
Abstract
Background
Cocaine-and amphetamine regulated transcript (CART) is an endogenous neuropeptide, which is widespread in animals, plays a key role in regulation of follicular atresia in cattle and sheep. Among animal ovaries, CART mRNA was firstly found in the cattle ovaries. CART was localized in the antral follicles oocytes, granulosa and cumulus cells by immunohistochemistry and in situ hybridization. Further research found that secretion of E2 was inhibited in granulosa cells with a certain dose of CART, the effect depends on the stage of cell differentiation, suggesting that CART could play a crucial role in regulating follicle atresia. The objective of this study was to characterize the CART expression model and hormones secretion in vivo and vitro in pig follicle granulosa cells, preliminarily studied whether CART have an effect on granulosa cells proliferation and hormones secretion in multiparous animals such as pigs.
Methods
The expression levels of CART mRNA in granulosa cells of different follicles were analyzed using qRT-PCR technology. Immunohistochemistry technology was used to localize CART peptide. Granulosa cells were cultured in medium supplemented with different concentrations of CART and FSH for 168 h using Long-term culture system, and observed using a microscope. The concentration of Estradiol (E2) and progesterone (P) in follicular fluids of different test groups were detected by enzyme linked immunosorbent assay (ELISA).
Results
Results showed that expression level of CART mRNA was highest in medium follicles, and significantly higher than that in large and small follicles (P < 0.05). Immunohistochemical results showed that CART were expressed both in granulosa cells and theca cells of large follicles, while CART were detected only in theca cells of medium and small follicles. After the granulosa cells were cultured for 168 h, and found that concentrations of E2 increase with concentrations of follicle-stimulating hormone (FSH) increase when the CART concentration was 0 μM. And the concentration of FSH reached 25 ng/mL, the concentration of E2 is greatest. It shows that the production of E2 needs induction of FSH in granulosa cells of pig ovarian follicles. With the increasing of CART concentrations (0.01, 0.1, 1 μM), E2 concentration has a declining trend, when the FSH concentrations were 25 and 50 ng/mL in the medium, respectively.
Conclusions
These results suggested that CART plays a role to inhibit granulosa cells proliferation and E2 production, which induced by FSH in porcine ovarian follicular granulosa cells in vitro, but the inhibition effect is not significant. So we hypothesis CART maybe not a main local negative regulatory factor during porcine follicular development, which is different from the single fetal animals.
Similar content being viewed by others


Background
In mammals, ovarian follicular development is a continuous process during reproductive life span. Follicles develop through the primordial, primary, and secondary stage before acquiring an antral cavity, after which tertiary and preovulatory follicles successively form, and then oocytes are released after LH peak stimulation. In fact, only a few follicles undergo ovulation, most of the developing follicles will undergo atresia [1,2,3,4].
Ovarian follicles develop in a wave-like pattern in some species, such as pigs, human and cattle, each follicular wave is initiated by a transient elevation of FSH, then a cohort of follicles begin to grow, among which some follicles growing fast transform into the dominant follicles, while the rest become the subordinate follicles that will undergo atresia [5,6,7,8,9].
Cocaine-and amphetamine regulated transcript (CART), discovered initially by Douglass et al. via differential display RT-PCR analysis of brains of rats administered cocaine [10], is expressed mainly in central nervous system or neuronal origin cells [11], which is involved in a wide range of behaviors, such as regulation of food intake, energy homeostasis, and reproduction [12,13,14,15]. CART, as a potent anorectic peptide in hypothalamus, is regulated by leptin [16]. Since leptin stimulation of GnRH release can be blocked by CART protein antibody in vitro, the action of leptin on reproductive neuroendocrine axis may be mediated by CART [17]. As specific CART receptors have not been identified, the regulatory mechanisms of CART biological activities are still unknown.
