
Abstract
Background
ADP-glucose pyrophosphorylase (AGPase), the key enzyme in plant starch biosynthesis, is a heterotetramer composed of two identical large subunits and two identical small subunits. AGPase has plastidial and cytosolic isoforms in higher plants, whereas it is mainly detected in the cytosol of grain endosperms in cereal crops. Our previous results have shown that the expression of the TaAGPL1 gene, encoding the cytosolic large subunit of wheat AGPase, temporally coincides with the rate of starch accumulation and that its overexpression dramatically increases wheat AGPase activity and the rate of starch accumulation, suggesting an important role.
Methods
In this study, we performed yeast one-hybrid screening using the promoter of the TaAGPL1 gene as bait and a wheat grain cDNA library as prey to screen out the upstream regulators of TaAGPL1 gene. And the barley stripe mosaic virus-induced gene-silencing (BSMV-VIGS) method was used to verify the functional characterization of the identified regulators in starch biosynthesis.
Results
Disulfide isomerase 1-2 protein (TaPDIL1-2) was screened out, and its binding to the TaAGPL1-1D promoter was further verified using another yeast one-hybrid screen. Transiently silenced wheat plants of the TaPDIL1-2 gene were obtained by using BSMV-VIGS method under field conditions. In grains of BSMV-VIGS-TaPDIL1-2-silenced wheat plants, the TaAGPL1 gene transcription levels, grain starch contents, and 1000-kernel weight also significantly increased.
Conclusions
As important chaperones involved in oxidative protein folding, PDIL proteins have been reported to form hetero-dimers with some transcription factors, and thus, our results suggested that TaPDIL1-2 protein could indirectly and negatively regulate the expression of the TaAGPL1 gene and function in starch biosynthesis.
Similar content being viewed by others

Background
Bread wheat is one of the most important cereal crops, accounting for approximately 20% of the daily calorie consumption worldwide, and starch is composed of approximately 70% wheat grains and is the most abundant storage carbohydrate in this organ [1]. AGPase uses the substrates glucose-1-phosphate (G-1-P) and ATP to produce pyrophosphate and ADP-glucose, the starch precursor, and is thought to be the key enzyme in starch synthesis [2]. In plants, AGPase is a heterotetramer composed of two large/modulatory subunits (LSU, or AGPL) and two small/catalytic subunits (SSU or AGPS) [3]. According to its cellular localization, AGPases have cytosolic and plastidial isoforms and these two types function independently to produce starch in cytosol and plastid, respectively. Thus, in plant cells, there are four types of AGPase subunits: cytosolic SSU, plastidial SSU, cytosolic LSU and plastidial LSU [4]. In the plant cells of most dicotyledonous plants, AGPase is active exclusively in plastids, but it is mainly present in the cytosol in the grain endosperm of the grass family, demonstrating approximately 85%, 90%, and 95% of the total AGPase activity in barley, rice, and maize, respectively [5,6,7], suggesting a crucial role of cytosolic AGPase in starch synthesis in these species.
The number of AGPase subunits is species-specific. For example, four AGPase large subunits (AGPL1, AGPL2, AGPL3, and AGPL4) have been identified in rice and maize, only two subunits, AGPL1 and AGPL2, are present in barley, and the extents to which they play roles in starch biosynthesis are different [8, 9]. Our previous studies have demonstrated that the expression of the TaAGPL1 gene encoding the cytosolic large subunit of wheat AGPase shows similar changes to the rate of starch accumulation, and its transcripts in wheat grains are significantly and positively related to the activity of AGPase and the accumulation rate of grain starch [10]. Moreover, overexpression of TaAGPL1 markedly enhanced AGPase activity and the rate of starch accumulation in wheat grains [11]. These data demonstrate that TaAGPL1 plays an important role in AGPase enzyme and starch biosynthesis in bread wheat and could have potential applications in wheat breeding to develop high-yield wheat cultivars [11]. However, the upstream regulatory mechanism of the TaAGPL1 gene family remains elusive.
Here we used the TaAGPL1 promoter as bait to screen an immature wheat grain cDNA library using a yeast one-hybrid (Y1H) assay to isolate upstream regulators. Among the identified proteins, one protein disulfide isomerase 1-2 (TaPDIL1-2) was selected to evaluate its binding to the TaAGPL1 promoter and test its potential function in regulating TaAGPL1 expression and starch biosynthesis.
Methods
Plant materials
Two semi-winter bread wheat cultivars, Zhoumai 18 and Bainong 207, characterized with high yield potentials, have widely been planted in the Yellow and Huai Valleys Winter Wheat Zone. The former was used in our previous studies [10, 12], and in this study, its leaf DNA was extracted to isolate the TaAGPL1-1D promoter and its grains were sampled for endosperm cDNA library synthesis and for detection of the TaAGPL1-1D promoter activity. The latter is relatively sensitive to plant viruses and was used for the BSMV-VIGS assay [13].
