (2016) Differential responses of soil respiration to soil warming and experimental throughfall reduction in a transitional oak forest in Central China. Agric For Meteorol 226–227:186–198
Liu YR, Delgado-Baquerizo M, Wang JT, Hu HW, Yang Z, He JZ (2018) New insights into the role of microbial community composition in driving soil respiration rates. Soil Biol Biochem 118:35–41
Lloyd J, Taylor JA (1994) On the temperature dependence of soil respiration. Funct Ecol 8:315–323
Lu HB, Liu SR, Wang H, Luan JW, Schindlbacher A, Liu YC, Wang Y (2017) Experimental throughfall reduction barely affects soil carbon dynamics in a warm-temperate oak forest, Central China. Sci Rep 7:15099
Luan JW, Liu SR, Wang JX, Zhu XL, Shi ZM (2011) Rhizospheric and heterotrophic respiration of a warm-temperate oak chronosequence in China. Soil Biol Biochem 43:503–512
Luan JW, Liu SR, Zhu XL, Wang JX, Liu K (2012) Roles of biotic and abiotic variables in determining spatial variation of soil respiration in secondary oak and planted pine forests. Soil Biol Biochem 44:143–150
Luo Y, Zhou X (2006) Soil respiration and the environment. Academic, Elsevier
Metcalfe DB, Meir P, Aragão LEOC, Malhi Y, da Costa ACL, Braga A, Gonçalves PHL, de Athaydes J, de Almeida SS, Williams M (2007) Factors controlling spatio-temporal variation in carbon dioxide efflux from surface litter, roots, and soil organic matter at four rain forest sites in the eastern Amazon. J Geophys Res-Biogeo 112:G04001
Moore JAM, Sulman BN, Mayes MA, Patterson CM, Classen AT (2020) Plant roots stimulate the decomposition of complex, but not simple, soil carbon. Funct Ecol 34:899–910
Moser G, Schuldt B, Hertel D, Horna V, Coners H, Barus H, Leuschner C (2014) Replicated throughfall exclusion experiment in an Indonesian perhumid rainforest: wood production, litter fall and fine root growth under simulated drought. Glob Chang Biol 20:1481–1497
Naylor D, Coleman-Derr D (2018) Drought stress and root-associated bacterial communities. Front Plant Sci 8:2223
Riggs C, Hobbie S, Bach E, Hofmockel K, Kazanski C (2015) Nitrogen addition changes grassland soil organic matter decomposition. Biogeochemistry 125:203–219
Risk D, Nickerson N, Phillips C, Kellman L, Moroni M (2012) Drought alters respired δ13 CO2 from autotrophic: but not heterotrophic soil respiration. Soil Biol Biochem 50:26–32
Schindlbacher A, Wunderlich S, Borken W, Kitzler B, Zechmeister-Boltenstern S, Jandl R (2012) Soil respiration under climate change: prolonged summer drought offsets soil warming effects. Glob Chang Biol 18:2270–2279
Selsted MB, van der Linden L, Ibrom A, Michelsen A, Larsen KS, Pedersen JK, Mikkelsen TN, Pilegaard K, Beier C, Ambus P (2012) Soil respiration is stimulated by elevated CO2 and reduced by summer drought: three years of measurements in a multifactor ecosystem manipulation experiment in a temperate heathland (CLIMAITE). Glob Chang Biol 18:1216–1230
Singh BK, Bardgett RD, Smith P, Reay DS (2010) Microorganisms and climate change: terrestrial feedbacks and mitigation options. Nat Rev Microbiol 8:779–790
Sotta ED, Veldkamp E, Schwendenmann L, Guimaraes BR, Paixao RK, Ruivo M, da Costa L, Carlos A, Meir P (2007) Effects of an induced drought on soil carbon dioxide (CO2) efflux and soil CO2 production in an eastern Amazonian rainforest, Brazil. Glob Chang Biol 13:2218–2229
Street LE, Garnett MH, Subke JA, Baxter R, Dean JF, Wookey PA (2020) Plant carbon allocation drives turnover of old soil organic matter in permafrost tundra soils. Glob Chang Biol 26:4559–4571
Sun SQ, Lei HQ, Chang SX (2019) Drought differentially affects autotrophic and heterotrophic soil respiration rates and their temperature sensitivity. Biol Fertil Soils 55:275–283
Tian H, Melillo JM, Kicklighter DW, McGuire AD, Helfrich J, Moore B, Vorosmarty CJ (2000) Climatic and biotic controls on annual carbon storage in Amazonian ecosystems. Glob Ecol Biogeogr 9:315–335
Trivedi P, Anderson IC, Singh BK (2013) Microbial modulators of soil carbon storage: integrating genomic and metabolic knowledge for global prediction. Trends Microbiol 21:641–651
Trivedi P, Delgado-Baquerizo M, Trivedi C, Hu HW, Anderson IC, Jeffries TC, Zhou J, Singh B (2016) Microbial regulation of the soil carbon cycle: evidence from gene-enzyme relationships. ISME J 10:2593–2604
Vance E, Brookes P, Jenkinson D (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707
Vidon P, Marchese S, Welsh M, McMillan S (2016) Impact of precipitation intensity and riparian geomorphic characteristics on greenhouse gas emissions at the soil-at-mosphere interface in a water-limited riparian zone. Water Air Soil Pollut 227:1–12
Vincent G, Shahriari AR, Lucot E, Badot PM, Epron D (2006) Spatial and seasonal variations in soil respiration in a temperate deciduous forest with fluctuating water table. Soil Biol Biochem 38:2527–2535
