
Abstract
Background
Fuchs endothelial corneal dystrophy is a hereditary disease and the most frequent cause of corneal transplantation in the worldwide. Its main clinical signs are an accelerated decrease in the number of endothelial cells, thickening of Descemet’s membrane and formation of guttae in the extracellular matrix. The cornea’s ability to maintain stromal dehydration is impaired, causing painful epithelial bullae and loss of vision at the point when the amount of corneal endothelial cells cannot be compensated. At present, apart from corneal transplantation, there is no other effective treatment that prevents blindness.
Main text
In this review, we first summarized the mutations of COL8A2, TCF4, TCF8, SLC4A11 and AGBL1 genes in Fuchs endothelial corneal dystrophy. The molecular mechanisms associated with Fuchs endothelial corneal dystrophy, such as endoplasmic reticulum stress and unfolded protein response pathway, oxidative stress, mitochondrial dysregulation pathway, apoptosis pathway, mitophagy, epithelial-mesenchymal transition pathway, RNA toxicity and repeat-associated non-ATG translation, and other pathogenesis, were then explored. Finally, we discussed several potential treatments related to the pathogenesis of Fuchs endothelial corneal dystrophy, which may be the focus of future research.
Conclusions
The pathogenesis of Fuchs endothelial corneal dystrophy is very complicated. Currently, corneal transplantation is an important method in the treatment of Fuchs endothelial corneal dystrophy. It is necessary to continuously explore the pathogenesis of Fuchs endothelial corneal dystrophy and establish the scientific foundations for the development of next-generation corneal therapeutics.
Similar content being viewed by others

Background
Fuchs endothelial corneal dystrophy (FECD) is a genetically heterogenous disease accompanied by irreparable damage to the corneal endothelium [1]. FECD can occur via autosomal dominant inheritance, but it is usually a sporadic disease [2, 3]. In 2016, FECD accounted for 36% of corneal transplantation in United States [4]. FECD is characterized by a thickening of Descemet’s membrane (DM) and the appearance of guttae [5]. The accelerated loss of corneal endothelial cells (CECs) first appears in the center of the cornea, and the same clinical signs appear on the periphery of the cornea. The mosaic defect of the corneal endothelium due to cell loss causes living cells to respond by proliferating and migrating, resulting in abnormalities in uniform size (polymegathism) and variations in hexagonal shape (pleomorphism) [6, 7].
The most recent International Classification of Corneal Dystrophies categorizes FECD into two types: 1) rare early-onset FECD, and 2) more common late-onset FECD. Early-onset FECD, ascribed to mutations in the collagen type VIII alpha 2 chain (COL8A2, MIM 12052) [8], usually begins in the first decade of life. On average, late-onset FECD manifests in the fifth decade of life and implicates rare mutations in solute carrier family 4 sodium borate transporter member 11 (SLC4A11, MIM 610206) [9, 10], transcription factor 8 gene (TCF8, MIM 189909) [11], transcription factor 4 gene (TCF4, MIM 602272) [12], lipoxygenase homology domains 1 (LOXHD1, MIM 613072) [13] and ATP/GTP binding protein like 1 (AGBL1, MIM 615496) [4].
The emerging clinical technologies prompted us to learn more about the clinical manifestations of FECD. As the disease develops, the epithelium does not change significantly during the early stage to the painful epithelial bullae caused by damage to the corneal pump function [1]. Confocal microscopy reveals that the Bowman layer is bright and reflective, with increased reflectivity in the basal epithelial and anterior stromal layers in FECD patients [14]. The collagen fibers in the posterior stroma with fewer fibrous connections are looser than in the anterior stroma. This corneal edema leads to an increase in the posterior corneal hydration, causing the posterior stroma to swell to the anterior chamber and the central cornea to thicken [15]. In normal corneal tissue, DM comprises two layers and is secreted by CECs. In adults, an anterior banded layer has a constant thickness of 3 μm, and a posterior non-banded layer has an approximate thickness of 10 μm throughout its lifespan [16]. Guttae, the focal nodules in DM, are deposited in the center of the cornea, spreading out from this point in FECD. As the density of endothelial cells decreases, the shape and size of the endothelial cell changes. Overall, having a better understanding of FECD’s genetic mutations and molecular mechanisms provides us with unique insights into FECD development and potential treatment options. The purpose of this article is to review gene mutations, molecular mechanisms and future therapies of FECD, which form the basis and rationale for the proposed management for FECD.
Main text
FECD genetic mutations
COL8A2 gene mutations in FECD
The COL8A2 gene is found on chromosome 1; it encodes the α2 chain of short-chain collagen VIII, which is an extracellular matrix (ECM) protein and constitutes a major component of DM [8]. The COL8A2 gene is closely related to early-onset FECD. Here, the substitution of glutamine for lysine is caused by a point mutation (p.Q455K) that was traced to an English family lineage [17]. Subsequently, a study of FECD patients showed a leucine-to-tryptophan substitution (p.L450W) in COL8A2 [8]. A study that genotyped FECD from Korean patient revealed a glutamine-to-valine (p.Q455V) variation, which was projected to disrupt the interplay between COL8A2 and COL8A1 [18]. Mutations of the COL8A2 found in recent FECD studies are shown in Table 1 [8, 17,18,19,20,21].
TCF4 gene mutations in FECD
The TCF4 gene, also known as E2–2, is found on chromosome 18 and encodes the E2–2 protein, which belongs to a family of basic helix-loop-helix transcription factors. TCF4 plays a valuable role in many developmental processes and is related to transforming growth factor-β (TGF-β) signaling pathways and epithelial-mesenchymal transition (EMT) and programmed cell death [22,23,24]. TCF4 gene mutation is the leading factor causing FECD. A genome-wide association study analysis revealed four single-nucleotide polymorphisms (SNPs) (rs17595731, rs613872, rs9954153, and rs2286812) in TCF4 that were independently correlated with FECD [12]. Subsequent studies confirmed that TCF4 (rs613872) has a significant correlation with FECD [25,26,27,28]. Of note, a recent genome-wide association study reported that TCF4 SNP rs784257 was the most influential SNP in FECD among the discovery specimens; it had a strong correlation imbalance with rs613872 [29]. Another study, the results of which remain to be verified, reported that a SNP was situated in close proximity to the CTG repeat sequence [30]. Moreover, the genome-wide association study identified three novel loci meeting genome-wide significance (P < 5 × 10−8): KANK4 rs79742895, LAMC1 rs3768617 and LINC00970/ATP1B1 rs1200114 [29]. Interestingly, the mutated genes were sex-specific, with LAMC1 mutation at a higher risk in women and TCF4 mutation at a higher risk in men [29].
TCF4 was first found to be caused by the CTG trinucleotide repeat (TNR) amplification in the third intron [31]. Wieben et al.’s study showed that 79% of the Caucasian FECD patients carried repeat lengths of more than 50. In contrast, there were usually only less than 40 repeats in unaffected individuals included in the study. Using genotyping, CTG18.1 allele amplified in a Chinese population was found to have a strong correlation with FECD. Indeed, it may be the main pathogenic variant of FECD in this population [32]. In the Caucasian population, the severity of the disease may be directly related to the repeat length, whereas no such association was found in a Japanese cohort [33,34,35]. Although the correlation between CTG repeat amplified polymorphism and FECD is stronger than that of the TCF4 rs613872 polymorphism, the combination of the two may better predict susceptibility to FECD [27].