GUS activity of the TaAGPL1-1D gene assay
The sequence of TaAGPL1-1D gene was used as a query to search for its upstream promoter sequence in the International Wheat Genome Sequencing Consortium (IWGSC) database. A 1280-bp fragment was retrieved and used as its promoter sequence. A pair of primers were designed to amplify this fragment from cv. Zhoumai 18 (Additional file 1: Table S1), and then a series of nested 5′ deletions of this promoter were inserted into the pCAMBIA1301 vector containing the CaMV35S promoter to drive expression of the GUS gene, forming the Pro-1: GUS (− 1269 bp to + 11 bp), Pro-2: GUS (− 870 bp to + 11 bp), and Pro-3: GUS (− 483 bp to +11 bp) recombinant vectors, respectively. GUS activity of TaAGPL1-1D was assessed using a transient transformation system in N. benthamiana plants or immature wheat grains. In tobacco plants, the recombinant plasmids of Pro-1: GUS, Pro-2: GUS, and Pro-3: GUS were transferred into the Agrobacterium EHA105 lines, and then the EHA105 lines were coinfiltrated into the N. benthamiana leaves [14]. For wheat grains, immature grains sampled at 20 days after anthesis were used to qualitatively and quantitatively determine the TaAGPL1-1D promoter activity using gene gun-mediated transient transformation protocol [15]. The recombinant plasmids of Pro-1: GUS, Pro-2: GUS, and Pro-3: GUS were transfected using the PDS-1000/He Particle Delivery SystemTM (Bio-Rad, Hercules, CA, USA). Stained samples were photographed using a Canon Digital Camera EOS 70 D (Canon Co., Ltd., Zhuhai, China). Quantitative GUS activity was subsequently measured [16].
Identification of TaPDIL1-2 protein by Y1H screening
Approximately 2 mg of high-quality total RNA from wheat endosperm sampled at 5, 10, 15, 20, 25, 30, and 35 days after anthesis was mixed and extracted with TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Wheat endosperm cDNA library was constructed using the Advantage 2 PCR Kit [Cat No. 639206, TaKaRa Biotechnology Co., Ltd., Dalian, China]. The TaAGPL1-1D promoter was used to construct the bait vector (pAbAi-TaAGPL1). The pAbAi-TaAGPL1 vector was then linearized and integrated into the genome of yeast strain Y1HGold. The bait strain Y1HGold/TaAGPL1-pro-pAbAi survived on SD/-Ura medium. The plasmids of the wheat endosperm cDNA library were next transformed into competent cells of the bait strain, and the transformed yeast cells grew on plates of SD/-Leu, -Ura, +AbA100 at 30 °C for 2–4 days. The positive yeast colonies were isolated and confirmed by PCR amplification with 5′ T7-F and 3′ AD-R primers (Additional file 1: Table S1).
The interaction between TaPDIL1-2 protein and TaAGPL1-1D promoter was measured using another Y1H assay. Three fragments (Pro-1, 1280 bp, Pro-2, 881 bp and Pro-3, 494 bp) of the TaAGPL1-1D promoter were separately ligated into the pAbAi vector. ORF (open reading frame) sequence of TaPDIL1-2 gene was cloned and then ligated into the pGADT7 vector to produce the prey vector. Two concentrations (0 and 100 ng/mL) of AbA were used, and the TaPDIL1-2 prey vector was transformed into yeast Y1HGold strains containing the Bait-Reporter vectors. pGADT7-TaPDIL1-2 and pAbAi-TaAGPL1 promoters were co-transformed into the Y1HGold yeast strains. The transformed cells were grown on SD/-Leu, -Ura selective medium with 100 ng/mL AbA at 30 °C for 2–4 days.
Transcriptional expression data for the TaAGPL1-1D gene in grain, spike, leaf/stem at reproductive and vegetative stages were obtained from the expVIP database and then viewed as a heat map using the HemI-1.0 tool (Heatmap illustrator) [17].
BSMV-VIGS experiment
To alleviate functional complementation and allow complete silencing, we selected a conserved cDNA fragment (190 bp) of the TaPDIL1-2 gene, three homoeologs of which shared 99.1% similarity, possibly enabling simultaneous silencing using the BSMV-VIGS method. The primers used for the BSMV-TaPDIL1-2 vector are shown in Additional file 1: Table S1. Viral vector construction, viral RNA transcription, and viral inoculation of BSMV-VIGS vectors were performed as described in our previous studies [13, 18]. BSMV-GFP-inoculated wheat seedlings were used as the negative control, and the appearance of chlorosis on the inoculated wheat spikes confirmed the successful BSMV inoculation [19]. In the field, BSMV inoculation was performed in the spike of wheat plants at the heading stage, and a total of 142 and 167 spikes were inoculated with BSMV-TaPDIL1-2 or BSMV-GFP, respectively, using a 20-µL transcript mixture for each spike.