Follicular growth and development is a complex process, which is not only precisely regulated by some endocrine factors, but also some locally produced intraovarian factors [3, 18,19,20]. A previous study reported that CART mRNA and CART peptide are expressed in bovine oocyte, and ovarian cells such as cumulus cells, and granulosa cell layer of antral follicles but not preantral follicles, suggesting a potential role of CART in the atresia of antral follicles [21]. Based on these findings, we hypothesized that CART may also play a role as a potential local regulator in the process of porcine follicular development. To investigate the relationship between CART and pig follicular development, we determined CART mRNA expression level in porcine follicles of different sizes and the concentrations of E2 and P in follicular fluid of those follicles, the localization of CART peptide was also detected by immunohistochemistry technology, and explored the effects of CART on granulosa cells proliferation and E2 secretion by in vitro culture. Our results indicated that CART may be involved in the process of porcine antral follicle development, yet its role may not be mediated by regulating the concentration of E2 and P.
Methods
Animal care
All animal procedures were performed with strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
Follicles and granulosa cells collection
Ten ovaries were collected from five female Large White pigs at the local slaughterhouse (Taigu, Shanxi, China). Follicles of 2–8 mm were dissected free from ovarian stroma and washed in 70% alcohol and DPBS solution for a few seconds. The follicles were classified into large (diameter > 5 mm), medium (3 mm < diameter < 5 mm) and small (diameter < 3 mm) groups, and placed in culture medium. Follicular fluid from each group follicles was aspirated and frozen on dry ice and stored at − 20 °C until hormone detection. Follicles were cut in half, and granulosa cells were collected from follicle internal wall and frozen in liquid nitrogen and stored at − 80 °C until RNA isolation.
RNA isolation and cDNA synthesis
Total RNA was isolated using Trizol (Takara, Dalian, China) according to the manufacturer’s instructions. Isolated RNA was dissolved in 30 µL of RNase free water. Before cDNA synthesis, 2 µL of total RNA were mixed with 2 µL of 5 × gDNA Eraser Buffer, 1 μL of gDNA Eraser (Takara, Dalian, China) and 5 μL of RNase free water and incubated at 42 °C for 2 min to remove genomic DNA. 0.8 μg RNA was mixed with 4 μL of 5 × PrimeScript® Buffer 2, 1 μL of RT Primer Mix, 1 μL of PrimeScript® RT Enzyme Mix I and 4 μL of RNase Free water (Takara, Dalian, China), and then incubated at 37 °C for 15 min followed by incubation at 85 °C for 5 s to synthesize cDNA, which was stored at − 20 °C until use.
Quantitative real-time PCR (qRT-PCR)
The relative expression level of CART mRNA in pig follicles of different sizes was measured by qRT-PCR. qRT-PCR was performed using 20 µL reaction volume containing 10 µL of SYBR® Green premix Ex Taq™II, 0.4 µL of ROX Reference Dye II(Takara, Dalian, China), 0.8 µL of forward and reverse primer, respectively, 2 µL of cDNA and 6 µL nuclease free water. Reactions were run on a 7500 Real Time PCR system (Thermo Scientific, Beijing, China) for 45 cycles at 95 °C for 15 s followed by 60 °C for 1 min. β-Actin gene was used as the endogenous control. Primers were designed using Primer 3 (http://primer3.ut.ee/), porcine CART primer was designed according to Sus scrofa CART mRNA, the primers are listed in Table 1. The relative mRNA expression level of CART was calculated using the comparative 2−ΔΔCT method [22]. The CART standard curve had a slope of − 3.124 (Eff. = 109.0%). The β-Actin standard curve had a slope of − 3.166 (Eff. = 106.9%).
Immunohistochemical localization of CART
Samples of adult ovary stroma were collected at a local abattoir from ovaries of three different animals. Samples were placed in a plastic tissue cassette, fixed with 4% (vol/vol) paraformaldehyde solution, and embedded in paraffin. Immunohistochemical localization of the CART peptide was performed using previously described procedures [23]. Rabbit anti-rat CART (55–102) polyclonal antisera (Phoenix Pharmaceuticals, Inc., Belmont, CA) (1:1000 dilution) was used in the analysis. Parallel controls were used, including sections incubated with a similar dilution of normal rabbit serum or rabbit anti-CART serum that had been pre-incubated overnight at 4 °C with 10 μg/mL rat CART (55–102) peptide (American Peptide Co., Sunnyvale, CA). Four serial sections from each sample were examined.