TCF8 gene mutations in FECD
The TCF8 gene, also called the zinc finger E-box binding homeobox 1 (ZEB1, MIM 189909), is situated on chromosome 10 and encodes the ZEB1 protein, which can be up-regulated by TCF4 expression [36]. It restrains collagen I expression and mediates EMT; here, the characteristics of epithelial cells disappear, such as cell-cell interaction, and migration and mesenchymal phenotypes are gained [24]. A study examining TCF8 variants in FECD patients (55 women, 19 men) in China found that heterozygous mutations (p.N696S) in TCF8 were present in only one of the patients [37]. Subsequently, another study on a FECD cohort of Caucasian adult males and females identified five missense mutations (p.N78T, p.Q810P, p.Q840P, p.A905G, and p.P649A) in TCF8; a change of one single nucleotide caused a different amino acid to be inserted into the resulting protein [11]. The mutations of the TCF8 gene found in recent FECD studies are shown in Table 2 [11, 19, 37,38,39].
LOXHD1 gene mutations in FECD
The LOXHD1 gene is situated on chromosome 18 and encodes the LOXHD1 protein, which is believed to direct proteins to the plasma membrane [40]. LOXHD1 was initially related to human autosomal recessive and progressive hearing loss [41, 42]. A missense mutation (p.R547C) in LOXHD1 was first confirmed in three large families with FECD. Moreover, a study on a cohort of sporadically affected individuals revealed 14 additional nonsynonymous coding variants and a missense variant (p.L635P); these were absent from control chromosomes [13]. The mutations of the LOXHD1 gene found in recent FECD studies about are shown in Table 3 [13, 38].
SLC4A11 gene mutations in FECD
The SLC4A11 gene is situated on chromosome 20 and encodes the protein SLC4A11, which is usually situated on the cell surface and performs membrane transport functions (OH−/H+/NH3/H2O) [43,44,45]. SLC4A11 is associated not only with FECD but with other types of corneal dystrophy, such as congenital hereditary corneal dystrophy type 2 and Harboyan syndrome [46]. In 2008, three missense variations (p.E399K, p.G709E and p.T754M) and a deletion variation (c.99-100delTC) that did not show up in matched controls were confirmed in FECD patients from India and China- all the variants are presumed to be pathogenic mutations [47]. The study also reported 15 non-pathogenic mutations (ten silent mutations and five missense mutations) [47]. In 2010, seven heterozygous missense novel variations (p.E167D, p.R282P, p.G583D, p.G742R, p.Y526C, p.V575M and p.G834S) resulting in the pathogenesis of adult FECD were identified in a study of 192 sporadic cases and a three-generation family [9]. The mutations of the SLC4A11 gene found in recent FECD studies are shown in Table 4 [9, 19, 20, 47,48,49].
AGBL1 gene mutations in FECD
The AGBL1 gene, also called cytosolic carboxypeptidase 4, is situated on chromosome 15 and encodes the AGBL1 protein [49]. A genome-wide linkage scan of 92 individuals from 22 families with FECD revealed that chromosome 15 was related to FECD [50]. Sequencing of AGBL1 confirmed a causal nonsense mutation (p.R1028*) in a three-generation FECD family. Further sequencing of two FECD-affected cases confirmed this result, and another missense mutation (p.C990S) was found in three unrelated individuals (see Table 5) [51]. Further, an immunoprecipitation assay suggested that AGBL1 protein bound to TCF4 but not to TCF8-a mutation of AGBL1 significantly reduced binding affinity to TCF4 [51].
Molecular mechanisms
Endoplasmic reticulum stress and unfolded protein response pathway
Proper protein folding is critical to overall cellular functioning. Cells have a conservation mechanism in the endoplasmic reticulum (ER) that allows them to avoid protein misfolding and deal with cytotoxic misfolded proteins, of which the excessive accumulation results in ER stress. Unfolded protein response (UPR) is a pro-survival response; it reduces unfolded protein accumulation and restores normal ER function. However, if protein aggregation persists and stress cannot be resolved, the signal changes from pro-survival to pro-apoptotic. COL8A2 accumulates in the ER of the corneal endothelia of FECD patients with L450W COL8A2 mutations [52]. Two studies confirmed that a COL8A2 transgenic knock-in mouse model of FECD exhibited dilated rough ER, overexpression of UPR-associated genes and proteins and UPR-associated apoptosis [53, 54]. A subsequent study confirmed that the missense mutations of SLC4A11 in FECD patients resulted in mutant proteins that accumulated in the ER [47]. Moreover, the LOXHD1 proteins aggregate in corneal cells carrying LOXHD1 mutations in FECD [13]. Analysis of the corneal endothelium from the FECD patients showed enlargement of rough ER and upregulated UPR markers, including the α subunit of the eukaryotic initiation factor 2 (eIF2α), a glucose-regulated protein and a C/EBP homologous protein (CHOP) [55]. TGF-β signaling in the CECs of FECD contributes to the abnormal accumulation of ECM protein in the ER, eventually leading to the formation of unfolded protein and resulting in apoptosis via UPR [23]. One study demonstrated that the deposition of unfolded protein continuously stimulates ER stress, which in turn activates UPR. When the UPR cannot balance with the unfolded protein, it triggers the apoptosis mechanism via three signal transducers: activating transcription factor 6 (ATF6), pancreatic endoplasmic reticulum kinase (PKR)-like endoplasmic reticulum kinase (PERK) and inositol-requiring enzyme 1 (IRE1) (Fig. 1a) [56]. ATF6 is transferred to the Golgi body, where it is hydrolyzed by the Golgi site-1 and site-2 proteases, which subsequently activates the ATF6. Activated ATF6 leads to an increased activity of the CHOP protein (Fig. 1a) [57], which is also activated by activated PERK via phosphorylation of eIF2α (Fig. 1a) [58]. The activation of CHOP can lead to apoptosis [59] and down-regulates B-cell lymphoma-2 (Bcl-2) known as anti-apoptotic protein (Fig. 1a) [60]. Phosphorylation of the c-Jun N-terminal kinase induced by IRE1, subsequently inhibits antiapoptotic genes such as Bcl-2, and thus inhibits apoptosis (Fig. 1a) [61]. Meanwhile, ER stress stimulates the release of Ca2+, which comes into the mitochondria, contributing to mitochondria producing more ATP. At the same time, more reactive oxygen species (ROS) are produced, when the increase of ROS level exceeds a certain threshold, mitochondria will release the mitochondrial cytochrome c, which can activate the apoptosis of CECs by caspase-9 and caspase-3 (Fig. 1a) [62].

Possible pathways leading to the loss of FECD CECs. a Gene mutations lead to the accumulation of unfolded proteins, which continue to activate ER stress, and further induce apoptosis through the three UPR pathways (ATF6, PERK, IRE1). Meanwhile, sustained ER stress can induce cell apoptosis through the mitochondria. b Ca2+ overload in FECD CECs may lead to apoptosis and SLC4A11 mutations are likely to result in CECs edema and rupture. eIF2α: α-subunit of eukaryotic translation initiation factor 2; JNK: c-Jun N-terminal kinase; S1P: site-1 protease; S2P: site-2 protease. The pieces of DNA in red represent the missense mutations of COL8A2 and/or SLC4A11 and/or LOXHD1. The purple cells represent dysfunctional CECs in FECD. The blue moons represent the guttae-the focal excrescences of DM
Oxidative stress
The corneal endothelium is particularly vulnerable to oxidative stress because it is exposed to light and vigorous metabolic activity caused by a high oxygen demand. Increased nitrotyrosine, a by-product of ROS, suggests that oxidative damage indeed occurs in FECD [63]. Because CECs are rich in mitochondria when they are stopped after mitosis, they are highly susceptible to oxidative damage due to their metabolically active pump and barrier functions. Peroxiredoxin are antioxidants that are anti-apoptotic, and Prx-2, -3, -5, and -6 are expressed in different parts of human corneal endothelial cells. A proteomic study confirmed the downregulation of peroxiredoxin (Prx-2, -3 and -5) in corneal endothelia, hinting at an incremental susceptibility to oxidant-induced damage [64]. NADPH quinone oxidoreductase 1 (NQO1) is a highly inducible and cell-protective flavoprotein that restrains the occurrence of ROS and free radicals in a cellular condition [65]. The expression of NQO1 is up-regulated by the nuclear factor erythroid 2-related factor 2 (Nrf2) transcription factor via binding to the sequence of antioxidant reaction elements in the upstream promoter region of NQO1 [66,67,68]. The antioxidant response related to protective Nrf2 is significantly decreased, resulting in the oxidant-antioxidant imbalance that characterizes FECD [66]. A recent report detected that the NQO1 protein levels in FECD specimens down-regulated quite dramatically, a reduction that was further confirmed in FECD patient-derived immortalized cell lines that presented with oxidative DNA damage [69]. The overexpression of NQO1 significantly decreases ROS levels and DNA damage caused by menadione (MN) and catechol estrogen stressors [69]. Moreover, Liu et al. [70] established a nongenetic FECD animal model by exposure to ultraviolet A, which caused greater mitochondrial DNA (mtDNA) and nuclear DNA damage in female mice. The sex-dependent effect of UVA was driven by the activation of estrogen-metabolizing enzyme CYP1B1 and formation of reactive estrogen metabolites and estrogen-DNA adducts in female but not male mice, causing CYP1B1-mediated estrogen genotoxicity. Together, these data confirm that an oxidant-antioxidant imbalance in FECD can induce oxidative DNA damage and apoptosis.
