
Abstract
Protein-encoding genes only constitute less than 2% of total human genomic sequences, and 98% of genetic information was previously referred to as “junk DNA”. Meanwhile, non-coding RNAs (ncRNAs) consist of approximately 60% of the transcriptional output of human cells. Thousands of ncRNAs have been identified in recent decades, and their essential roles in the regulation of gene expression in diverse cellular pathways associated with fundamental cell processes, including proliferation, differentiation, apoptosis, and metabolism, have been extensively investigated. Furthermore, the gene regulation networks they form modulate gene expression in normal development and under pathological conditions. In this review, we integrate current information about the classification, biogenesis, and function of ncRNAs and how these ncRNAs support skeletal development through their regulation of critical genes and signaling pathways in vivo. We also summarize the updated knowledge of ncRNAs involved in common skeletal diseases and disorders, including but not limited to osteoporosis, osteoarthritis, rheumatoid arthritis, scoliosis, and intervertebral disc degeneration, by highlighting their roles established from in vivo, in vitro, and ex vivo studies.
Similar content being viewed by others

Introduction
Out of 2.85 billion nucleotides of the human genome, there are only 20,000–25,000 protein-encoding genes, which constitute less than 2% of total human genomic sequences [1]. The rest 98% of the genome was referred to as non-coding DNA, also known as “junk DNA”. The discovery of the central dogma of molecular biology states that genetic information is transcribed from DNA to RNAs and translated from messenger RNAs (mRNAs) to proteins, which facilitate all the biochemical reactions and cellular activities in cells [2]. The roles of functional RNAs, including mRNA, ribosomal RNA (rRNA), and transfer RNA (tRNA), were first identified in the process of protein synthesis. While mRNAs, which carry the genomic information for protein-encoding, are referred to as coding RNAs, non-coding RNAs (ncRNAs), such as rRNA and tRNA, with biological functions rather than encoding proteins have been known for more than sixty years [3].
In recent two decades, except for rRNA and tRNA, other functional ncRNAs, such as imprinted small nucleolar RNAs (snoRNAs), microRNAs (miRNAs), circular RNAs (circRNAs) and long non-coding RNAs (lncRNAs), have been identified and investigated. The first discoveries of small ncRNAs identified lineage defective 4 (lin-4) and lethal 7 (let-7) in C.elegans. They were reported decades ago, demonstrating that some ncRNAs are evolutionarily conserved and essential for development [4, 5]. With increasing numbers and varieties of ncRNAs identified, it became clear that functional products from the genome are not limited to proteins but also include functional ncRNAs. Some ncRNAs, such as miRNAs, snoRNAs, and circRNAs, are conserved across species, while others, including lncRNAs, normally lack evolutionary conservation. Functions of ncRNAs are various, including modifications of rRNAs, regulations of mRNA splicing and editing, altering the stability and translation of mRNAs, modulating chromatin functions, etc [6,7,8,9]. Many of these functions of ncRNAs ultimately affect gene expression in diverse cellular pathways and networks, leading to substantial alterations in fundamental cell processes, including cell proliferation, differentiation, apoptosis, and metabolism. Many ncRNAs have been confirmed to play important regulatory roles regarding gene expression in physiological processes and pathological conditions. MiRNAs, lncRNAs, circRNAs, and snoRNAs are involved in organelle development, stem cell differentiation, aging, tumorigenesis, cancers, metabolic disorders, etc. Some ncRNAs were identified as potential diagnostic biomarkers and therapeutic targets in cancers or other diseases.
The mammalian skeleton serves as a rigid internal frame of the body for supporting and protecting the soft internal organs, providing movement abilities. It is the critical site for blood cell formation. Skeletogenesis originates from migration and condensation of mesenchymal cells. The mesenchymal cells can directly differentiate into osteoblasts to form bone via a process referred to as intramembranous bone formation or into chondrocytes to form cartilage through endochondral ossification [10]. The skeletal system consists of bone, cartilage, joints, bone marrow, tendons, and ligaments. Dysregulation of gene expression during the development of the skeletal system leads to failure of bone formation, cartilage defects, mis-patterning of the skeleton, abnormalities of joint formation, etc. Loss of homeostasis, inappropriate external force, genetic defects, aging, and other factors may lead to skeletal disorders, including osteoporosis, osteoarthritis, rheumatoid arthritis, scoliosis, bone fracture, growth plate injuries, and intervertebral disc degeneration.
NcRNAs have been known to regulate the development of the nervous system, immune system, cardiovascular system, and other systems. In addition, accumulating evidence reveals that miRNAs play crucial roles in regulating key processes in skeletal development and are involved in osteoporosis, osteoarthritis, intervertebral disc degeneration, etc. In recent years, lncRNAs and circRNAs have also emerged as critical regulators of gene expression in bone and cartilage development and skeletal diseases. Interestingly, most of these regulatory ncRNAs are transcribed by RNA polymerase II (Pol II), whereas structural ncRNAs, like snRNAs, snoRNAs, and tRNAs, are primarily transcribed by polymerase III (Pol III). In this review, we integrate current information about the classification, biogenesis, and function of Pol II transcribed regulatory ncRNAs and how these ncRNAs support skeletal development through their regulation of key genes, signaling pathways, and networks in the skeleton. We also summarize the updated knowledge of ncRNAs involved in common skeletal diseases by highlighting the related in vivo studies and examining the evidence for ex-vivo and in vitro studies as well.
Classification, biogenesis, and function of ncRNAs
Though protein-coding genes constitute only around 2% of the human genome sequences, modern transcriptome sequencing technology has revealed that at least 90% of the genome is actively transcribed [11, 12]. NcRNAs consist of around 60% of the transcriptional output of human cells. Large-scale transcriptome studies of mouse genomes also showed that ncRNAs are commonly transcribed. ncRNAs are generally classified via their functional and structural similarities: structural ncRNAs and regulatory ncRNAs. Structural ncRNAs have well-established structural functions, including rRNAs, tRNAs, spliceosomal small nuclear RNAs (snRNAs), and snoRNAs. Regulatory ncRNAs consist of miRNAs, interfering RNA, piwi-RNAs, long ncRNAs, and long intergenic ncRNAs, and they play roles in regulating gene expression. In this review, we mainly focus on the regulatory ncRNAs and also touch on several structural ncRNAs that are involved in skeletal development and diseases.
miRNAs
MiRNAs are highly conserved small ncRNAs with an average of 22 nucleotides in length. The first miRNA, lin4, was reported by Ruvkun and Ambros’ labs in 1993. Lin-4 was initially characterized as a gene that regulates the temporary development of C. elegans larvae in the 1980s [13, 14]. However, after more than ten years of research, it was demonstrated by Ruvkun and Ambros that instead of a protein-coding gene, lin-4 is a small ncRNA that contains a complementary sequence to that of the 3’untranslated region (3’UTR) of lin-14 gene. Furthermore, the lin-14 protein was down-regulated at the post-transcriptional level through its 3’UTR [4, 15], indicating that lin-4 RNA regulates the expression of lin-14 protein at the post-transcriptional level. Since this revolutionary discovery, significant progress has been made in searching for miRNAs in different species and predicting and identifying their protein targets and functions.
In mammals, about half of all currently identified miRNAs are intergenic, which are regulated by their own promoters and transcribed independently. The remaining miRNAs are intragenic and mostly located in the introns of their host protein-coding genes. Most miRNAs are transcribed by RNA polymerase II [16] from DNA sequences into primary miRNAs (pri-miRNAs) transcripts. In the nucleus, pri-miRNAs undergo a processing step by the microprocessor complex, which includes RNA binding proteins (RBP), such as Drosha and DGCR8 [16,17,18]. This processing generates a hairpin precursor miRNAs (pre-miRNAs) with a length of ~ 70 nucleotides. Then, the pre-miRNAs are exported into the cytoplasm [19, 20], where they are cleaved by the RNase III enzyme Dicer, producing a miRNA duplex of approximately 22 nucleotides [21]. Finally, one strand of miRNA duplex is loaded into Argonaute proteins (AGO) to form the RNA-induced silencing complex (RISC) [22, 23]. The latest release (Version 22.1) of the miRbase, which is one of the most authoritative online resources for miRNA sequences and annotation established in 2002, contains around 48,000 mature miRNA sequences from more than 200 organisms, and in which about 2600 mature miRNAs from ~ 1900 hairpin precursors were identified in the human genome [24, 25].
Most miRNAs regulate gene expression by interacting with 3’UTR of target mRNAs to suppress protein translation and induce mRNA degradation [26, 27]. miRNAs can also mediate cell-cell communications by being secreted out of cells and transported to target cells by vesicles, including exosomes [28]. Abnormal expression of miRNAs is involved in various biological processes and pathological conditions, and miRNAs are crucial for animal development. For example, miRNA-mediated gene regulations play an essential role during skeletal development, bone and cartilage homeostasis, and the initiation and progress of skeletal disorders.
LncRNAs
LncRNAs are RNAs longer than 200 nucleotides that are not translated into proteins. They are transcribed by RNA polymerase I (Pol I), Pol II, and Pol III, and some of them are from processed introns [29]. The number of lncRNA is numerous. More than 100,000 lncRNAs have been found in the human genome [30, 31]. They are widely expressed and have specific expression patterns in physiological and pathological conditions. The biogenesis of most lncRNAs is similar to mRNAs. First, they are transcribed by polymerase II (Pol II), then undergo capping, polyadenylating, and splicing, producing transcripts similar to mRNAs. However, they are less abundant and conserved than the mRNAs [32]. Moreover, different from the cytoplasmic localization of mRNAs, a large proportion of lncRNAs are localized in the nucleus, which might be mediated by their less efficient processing rate [33, 34], nuclear retention element [35], and being tethered to chromatin [36]. A large fraction of lncRNAs is exported to the cytosol like mRNAs. Upon arrival in the cytoplasm, they are sorted into different organelles, such as mitochondria, or distributed in the cytoplasm. It is unclear how the ribosome binding property of cytoplasmic lncRNAs contributes to their functions.
The structure of lncRNAs is very flexible, making them capable of interacting with DNA, RNAs, and proteins [6]. Through these interactions, lncRNAs control gene expression at multiple levels. A large number of lncRNAs localize on chromatin, where they can interact with DNA, RNAs, or proteins, activating or silencing the transcription of the targeted chromatin [37,38,39]. One of the most prominent examples of chromatin localization lncRNAs is lncRNA XIST, which silences the gene expression of the whole X chromosome [40]. LncRNAs also regulate gene expression at the post-transcriptional and post-translational levels. They control the splicing, turnover, and translation of the mRNAs [41,42,43]. By directly binding to target proteins, lncRNAs modulate their subcellular localization, stability, and modification [44,45,46]. Benefitting from the long and flexible structure, several abundant lncRNAs function as scaffolds and assemble membrane-less organelles, e.g., lncRNA NEAT1 underlies the complex organization of paraspeckles [47], lncRNA HSAIII mediates the formation of nuclear stress bodies [47]. By virtue of these robust regulation mechanisms, lncRNAs regulate almost all biological processes, such as cell differentiation, proliferation, apoptosis, senescence, translation, and metabolism [6, 29]. LncRNAs often have a modular structure and are rich in repeats, which are increasingly shown to be relevant to their function [29]. The dysregulation of lncRNAs could lead to severe diseases, such as neuronal disorders, cancers, and bone diseases [48, 49].