Granulosa cells cultured in vitro
Long-term culture system was performed using our previously procedures [24, 25]. Granulosa cells were cultured in a humidified environment of 5% CO2 and air at 37 °C for 168 h, medium was replaced with fresh medium every 48 h. Granulosa cells were harvested after termination of culture, washed by DPBS and digested using tryptase, and cell numbers were determined [26]. The situation of cells were observed and collected images after cultured for 48, 96, 144 and 168 h. At the end of culture, 340 μL medium were pooled from 2 adjacent wells per treatment, stored at − 20 °C for measurement of hormone.
Estradiol and progesterone enzyme-linked immunosorbent assay (ELISA)
Concentrations of E2 and P in follicular fluid of follicles at different sizes were detected using pig free estradiol and progesterone ELISA kit (Blue gene, Shanghai, China) according to the manufacturer’s instructions. The ELISA plates were read with a microplate reader (Thermo Scientific, Shanghai, China) to record the optical densities and the concentration of E2 and P derived from standard curve. The assay sensitivity was set as 0.5 pg/mL. The inter- and intra-assay CVs for E2 were 8.3 and 7.6%, respectively, and the inter- and intra-assay CVs for P were 9.2 and 5.8%, respectively.
The determination method of E2 concentration in medium was same to that in follicular fluid, standard curve was drew, concentration of each group was calculated。
Statistical analysis
Three biological reduplicates were used in all experiments mentioned above. The expression level of CART mRNA in granulosa cells of porcine follicles at different sizes and concentrations of E2 and P in follicular fluid were analyzed by one way ANOVA using SPSS computer software (IBM, USA). E2 concentration in culture medium was determined by Duncan-method of multiple comparisons. Data were presented as mean ± SE.
Results
Detection of CART mRNA in granulosa cells of porcine follicles
RT-PCR detection of CART mRNA and β-Actin gene in granulosa cells of porcine follicles at different sizes were shown through agarose gel electrophoresis. The products of β-Actin gene amplification in large, medium and small follicles were all 93 bp in size. CART mRNA amplification products in large, medium and small follicles were 102 bp in size.
The nucleotide sequence of the porcine CART and β-Actin cDNAs derived from RNA of follicular granulosa cells was 102 and 93 bp, respectively (data not shown). The nucleotide sequence of porcine CART shared 100% homology with that of Sus scrofa CART.
CART mRNA expression levels in large, medium and small follicles
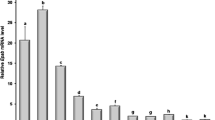
Quantitative real time PCR analysis revealed that the expression level of CART mRNA was significantly higher in medium follicles than that in large and small follicles (P < 0.05) (Fig. 1).

Quantitative real-time PCR analysis of CART mRNA expression in big, medium and small follicles. Superscript small letters indicate significantly different at the level of 0.05, same letters mean no significantly difference while different letters mean significantly difference. (n = 3 each; least square mean ± SEM)
Concentrations of E2 and P in follicular fluid of large, medium and small follicles
The concentrations of E2 and P in follicular fluid of large, medium and small follicles are showed in Fig. 2. No significant differences in the concentrations of each hormone (E2 and P) were observed in follicles of different sizes.

Estradiol and progesterone concentrations in big, medium and small follicles. Superscript small letters indicate significantly different at the level of 0.05, same letters mean no significantly difference while different letters mean significantly difference. (n = 3 each; least square mean ± SEM)
Intraovarian expression of CART protein
The intra-ovarian localization of CART peptide was determined by immunohistochemistry (Fig. 3). CART immunoreactivity was localized to the granulosa cells of big, medium and small follicles (Fig. 3b, e, h). Significant immunoreactivity in the granulosa cells was not detected when adjacent sections were incubated with normal rabbit serum (Fig. 3a, d, g) or when the CART antiserum was pre-absorbed with excess CART peptide (Fig. 3c, f, i).