Mitochondrial dysregulation pathway
Mitochondria, found in eukaryotic cells, are organelles covered by two membranes. They are the main site of ATP production within the electron transport chain. In addition to energy supply, mitochondria participate in other cell processes such as regulating calcium levels and apoptosis. The cornea is highly exposed to external elements; it receives a large amount of atmospheric oxygen and sunlight, including ultraviolet light, which prompts ROS generation in the cornea, mitochondrial dysfunction and oxidative damage to the cells. Furthermore, due to its post-mitotic nature, the corneal endothelium tends to accumulate mtDNA damage. Corneal tissue from FECD patients revealed an increase in ROS and 8-hydroxy-2′-deoxyguanosine (both of which are signs of oxidative damage) mainly in the mtDNA of CECs gathered around the rosettes [66]. In one study, FECD patients presented with significantly more mtDNA damage and lower DNA repair efficacy compared to normal controls [71]. Compared with normal corneal specimens, FECD human corneal endothelial cell lines and FECD CECs showed extensive mtDNA and nuclear DNA damage, as measured via quantitative polymerase chain reactions [72]. Subsequent studies increased endogenous cellular oxidative stress with MN, a quinone metabolized by a 1-electron reducing enzyme to produce intracellular superoxide and an unstable semiquinone radical that increases intracellular ROS, to model the pathognomonic rosette formation-a characteristic morphological change of FECD. MN induced rosette formation and damaged the mtDNA and nuclear DNA, which were rescued with N-acetyl-cysteine pre-treatment [72]. One study featuring the FECD explants reported that mtDNA levels increased and telomeres shortened [73]. FECD does not evenly affect the integrity of CECs. Some of the surviving FECD CECs were in a compensatory state of function. The surviving CECs may compensatively increase the mitochondrial content and produce ATP required by the Na/K-ATPase ion pump to maintain the relative dehydrating state of the corneal stroma. Therefore, the level of mtDNA is increased in the surviving FECD CECs. Interestingly, cell culture can rehabilitate mtDNA levels, telomere length, oxidant-antioxidant gene expression balance, and sensitivity to oxidative stress-induced cell death. Therefore, choosing the more functional FECD CECs for cell culture may provide a basis for future treatments [73]. Méthot et al. [74] studied a series of events leading to mitochondrial exhaustion as follows: FECD CECs initially helped to generate more ATP by adding mitochondrial calcium to compensate for endothelial cell loss, leading to increased oxidation. The CECs then increased mitochondrial mass, mitochondrial calcium and mitochondrial membrane potential in response to the continuous demand for more ATP. In this phase, the CECs peaked at their maximum capacity and began to respond to irreversible oxidative damage leading to mitochondrial burnout, causing cell death via apoptosis. Meanwhile, the mitochondrial membrane potential decreased, and calcium was released from the mitochondria.
Apoptosis pathway
Apoptosis, the spontaneous and orderly death of genetically controlled cells, is characterized by cell shrinkage, membrane blebbing, chromatin condensation and DNA fragmentation [75]. Apoptosis is considered to be an important mechanism of FECD. Analysis of endothelium flatmounts showed apoptotic CECs in the FECD group, and the control group showed no apoptotic cells. Further, the observed percentage of apoptotic endothelial cells was much higher in the FECD group compared to the controls. These are the first findings that demonstrate that apoptosis plays a significant role in endothelial cell death in FECD [76]. Subsequently, intense Fas, FasL and Bax staining were found in FECD patients, and faint staining of Bcl-2 was observed occasionally in FECD patients, all of which points to a disturbance in apoptotic regulatory molecules [77]. Eleven of the 14 corneas with FECD showed positive TUNEL labelling, indicating the activation of apoptosis [78]. A recent study detected mitochondrial dysfunction, which can lead to cytochrome c release and subsequent caspase breakage, promoting cell death by apoptosis [72]. Hence, these studies prove that apoptosis is closely related to FECD pathogenesis. Previous studies suggest that CECs’ death may also be caused by other mechanisms. For example, apoptotic CECs in FECD show an increase in mitochondrial Ca2+ [74]. A massive and/or a prolonged accumulation of Ca2+ in the mitochondria can lead to the release of cytochrome c, which drives the activation of caspase-3 [79]. Therefore, an increase in mitochondrial Ca2+ of FECD CECs probably promotes apoptosis (Fig. 1b). Moreover, SLC4A11, localized at the basolateral surface of CECs, contributes to osmotically-driven water flux from the stroma to aqueous humor to maintain the relative dehydration of the cornea [80], and four mutations associated with corneal endothelial dystrophy can impair solute transport (water flux) function [80, 81]. Therefore, SLC4A11 gene mutations potentially result in FECD CECs edema and rupture (Fig. 1b).
Mitophagy
Autophagy is a cellular process featuring ER stress and oxidative stress. Forming autophagosomes and combining them with lysosomes, autophagosomes phagocytose the resulting substance and the organelles in the cell. Autophagy plays an important protective role in cells. However, autophagy defects or overactivation can lead to cell death [82]. In animal models of FECD, DNA-damage regulated autophagy marker 1 was up-regulated [54]. Lithium therapy increased autophagy in mice subjects with FECD, possibly contributing to enhanced endothelial cell survival [83]. Mitochondrial health is determined by mitochondrial quality control namely, fission, fusion and mitophagy [84]. When mitochondrial fission and fusion occurs in repetitive cycles but cannot reduce mitochondrial damage, mitophagy is activated to clear the mitochondria [84]. Increased numbers of autophagic vacuoles were found in FECD tissues containing degraded and swollen mitochondria with cristolysis [85]. In the same study, the elevated autophagosome components (microtubule-associated protein 1 light chain 3-II and lysosomal-associated membrane protein 1) and the downregulation of mitochondrial fusion protein mitofusin 2 in mitochondrial fractions suggested a loss of mitochondrial fusion ability; here, fragmented mitochondria entered the pre-autophagic pool and activated autophagy [85]. Furthermore, in FECD, intracellular oxidative stress induces Parkin-mediated mitochondrial fragmentation whereby endogenous Dynamin-related protein 1 and PTEN-induced putative kinase 1 are segregated for degradation via mitophagy in the process of degenerative cell loss after mitosis of ocular tissue [86].