CircRNAs
CircRNAs are a large class of widespread covalently closed single-stranded RNAs with tissue-specific and cell-specific expression patterns [50, 51]. More than 420,000 circRNAs have been identified in the human genome [52]. They are mainly produced from the back-splicing of the exons of mRNA precursors, with lengths varying from 100 nt to more than 4kb [53]. The intronic complementary sequences (ICSs) in introns flanking the circle-forming exons promote back-splicing by forming the transient pairs [54,55,56], which is mediated by the binding of RNA-binding proteins (RBPs) [57, 58]. Notably, most ICSs are Alu elements in humans. A small subset of circRNAs is also generated from the debranching failure of the intron lariats [59, 60]. Compared to linear RNAs, circRNAs prefer to localize in the cytoplasm [61,62,63], have lower expression levels [63,64,65], and are more stable [66].
circRNAs can also be divided into coding and non-coding circRNAs, like linear RNAs. Similar to lncRNAs, non-coding circRNAs exert their functions by interacting with DNA, RNA, and proteins. They can regulate gene transcription by directly binding to DNA. For example, CircRNA Ci-ankrd52 competes with the linear RNA cognates to form an R-loop with the second intron of the ANKRD52 locus. It promotes the transcriptional elongation of the linear RNA cognates [67]. Non-coding circRNAs can act as miRNA or protein sponges to regulate gene expression or signaling transduction. circRNA CDR1as contains 73 conserved miR-7 binding sites. It inhibits miR-7 by serving as a miR-7 binding platform [63, 68]. circRNA cia-cGAS is a circRNA antagonist for cGAS. It shows a more substantial binding capacity to cGAS than self-DNA. Thus, it inhibits the activation of cGAS by preventing it from binding to self-DNA [69]. Non-coding circRNAs can also form functional circRNP complexes to modulate signaling pathways, e.g., mitochondrial abundant circRNA SCAR forms an RNP complex with ATP5B, a regulator of the mitochondrial permeability transition pore (mPTP) complex. CAR-ATP5B RNP blocks the mPTP opening and inhibits the output of the mitochondrial reactive oxygen species (mROS) [70]. circVAMP3 promotes the formation of stress granules (SGs) by interacting with G3BP1 and CAPRIN1 [71].
Coding circRNAs are similar to linear mRNAs that function by generating proteins. Since the lack of a 5’ cap and 3’ poly (A) tail, circRNAs can only use the cap-independent translation elements, such as the embedded internal ribosome entry sites (IRESs), to initiate their translation [72,73,74]. N6-methyladenosine (m6A) modification can also drive the translation initiation of circRNAs [75]. Polysome profiling combined with mass spectrometry suggests that hundreds of circRNAs undergo m6A-driven translation [75, 76]. The cap-independent translation in normal conditions is inefficient, resulting in a low abundance of proteins translated from the circRNAs [72, 77]. Intriguingly, stress conditions significantly promote cap-independent translation efficiency [72, 75, 77], highlighting the regulatory roles of coding circRNA-derived proteins.
The stable advantage of research on circRNAs encourages people to develop RNA circle-based therapy technologies. RNA circles are engineered to aptamers [78, 79], antisense RNAs [80], miRNA and protein sponges [81,82,83,84], and protein synthesized templates [85], which show superior in their activity to their linear counterparts, to fulfill therapeutic purposes. In summary, circRNAs are essential regulators of physiological and pathological processes, and the accumulating knowledge of circRNAs helps identify novel circRNA therapeutic targets and biomarkers and develop novel therapeutic strategies.
Other ncRNAs
There is another group of ncRNAs that are around 70 to 200 nucleotides (nts) long and are transcribed by RNA polymerase III (Pol III) [86]. Examples of these classic ncRNAs include transfer RNAs (tRNAs), 5 S ribosomal RNA (5S rRNA), and U6 small nuclear RNA (U6 snRNA). These ncRNAs play crucial roles in cellular metabolism, and their varying expression is hard to understand. As a result, it has been generally considered that Pol III-transcribed genes are expressed constantly. Mature tRNAs have high and mostly constant expression levels due to extensive nucleotide modifications and secondary structure. Similarly, 5 S rRNA and U6 snRNA are abundant and expressed almost constantly. The Pol III transcription products consist of short, highly structured ncRNA and plays a crucial role in various cellular activities such as nuclear gene regulation, splicing, RNA maturation and stability, cytoplasmic protein targeting, and translation [87]. In higher eukaryotes, the Pol III transcriptome has expanded to include newly evolved ncRNA species that regulate autophagy, immune signaling cascades, and translation [88]. Dysregulation of Pol III transcription and dysfunction of ncRNA species are frequently observed in human diseases, such as severe viral infection outcomes, autoimmunity, and tumor progression.
Complex lncRNA/circRNA-miRNA post-transcriptional regulatory network
LncRNAs, circRNAs, and miRNAs are the most critical regulatory ncRNAs in cells. The binding of lncRNAs and circRNAs by miRNAs connects these three regulatory pathways and forms a post-transcriptional regulatory network. LncRNA/circRNA-miRNA regulation network seems to be unidirectional, in which lncRNAs and circRNAs inhibit miRNA activity, delivering the activation signals to mRNAs. However, RNA-RNA interactome studies identified tens of thousands of miRNA-lncRNA and miRNA-circRNA interaction pairs [89]. In these interaction pairs, one miRNA can target multiple lncRNA/circRNA targets, reminiscent of multiple mRNA targets of miRNAs. The RNA-RNA interactome study offers the possibility that miRNAs can regulate the function of lncRNAs/circRNAs, which allows the transfer of genetic information flow from miRNAs to lncRNAs/circRNAs. In addition, the binding of miRNAs in coding circRNAs may also inhibit their translation, which is a future direction worth exploring. Therefore, the lncRNA/circRNA-miRNA may form a complex regulatory network whose regulatory role is emerging (Fig. 1).

Biogenesis and functions of circRNAs, miRNAs, and lncRNAs. The precursors of most circRNAs, miRNAs, and lncRNAs are transcribed by polymerase II (Pol II) in a manner similar to that of linear mRNAs. During transcription, most human genes undergo competition between linear splicing and backsplicing of exons. Backsplicing is often favored due to factors, such as long flanking introns, inverted repeat elements (such as Alu), and RBPs [53]. The close proximity of a downstream splice-donor site (SD) with an upstream splice-acceptor site (SA) is achieved through the base pairing of inverted repeat elements or the dimerization of RBPs, leading to backsplicing. This process involves an upstream branch point (BP) attacking a downstream SD site, which in turn attacks an upstream SA site, ultimately resulting in the formation of exon-intron circRNAs or exonic circRNAs. CircRNAs have been shown to modulate cellular signaling transduction by interacting with signaling proteins, inhibiting miRNA functions, and serving as a template for the production of functional proteins. Within the nucleus, primary miRNA transcripts (pri-mRNAs) undergo cleavage mediated by the microprocessor complex Drosha–DGCR8, generating precursor miRNAs (pre-miRNAs) [16]. These pre-miRNAs are subsequently transported to the cytoplasm by exportin 5 [19, 20]. Once in the cytoplasm, pre-miRNAs undergo further cleavage by the enzyme Dicer, resulting in the formation of miRNA duplexes. These duplexes then associate with AGO proteins, thereby assembling the RNA-induced silencing complex (RISC). Once loaded onto RISC, miRNAs may undergo further processing, ultimately leading to their engagement with target messenger RNA (mRNA). Such interactions may result in mRNA destabilization or translational repression, representing key steps in post-transcriptional gene regulation. LncRNAs are transcribed and processed like mRNAs. However, many lncRNAs are retained in the nucleus to regulate the mRNA metabolism or form subnuclear domains. In the cytoplasm, lncRNA also modulates cellular signaling transduction by interacting with signaling proteins, inhibiting miRNA functions, and serving as a template for the production of functional peptides [6]
ncRNAs in skeletal development
Small ncRNAs are essential for regulating the embryonic development of vertebrates. Deleting the critical miRNA-biogenesis enzyme Dicer in zebrafish and mice results in a lack of accumulation of miRNAs, leading to early embryonic lethality [90, 91]. Removal of Dicer in the limb bud mesenchymal cells by Prx1-Cre caused the developmental delay of limbs without obviously affecting their shape, suggesting Dicer is required for morphogenesis but not patterning of the vertebrate limbs [92].
Regulation of ncRNAs in endochondral ossification
Endochondral ossification is a developmental process that forms long bones in skeletal systems [93]. It begins with the condensations of mesenchymal stem cells, which differentiate into chondrocytes expressing transcription factor Sox9, extracellular matrix protein Col2a1, and polyproteoglycans (Aggrecan) to form cartilage templates. Chondrocytes continue to expand the cartilage template through continuous proliferation. Then, the cells in the middle of the cartilage template stop proliferating and differentiate into hypertrophic chondrocytes, expressing the extracellular matrix type X collagen encoded by the Col10a1 gene. As blood vessels begin to invade the cartilage template and are accompanied by the calcification of hypertrophic chondrocytes, hypertrophic chondrocytes are subsequently replaced by osteogenesis and form a primary ossification center. The growth plate is a temporary cartilage tissue located near both ends of the long bone between the primary and secondary ossification center. The proliferation and differentiation of growth plate chondrocytes contribute to the lengthening of the long bones.
Dicer-related signaling pathway plays an important role in the regulation of skeletal development and growth in mice. Conditionally deleting Dicer in mouse growth plates led to the lethality around weaning time with skeletal growth defects caused by a reduction of proliferation and accelerated terminal differentiation of chondrocytes [94]. Specifical deletion of Dicer in mature osteoblasts by the Osteocalcin-Cre increased bone mass in adult mice from 2 months to 8 months in long bones and vertebrae [95].
Among hundreds of miRNAs, miR-140 is one of the most extensively studied miRNAs that are closely associated with cartilage development. miR-140 is believed to be expressed explicitly in cartilage but not in other tissues based on whole-mount in situ hybridization data in zebrafish and mouse embryos [96, 97]. MiR-140 is highly expressed in proliferating chondrocytes and some pre-hypertrophic chondrocytes but not in hypertrophic chondrocytes in post-natal growth plates in mice (Fig. 2). Global knockout of miR-140 in mice caused shortened body length and tail length with a slightly shortened proliferation zone and proportionally expanded resting zone of the growth plate [98]. Interestingly, mice with both miR-140 deficiency and suppression of let 7 by overexpression of Lin28a led to dramatic skeletal growth defects. In contrast, overexpression of Lin28 only results in a mild abnormality of cartilage and bone in mice, suggesting lin-7 and miR-140 can work coordinately to regulate cartilage development in vivo [99]. MiR-140 was reported to regulate palatogenesis by targeting 3’UTR of Pdgf-receptor alpha gene and down-regulating Pdgfa protein to inhibit the accumulation of cranial neural crest cells to the oral ectoderm, and its deficiency caused an altered shape of the palate in zebrafish [100]. A single nucleotide mutation in the seed region of pre-miR-140 was reported to generate a gain of functions of miR-140-5p and to be associated with autosomal dominant human skeletal dysplasia characterized by disproportionate short stature with smaller limbs, which is the first reported human disease caused by the gain-of-function mutations in a highly conserved miRNA [101]. Genetic evidence in zebrafish shows that Sox9 is upstream of miR-140 to regulate chondrogenesis [102]. However, the direct target genes and signaling pathways regulated by miR-140 controlling the development of growth plates remain to be determined in vivo.

miRNAs regulating growth plate development. MiR-140 is expressed in the proliferating chondrocytes regulated by transcription factor Sox9. MiR-140 coordinately functions with let7, which is regulated by Lin28, to regulate chondrocyte proliferation in the growth plate. miR-1 is mainly expressed in the hypertrophic chondrocytes of growth plates in mice. miR-1 may regulate hedgehog signaling in hypertrophic chondrocytes
miR-1 is detected by in situ hybridization to be highly expressed in the hypertrophic chondrocytes compared to that in proliferating chondrocytes in pre-natal chicken growth plates [103]. Transgenic mice overexpressing miR-1 in resting and proliferating chondrocytes in postnatal growth plates resulted in smaller stature with shortened limbs by decreasing the proliferation rate of growth plate chondrocytes, disrupting their hypertrophic differentiation and inhibiting the apoptosis of chondrocytes in lower hypertrophic zone [104]. Hedgehog signaling is significantly decreased in the pre-hypertrophic chondrocytes of growth plates in the miR1-overexpressing mice. However, the direct target evidence has not been reported in vivo.