Immunohistochemical localization of CART peptide within the porcine ovary. a–c, Adjacent section micrograph of big follicle; d–f, adjacent section micrograph of medium follicle; g–i, adjacent section micrograph of small follicle. a, d, g, Pre-immune serum; b, e, h, rabbit anti-rat CART; c, f, i, pre-absorbed rabbit anti-rat CART. GC, granulosa cell layer; TC, thecal cell layer; a–f: Magnification, ×400; scale bar, 20 µM
Response doses effect of CART on E2 production and granulosa cells proliferation under long-term culture with FSH treatment
Comparing with control group, with the concentrations of FSH were increasing (5, 25, 50 ng/mL) in medium, the concentrations of E2 are on the rise, when the CART concentration was 0 μM. And the concentration of FSH reached 25 ng/mL, the secretion of E2 is greatest. It shows that the production of E2 needs induction of FSH in granulosa cells of pig ovarian follicles. With the increasing of CART concentrations (0.01, 0.1, 1 μM), E2 concentration has a declining trend, when the FSH concentrations were 25 and 50 ng/mL in the medium, respectively, but the generation of E2 is significantly suppressed when pretreatment with 25 ng/mL FSH, 0.1 and 1 μM CART (P < 0.05) (Fig. 4).

Effects of CART on FSH induced estradiol production of pig follicular granulosa cells after 168 h in vitro culture. Superscript small letters and capital letters indicate significantly different at the level of 0.05 and 0.01, Values with the same letters were not significantly different and values with the different letters were significantly different at the level of 0.01 or 0.05. (n = 3 each; least square mean ± SEM)
The images of granulosa cells in vitro culture shows that clusters of granulosa cells were obviously decrease with the concentration of CART increase at FSH (25 ng/mL) in culture system (Fig. 5). This is consistent with the detection results of E2 (Fig. 4), indicating that decrease of granulosa cells numbers lead to the decrease of E2 secretion.

The micrograph of granulosa cells after 168 h in vitro culture with FSH (25 ng/mL) in culture system, a CART 0 µM; b CART 0.01 µM; c CART 0.1 µM; d CART 1 µM. Magnification, ×100
Discussion
The process of follicle growth and development is regulated by various endocrine factors and intra-ovarian factors. As our understanding of the well-established endocrine regulation of antral follicle growth and development increasing, attention in recent years has been focused on both identification and subsequent contribution of locally produced regulatory molecules that are involved in antral follicle growth and developmental regulation. Results of the present study demonstrated that the previously described anorectic neuropeptide CART had a higher expression level in the granulosa cells of medium follicles than that of big and small follicles. CART expression was potentially associated with follicle development. However, other results of the present study indicated that the concentrations of E2 and P in follicular fluid of large, medium and small follicles had no significant difference. Results support potential local regulatory role for CART in porcine follicular development without regulating the production of E2 and P.
Our results indicate that E2 secretion decrease with the increase of CART under same FSH concentration, suggested that CART might plays an suppressive role on the proliferation of pig ovarian follicle granulosa cells. Previous research found CART can restrain E2 secretion of bovine follicular granulosa cells in vitro, and plays a negative regulatory role during follicular development [27, 28]. Our study confirmed CART can inhibit the proliferation of pig ovarian follicle granulosa cells, meanwhile CART could promote granulosa cell apoptosis of porcine ovarian follicles [29]. But inhibition effect of CART is not significant.
Being classified as a somatostatin-like peptide, the amino acid sequence of CART peptide was first reported for sheep in hypothalamus [30]. Subsequently more and more researches were focused on the identity of CART. Until 1995 when Douglass and his co-workers [10] identified CART as a mRNA species whose expression increased acutely after psychomotor stimulant administration. The nucleotide and predicted amino acid sequences of CART from rat [10], human [31], mouse [32], sheep [33] and cattle [34] have been reported previously and were highly homologous among different species. However, the expression of CART mRNA in the porcine ovary has not been reported.