Epithelial-mesenchymal transition pathway
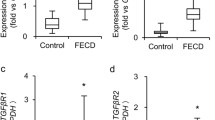
EMT is not only associated with embryonic development, but it is also related to wound healing, organ fibrosis and tumor occurrence and development [87]. The up-regulation of EMT-related genes ZEB1 and SNAI1 by TGF-β in CECs in FECD patients was associated with the deposition of ECM proteins [88]. Furthermore, the TGF-βR1 inhibitor (SB431542) suppressed the expression of ZEB1 and SNAI1, leading to a decrease in ECM. This suggests that blocking the TGF-β signaling pathway is helpful for FECD treatment [88]. A study using an in vitro model of FECD provided the first evidence that oxidative stress induced by MN resulted in EMT, leading to increased expression of SNAI1, ZEB1, fibronectin and N-cadherin in CECs [89]. The overexpression of both isoforms of E2–2 factors (E2–2A and E2–2B) in MDCK cells can induce EMT [90]. Snail1 up-regulates E2–2 expression, which up-regulates ZEB1 expression [24], suggesting that mutations in both TCF4 and TCF8 may share a common pathologic pathway. Moreover, decreased expression of miR29 family members in CECs from FECD may increase the deposition of ECM, including collagen I, collagen IV and laminin [91, 92].
RNA toxicity and repeat-associated non-ATG translation
Wieben et al. [93] were the first to report that the corneal endothelium from FECD patients harbored a unique signature of mis-splicing events caused by CTG TNR expansion in the TCF4 gene. They demonstrated that TNR expansions in the TCF4 gene lead to FECD through a mechanism associated with sequestration of muscleblind-like protein 1 in the RNA foci. The length of the CTG triplet repeat allele seems to be associated with disease severity [35]. Furthermore, TGC repeat lengths > 50 was found in up to 79% individuals with FECD; they were found in only 3% of unrelated individuals, suggesting that trinucleotide amplification may predict disease risk [31]. Interestingly, Foja and colleagues [94] reported the CTG repeat expansion may reduce gene expression of TCF4. By contrast, Okumura and colleagues [95] reported that TCF4 mRNA is upregulated in FECD CECs, regardless of the presence or absence of TNR expansion, but the length of the TNR in cases with expansion tended to be positively correlated with TCF4 expression level. According to a recent study, the levels of TCF4 transcripts change bidirectionally in response to an expanded CTG TNR; a decrease of TCF4 expression of proximal downstream promoters linked to these 5′exons while an increase in the levels of TCF4 transcripts encoded by downstream alternative 5′exons distal to the CTG TNR, possibly indicating a compensatory mechanism, explaining why previous studies on the level of TCF4 transcripts in FECD showed different results [96]. Almost all of the TNR expansion diseases thus far have been directly related to rare neurologic or neuromuscular diseases. Among eye diseases, FECD was first found to be related to TNR expansion. The expanded CTG·CAG repeat initiates transcription and translation through non-ATG in the third intron of TCF4, which provides a basis for studying repeat-related non-ATG translation in the CECs of FECD patients [97]. RNA focal points co-localized with the splicing factor muscleblind-like protein 1 in CECs from FECD patients; mRNA splicing changes also occurred. Combined, these represent the first evidence of RNA toxicity and mismatch in common non-neuro/neuromuscular diseases associated with repetitive expansion [98].
Other pathogenesis
A recent study found that SLC4A11 is a cell adhesion molecule, mediating CECs adhesion to DM. Four FECD-causing mutations in SLC4A11 extracellular loop 3 (Y526C, T561M, S565L and V575M) lead to the destruction of the adhesion of CECs to DM, which may explain the loss of CECs in FECD patients [99]. Moreover, a study demonstrated that DNA methylation alterations are crucial to the pathogenesis of FECD [100]. This study showed that promoter DNA hypermethylation of SLC4A11, which is critical to water transport in FECD CECs. Promoters of genes involved in cytoskeletal organization, which plays an important role in the barrier integrity of the corneal endothelium and restrict fluid leakage into the corneal stroma, tend to be hypomethylated in FECD. Promoter DNA hypermethylation of genes involved in cellular metabolism plays an important energy metabolism role in FECD CECs. All of the DNA methylation changes in genes, associated with cytoskeletal organization, cellular metabolism, and ion transport occurred in FECD CECs, may contribute to the loss of corneal transparency in FECD through changing corneal endothelia biological processes [100]. Therefore, drugs targeting DNA methylation can be developed and used for FECD treatment.
Current therapeutic modalities
In the past few decades, penetrating keratoplasty has been an effective treatment option for FECD. However, with the innovation of surgical techniques, lamellar keratoplasty effectively utilizes limited corneal specimens. Endothelial keratoplasty has offered some distinguishing benefits such as better vision recovery, less damage to the corneal structure and reduced incidence of bleeding, infection and endothelial rejection. Endothelial keratoplasty includes Descemet-stripping endothelial keratoplasty [101], Descemet membrane endothelial keratoplasty [102] and Descemetorhexis without endothelial keratoplasty [103].
Future therapies
Cell-based therapy
Corneal cell therapy is a new treatment strategy for FECD; it refers to the in vitro culture and expansion of primary human corneal cells before transplantation. Compared with existing healthy cells, this cell culture can restore the molecular phenotype associated with oxidative stress by selecting the more functional FECD cells [73]. In this treatment, the operator injects human CECs that were cultured in vitro into the anterior chamber. The patient then performs different maneuvers, such as assuming the prone position, to promote adhesion of the CECs to the DM [104]. A recent significant study confirmed the therapeutic effect of corneal cell therapy in patients with bullous keratopathy that was mainly caused by FECD [105]. The study reported that injecting cultured human CECs into the anterior chamber successfully reversed corneal edema; the clinical results were stable in the 2 years following the operation.
Gene therapy
FECD is typically a sporadic disease, but it can also take the form of autosomal dominant inheritance [2, 3]. Despite FECD being a genetically heterogenous disease, most humans with FECD, at least among Caucasian patients, have a CTG TNR expansion sequence in chromosome 18q21 of TCF4 [106]. The mutant TCF4 transcript accumulates with the repeated amplification of CTG, and its pathological effects reflect the cumulation of RNA lesions and the isolation of RNA splicing factors in the nucleus. This is similar to myotonic dystrophy-1, which is a disease associated with TNR. Researchers must keep exploring effective treatments for these diseases. For example, in muscular dystrophy-1 cell lines, inactivated Cas9 can prevent the transcription from TNR amplification. Deactivated Cas9 enzyme (dCas9) could be designed to efficaciously connect with the trinucleotide DNA repeat sequence of myotonic dystrophy type 1 cells, and thus restrain the transcription of amplified mRNA molecules [107]. Further evidence suggest that these dCas9 molecules are able to complex with pathological elongated mRNA molecules and ameliorate deleterious effects, especially in short and intermediate repeat lengths [107]. All of the studies indicated that similar dCas9 strategies, especially targeting CECs through intracameral delivery, may be able to effectively tackle the genetic variation of FECD and restore its normal phenotype.
Other therapies
In FECD, oxidative stress causes excessive endothelial cell apoptosis [66]. Consequently, a potential therapy for FECD is to target this pathway. N-acetyl-cysteine, an antioxidant and free radical scavenger, has been shown to rescue CECs exposed to oxidative stress and ER stress not only in vitro but also among in vivo animal models with FECD [72, 89, 108]. Research on Nrf2-related antioxidant defense deficiency provides a basis for investigating whether Nrf2 stimulator play a cellular protective role in FECD [66]. Sulforaphane, a natural glucosinolate, was found in green cruciferous vegetables [109]; it has a cytoprotective function, can increase the nuclear translocation of Nrf2, reduce the production of ROS and up-regulate several antioxidants, and thereby reduce CECs apoptosis [110]. Furthermore, Nrf2 levels can be reinforced by many other compounds (e.g., 3H-1,2-dithiole-3-thione), which intervenes in Nrf2 degradation [111, 112]. Meanwhile, TGF-β, an important regulator of EMT, is up-regulated in FECD, inducing the deposition of ECM proteins and resulting in apoptosis via the UPR in FECD [23]. Inhibition of TGF-β can suppress aggregation accumulation and the UPR as well as the activation of apoptosis [23]. Moreover, a recent study identified a non-steroidal anti-inflammatory drug, glafenine, which can correct cell surface trafficking defects in some SLC4A11 mutants, leading to increased SLC4A11-mediated water flux in cells expressing the treated mutants, providing a framework for future personalized medicine approaches to correct SLC4A11 misfolding mutants present in FECD CECs [113]. Further studies are needed to demonstrate whether these approaches are feasible therapy options for FECD patients.