MiR-17 ~ 92 cluster is a classic polycistronic miRNA encoded by the MIR17HG gene (also referred to as Mirc1 gene) containing miR-17, miR-20a, miR-18a, miR-19a, miR-19b-1, and miR-92a-1. Haploinsufficiency of miR-17 ~ 92 leads to a human autosomal dominant syndrome, referred to as Feingold syndrome, characterized by microcephaly, short stature, and severe digital abnormalities (Fig. 3). Homozygous deletion of the miR-17 ~ 92 cluster leads to prenatal lethality in mice with a severe delay of bone formation mimicking the defects in Feingold syndrome. Transgenic mice lacking one allele of the miR-17 ~ 92 cluster were viable but with significantly smaller stature than wild-type littermates [105, 106]. Different miRNAs in this cluster were demonstrated to functionally cooperate to control the patterning and development of the skeleton in vertebrates.

miRNAs controlling the proliferation of mesenchymal stem cells. Mutations of the miR-17 ~ 92 cluster are associated with human Feingold Syndrome. miR-17 ~ 92 inhibited TGFβ signaling, thus suppressing the proliferation of skeletal mesenchymal progenitors
MiR-322 is critical for the development and growth of tracheal cartilage. Lacking miR-322 in chondrocytes results in the upregulation of MEK1 protein, thus inhibiting the growth of tracheal cartilage, leading to respiration failure in mice [107]. Since lncRNA and circRNA have been identified only in recent years, there have been few reported mouse genetic studies on lncRNA or circRNA to show their roles in cartilage development in vivo. miR-34 was demonstrated to regulate bone formation by targeting SATB2 and inhibiting the proliferation and differentiation of osteoblasts in mice [108].
Regulation of ncRNAs in intramembrane ossification
Several miRNAs associated with intramembrane bone formation in vivo have been reported. Conditional deletion of Dicer in Osx-positive cells in mice resulted in a delay in suture closure time and a disorder of calvarial morphology in the skull development [109]. Specifically, deleting Dicer in mature osteoblasts by Osteocalcin-Cre did not change bone mass in the calvariae bone. Heterozygous miR-17 ~ 92 in humans was demonstrated to lead to microcephaly, which is one of the typical phenotypes of Feingold syndrome [105]. Lacking miR-92a in zebrafish led to obvious defects in craniofacial skeleton development. MiR-92a is highly expressed in the chondrogenic progenitor in zebrafish and regulates pharyngeal cartilage formation by directly targeting the noggin3-modulated Bmp signaling pathway [110]. Nardocci et al. [111] used mouse preosteoblastic cells and induced them to differentiate into osteoblasts. They performed RNA sequencing and identified lncRNA candidates that showed differential expression patterns during osteoblast differentiation. LncRNA-1 was found to be upregulated during osteogenesis and downregulated during myogenesis. Knockdown of lncRNA-1 expression inhibited osteogenic differentiation in primary mouse preosteoblasts, suggesting a new regulatory RNA involved in the early stages of osteogenesis.
ncRNAs in skeletal diseases
Epigenetic regulation is crucial for the appropriate development of the skeletal system. Emerging evidence indicates that ncRNAs are critical epigenetic factors that participate in bone development, homeostasis, and repair processes. Many downstream genes may be interfered with by ncRNAs and thereby alter cellular pathways and networks. Hence, changes in levels or activity of ncRNAs can impact cellular processes involved in proper skeletal development, including proliferation, metabolism, apoptosis, and differentiation. As epigenetic modulators are critically involved in the control of gene expression, ncRNAs are involved in the pathogenesis of a variety of skeletal diseases, such as osteoarthritis, osteoporosis, intervertebral disc degeneration, and rheumatoid arthritis.
ncRNAs in intervertebral disk degeneration
Intervertebral disc degeneration (IVDD) is a common degenerative musculoskeletal condition associated with many elements, including age, genetics, mechanics, and lifestyle [112]. Low back pain and reduced lumbar spine support are typical symptoms of IVDD, remarkably affecting the quality of life of patients and aggravating social burdens globally [113]. The intervertebral disc consists of the inner nucleus pulposus (NP) with the annulus fibrosus around it. Nucleus pulposus cells (NPCs) are the major type of cells residing in the NP, playing an essential role in maintaining disc health [114]. Excessive apoptosis of NPCs and degradation of their ECM result in IVDD.
Emerging evidence demonstrates that non‑coding RNAs are linked to the pathogenesis of IVDD [115, 116]. A number of aging-associated diseases are accompanied by epigenetic alterations [117]. NORAD is a lncRNA that regulates DNA damage and maintains genome integrity during cell growth and aging [83]. Li et al. [118] found that NORAD was significantly reduced as the development of NPC senescence increased, while elevating it alleviated NPC senescence (Fig. 4). They constructed NORAD global knockout mice and found that KO mice manifested premature aging with pronounced kyphosis at the age of 10 weeks. Similarly, the radiographic and histological analyses indicated that IVDD occurred in NORAD−/− mice at the age of 8 weeks, characterized by worse MRI manifestation, collapsed disc space, narrowed endplates height, and impaired cellularity of the NP. Furthermore, degeneration of NP and IVDD was more obvious in KO mice than in WT mice in an IVDD mouse model, suggesting a pivotal role of NORAD in the pathogenesis of IVDD. Mechanistically, NORAD was involved in m6A modification owing to increased WTAP expression, which was correlated to the trimethylation of H3K4. NORAD sequestered PUM1/2 and ameliorated the pro-decay of NPCs, and the role of NORAD was impaired in senescent NPCs by elevating m6A-mediated decay. In addition, PUM1/2 could target and degrade transcripts of E2F3, an essential factor of the cell cycle and cell proliferation, resulting in NPC senescence. Interruption of NORAD m6A modification could serve as an underlying therapeutic target of IVDD.

NcRNAs associated with IVDD. LncRNA NORAD regulates m6A modifications to regulate NP cells senescence. Inflammation factors are demonstrated to trigger the expression of miR-146a in NP cells to inhibit IVDD development
MiR-146a has been broadly investigated in the skeletal diseases [119,120,121,122,123]. Besides osteoarthritis and rheumatoid arthritis, it also seems to be correlated to the pathogenesis of IVDD. Gu et al. cultured disc segments from miR-146a−/− mice and WT mice under IL-1 challenge ex vivo [124]. IL-1 induced proteoglycan, MMP-13, and ADAMTS-5 expression in the disc organ cultures both in the WT group and KO group, which are correlated to progressive degenerative diseases, such as OA and IVDD. However, depletion of miR-146a resulted in more profound effects, indicating that stimulation of miR-146a might protect from IVDD and inflammation. In vivo experiments are required to verify the above results.
Numerous studies have demonstrated that ncRNAs have critical functions in modulating IVD development (as discussed in recently published review articles [115, 125]). Besides, some ncRNAs, including miR-15a-5p [126], miR-19b-3p [127], miR-25 [128], miR-26a-5p [129], miR-105-5p [130], miR-130b [131], miR-145-5p [132], miR-195 [133], miR-345-3p [134], miR-495-3p [135], miR-499a-5p [136], miR-502 [137], miR-660 [138], miR-2355-5p [139], miR-4478 [140], circKIF18A [141], hsa_circ_0083756 [142], hsa_circ_0059955 [143], circSPG21 [144], circPKNOX1 [145], circ-FAM169A [146], circERCC2 [147], circ-4099 [148], lncRNA XIST [149], and LncRNA NR2F1-AS1 [132] are revealed to be differently expressed in NP tissues and cells from IVDD patients. However, the investigation into the different mechanisms of IVDD in genetic animal models is extremely limited. Therefore, exploring the diverse mechanisms participating in NPC senescence is crucial for investigating IVDD, which will facilitate the development of novel therapeutic strategies.
ncRNAs in rheumatoid arthritis
Rheumatoid arthritis (RA) is a systemic inflammatory autoimmune disease without a clear etiology. RA is characterized by chronic, destructive arthritis due to multiple changes in the adaptive immune system and innate immune system [150]. T cells, B cells, osteoclasts, and many cytokines are involved in the pathogenesis of RA [151]. In addition, ncRNAs have been shown to exert powerful regulatory functions in the immune response, including the regulation of lymphocytes and dendritic cells.
MiR-155 is a widely investigated miRNA in autoimmune diseases, including RA. In RA patients, the expression of miR-155 was dramatically elevated in the peripheral blood mononuclear cells compared with that in the healthy counterparts [152]. MiR-155 globally knockout mice showed defective B cell immunity and aberrant production of pro-inflammatory cytokines, which were strongly correlated to the pathogenesis of RA [153]. In a collagen-induced RA (CIA) model, miR-115−/− mice developed much more moderate typical clinical and histological signs than WT mice [154]. This might be due to fewer anti-collagen antibodies and antigen-specific T cells in KO mice. In addition, the levels of interleukin-6 (IL-6), IL-17, and IL-22 were reduced, indicating that Th17 polarization of KO mice was hampered. Moreover, miR-155 could directly target human Src homology 2-containing inositol phosphatase-1 (SHIP-1) for degradation. Expression of SHIP-1, a potent antagonist of several inflammatory pathways, was markedly decreased in RA synovial CD14+ cells. On the contrary, suppression of miR-155 with antagomir dramatically elevated the expression level of SHIP-1 mRNA in synovial fluid CD14+ cells from RA patients. Chen et al. found that the bone marrow-derived dendritic cells (BMDCs) obtained from the KO mice generated fewer pro-inflammatory cytokines in the lipopolysaccharide (LPS) induced inflammation model [155]. Collectively, these findings suggest that miR-155 is critical for the pathogenesis of RA, and it might benefit the development of novel therapeutic strategies.
MiR-146a is a broadly investigated anti-inflammatory miRNA [156]. MiR-146a global knockout mice presented aggravated joint inflammation, while intravenous injection of miR-146a mimics prevented joint destruction in an RA model [119, 121]. Ammari et al. found that the expression of miR-146a was decreased in the Ly6Chigh monocytes of arthritic mice and CD14+ CD16− monocytes in RA patients. They administrated miR-146a mimics targeting Ly6Chigh monocytes through DMAPAP/DOPE cationic liposome. The administration reduced bone absorption and bone erosion in arthritic joints in KO mice. Therefore, overexpression of miR-146a in Ly6Chigh monocytes could alleviate joint destruction and avoid inflammation in RA simultaneously.
MiR-204 and miR-211, two homologous miRNAs with the same target genes, were reported to protect against OA progression [157]. Wang and coworkers continued to determine the effects of two miRNAs on synovial hyperplasia and inflammation [158]. First, they found that expression of miR204/211 was dramatically decreased in synoviocytes of CIA mice, affecting cell migration, apoptosis, proliferation, and inflammatory responses. Based on bioinformatics analysis and subsequent validation experiments, Ssrp1 was considered to be a target gene of miR-204 and miR-211. Next, they constructed miR204/211 double knockout mice and performed the CIA model. Double knockout mice presented higher Ssrp1 levels with stronger synovial hyperplasia and inflammation, which could be restored by injecting AAV- shSsrp1 intra-articularly. These results indicate that miR204/211 is essential in the progression of RA, which might provide a novel treatment strategy for RA.
Nuclear-enriched abundant transcript 1 (NEAT1) is a lncRNA that was reported to play a vital role in the progression of RA [159, 160]. The synovial tissue obtained from both mice and RA patients expressed higher NEAT1 [159]. Depleting NEAT1 with lenti-sh-NEAT1 in mice inhibited the infiltration of both CD4+ T cells and macrophages into synovial tissue [159, 160]. Moreover, it reduced the expression of inflammatory factors in peripheral blood and decreased the incidence/severity of RA in CIA mice [159]. Mechanistically, NEAT1 was activated by p-p65 and then promoted the p300/CBP/IL-18 signaling pathway, which eventually aggravated RA [159]. NEAT1 could also stimulate CD4+ T cells to differentiate into Th17 cells by enhancing the STAT3 expression [160].