Transcription levels of the TaPDIL1-2 gene in the wheat grains at 19, 24, 29, and 34 days after anthesis were measured using the quantitative real-time PCR (qPCR) [20]. The wheat Actin and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (GenBank accession no. AB181991 and EF592180) genes were used as internal controls.
Statistical analysis
One-way analysis of variance (ANOVA) was performed using SPSS version 17.0 software. Data are the mean ± standard deviation (SD) of at least three independent experiments. A multiple range test was used to compare the mean values at the P < 0.05 level.
Results and discussion
Isolation of the TaAGPL1 promoter
Bread wheat genome contains three closely related, yet distinct, subgenomes (AABBDD) and three homoeologs, with over 95% sequence identity in their coding regions for the majority of genes [17]. In bread wheat, rice and maize, AGPL1 gene transcripts showed high levels in endosperm, whereas they were not detected in leaves or other organs [7,8,9, 12], suggesting that the promoter could be endosperm-specific. The cDNA sequence of the TaAGPL1 gene, which was highly expressed in endosperm during the grain filling period in our previous study [10], was searched against the recently published IWGSC databases (RefSeq v1.0) [21], and a high level (100% identities) to a chromosome-located contig (TraesCS1D01G427400.1) was found, suggesting its localization on 1D chromosome.
The sequence of the TaAGPL1-1D gene was used as a query to search its upstream promoter sequence in the IWGSC database, and a 1280-bp fragment (− 1269 bp to + 11 bp) was retrieved and used to design the primer pairs to amplify its promoter sequence. The amplified TaAGPL1-1D promoter demonstrated high similarity (99.8%) to the retrieved IWGSC sequence (Additional file 2: Fig. S1). Using the classic transient transformation system [14], we did not detect histochemical GUS activity of the TaAGPL1-1D promoter in the leaves of transiently transformed-N. benthamiana plants (Additional file 3: Fig. S2), possibly due to its endosperm-specific expression profile. Alternatively, a gene gun-mediated transient transformation protocol for wheat grains was used to detect TaAGPL1-1D promoter activity [16]. Our experiment showed that the TaAGPL1-1D promoter qualitatively and quantitatively drove expression of the GUS gene (Fig. 1), arguing that this promoter contains cis-acting elements that regulate TaAGPL1-1D gene expression in wheat grains.

Qualitative and quantitative detection of the TaAGPL1 promoter-driven GUS activity in the wheat grains. a Qualitative staining observation on GUS driven by three fragments of the TaAGPL1 promoter. Upper panel, Pro-1, Pro-2, and Pro-3 represent 1280 bp (− 1269 bp to + 11 bp), 881 bp (− 870 bp to + 11 bp), and 494 bp (− 483 bp to + 11 bp) of the TaAGPL1 promoter, respectively; 35 S represents CaMV35S promoter; Lower panel, negative controls. b Quantitative detection on GUS activities driven by three different fragments of TaAGPL1 promoter or 35S promoter. Each value is mean ± standard deviation of three biological replicates, and different letters indicate significant differences (P < 0.05) relative to control by using one-way ANOVA of Duncan’s multiple range test
The binding between TaPDIL1-2 gene and TaAGPL1 promoter
Wheat grains were sampled at 5, 10, 15, 20, 25, 30, and 35 days after anthesis, endosperm was reserved and other organs were removed. The mRNA isolated from the sampled endosperm was used as template to synthesize the full-length double-stranded complementary DNA and together with the linearized pGADT7-Rec vector was co-transformed into yeast strain Y1HGold cells for directional combination through homologous recombination. The combined plasmids were extracted and subsequently transformed into E. coli strain stellar cells. The total capacity was 1.08 ± 0.3 × 106 cfu with more than 77.9% inserted fragments greater than 750 bp (Additional file 4: Fig. S3), which was suitable for yeast screening.
Using the above-amplified TaAGPL1-1D promoter as bait and wheat endosperm cDNA library as prey, we performed Y1H screening (Fig. 2a), and identified 39 positive clones, which were subsequently sequenced and functionally annotated (Additional file 5: Table S2). Of these clones, the TaPDIL1-2 gene was screened out. The complete open reading frame (ORF, 1539 bp) of the TaPDIL1-2 gene was subsequently cloned (Additional file 6: Fig. S4), and its prey vector was constructed and transformed into the bait strain Y1HGold containing three fragments (1280 bp, 881 bp, or 494 bp) of the TaAGPL1-1D promoter (Fig. 2b), respectively, to perform the second Y1H screening with 0 or 100 ng/mL AbA. Our experiment showed that the transformed yeast cells containing Y1HGold/Pro-1 (− 1269 bp to + 11 bp) grew optimally in SD/-Leu, -Ura, +AbA100 medium, followed by Y1HGold/Pro-2 (− 870 bp to + 11 bp) and Y1HGold/Pro-3 (− 483 bp to + 11 bp) (Fig. 2c). These data confirmed the binding between TaPDIL1-2 protein and TaAGPL1-1D promoter, with the potential binding region located in the promoter between − 483 and − 1269 bp.