Conclusions
This review reveals that our understanding of the pathogenesis of FECD and the development of molecular genetics is becoming more profound. The gene mutation site has been established and the molecular mechanisms are becoming clearer. However, many questions regarding the pathogenesis remain elusive. Overall, the data from the studies included here illuminate the molecular mechanisms associated with FECD and may help to optimize various therapeutic approaches.
Availability of data and materials
Not applicable.
Abbreviations
- FECD:
-
Fuchs endothelial corneal dystrophy
- COL8A2 :
-
Collagen Type VIII Alpha 2 Chain
- TCF4 :
-
Transcription factor 4 gene
- TCF8 :
-
Transcription factor 8 gene
- ZEB1 :
-
Zinc finger E-box binding homeobox 1
- LOXHD1 :
-
Lipoxygenase homology domains 1
- SLC4A11 :
-
Solute carrier family 4 sodium borate transporter member 11
- AGBL1 :
-
ATP/GTP binding protein like 1
- ER:
-
Endoplasmic reticulum
- CECs:
-
Corneal endothelial cells
- ECM:
-
Extracellular matrix
- TGF-β:
-
Transforming growth factor-β
- SNPs:
-
Single-nucleotide polymorphisms
- UPR:
-
Unfolded protein response
- PERK:
-
Pancreatic endoplasmic reticulum kinase (PKR)-like endoplasmic reticulum kinase
- ATF6:
-
Activating transcription factor 6
- IRE1:
-
Inositol requiring enzyme 1
- eIF2α:
-
α subunit of eukaryotic initiation factor 2
- CHOP:
-
C/EBP homologous protein
- S1P:
-
Site-1 protease
- S2P:
-
Site-2 protease
- Bcl-2:
-
B-cell lymphoma-2
- JNK:
-
c-Jun N-terminal kinase
- ROS:
-
Reactive oxygen species
- NQO1:
-
NADPH quinone oxidoreductase 1
- Nrf2:
-
Nuclear factor erythroid 2-related factor 2
- mtDNA:
-
Mitochondrial DNA
- EMT:
-
Epithelial-mesenchymal transition
- TNR:
-
Trinucleotide repeat
- DM:
-
Descemet’s membrane
- dCas9:
-
Deactivated Cas9 enzyme
References
Adamis AP, Filatov V, Tripathi BJ, Tripathi RC. Fuchs' endothelial dystrophy of the cornea. Surv Ophthalmol. 1993;38(2):149–68.
Cross HE, Maumenee AE, Cantolino SJ. Inheritance of Fuchs' endothelial dystrophy. Arch Ophthalmol. 1971;85(3):268–72.
Rosenblum P, Stark WJ, Maumenee IH, Hirst LW, Maumenee AE. Hereditary Fuchs' dystrophy. Am J Ophthalmol. 1980;90(4):455–62.
Sarnicola C, Farooq AV, Colby K. Fuchs endothelial corneal dystrophy: update on pathogenesis and future directions. Eye Contact Lens. 2019;45(1):1–10.
Waring GO 3rd, Bourne WM, Edelhauser HF, Kenyon KR. The corneal endothelium. Normal and pathologic structure and function. Ophthalmology. 1982;89(6):531–90.
Elhalis H, Azizi B, Jurkunas UV. Fuchs endothelial corneal dystrophy. Ocul Surf. 2010;8(4):173–84.
Hamill CE, Schmedt T, Jurkunas U. Fuchs endothelial cornea dystrophy: a review of the genetics behind disease development. Semin Ophthalmol. 2013;28(5–6):281–6.
Gottsch JD, Sundin OH, Liu SH, Jun AS, Broman KW, Stark WJ, et al. Inheritance of a novel COL8A2 mutation defines a distinct early-onset subtype of Fuchs corneal dystrophy. Invest Ophthalmol Vis Sci. 2005;46(6):1934–9.
Riazuddin SA, Vithana EN, Seet LF, Liu Y, Al-Saif A, Koh LW, et al. Missense mutations in the sodium borate cotransporter SLC4A11 cause late-onset Fuchs corneal dystrophy. Hum Mutat. 2010;31(11):1261–8.
Malhotra D, Loganathan SK, Chiu AM, Lukowski CM, Casey JR. Human corneal expression of SLC4A11, a gene mutated in endothelial corneal dystrophies. Sci Rep. 2019;9(1):9681.
Riazuddin SA, Zaghloul NA, Al-Saif A, Davey L, Diplas BH, Meadows DN, et al. Missense mutations in TCF8 cause late-onset Fuchs corneal dystrophy and interact with FCD4 on chromosome 9p. Am J Hum Genet. 2010;86(1):45–53.
Baratz KH, Tosakulwong N, Ryu E, Brown WL, Branham K, Chen W, et al. E2-2 protein and Fuchs's corneal dystrophy. N Engl J Med. 2010;363(11):1016–24.
Riazuddin SA, Parker DS, McGlumphy EJ, Oh EC, Iliff BW, Schmedt T, et al. Mutations in LOXHD1, a recessive-deafness locus, cause dominant late-onset Fuchs corneal dystrophy. Am J Hum Genet. 2012;90(3):533–9.
Patel SV, McLaren JW. In vivo confocal microscopy of Fuchs endothelial dystrophy before and after endothelial keratoplasty. JAMA Ophthalmol. 2013;131(5):611–8.
Eghrari AO, Riazuddin SA, Gottsch JD. Fuchs corneal dystrophy. Prog Mol Biol Transl Sci. 2015;134:79–97.
Wilson SE, Bourne WM. Fuchs' dystrophy. Cornea. 1988;7(1):2–18.
Biswas S, Munier FL, Yardley J, Hart-Holden N, Perveen R, Cousin P, et al. Missense mutations in COL8A2, the gene encoding the alpha2 chain of type VIII collagen, cause two forms of corneal endothelial dystrophy. Hum Mol Genet. 2001;10(21):2415–23.
Mok JW, Kim HS, Joo CK. Q455V mutation in COL8A2 is associated with Fuchs' corneal dystrophy in Korean patients. Eye (Lond). 2009;23(4):895–903.
Minear MA, Li YJ, Rimmler J, Balajonda E, Watson S, Allingham RR, et al. Genetic screen of African Americans with Fuchs endothelial corneal dystrophy. Mol Vis. 2013;19:2508–16.
Tang H, Zhang W, Yan XM, Wang LP, Dong H, Shou T, et al. Analysis of SLC4A11, ZEB1, LOXHD1, COL8A2 and TCF4 gene sequences in a multi-generational family with late-onset Fuchs corneal dystrophy. Int J Mol Med. 2016;37(6):1487–500.
Moschos MM, Diamantopoulou A, Gouliopoulos N, Droutsas K, Bagli E, Chatzistefanou K, et al. TCF4 and COL8A2 gene polymorphism screening in a Greek population of late-onset Fuchs endothelial corneal dystrophy. In Vivo. 2019;33(3):963–71.