To date, nearly 100 ncRNAs have been reported to be strongly associated with the pathogenesis of RA (presented in a recently published review article [161]), which still needs to be studied and validated further by genetic models. Taken together, the pivotal roles of ncRNAs as biomarkers and targets for RA require additional research.
ncRNAs in osteoporosis
Bone metabolism is a complicated process that includes anabolism and catabolism. Osteoblasts and osteoclasts play essential roles in maintaining the homeostasis of the formation and absorption of the bones. Once bone formation decreases and bone absorption increases, osteoporosis occurs and aggravates the fracture risk. Emerging evidence indicates that ncRNAs are closely involved in the incidence and development of osteoporosis. Jin et al. [162] found that more than 300 ncRNAs were aberrant in patients with postmenopausal osteoporosis, and regulating these related ncRNAs might be helpful in treating osteoporosis. In this section, we will review those pivotal ncRNAs whose roles have been verified through genetic animal models.
Bone formation
Positive regulators
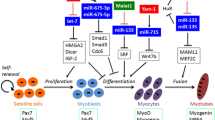
Osteoblasts, mainly differentiated from mesenchymal stem/stromal cells, regulate bone homeostasis by facilitating bone formation. Some ncRNAs have been investigated to facilitate osteoblastic differentiation in osteoporosis. MiR-143 is highly expressed in osteoblasts, and overexpression of it was reported to promote osteoblast differentiation [163]. Osteoblastic differentiation of miR-143−/− mice was markedly inhibited. Using mRNA-sequencing combined with target prediction and luciferase reporter experiment, miR-143 was found to directly target the 3’-UTR of HDAC7 and inhibit HDAC7 expression in osteoblasts. HDAC7-siRNA was injected into the bone marrow of miR-143 KO mice, and the symptoms caused by miR-143 depletion were significantly ameliorated. On the contrary, administration of agomiR-143 intravenously accelerated osteogenesis and inhibited bone loss in an aging-induced osteoporosis mice model. MiR-497 ~ 195 cluster tended to be expressed in CD31hi Emcnhi endothelial cells (a specific bone vessel subtype), which was decreased during aging [164]. The endothelial-specific knockout of miR-497 ~ 195 cluster using miR-497 ~ 195fl/fl; Cdh5 (PAC)-Cre mice manifested fewer CD31hiEmcnhi vessels and lower bone mass while intravenous injection of aptamer-agomiR-195 promoted bone formation in aged mice. Mechanistically, the miR-497 ~ 195 cluster maintained the Notch activity via targeting F-box and WD-40 domain protein (Fbxw7) and maintained the HIF-1α stability via targeting Prolyl 4-hydroxylase possessing a transmembrane domain (P4HTM) in the endothelial cells. LncRNA Bmncr determined the fate of aging BMSCs by regulating the osteogenic niche in the bone marrow [165]. Bmncr−/− mice presented redundant age-associated bone loss, while transgenic overexpression of Bmncr increased the bone mass. Bmncr served as a scaffold to promote the interaction between TAZ and ABL and then stimulated the assembly of the TAZ-RUNX2 and TAZ-PPARG transcriptional complex, resulting in bone formation. Another lncRNA, Crnde, also has a function of osteoblast proliferation and differentiation [166]. Crnde knockout mice possessed impaired osteoblast proliferation/differentiation and reduced bone mass phenotypes. This might be caused by decreased Alp, Runx2, and Osx expression via inhibiting the Wnt/β-catenin pathway, which was responsible for cell proliferation (Fig. 5).

A diagram briefly summarizing the ncRNAs that are involved in the dynamic balance between osteogenesis and osteoclastogenesis in transgenic mice models. The pathological progress of OP is due to the imbalance between bone formation and resorption mediated by osteoblasts and osteoclasts, respectively. Dysregulated ncRNAs participate in the development of OP
Negative regulators
Some other ncRNAs have been investigated to be negative regulatory factors of osteogenesis. Due to osteoporosis being an aging-related disease, Li and coworkers performed miRNA microarray analysis to compare bone marrow stromal cells (BMSCs) in young mice with those in aged mice and found that miR-188 expression was much higher in aged mice [167]. Global knockout of miR-188 or administration of aptamer-antagomiR-188 into bone marrow alleviated age-related bone loss and promoted adipogenesis. However, overexpression of miR-188 in osterix+ osteoprogenitors or BMSCs resulted in aggravated bone loss and adipogenesis of BMSCs. Moreover, HDAC9 and RICTOR were validated as the direct targets of miR-188. MiR-146a−/− mice have an enhanced osteoblastic activity and bone formation rate [122, 123]. MiR-146a-5p expression was increased in the bone tissue of both aging female mice and PMOP patients [122]. In addition, miR-146a−/− reversed bone loss while aging in female mice. Meanwhile, the multiplication of MSC was faster, and apoptosis of MSC was weaker in miR-146a−/− mice with higher expression of osteoblastic markers, including Alp and Ocn. Using target prediction, luciferase reporter experiment, and RNA immunoprecipitation assays in MC3T3-E1 cells, Zheng et al. [122] found Sirt1 could bind with miR-146a-5p in its 3’-UTR. Furthermore, the protein expression of Sirt1 was significantly elevated in miR-146a global KO mice. Thus, miR-146a was positively associated with age, and miR-146a ablation could prevent mice from OP. LncRNA Neat1 acted as a vital ncRNA in osteoblastic differentiation in mice subjected to mechanical loading [168]. Neat1−/− mice showed impaired bone structure and bone loss. In addition, Neat1 depletion inhibited bone formation induced by mechanical loading and blunted mechanical unloading-induced bone loss. Mechanistically, paraspeckles accelerated the nuclear retention of Smurf1 suppressed the translation, and ultimately suppressed ubiquitination-mediated degradation of Runx2, one of the targets of Smurf1.
Postmenopausal osteoporosis (PMOP) is a frequent bone disorder. Xu et al. [169] found miR-143 and miR-145 were potential treatment targets for PMOP, and overexpressing miR-143/145 hampered self-renewing and osteoblastic differentiation of BMSCs. On the contrary, miR-143/145 global knockout or antagomiR-143/145 significantly ameliorated estrogen-deficient OP in female mice. Specifically, cytoplasmic miR-143/145 and lncRNA MIR143HG, controlled by ERβ, cooperatively regulated pluripotency genes translation via canonical ceRNA pathway, and MIR143HG cooperated with miR-143 to nuclear translocation for co-activation of SOX2 transcription via opening promoter chromatin. In an ovariectomized (OVX) mouse model, miR-185 knockout ameliorated bone loss in an estrogen-deficient OP model, while miR-185 agomir reversed the increased bone formation in miR-185−/− mice after OVX [170]. Osteoblasts and MSCs obtained from miR-185−/− mice displayed enhanced bone formation. Mechanistically, miR-185 ablation activated the BMP/Smad pathway to regulate Bgn, the knockdown of which reversed the increase of Alp, Bmp2, and p-Smad1/5/8. MiR-338 cluster (including miR-338-3p and miR-3065-5p) was highly enriched in PMOP patients and OVX-induced mice [171]. Either miR-338-3p or miR-3065-5p inhibitor could dramatically alleviate the symptoms of OP in OVX mice by suppressing the expression and secretion of the miR-338 cluster. Moreover, the deletion of miR-338 could relieve or even arrest OP progression. BMSCs obtained from the femurs of miR-338 global knockout mice expressed higher Runx2, Opn, Osx, and Ocn with a greater number of mineralization nodules after osteogenic induction. Dual luciferase assay results validated that overexpression of miR-338-3p and miR-3065-5p in MC3T3-E1 cells inhibited osteoblastic differentiation by directly targeting Runx2 and Sox4. Lu et al. [172] found that miR-451a was dramatically elevated post-OVX operation. Primary osteoblasts extracted from miR-451a−/− mice possessed the stronger proliferative ability, Alp activity, and the ability to form a mineralized matrix. The gene and protein levels of osteoblast markers were dramatically increased in osteoblasts harvested from miR-451a−/− mice. Additionally, ablation of miR-451a alleviated the bone loss in estrogen-deficient OP, which was abrogated by miR-451a agomir administration. Using database prediction and further validation of biological experiments, Bmp6 was considered a direct target of miR-451a. Table 1 summarizes the role of some ncRNAs in osteoporosis and their mechanisms through bone formation validated in the transgenic mice model.
Bone resorption
Osteoclasts are differentiated from macrophage/monocyte precursor cells in the presence of RANKL and other osteoclastogenic factors. Understanding aberrant bone resorption is critical for investigating the diagnosis, treatment, and prevention of osteoporosis. Mir-21 is the first investigated ncRNA in regulating bone resorption in mice. miR-21−/− mice showed normal skeletal structure and osteogenesis while inhibited osteoclastogenesis [173]. However, miR-21−/− mice exhibited inhibited bone resorption in tartrate-resistant acid phosphatase (TRAP) staining and C-telopeptide of type 1 collagen (CTX-1) ELISA test. Mir-21 ablation also attenuated OVX-induced osteoporosis and aging-induced osteoporosis, which was validated in osteoporosis patients. PDCD4, a regulator of osteoclast differentiation, was proved to be a direct target of miR-21 in supporting osteoclastic function. Mir-128 was regarded as an essential ncRNA in aging and inflammatory-related diseases involving osteoporosis [174]. MiR-128 was highly enriched in PMOP patients and OVX mice, which was further proved to promote osteoclastogenesis. Depletion of miR-128 in the osteoclasts (using miR-128flox/flox; LysM-Cre mice) dramatically decreased osteoclastogenesis and relieved OVX-induced bone loss, as evidenced by TRAP staining, qPCR analysis, and ELISA assay. MiR-128 promoted osteoclastogenesis by targeting Sirt1 and then activating NF-κB signaling by acetylation modifications. Xu et al. [169] demonstrated that miR-143/145 was notably increased in the serum, saliva, and BMSCs from postmenopausal women compared to premenopausal counterparts. In addition, the self-renewing and differentiation of BMSCs were impaired in postmenopausal women. Depletion of miR-143/145 or injection of antagomiR-143/145 arrested bone loss and retained bone regeneration in the estrogen-deficient OP, while agomiR-143/145 exacerbated it. That was due to extracellular vesicles from BMSCs loading miR-143/145 and being transferred to osteoclasts to affect osteoclastic activity and function by targeting Cd226 and Srgap2. Besides miR-143/145, miR-146a, miR-148a, and miR-301-b were also regarded as therapeutic targets for estrogen deficiency-induced osteoporosis. MiR-146a was elevated in OVX-induced OP mice, the deletion of which protected the mice from bone loss [123]. Osteoclast activities were hampered in the miR-146a global knockout mice subjected to OVX. Macrophage colony-stimulating factor (M-CSF) and RANKL/OPG ratio from the bone microenvironment regulated osteoclastogenesis under estrogen deficiency. Pan et al. revealed that miR-148a−/− mice showed an increased bone mass and decreased bone resorption by targeting NRP1 [175]. In an estrogen-deficient OP model and a calvarial osteolysis model, depletion of miR-148a prevented mice from redundant bone absorption, while injection of agomiR-148a intravenously or AAV-shNRP1 via bone marrow cavity accelerated pathogenesis of OP. More importantly, β-CTX secretion in PMOP patients was positively related to the expression of miR-148a. MiR-301-b was elevated in the bone tissue of either PMOP patients or OVX-induced mice [176]. Osteoclastic conditional knockout of miR-301 (miR-301-bflox/flox; LysM-Cre mice) led to reduced osteoclastogenesis and osteoclast number, exerting a strong bone protection effect. Further studies revealed that CYLD, an anti-inflammatory factor, is the direct target of miR-301-b. CYLD governed osteoclastogenesis through activating NF-κB signaling.