Wang JS, Song B, Ma FF, Tian DS, Li Y, Yan T, Quan Q, Zhang FY, Li ZL, Wang BX, Gao Q, Chen WN, Niu SL (2019) Nitrogen addition reduces soil respiration but increases the relative contribution of heterotrophic component in an alpine meadow. Funct Ecol 33:2239–2253
Wang Q, Garrity GM, Tiedje JM, Cole JR (2007) Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol 73:5261–5267
Wang X, Liu LL, Piao SL, Janssens IA, Tang JW, Liu WX, Chi Y, Wang J, Xu S (2014) Soil respiration under climate warming: differential response of heterotrophic and autotrophic respiration. Glob Chang Biol 20:3229–3237
Wei H, Peng CH, Yang B, Song HX, Li Q, Jiang L, Wei G, Wang KF, Wang H, Liu XJ, Chen DX, Li YD, Wang M (2018) Contrasting soil bacterial community, diversity, and function in two forests in China. Front Microbiol 9:1693
Wu J, Joergensen R, Pommerening B, Chaussod R, Brookes P (1990) Measurement of soil microbial biomass C by fumigation-extraction - an automated procedure. Soil Biol Biochem 22:1167–1169
Yan LM, Chen SP, Huang JH, Lin GH (2010) Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe. Glob Chang Biol 16:2345–2357
Yusoff MZM, Hu A, Feng C, Maeda T, Shirai Y, Hassan MA, Yu CP (2013) Influence of pretreated activated sludge for electricity generation in microbial fuel cell application. Bioresour Technol 145:90–96
Zhang C, Niu D, Hall SJ, Wen H, Li X, Fu H, Wan C, Elser JJ (2014) Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biol Biochem 75:113–123
Zhang X, Zhang Y, Sha L, Wu C, Tan Z, Song Q, Liu Y, Dong L (2015) Effects of continuous drought stress on soil respiration in a tropical rainforest in Southwest China. Plant Soil 394:343–353
Zhou GY, Zhou XH, Liu RQ, Du ZG, Zhou LY, Li SS, Liu HY, Shao JJ, Wang JW, Nie YY, Gao J, Wang MH, Zhang MY, Wang XH, Bai SH (2020) Soil fungi and fine root biomass mediate drought-induced reductions in soil respiration. Funct Ecol 34:2634–2643
Zhou L, Liu Y, Zhang Y, Sha L, Song Q, Zhou W, Li J (2019) Soil respiration after six years of continuous drought stress in the tropical rainforest in Southwest China. Soil Biol Biochem 138:1–11
Acknowledgements
We would like to thank the Baotianman Forest Ecosystem Research Station for experimental maintenance and Xiaodong Niu and Xiang Li for assistance in fieldwork.
Funding
This study was supported by the National Key R&D Program of China (No. 2018YFC0507301) and by Research and Development Project of RIFEEP, Chinese Academy of Forestry (99802–2020).
Author information
Authors and Affiliations
Contributions
Conceived and design the experiment: Shirong Liu, Jinglei Zhang. Obtained data and samples in the field: Jinglei Zhang, Cuiju Liu, Xiaojing Liu. Processed samples in the lab: Jinglei Zhang, Xingwei Guo, Baoliang Niu. Analyzed the data and wrote the manuscript: Jinglei Zhang, Shirong Liu, Junwei Luan, Hui Wang, Cuiju Liu. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Supplementary Information
Additional file 1:
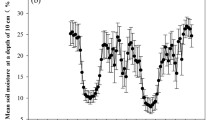
Fig. S1. Seasonal variation of daily (black bars) and monthly (grey bars) precipitation under ambient environment (a); daily averages of automatically measured (30 min interval) volumetric soil moisture content (b) and daily averages of automatically measured (30 min interval) soil temperature (c) to a depth of 40 cm under Control and throughfall reduction (TFR) conditions. Table S1. Results (F and P values) of two-way ANOVA on the effects of throughfall reduction (TFR), year (Y) and their interactions (TFR × Y) on soil and fine root properties. Table S2. Pearson correlation (R) between HR and measured variables. ST: soil temperature, SM: soil moisture, BR: bacterial richness, FR: fungal richness, MBC: microbial biomass carbon, MBN: microbial biomass nitrogen, BG: β-glucosidase, POP: polyphenolase, POD: peroxidase, AL: amylase. Table S3. Pearson correlation (R) between AR and measured variables. ST: soil temperature, SM: soil moisture, FRB: fine root biomass, NSC: non-structural carbohydrate, RC: root carbon content, RN: root nitrogen content, RP: root phosphorus content. Table S4. Relative abundance of microbial genera that showed a significant response (P < 0.05) to TFR. Fig. S2. Random Forest regression model shows the main microbial drivers of HR. MSE, is the mean square error. *P < 0.05, **P < 0.01.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, J., Liu, S., Liu, C. et al. Different mechanisms underlying divergent responses of autotrophic and heterotrophic respiration to long-term throughfall reduction in a warm-temperate oak forest. For. Ecosyst. 8, 41 (2021). https://doi.org/10.1186/s40663-021-00321-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40663-021-00321-z