Forrest MP, Waite AJ, Martin-Rendon E, Blake DJ. Knockdown of human TCF4 affects multiple signaling pathways involved in cell survival, epithelial to mesenchymal transition and neuronal differentiation. PLoS One. 2013;8(8):e73169.
Okumura N, Hashimoto K, Kitahara M, Okuda H, Ueda E, Watanabe K, et al. Activation of TGF-beta signaling induces cell death via the unfolded protein response in Fuchs endothelial corneal dystrophy. Sci Rep. 2017;7(1):6801.
Cano A, Portillo F. An emerging role for class I bHLH E2-2 proteins in EMT regulation and tumor progression. Cell Adhes Migr. 2010;4(1):56–60.
Riazuddin SA, McGlumphy EJ, Yeo WS, Wang J, Katsanis N, Gottsch JD. Replication of the TCF4 intronic variant in late-onset Fuchs corneal dystrophy and evidence of independence from the FCD2 locus. Invest Ophthalmol Vis Sci. 2011;52(5):2825–9.
Rao BS, Tharigopala A, Rachapalli SR, Rajagopal R, Soumittra N. Association of polymorphisms in the intron of TCF4 gene to late-onset Fuchs endothelial corneal dystrophy: an Indian cohort study. Indian J Ophthalmol. 2017;65(10):931–5.
Li D, Peng X, Sun H. Association of TCF4 polymorphisms and Fuchs' endothelial dystrophy: a meta-analysis. BMC Ophthalmol. 2015;15:61.
Okumura N, Hayashi R, Nakano M, Tashiro K, Yoshii K, Aleff R, et al. Association of rs613872 and trinucleotide repeat expansion in the TCF4 gene of German patients with Fuchs endothelial corneal dystrophy. Cornea. 2019;38(7):799–805.
Afshari NA, Igo RP Jr, Morris NJ, Stambolian D, Sharma S, Pulagam VL, et al. Genome-wide association study identifies three novel loci in Fuchs endothelial corneal dystrophy. Nat Commun. 2017;8(1):14898.
Wieben ED, Aleff RA, Eckloff BW, Atkinson EJ, Baheti S, Middha S, et al. Comprehensive assessment of genetic variants within TCF4 in Fuchs' endothelial corneal dystrophy. Invest Ophthalmol Vis Sci. 2014;55(9):6101–7.
Wieben ED, Aleff RA, Tosakulwong N, Butz ML, Highsmith WE, Edwards AO, et al. A common trinucleotide repeat expansion within the transcription factor 4 (TCF4, E2-2) gene predicts Fuchs corneal dystrophy. PLoS One. 2012;7(11):e49083.
Xing C, Gong X, Hussain I, Khor CC, Tan DT, Aung T, et al. Transethnic replication of association of CTG18.1 repeat expansion of TCF4 gene with Fuchs' corneal dystrophy in Chinese implies common causal variant. Invest Ophthalmol Vis Sci. 2014;55(11):7073–8.
Nakano M, Okumura N, Nakagawa H, Koizumi N, Ikeda Y, Ueno M, et al. Trinucleotide repeat expansion in the TCF4 gene in Fuchs' endothelial corneal dystrophy in Japanese. Invest Ophthalmol Vis Sci. 2015;56(8):4865–9.
Wieben ED, Aleff RA, Tang X, Kalari KR, Maguire LJ, Patel SV, et al. Gene expression in the corneal endothelium of Fuchs endothelial corneal dystrophy patients with and without expansion of a trinucleotide repeat in TCF4. PLoS One. 2018;13(7):e0200005.
Soliman AZ, Xing C, Radwan SH, Gong X, Mootha VV. Correlation of severity of Fuchs endothelial corneal dystrophy with triplet repeat expansion in TCF4. JAMA Ophthalmol. 2015;133(12):1386–91.
Vedana G, Villarreal G Jr, Jun AS. Fuchs endothelial corneal dystrophy: current perspectives. Clin Ophthalmol. 2016;10:321–30.
Mehta JS, Vithana EN, Tan DT, Yong VH, Yam GH, Law RW, et al. Analysis of the posterior polymorphous corneal dystrophy 3 gene, TCF8, in late-onset Fuchs endothelial corneal dystrophy. Invest Ophthalmol Vis Sci. 2008;49(1):184–8.
Rao BS, Ansar S, Arokiasamy T, Sudhir RR, Umashankar V, Rajagopal R, et al. Analysis of candidate genes ZEB1 and LOXHD1 in late-onset Fuchs' endothelial corneal dystrophy in an Indian cohort. Ophthalmic Genet. 2018;39(4):443–9.
Gupta R, Kumawat BL, Paliwal P, Tandon R, Sharma N, Sen S, et al. Association of ZEB1 and TCF4 rs613872 changes with late onset Fuchs endothelial corneal dystrophy in patients from northern India. Mol Vis. 2015;21:1252–60.
Zhang J, McGhee CNJ, Patel DV. The molecular basis of Fuchs' endothelial corneal dystrophy. Mol Diagn Ther. 2019;23(1):97–112.
Grillet N, Schwander M, Hildebrand MS, Sczaniecka A, Kolatkar A, Velasco J, et al. Mutations in LOXHD1, an evolutionarily conserved stereociliary protein, disrupt hair cell function in mice and cause progressive hearing loss in humans. Am J Hum Genet. 2009;85(3):328–37.
Edvardson S, Jalas C, Shaag A, Zenvirt S, Landau C, Lerer I, et al. A deleterious mutation in the LOXHD1 gene causes autosomal recessive hearing loss in Ashkenazi Jews. Am J Med Genet A. 2011;155A(5):1170–2.
Badior KE, Alka K, Casey JR. SLC4A11 three-dimensional homology model rationalizes corneal dystrophy-causing mutations. Hum Mutat. 2017;38(3):279–88.
Zhang W, Ogando DG, Bonanno JA, Obukhov AG. Human SLC4A11 is a novel NH3/H+ co-transporter. J Biol Chem. 2015;290(27):16894–905.
Jalimarada SS, Ogando DG, Vithana EN, Bonanno JA. Ion transport function of SLC4A11 in corneal endothelium. Invest Ophthalmol Vis Sci. 2013;54(6):4330–40.
Loganathan SK, Casey JR. Corneal dystrophy-causing SLC4A11 mutants: suitability for folding-correction therapy. Hum Mutat. 2014;35(9):1082–91.
Vithana EN, Morgan PE, Ramprasad V, Tan DT, Yong VH, Venkataraman D, et al. SLC4A11 mutations in Fuchs endothelial corneal dystrophy. Hum Mol Genet. 2008;17(5):656–66.
Soumittra N, Loganathan SK, Madhavan D, Ramprasad VL, Arokiasamy T, Sumathi S, et al. Biosynthetic and functional defects in newly identified SLC4A11 mutants and absence of COL8A2 mutations in Fuchs endothelial corneal dystrophy. J Hum Genet. 2014;59(8):444–53.
Zhang J, Wu D, Li Y, Fan Y, Chen H, Hong J, et al. Novel mutations associated with various types of corneal dystrophies in a Han Chinese population. Front Genet. 2019;10:881.
Afshari NA, Li YJ, Pericak-Vance MA, Gregory S, Klintworth GK. Genome-wide linkage scan in fuchs endothelial corneal dystrophy. Invest Ophthalmol Vis Sci. 2009;50(3):1093–7.
Riazuddin SA, Vasanth S, Katsanis N, Gottsch JD. Mutations in AGBL1 cause dominant late-onset Fuchs corneal dystrophy and alter protein-protein interaction with TCF4. Am J Hum Genet. 2013;93(4):758–64.
Zhang C, Bell WR, Sundin OH, De La Cruz Z, Stark WJ, Green WR, et al. Immunohistochemistry and electron microscopy of early-onset fuchs corneal dystrophy in three cases with the same L450W COL8A2 mutation. Trans Am Ophthalmol Soc. 2006;104:85–97.