Investigations of osteoporosis on lncRNA or circRNA were limited in vivo. LncRNA Nron is highly enriched in bone tissue and markedly decreased in OVX mice [177]. Osteoclasts isolated from OVX mice or osteoclast-specific Nron knockout mice (Nronflox/flox; Ctsk-Cre mice) also expressed lower Nron with higher bone resorption. On the contrary, osteoclast-specific Nron overexpression transgenic mice (using Ctsk promoter) or pharmacological overexpression of Nron developed higher bone mass owing to suppressed osteoclastogenesis. Mechanistically, Nron interacted with CUL4B via its functional motif NCM2 to regulate the Erα pathway through ubiquitination in osteoclasts (Fig. 5).
In summary, numerous studies have shown that ncRNAs are recognized as key regulators of osteoporosis. In this section, we mainly reviewed some lncRNAs and miRNAs with essential roles in osteogenesis and osteoclastogenesis in vivo and their potential molecular mechanisms and translational significance. Nevertheless, there are few studies investigating the gain- or loss-of-function of circRNAs in transgenic animals until the submission of the manuscript. NcRNA-targeted genes affect different signaling pathways, which are important in the regulation of bone formation and absorption. More research is needed to study the manipulation of ncRNA expression in the future, which will establish the pathogenesis of osteoporosis and provide a potential therapeutic strategy for osteoporosis. An in-depth understanding of the regulation of those ncRNAs will be instrumental in exploring explicit targets to diagnose and treat osteoporosis. Table 2 summarizes the role of some ncRNAs in osteoporosis and their mechanisms through bone absorption validated in the transgenic mice model.
ncRNAs in osteoarthritis
Osteoarthritis (OA) is one of the prevalent diseases triggering pain and disability, which has a profound impact on the quality of life and socio-economic burden/cost throughout the world [178]. It usually affects joints in the hips, knees, and fingers, with characteristics of cartilage destruction, subchondral bone remodeling, and osteophyte formation. The incidence rate of OA has increased in recent years, affecting over 25% of the population over 18 years of age [179]. Most patients with OA have not received adequate treatment and can only alleviate symptoms. Identifying specific biomarkers for early diagnosis, subgroup classification, stage, and disease prognosis of OA is urgent [180]. A comprehensive understanding of molecular mechanisms of pathogenic OA will help discover new biomarkers so as to prevent or delay the deterioration of the disease [179]. It is well-known that ncRNAs can act as epigenetic regulators in cartilage formation and homeostasis by mediating the proliferation, differentiation, and ECM biosynthesis of chondrocytes [180]. The abnormal expression of ncRNAs may result in ECM degradation, chondrocyte hypertrophy, and chondrocyte apoptosis and subsequently lead to OA. A growing number of in vivo studies have highlighted the vital functions of ncRNAs in maintaining cartilage homeostasis.
Positive miRNA regulators for articular cartilage homeostasis
Recent studies revealed that Sox9 could act as a critical transcription factor during cartilage formation and pathogenesis of OA [181, 182]. MiR-140 is a cartilage-specific ncRNA in mouse embryos and zebrafish, which is regulated by Sox9 [98]. MiR-140 global knockout mice showed short stature and lower body weight in the early stages [98]. However, these mice started to present OA-like pathology in the knee joints after 12 months old. Consistent with the aging-induced OA model, miR-140−/− mice also exhibited accelerated articular cartilage injury in a surgical arthritis model. In addition, in an antigen injection-induced arthritis model, cartilage-specific miR-140 overexpression transgenic mice exhibited milder OA symptoms. DNA array analysis, together with further in vitro studies and in vivo studies, validated that miR-140 directly inhibited Adamts-5 expression from postponing aggrecan degradation and OA progression. MiR-455, an intensely expressed ncRNA in human and mouse chondrocytes, could also maintain cartilage homeostasis governed by Sox9 [183, 184]. MiR-455-3p−/− mice presented accelerated cartilage degeneration at both 5 months and 12 months old [184]. Consistent with this, the knee joints of miR-455−/− mice developed OA-like pathology at 6 months of age. Administration of miR-455-3p and miR-455-5p mimics into the knee cartilage of mice subjected to DMM surgery significantly inhibited cartilage degeneration. HIF-2α, encoded by EPAS1, was validated as a target of miR-455-5p and − 3p. HIF-2α was a catabolic factor for maintaining cartilage homeostasis, knockdown of which using siRNA in vivo injection into the knee cartilage of miR-455−/− mice dramatically alleviated the cartilage degeneration.
Runx2 is a dominant regulator of chondrocyte hypertrophy and OA progression, overexpression of which leads to hypertrophic differentiation of normal chondrocytes [185, 186]. MiR-204 and miR-211 are homologous miRNAs, maintaining joint homeostasis via a Runx2-dependent manner to inhibit OA pathogenesis [157]. Global knockout of miR204/211 (miR-204fl/fl; miR-211fl/fl; CMV-Cre mice) resulted in spontaneous OA at the age of 15 weeks. Mesenchymal progenitor conditional knockout of miR204/211 (miR-204fl/fl; miR-211fl/fl; Prx1-Cre mice) presented OA-like phonotypes gradually with age. Depletion of miR-204/211 in mesenchymal tissues led to Runx2 accumulation and catabolic activities in articular cartilage degradation. Additionally, mesenchymal progenitor cells in dKO mice produced excessive mesenchymal cells by activating the Ngf/Akt pathway and contributed to synovial hyperplasia. Strikingly, these OA phenotypes could be ameliorated by intra-articular injection of AAV5-miR-204 in OA mice or knocking down Runx2 in mice (miR-204fl/fl; miR-211fl/fl; Runx2fl/+; Prx1-Cre). Table 3 summarizes the role of some ncRNAs in osteoarthritis and their mechanisms validated in the transgenic mice model.
Negative miRNA regulators for cartilage homeostasis
Utilizing Gene Expression Omnibus (GEO) datasets and further independent cohort and molecular biological validation, Wang and coworkers identified that miR-21-5p was markedly upregulated in OA patients [187]. They subsequently generated cartilage-specific miR-21-5p knockout mice by crossing miR-21-5pfl/fl and Col2a1-CreERT2 mice. The cKO mice inhibited mice spontaneous OA at the 12th month of age, and the destabilization of the medial meniscus (DMM) induced OA. Intra-articular administration of antagomiR-21-5p significantly ameliorated articular cartilage degradation in the mice DMM model, while agomiR-21-5p aggravated it. Further studies indicated that FGF18 was a direct target of miR-21-5p, which was validated in vivo that the protein level of FGF18 in cKO mice was increased. Mir-21-5p was also related to the temporomandibular joint osteoarthritis (TMJOA) [188]. In a unilateral anterior crossbite model, miR-21-5p−/− mice alleviated the progression of TMJOA, expressing less inflammatory-related genes and proteins. MiRNA target databases with further experiments validated that Spry1 was a target gene of miR-21-5p. MiR-141/200c was also found to be enhanced in OA patients identified by microarray and further independent cohort [189]. Cartilage conditional knockout of miR-141/200c using miR-141/200cfl/fl; Col2a1- CreERT2 mice alleviated either aging or surgery-induced OA. Intra-articular administration of miR-141/200c delivered by chondrocyte-specific nanoparticles execrated OA pathogenesis while corresponding miR-141/200c inhibitor reagent exhibited chondroprotective effects. Specifically, miR-141/200c targeted SIRT1, a deacetylase, thus activating IL-6/STAT3 signaling to aggravate OA development.
MiR-146a was reported to be a biomarker of OA, which was elevated in the articular cartilage of early OA patients [190] and mice subjected to DMM surgery [191]. MiR-146a−/− mice suppressed both spontaneous OA and knee destabilization-induced OA [191]. Lenti-miR-146a-mimic inhibited cartilage matrix-related Sox9 and Col2a1 expression in the primary mouse articular chondrocytes, while Lenti-miR-146a-inhibitor imparted a reverse effect. This result was validated in the DMM-induced mice model, indicating that suppressing miR-146a has a potential therapeutic effect on OA. The regulation of miR-146a on cartilage anabolism was considered as miR-146a targeted several genes in vitro and in vivo through an NF-кB-dependent signaling, including Tgif1, Camk2d, and Ppp3r2. The expression of miR-483-5p was increased either in the articular cartilage of OA patients or DMM-induced OA mice [192]. Bai group generated doxycycline-inducible miR-483 transgenic mice to investigate the pivotal role of miR-483-5p on OA [192, 193]. These mice exhibited more severe symptoms both in DMM-induced OA and aging-induced OA. Particularly, miR-483-5p was responsible for the expression of mTORC1, which was HDAC4 dependent. In addition, miR-483-5p directly targeted Matn3 and Timp2 to promote chondrocyte hypertrophy, extra-cellular matrix degradation, and cartilage angiogenesis. MiR-34a-5p was a p53-mediated ncRNA that participated in various human diseases, including OA. MiR-34a-5p was remarkably elevated in the plasma, cartilage, and synovium of OA patients and DMM-induced OA mice, which was correlated to obesity. Intra-articular administration of miR-34a-5p antisense oligonucleotide or miR-34a-5p global knockout protected articular cartilage from DMM-induced damage with or without a high-fat diet. Using RNA-sequencing with bioinformatics analysis, Pparg was predicted as a target of miR-34a-5p, which still needs to be validated further.
CircRNAs in osteoarthritis
Due to the rapid development of high-throughput RNA-sequencing methods, more than 30,000 circRNAs have been reported in the last few years [62]. The research on circRNAs in OA has also been constantly emerging. Expression of circNFKB1 [194] and circGCN1L1 [195] has been reported to be up-regulated in human chondrocytes via RNA sequencing. Knockdown of circNFKB1 inhibited ECM catabolism and promoted osteoarthritis progression through interacting with ENO1 and sustaining the NF-κB signaling pathway. Ad-circNFKB1 was further intra-articularly injected into DMM-induced OA mice to determine its role in vivo, and the results showed that circNFKB1 aggravated OA progression, as manifested by cartilage destruction and osteophyte formation. While circGCN1L1 facilitated chondrocyte apoptosis, induced inflammation in synoviocytes, and decreased the anabolism of ECM via sponging miR-330-3p and targeting TNF-α in TMJ OA. CircRNAs can also be protective for the progression of OA. For example, circSERPINE2 [196], circPDE4B [197], and circFOXO3 [198] were downregulated in OA cartilage. circSERPINE2 protected articular cartilage via targeting miR-1271 and E26 transformation-specific-related gene (ERG). It could alleviate chondrocyte apoptosis and promote anabolism of ECM through the miR-1271-ERG pathway. Furthermore, intra-articular injection of adeno-associated virus overexpressing circSERPINE2 alleviated OA phenotype in a rabbit model. CircPDE4B regulated chondrocyte cell proliferation and ECM metabolism; the circPDE4B–RIC8A axis played an important role in regulating downstream p38 MAPK signaling. circFOXO3 activated PI3K/AKT-mediated autophagy, further alleviating apoptosis of chondrocytes and promoting anabolism of the ECM. As cartilage destruction and osteophytes decreased in Lv-circFOXO3 DMM mice, the anti-OA effect of circFOXO3 in vivo was also validated. CircPARD3B was downregulated in synovial tissues in OA, and it inhibited synovial angiogenesis via targeting miR-326 and SIRT1 [199].