Previous studies have reported that CART was expressed in many tissues, including pituitary gland, adrenal gland [11, 35, 36], stomach [36], and intestines [36, 37]. However, reports about CART on mammalian gonads are rare. Murphy et al. [36, 38] did not detect immunoreactive CART peptide in rat ovaries and testis. Within gonadal tissues, CART expression has been reported in the goldfish ovary [38] and in nerves that innervate the epididymis of rat testis [39]. However, the present study detected CART mRNA and peptide expression in granulosa cells of porcine antral follicles. Smith, et al. [12] also found that both CART mRNA and protein were expressed in oocyte, cumulus cells, and granulosa cells of antral follicles in bovine ovary.
Our study demonstrated that the CART mRNA and peptide expressed in granulosa cells of porcine antral follicles at various development stages and the medium follicles had the highest expression level, suggesting that CART may play an important role in antral follicle development by regulating differentiation of granulosa cells. The exact mechanisms about how CART exerts its regulatory role in granulosa cells need to be further explored.
It is acknowledged that study design was not optimal due to follicles (large, medium, small) were used in CART mRNA expression and hormone determination. For the demarcation of follicles in different stages of multiparous animals, unlike single fetal animals whose follicles can be clearly divided into dominant and subordinate follicles. Despite such limitations, our results indicates that the concentrations of E2 and P in follicular fluid of large, medium and small follicles had no significant difference, which suggests that CART has no direct regulatory effect on the production of these two hormones in porcine antral follicles. Follicular growth and development are regulated by endocrine factors [40, 41], including E2 and P, which are important indicators to reflect the state of follicular growth. Smith et al. [12] demonstrated that CART had an inhibitory effect on in vitro production of E2 by granulosa cells.
Conclusion
CART leads to decrease of E2 production by inhibiting granulosa cells proliferation, which induced by FSH in porcine ovarian follicular granulosa cells. We hypothesis CART maybe not a main local negative regulatory factor during porcine follicular development, which is different from the single fetal animals.
References
McGee EA, Hsueh AJ. Initial and cyclic recruitment of ovarian follicles. Endocr Rev. 2000;21(2):200–14.
Barnett KR, Schilling C, Greenfeld CR, Tomic D, Flaws JA. Ovarian follicle development and transgenic mouse models. Hum Reprod Update. 2006;12(5):537–55.
Knight PG, Glister C. TGF-beta superfamily members and ovarian follicle development. Reproduction. 2006;132(2):191–206.
Dayal M, Sagar S, Chaurasia A, Singh U. Anti-Mullerian hormone: a new marker for ovarian function. J Obstet Gynaecol India. 2014;64(2):130–3.
Lucy MC, Liu J, Boyd CK, Bracken CJ. Ovarian follicular growth in sows. Reprod Suppl. 2001;58:31–45.
Baerwald AR, Adams GP, Pierson RA. Characterization of ovarian follicular wave dynamics in women. Biol Reprod. 2003;69(3):1023–31.
Lucy MC, Savio JD, Badinga L, De-La-Sota RL, Thatcher WW. Factors that affect ovarian follicular dynamics in cattle. J Anim Sci. 1992;70(11):3615–26.
Fortune JE. Ovarian follicular growth and development in mammals. Biol Reprod. 1994;50(2):225–32.
Ginther OJ. The theory of follicle selection in cattle. Domest Anim Endocrinol. 2016;57:85–99.
Douglass J, McKinzie AA, Couceyro P. PCR differential display identifies a rat brain mRNA that is transcriptionally regulated by cocaine and amphetamine. J Neurosci. 1995;15(3 Pt 2):2471–81.