Jun AS, Meng H, Ramanan N, Matthaei M, Chakravarti S, Bonshek R, et al. An alpha 2 collagen VIII transgenic knock-in mouse model of Fuchs endothelial corneal dystrophy shows early endothelial cell unfolded protein response and apoptosis. Hum Mol Genet. 2012;21(2):384–93.
Meng H, Matthaei M, Ramanan N, Grebe R, Chakravarti S, Speck CL, et al. L450W and Q455K Col8a2 knock-in mouse models of Fuchs endothelial corneal dystrophy show distinct phenotypes and evidence for altered autophagy. Invest Ophthalmol Vis Sci. 2013;54(3):1887–97.
Engler C, Kelliher C, Spitze AR, Speck CL, Eberhart CG, Jun AS. Unfolded protein response in fuchs endothelial corneal dystrophy: a unifying pathogenic pathway? Am J Ophthalmol. 2010;149(2):194–202 e2.
Okumura N, Kitahara M, Okuda H, Hashimoto K, Ueda E, Nakahara M, et al. Sustained activation of the unfolded protein response induces cell death in Fuchs' endothelial corneal dystrophy. Invest Ophthalmol Vis Sci. 2017;58(9):3697–707.
Shen J, Chen X, Hendershot L, Prywes R. ER stress regulation of ATF6 localization by dissociation of BiP/GRP78 binding and unmasking of Golgi localization signals. Dev Cell. 2002;3(1):99–111.
Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol. 2007;8(7):519–29.
McCullough KD, Martindale JL, Klotz LO, Aw TY, Holbrook NJ. Gadd153 sensitizes cells to endoplasmic reticulum stress by down-regulating Bcl2 and perturbing the cellular redox state. Mol Cell Biol. 2001;21(4):1249–59.
Tabas I, Ron D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat Cell Biol. 2011;13(3):184–90.
Chen Y, Brandizzi F. IRE1: ER stress sensor and cell fate executor. Trends Cell Biol. 2013;23(11):547–55.
Lee HC, Wei YH. Mitochondrial role in life and death of the cell. J Biomed Sci. 2000;7(1):2–15.
Buddi R, Lin B, Atilano SR, Zorapapel NC, Kenney MC, Brown DJ. Evidence of oxidative stress in human corneal diseases. J Histochem Cytochem. 2002;50(3):341–51.
Jurkunas UV, Rawe I, Bitar MS, Zhu C, Harris DL, Colby K, et al. Decreased expression of peroxiredoxins in Fuchs' endothelial dystrophy. Invest Ophthalmol Vis Sci. 2008;49(7):2956–63.
Dinkova-Kostova AT, Talalay P. NAD(P)H:quinone acceptor oxidoreductase 1 (NQO1), a multifunctional antioxidant enzyme and exceptionally versatile cytoprotector. Arch Biochem Biophys. 2010;501(1):116–23.
Jurkunas UV, Bitar MS, Funaki T, Azizi B. Evidence of oxidative stress in the pathogenesis of fuchs endothelial corneal dystrophy. Am J Pathol. 2010;177(5):2278–89.
Favreau LV, Pickett CB. The rat quinone reductase antioxidant response element. Identification of the nucleotide sequence required for basal and inducible activity and detection of antioxidant response element-binding proteins in hepatoma and non-hepatoma cell lines. J Biol Chem. 1995;270(41):24468–74.
Nioi P, McMahon M, Itoh K, Yamamoto M, Hayes JD. Identification of a novel Nrf2-regulated antioxidant response element (ARE) in the mouse NAD(P)H:quinone oxidoreductase 1 gene: reassessment of the ARE consensus sequence. Biochem J. 2003;374(Pt 2):337–48.
Miyajima T, Melangath G, Zhu S, Deshpande N, Vasanth S, Mondal B, et al. Loss of NQO1 generates genotoxic estrogen-DNA adducts in Fuchs endothelial corneal dystrophy. Free Radic Biol Med. 2020;147:69–79.
Liu C, Miyajima T, Melangath G, Miyai T, Vasanth S, Deshpande N, et al. Ultraviolet A light induces DNA damage and estrogen-DNA adducts in Fuchs endothelial corneal dystrophy causing females to be more affected. Proc Natl Acad Sci U S A. 2020;117(1):573–83.
Czarny P, Seda A, Wielgorski M, Binczyk E, Markiewicz B, Kasprzak E, et al. Mutagenesis of mitochondrial DNA in Fuchs endothelial corneal dystrophy. Mutat Res. 2014;760:42–7.
Halilovic A, Schmedt T, Benischke AS, Hamill C, Chen Y, Santos JH, et al. Menadione-induced DNA damage leads to mitochondrial dysfunction and fragmentation during rosette formation in Fuchs endothelial corneal dystrophy. Antioxid Redox Signal. 2016;24(18):1072–83.
Gendron SP, Thériault M, Proulx S, Brunette I, Rochette PJ. Restoration of mitochondrial integrity, telomere length, and sensitivity to oxidation by in vitro culture of Fuchs' endothelial corneal dystrophy cells. Invest Ophthalmol Vis Sci. 2016;57(14):5926–34.
Méthot SJ, Proulx S, Brunette I, Rochette PJ. Chronology of cellular events related to mitochondrial burnout leading to cell death in Fuchs endothelial corneal dystrophy. Sci Rep. 2020;10(1):5811.
Kerr JF, Wyllie AH, Currie AR. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer. 1972;26(4):239–57.
Borderie VM, Baudrimont M, Vallée A, Ereau TL, Gray F, Laroche L. Corneal endothelial cell apoptosis in patients with Fuchs' dystrophy. Invest Ophthalmol Vis Sci. 2000;41(9):2501–5.
Li QJ, Ashraf MF, Shen DF, Green WR, Stark WJ, Chan CC, et al. The role of apoptosis in the pathogenesis of Fuchs endothelial dystrophy of the cornea. Arch Ophthalmol. 2001;119(11):1597–604.
Szentmary N, Szende B, Süveges I. Epithelial cell, keratocyte, and endothelial cell apoptosis in Fuchs' dystrophy and in pseudophakic bullous keratopathy. Eur J Ophthalmol. 2005;15(1):17–22.
Jeong SY, Seol DW. The role of mitochondria in apoptosis. BMB Rep. 2008;41(1):11–22.
Vilas GL, Loganathan SK, Liu J, Riau AK, Young JD, Mehta JS, et al. Transmembrane water-flux through SLC4A11: a route defective in genetic corneal diseases. Hum Mol Genet. 2013;22(22):4579–90.
Li S, Hundal KS, Chen X, Choi M, Ogando DG, Obukhov AG, et al. R125H, W240S, C386R, and V507I SLC4A11 mutations associated with corneal endothelial dystrophy affect the transporter function but not trafficking in PS120 cells. Exp Eye Res. 2019;180:86–91.
Nikoletopoulou V, Markaki M, Palikaras K, Tavernarakis N. Crosstalk between apoptosis, necrosis and autophagy. Biochim Biophys Acta. 2013;1833(12):3448–59.
Kim EC, Meng H, Jun AS. Lithium treatment increases endothelial cell survival and autophagy in a mouse model of Fuchs endothelial corneal dystrophy. Br J Ophthalmol. 2013;97(8):1068–73.
Youle RJ, van der Bliek AM. Mitochondrial fission, fusion, and stress. Science. 2012;337(6098):1062–5.
Benischke AS, Vasanth S, Miyai T, Katikireddy KR, White T, Chen Y, et al. Activation of mitophagy leads to decline in Mfn2 and loss of mitochondrial mass in Fuchs endothelial corneal dystrophy. Sci Rep. 2017;7(1):6656.