LncRNAs in osteoarthritis
LncRNAs do not translate into polypeptides; however, they have essential effects on regulating the expression of specific genes. During the pathogenesis of OA, many lncRNAs have been discovered to be dysregulated in cartilage or synovial fluid. The function of lncRNAs in the pathogenesis of OA has been revealed with more details in recent research. HOTAIR [200] and LINC02288 [201] were found to be upregulated in human OA cartilage, and LINC02288 promoted chondrocyte apoptosis and inflammation by targeting the miR-347a-3p/RTN axis. HOTAIR directly bound to miR-17-5p and indirectly upregulated the expression of FUT2 and exacerbated chondrocyte apoptosis and ECM degradation. HOTAIR/miR-17-5p/FUT2 axis contributed to OA progression via the Wnt/β-catenin pathway. Intra-articular injection of the lentivirus expressing HOTAIR or FUT2 showed more severe damage to the cartilage. Another study also revealed that HOTAIR [202] promoted the progression of OA by inhibiting the proliferation of chondrocytes and promoting apoptosis and ECM degradation via regulating the miR-20b/PTEN axis. On the contrary, MM2P [203] was protective for the differentiation of chondrocytes, enhanced the expression of Colla2 and Acan and promoted the secretion of proteoglycan and type II collagen in chondrocytes by inducing M2 macrophage polarization, and accelerating the delivery of M2-derived exosomal SOX9 into chondrocytes. Another lncRNA, linc-ROR [204], also promoted mesenchymal stem cells chondrogenesis and cartilage formation via regulating SOX9 expression, linc-ROR functioned as a sponge for miR-138 and miR-145. They both suppressed BMSCs chondrogenesis activity and SOX9 expression, while co-expression of linc-ROR showed a rescuing effect. Figure 6 summarizes the roles of some ncRNAs that have been proved in transgenic mice models as regulators in the molecular mechanism of OA.

Illustration of the roles of ncRNAs that are experimentally proved in transgenic mice models as key regulators in the molecular mechanism of OA. Nc RNAs contribute to many pathologic processes in OA, including (a) ECM degradation, (b) chondrocyte proliferation and differentiation, (c) autophagy and apoptosis, (d) angiogenesis, and (e) inflammatory events
Conclusions
Based on the evidence from hundreds of in vivo studies, it has been clearly demonstrated that ncRNAs play essential roles in skeletal development and diseases by regulating gene expression. The difficulty of mapping the interplay regulation networks of ncRNAs in the development and diseases of the skeletal system is identifying the critical ones among thousands of ncRNAs abundant in the skeletal tissues. Tissue-specific ncRNAs should be identified and might be critical in vivo systems. Thousands of in vitro studies are informative in terms of identifying the direct targets and related functions of each ncRNA in a specific cell type. However, the functional validation in vivo finally pins down the roles of specific ncRNAs in the physiological system.
Data availability
Not applicable.
References
International Human Genome Sequencing. Finishing the euchromatic sequence of the human genome. Nature. 2004;431(7011):931–45.
Crick F. Central dogma of molecular biology. Nature. 1970;227(5258):561–3.
Eddy SR. Non-coding RNA genes and the modern RNA world. Nat Rev Genet. 2001;2(12):919–29.
Lee RC, Feinbaum RL, Ambros V. The C. Elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75(5):843–54.
Reinhart BJ, et al. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. 2000;403(6772):901–6.
Statello L, et al. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118.
Greene J, et al. Circular RNAs: Biogenesis, function and role in Human diseases. Front Mol Biosci. 2017;4:38.
Gebert LFR, MacRae IJ. Regulation of microRNA function in animals. Nat Rev Mol Cell Biol. 2019;20(1):21–37.
Bratkovic T, Bozic J, Rogelj B. Functional diversity of small nucleolar RNAs. Nucleic Acids Res. 2020;48(4):1627–51.
Salhotra A, et al. Mechanisms of bone development and repair. Nat Rev Mol Cell Biol. 2020;21(11):696–711.
Consortium EP, et al. Expanded encyclopaedias of DNA elements in the human and mouse genomes. Nature. 2020;583(7818):699–710.
She X, et al. Shotgun sequence assembly and recent segmental duplications within the human genome. Nature. 2004;431(7011):927–30.
Horvitz HR, Sulston JE. Isolation and genetic characterization of cell-lineage mutants of the nematode Caenorhabditis elegans. Genetics. 1980;96(2):435–54.
Chalfie M, Horvitz HR, Sulston JE. Mutations that lead to reiterations in the cell lineages of C. Elegans. Cell. 1981;24(1):59–69.
Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. Elegans. Cell. 1993;75(5):855–62.
Denli AM, et al. Processing of primary microRNAs by the Microprocessor complex. Nature. 2004;432(7014):231–5.
Lee Y, et al. The nuclear RNase III Drosha initiates microRNA processing. Nature. 2003;425(6956):415–9.
Han J, et al. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004;18(24):3016–27.
Bohnsack MT, Czaplinski K, Gorlich D. Exportin 5 is a RanGTP-dependent dsRNA-binding protein that mediates nuclear export of pre-miRNAs. RNA. 2004;10(2):185–91.
Lund E, et al. Nuclear export of microRNA precursors. Science. 2004;303(5654):95–8.
Bernstein E, et al. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409(6818):363–6.
Hammond SM, et al. Argonaute2, a link between genetic and biochemical analyses of RNAi. Science. 2001;293(5532):1146–50.
Mourelatos Z, et al. miRNPs: a novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev. 2002;16(6):720–8.
Kozomara A, Birgaoanu M, Griffiths-Jones S. miRBase: from microRNA sequences to function. Nucleic Acids Res. 2019;47(D1):D155–62.
Alles J, et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019;47(7):3353–64.
Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014;15(8):509–24.
Treiber T, Treiber N, Meister G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat Rev Mol Cell Biol. 2019;20(1):5–20.
O’Brien J, et al. Overview of MicroRNA Biogenesis, mechanisms of actions, and circulation. Front Endocrinol (Lausanne). 2018;9:402.
Mattick JS, et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat Rev Mol Cell Biol. 2023;24(6):430–47.
Uszczynska-Ratajczak B, et al. Towards a complete map of the human long non-coding RNA transcriptome. Nat Rev Genet. 2018;19(9):535–48.
Fang S, et al. NONCODEV5: a comprehensive annotation database for long non-coding RNAs. Nucleic Acids Res. 2018;46(D1):D308–14.
Ulitsky I, Bartel DP. lincRNAs: genomics, evolution, and mechanisms. Cell. 2013;154(1):26–46.
Mele M, et al. Chromatin environment, transcriptional regulation, and splicing distinguish lincRNAs and mRNAs. Genome Res. 2017;27(1):27–37.
Zuckerman B, Ulitsky I. Predictive models of subcellular localization of long RNAs. RNA. 2019;25(5):557–72.
Azam S, et al. Nuclear retention element recruits U1 snRNP components to restrain spliced lncRNAs in the nucleus. RNA Biol. 2019;16(8):1001–9.
Pintacuda G, et al. hnRNPK recruits PCGF3/5-PRC1 to the Xist RNA B-Repeat to establish polycomb-mediated chromosomal silencing. Mol Cell. 2017;68(5):955–69. e10.
Chu C, et al. Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions. Mol Cell. 2011;44(4):667–78.
Bonetti A, et al. RADICL-seq identifies general and cell type-specific principles of genome-wide RNA-chromatin interactions. Nat Commun. 2020;11(1):1018.
Li X, et al. GRID-seq reveals the global RNA-chromatin interactome. Nat Biotechnol. 2017;35(10):940–50.
Loda A, Heard E. Xist RNA in action: past, present, and future. PLoS Genet. 2019;15(9):e1008333.
Engreitz JM, et al. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature. 2016;539(7629):452–5.
Yang L, et al. ncRNA- and Pc2 methylation-dependent gene relocation between nuclear structures mediates gene activation programs. Cell. 2011;147(4):773–88.
Vendramin R, et al. SAMMSON fosters cancer cell fitness by concertedly enhancing mitochondrial and cytosolic translation. Nat Struct Mol Biol. 2018;25(11):1035–46.
Xiang JF, et al. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014;24(5):513–31.
Noh JH, et al. HuR and GRSF1 modulate the nuclear export and mitochondrial localization of the lncRNA RMRP. Genes Dev. 2016;30(10):1224–39.
Latos PA, et al. Airn transcriptional overlap, but not its lncRNA products, induces imprinted Igf2r silencing. Science. 2012;338(6113):1469–72.
West JA, et al. Structural, super-resolution microscopy analysis of paraspeckle nuclear body organization. J Cell Biol. 2016;214(7):817–30.
Huarte M. The emerging role of lncRNAs in cancer. Nat Med. 2015;21(11):1253–61.
Silva AM, et al. Long noncoding RNAs: a missing link in osteoporosis. Bone Res. 2019;7:10.
Liu CX, Chen LL. Circular RNAs: characterization, cellular roles, and applications. Cell. 2022;185(13):2390.
Kristensen LS, et al. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20(11):675–91.
Wu W, Ji P, Zhao F. CircAtlas: an integrated resource of one million highly accurate circular RNAs from 1070 vertebrate transcriptomes. Genome Biol. 2020;21(1):101.
Szabo L, Salzman J. Detecting circular RNAs: bioinformatic and experimental challenges. Nat Rev Genet. 2016;17(11):679–92.
Aktas T, et al. DHX9 suppresses RNA processing defects originating from the Alu invasion of the human genome. Nature. 2017;544(7648):115–9.
Zhang XO, et al. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016;26(9):1277–87.
Jeck WR, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. 2013;19(2):141–57.
Errichelli L, et al. FUS affects circular RNA expression in murine embryonic stem cell-derived motor neurons. Nat Commun. 2017;8:14741.
Li X, et al. Coordinated circRNA Biogenesis and function with NF90/NF110 in viral infection. Mol Cell. 2017;67(2):214–e2277.
Zhang Y, et al. Circular intronic long noncoding RNAs. Mol Cell. 2013;51(6):792–806.
Talhouarne GJ, Gall JG. Lariat intronic RNAs in the cytoplasm of Xenopus tropicalis oocytes. RNA. 2014;20(9):1476–87.
Hsu MT, Coca-Prados M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature. 1979;280(5720):339–40.
Salzman J, et al. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE. 2012;7(2):e30733.
Memczak S, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–8.
Capel B, et al. Circular transcripts of the testis-determining gene sry in adult mouse testis. Cell. 1993;73(5):1019–30.
Fan X, et al. Single-cell RNA-seq transcriptome analysis of linear and circular RNAs in mouse preimplantation embryos. Genome Biol. 2015;16(1):148.
Enuka Y, et al. Circular RNAs are long-lived and display only minimal early alterations in response to a growth factor. Nucleic Acids Res. 2016;44(3):1370–83.
Li X, et al. Linking circular intronic RNA degradation and function in transcription by RNase H1. Sci China Life Sci. 2021;64(11):1795–809.
Hansen TB, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495(7441):384–8.
Xia P, et al. A circular RNA protects dormant hematopoietic stem cells from DNA sensor cGAS-Mediated exhaustion. Immunity. 2018;48(4):688–e7017.
Zhao Q, et al. Targeting Mitochondria-located circRNA SCAR alleviates NASH via reducing mROS output. Cell. 2020;183(1):76–e9322.
Wu X, et al. Kindlin-2 preserves integrity of the articular cartilage to protect against osteoarthritis. Nat Aging. 2022;2(4):332–47.
Chen CK, et al. Structured elements drive extensive circular RNA translation. Mol Cell. 2021;81(20):4300–e431813.
Legnini I, et al. Circ-ZNF609 is a circular RNA that can be translated and functions in Myogenesis. Mol Cell. 2017;66(1):22–e379.
Yang Y, et al. Novel role of FBXW7 circular RNA in repressing Glioma Tumorigenesis. J Natl Cancer Inst. 2018;110(3):304–15.
Yang Y, et al. Extensive translation of circular RNAs driven by N(6)-methyladenosine. Cell Res. 2017;27(5):626–41.
Tang C, et al. M(6)A-dependent biogenesis of circular RNAs in male germ cells. Cell Res. 2020;30(3):211–28.