Y1H assay between TaPDIL1-2 protein and TaAGPL1-1D promoter. a cDNA library screen of TaAGPL1-1D promoter in yeast cells; b different fragments (Pro-1, 1280 bp, Pro-2, 881 bp, and Pro-3, 494 bp) of TaAGPL1-1D promoter using Y1H assay; c yeast cells transformed with both the bait vectors containing the above three promoter fragments fused to a pAbAi reporter gene, and a prey vector containing TaPDIL1-2 fused to a pGADT7 activation domain. Yeast cells were grown in liquid media to an OD600 of 0.02, and diluted in a 10 × dilution series (from 100 to 10−3). Each dilution 5 μL was spotted on media selecting for plasmids (SD/-Leu, -Ura), supplemented with 0 and 100 ng/mL aureobasidin A (AbA)
Characterization and expression of TaPDIL1-2
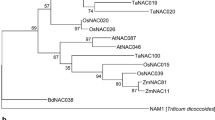
By searching the IWGSC databases, the isolated TaPDIL1-2 gene was localized to 4BS, and its ORF encoded a 512-amino-acid protein of 56.43 kDa with a predicted pI of 5.03. The deduced TaPDIL1-2 protein contained 4 classical thioredoxin domains, including 2 redox CGHC active sites (a and a′), 2 inactive domains (b and b′), and 1 C-terminal KDEL signal sequence (KDEL), which is a classcial endoplasmic reticulun (ER)-retention signal supporting ER-localization (Fig. 3a). Phylogenetic analysis indicated that the thioredoxin domain of TaPDIL1-2 shared more than 75.7% identity with those of AtPDIL1-2, OsPDIL1-1, OsPDIL1-2 and ZmPDIL1-2, belonging to one sub-branch of group I in the PDIL subfamily (Fig. 3b). Some PDIL proteins have been identified and functionally characterized in the wheat genome [22]. To our knowledge, however, there have been no reports regarding TaPDIL1-2 or its orthologous genes, and the amino acid sequence of TaPDIL1-2 had low similarities (< 24.3%) to other wheat PDIL proteins (Fig. 3b).

Domains, phylogenetic relationships and expression profiles of TaPDIL1-2. a Putative amino acid sequence of the protein encoded by the isolated TaPDIL1-2 in this study. a, a′, b, b′ Represent two redox active site CGHC and two inactive domains, respectively, and they are underlined with black. C-terminal KDEL signal sequence is marked with green color; b phylogenetic analysis of plant PDILs. The phylogenetic tree was constructed using neighbor-joining phylogeny of MEGA 5.0 with default parameter; c In silico expression profiling of TaPDIL1-2 homoeologs in different organs at different growth stages of cv. Chinese Spring wheat. The expression data were generated from the expVIP database. L/S, leaf/stem; Sp, spike; G, grain; Ve, vegetative stage; Re, reproductive stage; 4B, 4D, and 4A represent TaPDIL1-2 4B, 4D and 4A homoeologs, respectively
At the transcriptional level, the in silico expression data revealed three homoeologs of the TaPDIL1-2 gene that were more highly expressed during the reproductive stage than the vegetative stages, and during the reproductive stage, their expression level were higher in the endosperm/grain and spikes than in stem and leaf (Fig. 3c). The bread wheat endosperm mainly accumulates starch (≥ 70%) and protein (10–14%), forming the bulk of the grain. These findings suggest that TaPDIL1-2 homoeologs can function in starch biosynthesis in wheat grain.
Negative regulation of the TaPDIL1-2 gene in wheat starch biosynthesis
Gene function in higher plants is often explored through the use transgenic and mutational assays. In bread wheat, however, the multiple copy insertions, low transformation efficiency, cultivar specificity, time consumption, and high cost of transgenic approaches have greatly limited gene function studies in this species [23]. The functional redundancy among homoeologs in this species also causes some difficulties in terms of generating null mutants [24]. BSMV-VIGS method facilitates the rapid generation of gene knockdown phenotypes in polyploid species because plant transformation is not required, accelerating the characterization of target genes [25]. In this study, this method was used to evaluate the function of the TaPDIL1-2 gene. We constructed BSMV-TaPDIL1-2 and BSMV-GFP vectors, and the latter was used as the control. To prevent functional complementation and allow complete silencing, we selected a conserved cDNA fragment (190 bp) of the TaPDIL1-2 gene, three homoeologs of which shared 99.1% similarity (Additional file 7: Fig. S5), potentially enabling the simultaneous silencing of its three homoeologs using the BSMV-VIGS method.