Thim L, Kristensen P, Nielsen PF, Wulff BS, Clausen JT. Tissue-specific processing of cocaine-and amphetamine-regulated transcript peptides in the rat. Proc Natl Acad Sci USA. 1999;96(6):2722–7.
Smith SM, Vaughan JM, Donaldson CJ, Rivier J, Li C, Chen A, Vale WW. Cocaine-and amphetamine-regulated transcript activates the hypothalamic-pituitary-adrenal axis through a corticotropin-releasing factor receptor-dependent mechanism. Endocrinology. 2004;145(11):5202–9.
Boone EM, Hawks BW, Li W, Garlow SJ. Genetic regulation of hypothalamic cocaine and amphetamine-regulated transcript (CART) in BxD inbred mice. Brain Res. 2008;1194:1–7.
Lima FB, Henderson JA, Reddy AP, Tokuyama Y, Hubert GW, Kuhar MJ, Bethea CL. Unique responses of midbrain CART neurons in macaques to ovarian steroids. Brain Res. 2008;1227:76–88.
Derks NM, Gaszner B, Bernhardt K, Roubos EW, Kozicz T. Sex-specific expression of BDNF and CART in the midbrain non-preganglionic Edinger–Westphal nucleus in the rat. Peptides. 2009;30(12):2268–74.
Kristensen P, Judge ME, Thim L, Ribel U, Christjansen KN, Wulff BS, Clausen JT, Jensen PB, Madsen OD, Vrang N, Larsen PJ, Hastrup S. Hypothalamic CART is a new anorectic peptide regulated by leptin. Nature. 1998;393(6680):72–6.
Van Vugt DA, Lujan ME, Froats M, Krzemien A, Couceyro PR, Reid RL. Effect of fasting on cocaine-amphetamine-regulated transcript, neuropeptide Y, and leptin receptor expression in the non-human primate hypothalamus. Neuroendocrinology. 2006;84(2):83–93.
Demeestere I, Centner J, Gervy C, Englert Y, Delbaere A. Impact of various endocrine and paracrine factors on in vitro culture of preantral follicles in rodents. Reproduction. 2005;130(2):147–56.
Knight PG, Glister C. Local roles of TGF-beta superfamily members in the control of ovarian follicle development. Anim Reprod Sci. 2003;78(3–4):165–83.
Silva GM, Brito IR, Sales AD, Aguiar FL, Duarte AB, Araújo VR, Vieira LA, Magalhães-Padilha DM, Lima LF, Alves BG, Silveira LB, Lo Turco EG, Rodrigues AP, Campello CC, Wheeler MB, Figueiredo JR. In vitro growth and maturation of isolated caprine preantral follicles: influence of insulin and FSH concentration, culture dish, coculture, and oocyte size on meiotic resumption. Theriogenology. 2017;90:32–41.
Smith GW, Sen A, Folger JK, Ireland JJ. Putative role of cocaine- and amphetamine-regulated transcript (CARTPT) in dominant follicle selection in cattle. Soc Reprod Fertil Suppl. 2010;67:105–17.
Shi L, Zhao H, Ren Y, Yao X, Song R, Yue W. Effects of different levels of dietary selenium on the proliferation of spermatogonial stem cells and antioxidant status in testis of roosters. Anim Reprod Sci. 2014;149(3–4):266–72.
Bakke LJ, Li Q, Cassar CA, Dow MP, Pursley JR, Smith GW. Gonadotropin surge-induced differential upregulation of collagenase-1 (MMP-1) and collagenase-3 (MMP-13) mRNA and protein in bovine preovulatory follicles. Biol Reprod. 2004;71(2):605–12.
Sen A, Lv L, Bello N, Ireland JJ, Smith GW. Cocaine-and amphetamine-regulated transcript accelerates termination of follicle-stimulating hormone-induced extracellularly regulated kinase 1/2 and Akt activation by regulating the expression and degradation of specific mitogen-activated protein kinase phosphatases in bovine granulosa cells. Mol Endocrinol. 2008;22(12):2655–76.