Pamudurti NR, et al. Translation of CircRNAs. Mol Cell. 2017;66(1):9–e217.
Wurster SE, Maher LJ 3. Selection and characterization of anti-NF-kappaB p65 RNA aptamers. RNA. 2008;14(6):1037–47.
Litke JL, Jaffrey SR. Highly efficient expression of circular RNA aptamers in cells using autocatalytic transcripts. Nat Biotechnol. 2019;37(6):667–75.
Pfafenrot C, et al. Inhibition of SARS-CoV-2 coronavirus proliferation by designer antisense-circRNAs. Nucleic Acids Res. 2021;49(21):12502–16.
Jost I, et al. Functional sequestration of microRNA-122 from Hepatitis C Virus by circular RNA sponges. RNA Biol. 2018;15(8):1032–9.
Lavenniah A, et al. Engineered Circular RNA sponges act as miRNA inhibitors to attenuate pressure overload-Induced Cardiac Hypertrophy. Mol Ther. 2020;28(6):1506–17.
Elguindy MM, Mendell JT. NORAD-induced Pumilio phase separation is required for genome stability. Nature. 2021;595(7866):303–8.
Schreiner S, et al. Design and application of circular RNAs with protein-sponge function. Nucleic Acids Res. 2020;48(21):12326–35.
Qu L, et al. Circular RNA vaccines against SARS-CoV-2 and emerging variants. Cell. 2022;185(10):1728–e174416.
Zhou S, Van Bortle K. The Pol III transcriptome: basic features, recurrent patterns, and emerging roles in cancer. Wiley Interdiscip Rev RNA. 2023;14(5):e1782.
Lee YS, Lee YS. nc886, an RNA polymerase III-Transcribed noncoding RNA whose expression is dynamic and regulated by intriguing mechanisms. Int J Mol Sci, 2023. 24(10).
Naesens L, Haerynck F, Gack MU. The RNA polymerase III-RIG-I axis in antiviral immunity and inflammation. Trends Immunol. 2023;44(6):435–49.
Li JH, et al. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42(Database issue):D92–7.
Bernstein E, et al. Dicer is essential for mouse development. Nat Genet. 2003;35(3):215–7.
Wienholds E, et al. The microRNA-producing enzyme Dicer1 is essential for zebrafish development. Nat Genet. 2003;35(3):217–8.
Harfe BD, et al. The RNaseIII enzyme Dicer is required for morphogenesis but not patterning of the vertebrate limb. Proc Natl Acad Sci U S A. 2005;102(31):10898–903.
Berendsen AD, Olsen BR. Bone development. Bone. 2015;80:14–8.
Kobayashi T, et al. Dicer-dependent pathways regulate chondrocyte proliferation and differentiation. Proc Natl Acad Sci U S A. 2008;105(6):1949–54.
Gaur T, et al. Dicer inactivation in osteoprogenitor cells compromises fetal survival and bone formation, while excision in differentiated osteoblasts increases bone mass in the adult mouse. Dev Biol. 2010;340(1):10–21.
Wienholds E, et al. MicroRNA expression in zebrafish embryonic development. Science. 2005;309(5732):310–1.
Tuddenham L, et al. The cartilage specific microRNA-140 targets histone deacetylase 4 in mouse cells. FEBS Lett. 2006;580(17):4214–7.
Miyaki S, et al. MicroRNA-140 plays dual roles in both cartilage development and homeostasis. Genes Dev. 2010;24(11):1173–85.
Papaioannou G, et al. let-7 and miR-140 microRNAs coordinately regulate skeletal development. Proc Natl Acad Sci U S A. 2013;110(35):E3291–300.
Eberhart JK, et al. MicroRNA Mirn140 modulates Pdgf signaling during palatogenesis. Nat Genet. 2008;40(3):290–8.
Grigelioniene G, et al. Gain-of-function mutation of microRNA-140 in human skeletal dysplasia. Nat Med. 2019;25(4):583–90.
Nakamura Y, et al. Sox9 is upstream of microRNA-140 in cartilage. Appl Biochem Biotechnol. 2012;166(1):64–71.
Li P, et al. MicroRNA-1 regulates chondrocyte phenotype by repressing histone deacetylase 4 during growth plate development. FASEB J. 2014;28(9):3930–41.
Cong L, et al. MiR-1 is a critical regulator of chondrocyte proliferation and hypertrophy by inhibiting Indian hedgehog pathway during postnatal endochondral ossification in miR-1 overexpression transgenic mice. Bone. 2022;165:116566.
de Pontual L, et al. Germline deletion of the miR-17 approximately 92 cluster causes skeletal and growth defects in humans. Nat Genet. 2011;43(10):1026–30.
Han YC, et al. An allelic series of miR-17 approximately 92-mutant mice uncovers functional specialization and cooperation among members of a microRNA polycistron. Nat Genet. 2015;47(7):766–75.
Bluhm B, et al. miR-322 stabilizes MEK1 expression to inhibit RAF/MEK/ERK pathway activation in cartilage. Development. 2017;144(19):3562–77.
Wei J, et al. miR-34s inhibit osteoblast proliferation and differentiation in the mouse by targeting SATB2. J Cell Biol. 2012;197(4):509–21.
Atsawasuwan P, et al. Calvarial bone development and suture closure in Dicer-deficient mice. Orthod Craniofac Res. 2017;20(Suppl 1):26–31.
Ning G, et al. MicroRNA-92a upholds Bmp signaling by targeting noggin3 during pharyngeal cartilage formation. Dev Cell. 2013;24(3):283–95.
Nardocci G, et al. Identification of a novel long noncoding RNA that promotes osteoblast differentiation. J Cell Biochem. 2018;119(9):7657–66.
Sun K, et al. Mitophagy in degenerative joint diseases. Autophagy. 2021;17(9):2082–92.
Knezevic NN, et al. Low back pain. Lancet. 2021;398(10294):78–92.
Xu X, et al. Progerin accumulation in nucleus pulposus cells impairs mitochondrial function and induces intervertebral disc degeneration and therapeutic effects of sulforaphane. Theranostics. 2019;9(8):2252–67.
Jiang J, et al. The role of miRNA, lncRNA and circRNA in the development of intervertebral disk degeneration (review). Exp Ther Med. 2021;21(6):555.
Ohnishi T, Iwasaki N, Sudo H. Causes of and molecular targets for the Treatment of Intervertebral Disc Degeneration: a review. Cells, 2022. 11(3).
Sen P, et al. Epigenetic mechanisms of longevity and aging. Cell. 2016;166(4):822–39.
Li G, et al. WTAP-mediated m(6)a modification of lncRNA NORAD promotes intervertebral disc degeneration. Nat Commun. 2022;13(1):1469.
Ammari M, et al. Delivery of miR-146a to Ly6C(high) Monocytes inhibits pathogenic bone Erosion in inflammatory arthritis. Theranostics. 2018;8(21):5972–85.
Nakasa T, et al. Expression of microRNA-146 in rheumatoid arthritis synovial tissue. Arthritis Rheum. 2008;58(5):1284–92.
Lochhead RB, et al. MicroRNA-146a provides feedback regulation of lyme arthritis but not carditis during infection with Borrelia burgdorferi. PLoS Pathog. 2014;10(6):e1004212.
Zheng M, et al. miR-146a-5p targets Sirt1 to regulate bone mass. Bone Rep. 2021;14:101013.
Zhao J, et al. MiR-146a deletion protects from bone loss in OVX mice by suppressing RANKL/OPG and M-CSF in bone microenvironment. J Bone Min Res. 2019;34(11):2149–61.
Gu SX, et al. MicroRNA-146a reduces IL-1 dependent inflammatory responses in the intervertebral disc. Gene. 2015;555(2):80–7.
Guo Y, et al. Preliminary construction of a regulatory network of miRNAs in the pathogenesis of nucleus pulposus degeneration - a review based on data mining. Am J Transl Res. 2021;13(9):9919–31.
Zhang S, et al. Role of microRNA-15a-5p/Sox9/NF-κB axis in inflammatory factors and apoptosis of murine nucleus pulposus cells in intervertebral disc degeneration. Life Sci. 2021;277:119408.
Zhao Y, Li A. miR-19b-3p relieves intervertebral disc degeneration through modulating PTEN/PI3K/Akt/mTOR signaling pathway. Aging. 2021;13(18):22459–73.
Lei C et al. MicroRNA–25 protects nucleus pulposus cells against apoptosis via targeting SUMO2 in intervertebral disc degeneration. Mol Med Rep, 2021. 24(4).
Yuan X, et al. Human umbilical cord mesenchymal stem cells deliver exogenous miR-26a-5p via exosomes to inhibit nucleus pulposus cell pyroptosis through METTL14/NLRP3. Mol Med. 2021;27(1):91.
Sun Y, Zhang W, Li X. Induced pluripotent stem cell-derived mesenchymal stem cells deliver exogenous mir-105-5p via small extracellular vesicles to rejuvenate senescent nucleus pulposus cells and attenuate intervertebral disc degeneration. Stem Cell Res Ther. 2021;12(1):286.
Wu T, et al. Inhibition of miR-130b-3p restores autophagy and attenuates intervertebral disc degeneration through mediating ATG14 and PRKAA1. Apoptosis. 2022;27(5–6):409–25.
Du L, et al. LncRNA nuclear receptor subfamily 2 group F member 1 antisense RNA 1 (NR2F1-AS1) aggravates nucleus pulposus cell apoptosis and extracellular matrix degradation. Bioengineered. 2022;13(2):2746–62.
Lin XL, et al. Expression of miR-195 and its target gene Bcl-2 in human intervertebral disc degeneration and their effects on nucleus pulposus cell apoptosis. J Orthop Surg Res. 2021;16(1):412.
Li Y, et al. Circ_0004354 might compete with circ_0040039 to induce NPCs death and inflammatory response by targeting miR-345-3p-FAF1/TP73 axis in intervertebral disc degeneration. Oxid Med Cell Longev. 2022;2022:p2776440.
Lin X, Lin Q. MiRNA-495-3p attenuates TNF-α Induced apoptosis and inflammation in human nucleus Pulposus cells by targeting IL5RA. Inflammation. 2020;43(5):1797–805.
Sun JC, et al. MiR-499a-5p suppresses apoptosis of human nucleus pulposus cells and degradation of their extracellular matrix by targeting SOX4. Biomed Pharmacother. 2019;113:108652.
Guo Z, et al. MiR-502 suppresses TNF-α-Induced Nucleus Pulposus Cell apoptosis by targeting TARF2. Biomed Res Int. 2021;2021:p5558369.
Zhang HJ, et al. Knockdown of miR-660 protects nucleus pulposus cells from TNF-a-induced apoptosis by targeting serum amyloid A1. J Orthop Surg Res. 2020;15(1):7.
Guo Y, et al. ERRFI1 inhibits proliferation and inflammation of Nucleus Pulposus and is negatively regulated by mir-2355-5p in intervertebral disc degeneration. Spine (Phila Pa 1976). 2019;44(15):E873–81.
Zhang J et al. MiR-4478 accelerates Nucleus Pulposus cells apoptosis Induced by oxidative stress by targeting MTH1. Spine (Phila Pa 1976), 2022.
Wang J, et al. Oxidative stress-induced circKIF18A downregulation impairs MCM7-mediated anti-senescence in intervertebral disc degeneration. Exp Mol Med. 2022;54(3):285–97.
Du X, et al. Circular RNA hsa_circ_0083756 promotes intervertebral disc degeneration by sponging miR-558 and regulating TREM1 expression. Cell Prolif. 2022;55(4):e13205.
Kong D, et al. Knockdown of hsa_circ_0059955 induces apoptosis and cell cycle arrest in Nucleus Pulposus cells via inhibiting itchy E3 ubiquitin protein ligase. Drug Des Devel Ther. 2020;14:3951–63.