In previous studies, the BSMV-VIGS method has mostly been applied to explore the function of candidate genes of cereal crops in controlled conditions considering responses to single or several environmental factors [26,27,28]. However, there are some differences (e.g. stronger plants, more yields) in growth and phenotypes of most higher plants including important crops grown between field conditions with multiple and variable factors (light, temperature, etc.) and under the controlled conditions [26]. Thus, field experiments can more efficiently explore the function of the target genes. By using arch plastic sheds combined with water-sprayed spikes, a few efficient and convenient approaches were designed in our previous study for application of the BSMV-VIGS method to wheat plants under field conditions [13]. In the present study, BSMV-TaPDIL1-2 and BSMV-GFP vectors were separately used to inoculate 142 and 167 wheat spikes at anthesis under field conditions. We observed that BSMV-VIGS-induced chlorosis occurred on inoculated-spikes at 19 days after anthesis, gradually extended throughout the entire spike, and persisted until the mature stage (Fig. 4a), suggesting that these two virus vectors were successfully inoculated into the wheat spikes.

Phenotypes and its transcription levels of TaPDIL1-2-silencing wheat plants. Grain starch contents and 1000-kernel weight of BSMV-TaPDIL1-2-silenced wheat plants, and transcription levels of TaAGPL1-1D gene. a Phenotypes of BSMV-TaPDIL1-2-, and BSVM-GFP-inoculated wheat spikes at 19 days after anthesis, respectively; b, c transcriptional levels of the TaPDIL1-2 gene at 19, 24, 29, and 34 days after anthesis detected by using the qPCR method with the wheat Actin (b) and GAPDH (c) genes as two internal controls; d and e, grain starch contents and 1000-kernel weight, respectively; f, g transcription levels of the TaAGPL1-1D genes in inoculated wheat grains at 19, 24, 29, and 34 days after anthesis with Actin (f) and GAPDH (g) genes as two internal controls, respectively. Each value is mean ± standard deviation of three biological replicates, and different letters indicate significant differences (P < 0.05) relative to control by using one-way ANOVA of Duncan’s multiple range test
At 19, 24, 29, and 34 days after anthesis, the transcription levels of the TaPDIL1-2 gene were determined by using qPCR, and the β-actin and GAPDH genes were used as two internal controls. Our results showed that transcription levels of the TaPDIL1-2 gene in grains of BSMV-TaPDIL1-2-inoculated spikes were markedly decreased by 42.2–79.2% (Fig. 4b, c), similar to previous studies [13, 29]. The starch contents and 1000-kernel weight in grains of BSMV-TaPDIL1-2-inoculated wheat spikes were increased by 16.6% and 6.8%, respectively (Fig. 4d, e), demonstrating that TaPDIL1-2 silencing caused enhanced starch biosynthesis in the wheat endosperm. Thus, TaPDIL1-2 might act as a negative regulator of starch biosynthesis. Similarly, the transcription levels of the TaAGPL1-1D gene in the endosperm of BSMV-TaPDIL1-2-infected wheat spikes at the four sampling time points increased remarkably (Fig. 4f, g). Because our previous studies have confirmed the important role of the TaAGPL1 gene in starch biosynthesis in wheat grain [11], we inferred that the TaPDIL1-2 gene could play a crucial role in wheat starch biosynthesis by negatively regulating TaAGPL1-1D gene expression.
Indirect regulation of TaPDIL1-2 gene on expression of TaAGPL1-1D gene
Most of PDIL proteins contain a KDEL at the C-terminus, and are localized in ER, where they combine with other protein substrates to form enzymatic complexes, in which their classical thioredoxin domains are responsible for interacting with substrates [30]. In the enzymatic complexes, PDIL proteins function as chaperones to bind to the unfolded or partially folded protein substrates by ascertaining their correct folding and preventing their aggregation [31]. Cereal endosperm storage proteins (e.g., gliadins and glutenins) have many disulfide bonds, which are bound to some PDIL proteins [32].