Miyai T, Vasanth S, Melangath G, Deshpande N, Kumar V, Benischke AS, et al. Activation of PINK1-Parkin-mediated mitophagy degrades mitochondrial quality control proteins in Fuchs endothelial corneal dystrophy. Am J Pathol. 2019;189(10):2061–76.
Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–96.
Okumura N, Minamiyama R, Ho LT, Kay EP, Kawasaki S, Tourtas T, et al. Involvement of ZEB1 and Snail1 in excessive production of extracellular matrix in Fuchs endothelial corneal dystrophy. Lab Invest. 2015;95(11):1291–304.
Katikireddy KR, White TL, Miyajima T, Vasanth S, Raoof D, Chen Y, et al. NQO1 downregulation potentiates menadione-induced endothelial-mesenchymal transition during rosette formation in Fuchs endothelial corneal dystrophy. Free Radic Biol Med. 2018;116:19–30.
Sobrado VR, Moreno-Bueno G, Cubillo E, Holt LJ, Nieto MA, Portillo F, et al. The class I bHLH factors E2-2A and E2-2B regulate EMT. J Cell Sci. 2009;122(Pt 7):1014–24.
Matthaei M, Hu J, Kallay L, Eberhart CG, Cursiefen C, Qian J, et al. Endothelial cell microRNA expression in human late-onset Fuchs' dystrophy. Invest Ophthalmol Vis Sci. 2014;55(1):216–25.
Toyono T, Usui T, Villarreal G Jr, Kallay L, Matthaei M, Vianna LM, et al. MicroRNA-29b overexpression decreases extracellular matrix mRNA and protein production in human corneal endothelial cells. Cornea. 2016;35(11):1466–70.
Wieben ED, Aleff RA, Tang X, Butz ML, Kalari KR, Highsmith EW, et al. Trinucleotide repeat expansion in the transcription factor 4 (TCF4) gene leads to widespread mRNA splicing changes in Fuchs' endothelial corneal dystrophy. Invest Ophthalmol Vis Sci. 2017;58(1):343–52.
Foja S, Luther M, Hoffmann K, Rupprecht A, Gruenauer-Kloevekorn C. CTG18.1 repeat expansion may reduce TCF4 gene expression in corneal endothelial cells of German patients with Fuchs' dystrophy. Graefes Arch Clin Exp Ophthalmol. 2017;255(8):1621–31.
Okumura N, Hayashi R, Nakano M, Yoshii K, Tashiro K, Sato T, et al. Effect of trinucleotide repeat expansion on the expression of TCF4 mRNA in Fuchs' endothelial corneal dystrophy. Invest Ophthalmol Vis Sci. 2019;60(2):779–86.
Sirp A, Leite K, Tuvikene J, Nurm K, Sepp M, Timmusk T. The Fuchs corneal dystrophy-associated CTG repeat expansion in the TCF4 gene affects transcription from its alternative promoters. Sci Rep. 2020;10(1):18424.
Soragni E, Petrosyan L, Rinkoski TA, Wieben ED, Baratz KH, Fautsch MP, et al. Repeat-associated non-ATG (RAN) translation in Fuchs' endothelial corneal dystrophy. Invest Ophthalmol Vis Sci. 2018;59(5):1888–96.
Du J, Aleff RA, Soragni E, Kalari K, Nie J, Tang X, et al. RNA toxicity and missplicing in the common eye disease fuchs endothelial corneal dystrophy. J Biol Chem. 2015;290(10):5979–90.
Malhotra D, Jung M, Fecher-Trost C, Lovatt M, Peh GSL, Noskov S, et al. Defective cell adhesion function of solute transporter, SLC4A11, in endothelial corneal dystrophies. Hum Mol Genet. 2020;29(1):97–116.
Khuc E, Bainer R, Wolf M, Clay SM, Weisenberger DJ, Kemmer J, et al. Comprehensive characterization of DNA methylation changes in Fuchs endothelial corneal dystrophy. PLoS One. 2017;12(4):e0175112.
Anshu A, Price MO, Tan DT, Price FW Jr. Endothelial keratoplasty: a revolution in evolution. Surv Ophthalmol. 2012;57(3):236–52.
Ham L, Dapena I, van Luijk C, van der Wees J, Melles GR. Descemet membrane endothelial keratoplasty (DMEK) for Fuchs endothelial dystrophy: review of the first 50 consecutive cases. Eye (Lond). 2009;23(10):1990–8.
Huang MJ, Kane S, Dhaliwal DK. Descemetorhexis without endothelial keratoplasty versus DMEK for treatment of Fuchs endothelial corneal dystrophy. Cornea. 2018;37(12):1479–83.
Mehta JS, Kocaba V, Soh YQ. The future of keratoplasty: cell-based therapy, regenerative medicine, bioengineering keratoplasty, gene therapy. Curr Opin Ophthalmol. 2019;30(4):286–91.
Kinoshita S, Koizumi N, Ueno M, Okumura N, Imai K, Tanaka H, et al. Injection of cultured cells with a ROCK inhibitor for bullous keratopathy. N Engl J Med. 2018;378(11):995–1003.
Soh YQ, Peh GS, Mehta JS. Evolving therapies for Fuchs' endothelial dystrophy. Regen Med. 2018;13(1):97–115.
Pinto BS, Saxena T, Oliveira R, Mendez-Gomez HR, Cleary JD, Denes LT, et al. Impeding transcription of expanded microsatellite repeats by deactivated Cas9. Mol Cell. 2017;68(3):479–90 e5.
Kim EC, Meng H, Jun AS. N-acetylcysteine increases corneal endothelial cell survival in a mouse model of Fuchs endothelial corneal dystrophy. Exp Eye Res. 2014;127:20–5.
Shapiro TA, Fahey JW, Wade KL, Stephenson KK, Talalay P. Human metabolism and excretion of cancer chemoprotective glucosinolates and isothiocyanates of cruciferous vegetables. Cancer Epidemiol Biomark Prev. 1998;7(12):1091–100.
Ziaei A, Schmedt T, Chen Y, Jurkunas UV. Sulforaphane decreases endothelial cell apoptosis in Fuchs endothelial corneal dystrophy: a novel treatment. Invest Ophthalmol Vis Sci. 2013;54(10):6724–34.
Manandhar S, Cho JM, Kim JA, Kensler TW, Kwak MK. Induction of Nrf2-regulated genes by 3H-1, 2-dithiole-3-thione through the ERK signaling pathway in murine keratinocytes. Eur J Pharmacol. 2007;577(1–3):17–27.
Soriano FX, Leveille F, Papadia S, Higgins LG, Varley J, Baxter P, et al. Induction of sulfiredoxin expression and reduction of peroxiredoxin hyperoxidation by the neuroprotective Nrf2 activator 3H-1,2-dithiole-3-thione. J Neurochem. 2008;107(2):533–43.
Alka K, Casey JR. Molecular phenotype of SLC4A11 missense mutants: setting the stage for personalized medicine in corneal dystrophies. Hum Mutat. 2018;39(5):676–90.
Acknowledgments
Not applicable.
Funding
This work was supported by the Heilongjiang Postdoctoral Fund (Grant No. LBH-Z18185) and the Heilongjiang Provincial Colleges and Universities Basic Scientific Research Fund (Grant No. 2018-KYYWF-0484).
Author information
Authors and Affiliations
Contributions
PL and LW conceived the idea for the article, TZ and CZ performed literature search and collected data, YZ and HL analyzed the data, XL and TZ drafted and critically revised the work for intellectual content. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, X., Zheng, T., Zhao, C. et al. Genetic mutations and molecular mechanisms of Fuchs endothelial corneal dystrophy. Eye and Vis 8, 24 (2021). https://doi.org/10.1186/s40662-021-00246-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40662-021-00246-2