Huang Y, et al. circSPG21 protects against intervertebral disc disease by targeting miR-1197/ATP1B3. Exp Mol Med. 2021;53(10):1547–58.
Huang Y, et al. Inhibition of intervertebral disc disease progression via the circPKNOX1-miR-370-3p-KIAA0355 axis. Cell Death Discov. 2021;7(1):39.
Li Y, et al. Identification of circ-FAM169A sponges miR-583 involved in the regulation of intervertebral disc degeneration. J Orthop Translat. 2021;26:121–31.
Xie L, et al. CircERCC2 ameliorated intervertebral disc degeneration by regulating mitophagy and apoptosis through miR-182-5p/SIRT1 axis. Cell Death Dis. 2019;10(10):751.
Wang H, et al. Circular RNA circ-4099 is induced by TNF-α and regulates ECM synthesis by blocking mir-616-5p inhibition of Sox9 in intervertebral disc degeneration. Exp Mol Med. 2018;50(4):1–14.
Chen W, Li S, Zhang F. Role of lncRNA XIST/microRNA-19/PTEN network in autophagy of nucleus pulposus cells in intervertebral disc degeneration via the PI3K/Akt signaling pathway. Cell Cycle. 2021;20(17):1629–41.
Rheumatoid arthritis. Nat Rev Dis Primers. 2018;4:18002.
Hu XX, et al. T-cells interact with B cells, dendritic cells, and fibroblast-like synoviocytes as hub-like key cells in rheumatoid arthritis. Int Immunopharmacol. 2019;70:428–34.
Li X, Tian F, Wang F. Rheumatoid arthritis-associated microRNA-155 targets SOCS1 and upregulates TNF-α and IL-1β in PBMCs. Int J Mol Sci. 2013;14(12):23910–21.
Rodriguez A, et al. Requirement of bic/microRNA-155 for normal immune function. Science. 2007;316(5824):608–11.
Blüml S, et al. Essential role of microRNA-155 in the pathogenesis of autoimmune arthritis in mice. Arthritis Rheum. 2011;63(5):1281–8.
Chen S, et al. MicroRNA-155-deficient dendritic cells cause less severe GVHD through reduced migration and defective inflammasome activation. Blood. 2015;126(1):103–12.
Taganov KD, et al. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A. 2006;103(33):12481–6.
Huang J, et al. The microRNAs miR-204 and miR-211 maintain joint homeostasis and protect against osteoarthritis progression. Nat Commun. 2019;10(1):2876.
Wang QS et al. Mir204 and Mir211 suppress synovial inflammation and proliferation in rheumatoid arthritis by targeting Ssrp1. Elife, 2022. 11.
Guo T, et al. Long non-coding RNA NEAT1 Knockdown alleviates rheumatoid arthritis by reducing IL-18 through p300/CBP repression. Inflammation. 2022;45(1):100–15.
Shui X, et al. Knockdown of lncRNA NEAT1 inhibits Th17/CD4(+) T cell differentiation through reducing the STAT3 protein level. J Cell Physiol. 2019;234(12):22477–84.
Han JJ, Wang XQ, Zhang XA. Functional interactions between lncRNAs/circRNAs and miRNAs: insights into rheumatoid arthritis. Front Immunol. 2022;13:810317.
Jin D, et al. Systematic analysis of lncRNAs, mRNAs, circRNAs and miRNAs in patients with postmenopausal osteoporosis. Am J Transl Res. 2018;10(5):1498–510.
Wang R, et al. miR-143 promotes angiogenesis and osteoblast differentiation by targeting HDAC7. Cell Death Dis. 2020;11(3):179.
Yang M, et al. MiR-497∼195 cluster regulates angiogenesis during coupling with osteogenesis by maintaining endothelial notch and HIF-1α activity. Nat Commun. 2017;8:16003.
Li CJ, et al. Long noncoding RNA bmncr regulates mesenchymal stem cell fate during skeletal aging. J Clin Invest. 2018;128(12):5251–66.
Mulati M, et al. The long noncoding RNA crnde regulates osteoblast proliferation through the Wnt/β-catenin signaling pathway in mice. Bone. 2020;130:115076.
Li CJ, et al. MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation. J Clin Invest. 2015;125(4):1509–22.
Liu C, et al. The mechanosensitive lncRNA Neat1 promotes osteoblast function through paraspeckle-dependent Smurf1 mRNA retention. Bone Res. 2022;10(1):18.
Xu R, et al. Identification of the canonical and noncanonical role of miR-143/145 in estrogen-deficient bone loss. Theranostics. 2021;11(11):5491–510.
Cui Q, et al. Mmu-miR-185 depletion promotes osteogenic differentiation and suppresses bone loss in osteoporosis through the bgn-mediated BMP/Smad pathway. Cell Death Dis. 2019;10(3):172.
Lin C, et al. Circulating miR-338 cluster activities on osteoblast differentiation: potential diagnostic and therapeutic targets for postmenopausal osteoporosis. Theranostics. 2019;9(13):3780–97.
Lu XD, Han WX, Liu YX. Suppression of miR-451a accelerates osteogenic differentiation and inhibits bone loss via Bmp6 signaling during osteoporosis. Biomed Pharmacother. 2019;120:109378.
Hu CH, et al. miR-21 deficiency inhibits osteoclast function and prevents bone loss in mice. Sci Rep. 2017;7:43191.
Shen G, et al. miR-128 plays a critical role in murine osteoclastogenesis and estrogen deficiency-induced bone loss. Theranostics. 2020;10(10):4334–48.
Pan B, et al. MiR-148a deletion protects from bone loss in physiological and estrogen-deficient mice by targeting NRP1. Cell Death Discov. 2022;8(1):470.
Zhu J, Wang H, Liu H. Osteoclastic mir-301-b knockout reduces ovariectomy (OVX)-induced bone loss by regulating CYDR/NF-κB signaling pathway. Biochem Biophys Res Commun. 2020;529(1):35–42.
Jin F, et al. A functional motif of long noncoding RNA nron against osteoporosis. Nat Commun. 2021;12(1):3319.
Hunter DJ, Bierma-Zeinstra S. Osteoarthr Lancet. 2019;393(10182):1745–59.
Chen D, et al. Osteoarthritis: toward a comprehensive understanding of pathological mechanism. Bone Res. 2017;5:16044.
Rocha FAC, Ali SA. Soluble biomarkers in osteoarthritis in 2022: year in review. Osteoarthritis Cartilage; 2022.
Dy P, et al. Sox9 directs hypertrophic maturation and blocks osteoblast differentiation of growth plate chondrocytes. Dev Cell. 2012;22(3):597–609.
Nagata K, et al. Runx2 and Runx3 differentially regulate articular chondrocytes during surgically induced osteoarthritis development. Nat Commun. 2022;13(1):6187.
Ito Y, et al. Both microRNA-455-5p and – 3p repress hypoxia-inducible factor-2α expression and coordinately regulate cartilage homeostasis. Nat Commun. 2021;12(1):4148.
Hu S, et al. MicroRNA-455-3p promotes TGF-β signaling and inhibits osteoarthritis development by directly targeting PAK2. Exp Mol Med. 2019;51(10):1–13.
Yoon DS, et al. RUNX2 stabilization by long non-coding RNAs contributes to hypertrophic changes in human chondrocytes. Int J Biol Sci. 2023;19(1):13–33.
Chen D, et al. Runx2 plays a central role in Osteoarthritis development. J Orthop Translat. 2020;23:132–9.
Wang XB, et al. MicroRNA-21-5p as a novel therapeutic target for osteoarthritis. Rheumatology (Oxford); 2019.
Ma S, et al. MiR-21-5p regulates extracellular matrix degradation and angiogenesis in TMJOA by targeting Spry1. Arthritis Res Ther. 2020;22(1):99.
Ji ML, et al. Precise targeting of miR-141/200c cluster in chondrocytes attenuates osteoarthritis development. Ann Rheum Dis. 2021;80(3):356–66.
Yamasaki K, et al. Expression of MicroRNA-146a in osteoarthritis cartilage. Arthritis Rheum. 2009;60(4):1035–41.
Zhang X et al. miR-146a facilitates osteoarthritis by regulating cartilage homeostasis via targeting Camk2d and Ppp3r2 Cell Death Dis, 2017. 8(4): p. e2734.
Wang H, et al. Intra-articular delivery of Antago-Mir-483-5p inhibits osteoarthritis by modulating Matrilin 3 and tissue inhibitor of metalloproteinase 2. Mol Ther. 2017;25(3):715–27.
Wang H, et al. Chondrocyte mTORC1 activation stimulates mir-483-5p via HDAC4 in osteoarthritis progression. J Cell Physiol. 2019;234(3):2730–40.
Tang S, et al. Circular RNA circNFKB1 promotes osteoarthritis progression through interacting with ENO1 and sustaining NF-kappaB signaling. Cell Death Dis. 2022;13(8):695.
Zhu H, et al. CircGCN1L1 promotes synoviocyte proliferation and chondrocyte apoptosis by targeting mir-330-3p and TNF-alpha in TMJ osteoarthritis. Cell Death Dis. 2020;11(4):284.
Shen S, et al. CircSERPINE2 protects against osteoarthritis by targeting miR-1271 and ETS-related gene. Ann Rheum Dis. 2019;78(6):826–36.
Shen S, et al. circPDE4B prevents articular cartilage degeneration and promotes repair by acting as a scaffold for RIC8A and MID1. Ann Rheum Dis. 2021;80(9):1209–19.
Zhao C, et al. CircFOXO3 protects against osteoarthritis by targeting its parental gene FOXO3 and activating PI3K/AKT-mediated autophagy. Cell Death Dis. 2022;13(11):932.
Lin Z, et al. Potential predictive and therapeutic applications of small extracellular vesicles-derived circPARD3B in osteoarthritis. Front Pharmacol. 2022;13:968776.
Hu J et al. Long non-coding RNA HOTAIR promotes osteoarthritis progression via miR-17-5p/FUT2/beta-catenin axis. Cell Death Dis, 2018. 9(7): p. 711.
Fu Q, et al. LINC02288 promotes chondrocyte apoptosis and inflammation through miR-374a-3p targeting RTN3. J Gene Med. 2021;23(5):e3314.
Chen Y, et al. Long-chain non-coding RNA HOTAIR promotes the progression of osteoarthritis via sponging miR-20b/PTEN axis. Life Sci. 2020;253:117685.
Bai J, et al. LncRNA MM2P-induced, exosome-mediated transfer of Sox9 from monocyte-derived cells modulates primary chondrocytes. Cell Death Dis. 2020;11(9):763.
Feng L, et al. Linc-ROR promotes mesenchymal stem cells chondrogenesis and cartilage formation via regulating SOX9 expression. Osteoarthritis Cartilage. 2021;29(4):568–78.
Endisha H, et al. MicroRNA-34a-5p promotes Joint Destruction during Osteoarthritis. Arthritis Rheumatol. 2021;73(3):426–39.
Acknowledgements
Not applicable.
Funding
This work was supported, in part, by the National Key Research and Development Program of China Grants (2019YFA0906004), the National Natural Science Foundation of China Grants (82230081, 82250710175, 82172375, 81991513, and 82261160395), the Shenzhen Fundamental Research Program (JCYJ20220818100617036), the Shenzhen Key Laboratory of Cell Microenvironment Grant (ZDSYS20140509142721429) and the Guangdong Provincial Science and Technology Innovation Council Grant (2017B030301018).
Author information
Authors and Affiliations
Contributions
QY, LL, and GX conceptualized and wrote the outline of the manuscript. QY, TH, JL, XW, RL, and LL did the literature search and wrote the manuscript draft. GX reviewed and edited the manuscript. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yao, Q., He, T., Liao, JY. et al. Noncoding RNAs in skeletal development and disorders. Biol Res 57, 16 (2024). https://doi.org/10.1186/s40659-024-00497-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40659-024-00497-y