Transcription factors (TFs) function by binding to cis-acting elements preferentially located in the promoter region of the target genes [33], some TFs are also localized in ER to act as homo- or hetero-dimers with some chaperones (e.g. DnaK), where the cytoplasm-facing N-terminal regions are released by chaperones to be active TFs, and then, active TFs have been transferred from the ER into the nucleus, in which they activate the transcription of downstream genes [34]. For example, DRBEIII-1, a plant-specific TF in Brassica napus, was fused to a human PDIL protein to form a fusion protein complex (HDP), in which DREBIII-1 exhibited a highly soluble and biologically active form and HDP was confirmed to have the biological function of DRBEIII-1 [35]. Our sequence analysis indicated that the potential binding region was located from − 483 to − 1269 bp in the TaAGPL1-1D promoter containing the Auxin-responsive element and GCC-box-binding cis-acting elements (Additional file 8: Fig. S6). Previous studies have shown that a few PHD-, or AP2/ERF, bZIP-type TFs negatively regulate the expression of some starch biosynthesis genes [10, 36,37,38,39,40], and some members (e.g. bZIP28 and bZIP60) are localized in ER and possess the capacity to preferentially on the promoters of downstream target genes via binding to the above-mentioned cis-acting elements [41, 42]. Therefore, we speculated that functional TaPDIL1-2 protein could combine with TFs in ER to ascertain their correct folding to activate TFs, and then, active TFs are transferred into nucleus, in which they negatively regulate the expression of the TaAGPL1-1D gene. Based on these results, we propose one pathway for the TaPDIL1-2 chaperone to indirectly and negatively regulate the expression of the TaAGPL1 gene in starch biosynthesis (Fig. 5).

Schematic model of TaPDIL1-2 chaperone in a protein complex by modifying the folding of some transcription factors (TFs), which negatively regulates expression of the TaAGPL1 gene
Conclusions
We isolated the promoter of TaAGPL1-1DL gene and constructed a wheat endosperm cDNA library. Through a yeast one-hybrid screening by using the promoter of TaAGPL1-1DL gene as bait, and wheat grain cDNA library as prey, we further identified a protein disulfide isomerase (TaPDIL1-2), which bound to the promoter of TaAGPL1-1DL gene. By using BSMV-VIGS method in field experiments, we furthermore found that TaPDIL1-2 gene could negatively regulate the bread wheat starch biosynthesis.
Availability of data and materials
Please contact author for data requests.
Abbreviations
- AGPase:
-
ADP-glucose pyrophosphorylase
- BSMV-VIGS:
-
barley stripe mosaic virus-induced gene-silencing
- TaPDIL1-2:
-
wheat disulfide isomerase 1-2 protein
- G-1-P:
-
glucose-1-phosphate
- LSU or AGPL:
-
large/modulatory subunits of AGPase
- SSU or AGPS:
-
small/catalytic subunits of AGPase
- Y1H:
-
yeast one-hybrid
- IWGSC:
-
International Wheat Genome Sequencing Consortium
- qPCR:
-
quantitative real-time PCR
- GAPDH:
-
glyceraldehyde 3-phosphate dehydrogenase
- ANOVA:
-
one-way analysis of variance
- SD:
-
standard deviation
- ORF:
-
open reading frame
- TFs:
-
transcription factors
- ER:
-
endoplasmic reticulum
References
Morell MK, Rahman S, Abrahams SL, Appels R. The biochemistry and molecular biology of starch synthesis in cereals. Aust J Plant Physiol. 1995;22:647–60.
Jeon JS, Ryoo N, Hahn TR, Walia H, Nakamura Y. Starch biosynthesis in cereal endosperm. Plant Physiol Biochem. 2010;48:383–92.
Tetlow IJ, Davies EJ, Vardy KA, Bowsher CG, Burrell MM, Emes MJ. Subcellular localization of ADP glucose pyrophosphorylase in developing wheat endosperm and analysis of the properties of a plastidial isoform. J Exp Bot. 2003;54:715–25.
Burton RA, Johnson PE, Beckles DM, Fincher GB, Jenner HL, Naldrett MJ, Denyer K. Characterization of the genes encoding the cytosolic and plastidial forms of ADP-glucose pyrophosphorylase in wheat endosperm. Plant Physiol. 2002;130:1464–75.
Denyer K, Dunlap F, Thorbjørnsen T, Keeling P, Smith AM. The major form of ADP-glucose pyrophosphorylase in maize endosperm is extra-plastidial. Plant Physiol. 1996;112:779–85.
Rösti S, Rudi H, Rudi K, Opsahl-Sorteberg HG, Fahy B, Denyer K. The gene encoding the cytosilic small subunit of ADP-glucose pyrophosphorylase in barley endosperm also encodes the major plastidial small subunit in the leaves. J Exp Bot. 2006;57:3619–26.
Goren A, Ashlock D, Tetlow IJ. Starch formation inside plastids of higher plants. Protoplasma. 2018;255:1855–76.
Ohdan T, Francisco PB, Sawada JT, Hirose T, Terao T, Satoh H, Nakamura Y. Expression profiling of genes involved in starch synthesis in sink and source organs of rice. J Exp Bot. 2005;56:3229–44.
Radchuk VV, Borisjuk L, Sreenivasulu N, Merx K, Mock H, Rolletschek H, Wobus U, Weschke W. Spatiotemporal profiling of starch biosynthesis and degradation in the developing barley grain. Plant Physiol. 2009;150:190–204.
Kang G, Xu W, Liu G, Peng X, Guo T. Comprehensive analysis of the transcription of starch synthesis genes and the transcription factor RSR1 in wheat endosperm. Genome. 2013;56:115–22.
Kang G, Liu G, Peng X, Wei L, Wang C, Zhu Y, Ma Y, Jiang Y, Guo T. Increasing the starch content and grain weight of common wheat by overexpression of the cytosolic AGPase large subunit gene. Plant Physiol Biochem. 2013;73:93–8.
Kang G, Peng X, Wang L, Yang Y, Shao R, Xie Y, Ma D, Wang C, Zhu Y, Guo T, Zhu Y. Ultrastructural observation of mesophyll cells and temporal expression profiles of the genes involved in transitory starch metabolism in flag leaves of wheat after anthesis. Physiol Plant. 2015;153:12–29.
Liu G, Wu Y, Xu M, Gao TT, Wang P, Wang L, Guo TT, Kang G. Virus-induced gene silencing identifies an important role of the TaRSR1 transcription factor in starch synthesis in bread wheat. Int J Mol Sci. 2016;17:1557.
Yang YN, Li RG, Qi M. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 2000;22:543–51.
Wang K, Zhang X, Zhao Y, Chen F, Xia G. Structure, variation and expression analysis of glutenin gene promoters from Triticum aestivum cultivar Chinese Spring shows the distal region of promoter 1Bx7 is key regulatory sequence. Gene. 2013;527:484–90.
Jefferson RA, Kavanagh TA, Bevan MW. GUS fusions: betaglucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987;6:3901–7.
Borrill P, Ramirez-Gonzalez R, Uauy C. expVIP: a customisable RNA-seq data analysis and visualisation platform. Plant Physiol. 2016;170:2172–86.
Ma HZ, Liu GQ, Li CW, Kang GZ, Guo TC. Identification of the TaBTF3 gene in wheat (Triticum aestivum L.) and the effect of its silencing on wheat chloroplast, mitochondria and mesophyll cell development. Biochem Biophys Res Commun. 2012;426:608–14.
Lee WS, Rudd JJ, Kanyuka K. Virus induced gene silencing (VIGS) for functional analysis of wheat genes involved in Zymoseptoria tritici susceptibility and resistance. Fungal Genet Biol. 2015;79:84–8.
Li G, Wu Y, Liu G, Xiao X, Wang P, Gao T, Xu M, Han Q, Wang Y, Guo T, Kang G. Large-scale proteomics combined with transgenic experiments demonstrates an important role of jasmonic acid in potassium deficiency response in wheat and rice. Mol Cell Proteomics. 2017;16:1889–905.
International Wheat Genome Sequencing Consortium (IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science. 2018;361:661.
Kimura S, Higashino Y, Kitao Y, Masuda T, Urade R. Expression and characterization of protein disulfide isomerase family proteins in bread wheat. BMC Plant Biol. 2015;15:73.
Borrill P, Adamski N, Uauy C. Genomics as the key to unlocking the polyploid potential of wheat. New Phytol. 2015;208:1008–22.
Gao T, Li G, Wang C, Dong J, Yuan S, Wang Y, Kang G. Function of the ERFL1a transcription factor in wheat responses to water deficiency. Int J Mol Sci. 2018;19:1465.
Tufan HA, Stefanato FL, McGrann GRD, MacCormack R, Boyd LA. The barley stripe mosaic virus system used for virus-induced gene silencing in cereals differentially affects susceptibility to fungal pathogens in wheat. J Plant Physiol. 2011;168:990–4.
Ma M, Yan Y, Huang L, Chen M, Zhao H. Virus-induced gene-silencing in wheat spikes and grains and its application in functional analysis of HMW-GS-encoding genes. BMC Plant Biol. 2012;12:141.
Rong W, Qi L, Wang A, Ye X, Du L, Liang H, Xin Z, Zhang Z. The ERF transcription factor TaERF3 promotes tolerance to salt and drought stresses in wheat. Plant Biotechnol J. 2014;12:468–79.
Chen K, Li H, Chen Y, Zheng Q, Li B, Li Z. TaSCL14, a novel wheat (Triticum aestivum L.) GRAS gene, regulates plant growth, photosynthesis, tolerance to photooxidative stress, and senescence. J Genet Genom. 2015;42:21–32.
Scofield SR, Huang L, Brandt AS, Gill BS. Development of a virus-induced gene-silencing system for hexaploid wheat and its use in functional analysis of the lr21-mediated leaf rust resistance pathway. Plant Physiol. 2005;138:2165–73.
Edman JC, Ellis L, Blacher RW, Roth RA, Rutter WJ. Sequence of protein disulphide isomerase and implications of its relationship to thioredoxin. Nature. 1985;317:267–70.
Hayano T, Hirose M, Kikuchi M. Protein disulfide isomerase mutant lacking its isomerase activity accelerates folding in the cell. FEBS Lett. 1995;377:505–11.
Kim YJ, Yeu SY, Park BS, Koh HJ, Song JT, Seo HS. Protein disulfide isomeraselike protein 1-1 controls endosperm development through regulation of the amount and composition of seed proteins in rice. PLoS ONE. 2012;7:e44493.
Castrillo G, Turck F, Leveugle M, Lecharny A, Carbonero P, Coupland G, Paz-Ares J, Oñate-Sánchez L. Speeding cis–trans regulation discovery by phylogenomic analyses coupled with screening of an arrayed library of Arabidopsis transcription factors. PLoS ONE. 2011;6:e21524.
Fanata IWD, Lee SY, Lee KO. The unfolded protein response in plants: a fundamental adaptive cellular response to internal and external stresses. J Proteomics. 2013;93:356–68.
Liu Y, Zhao TJ, Yan YB, Zhou HM. Increase of soluble expression in Escherichia coli cytoplasm by a protein disulfide isomerase gene fusion system. Protein Expres Purif. 2005;44:155–61.
Fu FF, Xue HW. Coexpression analysis identifies rice starch regulator1, a rice AP2/EREBP family transcription factor, as a novel rice starch biosynthesis regulator. Plant Physiol. 2010;154:927–38.
Li H, Xiao Q, Zhang C, Du J, Li X, Huang H, Wei B, Li Y, Yu G, Liu H, Hu Y, Liu Y, Zhang J, Huang Y. Identification and characterization of transcription factor ZmEREB94 involved in starch synthesis in maize. J Plant Physiol. 2017;216:11–6.
Schmidt R, Schippers JHM, Mieulet D, Watanabe M, Hoefgen R, Guiderdoni E. Mueller-Roeber BSALT-RESPONSIVE ERF1 is a negative regulator of grain filling and gibberellin-mediated seedling establishment in rice. Mol Plant. 2014;7:404–21.
Liu C, Chen X, Ma P, Zhang S, Zeng C, Jiang X, Wang W. Ethylene responsive factor MeERF72 negatively regulates sucrose synthase 1 gene in cassava. Inter J Mol Sci. 2018;19:1281.
Ma P, Chen X, Liu C, Xia Z, Song Y, Zeng C, Li Y, Wang W. MePHD1 as a PHD-finger protein negatively regulates ADP-glucose pyrophosphorylase small subunit1a gene in cassava. Inter J Mol Sci. 2018;19:2831.
Yang ZT, Lu SJ, Wang MJ, Bi DL, Sun L, Zhou SF, Song ZT, Liu JX. A plasma membrane-tethered transcription factor, NAC062/ANAC062/NTL6, mediates the unfolded protein response in Arabidopsis. Plant J. 2014;79:1033–43.
Singh KB, Foley RC, Oñate-Sánchez L. Transcription factors in plant defense and stress responses. Curr Opin Plant Biol. 2002;5:430–6.
Acknowledgements
Not applicable.
Funding
This study was financially supported by the National Natural Science Foundation of China (31871550 and 31571575), the Henan Province Tackling Key Problems in Science and Technology (182102110102), and the Important Science and Technology Specific Project of Henan Province (151100110700).
Author information
Authors and Affiliations
Contributions
JD performed experimental works and data analysis; YZ, YF and JW participated in data analysis; YW and QZ designed the experiments; SY and YW participated experimental works; GK and GL wrote the article; GL, GK and XO provided the financial support. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Table S1.
The primer sequences used in this study.
Additional file 2: Fig. S1.
Isolated sequence of the TaAGPL1-1D promoter from bread wheat cv. Zhoumai 18.
Additional file 3: Fig. S2.
GUS histochemical staining and activity determination of the TaAGPL1-1DL promoter in N. benthamiana leaves.
Additional file 4: Fig. S3.
The identified quality of wheat grains cDNA library.
Additional file 5: Table S2.
The identified protein species via Y1H screening by using TaAGPL1-1D promoter as bait and wheat endosperm cDNA library as prey.
Additional file 6: Fig. S4.
The open reading frame (ORF) of the isolated TaPDIL1-2 gene.
Additional file 7: Fig. S5
The partial cDNA sequence of three homoeologs of the TaPDIL1-2 gene used for BSMV-VIGS experiment.
Additional file 8: Fig. S6.
The main cis-acting elements in partial fragment (786 bp, from − 483 to − 1269 bp) of TaAGPL1-1D promoter.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Dong, J., Zheng, Y., Fu, Y. et al. PDIL1-2 can indirectly and negatively regulate expression of the AGPL1 gene in bread wheat. Biol Res 52, 56 (2019). https://doi.org/10.1186/s40659-019-0263-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40659-019-0263-2