Lv L, Jimenez-Krassel F, Sen A, Bettegowda A, Mondal M, Folger JK, Lee KB, Ireland JJ, Smith GW. Evidence supporting a role for cocaine and amphetamine regulated transcript (CART) in control of granulosa cell estradiol production associated with dominant follicle selection in cattle. Biol Reprod. 2009;81(3):580–6.
Folger JK, Jimenez-Krassel F, Ireland JJ, Lv L, Smith GW. Regulation of granulosa cell cocaine and amphetamine regulated transcript (CART) binding and effect of CART signaling inhibitor on granulosa cell estradiol production during dominant follicle selection in cattle. Biol Reprod. 2013;89(6):137.
Rosales-Torres AM, Avalos-Rodríguez A, Vergara-Onofre M, Hernández-Pérez O, Ballesteros LM, García-Macedo R, Ortíz-Navarrete V, Rosado A. Multiparametric study of atresia in ewe antral follicles: histology, flow cytometry, internucleosomal DNA fragmentation and lysosomal enzyme activities in granulosa cells and follicular fluid. Mol Reprod Dev. 2000;55(3):270–81.
Wood JR, Strauss JF. Multiple signal transduction pathways regulate ovarian steroidogenesis. Rev Endocr Metab Disord. 2002;3(1):33–46.
Austin EJ, Mihm M, Evans AC, Knight PG, Ireland JL, Ireland JJ, Roche JF. Alterations in intrafollicular regulatory factors and apoptosis during selection of follicles in the first follicular wave of the bovine estrous cycle. Biol Reprod. 2001;64(3):839–48.
Spiess J, Villarreal J, Vale W. Isolation and sequence analysis of a somatostatin-like polypeptide from ovine hypothalamus. Biochemistry. 1981;20(7):1982–8.
Douglass J, Daoud S. Characterization of the human cDNA and genomic DNA encoding CART: a cocaine- and amphetamine-regulated transcript. Gene. 1996;169(2):241–5.
Kuhar MJ, Adams S, Dominguez G, Jaworski J, Balkan B. CART peptides. Neuropeptides. 2002;36(1):1–8.
Barrett P, Morris MA, Moar KM, Mercer JG, Davidson JA, Findlay PA, Adam CL, Morgan PJ. The differential regulation of CART gene expression in a pituitary cell line and primary cell cultures of ovine pars tuberalis cells. J Neuroendocrinol. 2001;13(4):347–52.
Kobayashi Y, Jimenez-Krassel F, Li Q, Yao J, Huang R, Ireland JJ, Coussens PM, Smith GW. Evidence that cocaine- and amphetamine-regulated transcript is a novel intraovarian regulator of follicular atresia. Endocrinology. 2004;145(11):5373–83.
Koylu EO, Couceyro PR, Lambert PD, Ling NC, DeSouza EB, Kuhar MJ. Immunohistochemical localization of novel CART peptides in rat hypothalamus, pituitary and adrenal gland. J Neuroendocrinol. 1997;9(11):823–33.
Murphy KG, Abbott CR, Mahmoudi M, Hunter R, Gardiner JV, Rossi M, Stanley SA, Ghatei MA, Kuhar MJ, Bloom SR. Quantification and synthesis of cocaine- and amphetamine-regulated transcript peptide (79–102)-like immunoreactivity and mRNA in rat tissues. J Endocrinol. 2000;166(3):659–68.
Kasacka I, Piotrowska Z. Evaluation of density and distribution of CART-immunoreactive structures in gastrointestinal tract of hypertensive rats. BioFactors. 2012;38(6):407–15.
Volkoff H, Peter RE. Characterization of two forms of cocaine- and amphetamine-regulated transcript (CART) peptide precursors in goldfish: molecular cloning and distribution, modulation of expression by nutritional status, and interactions with leptin. Endocrinology. 2001;142(12):5076–88.
Dun NJ, Dun SL, Wong PY, Yang J, Chang J. Cocaine-and amphetamine-regulated transcript peptide in the rat epididymis: an immunohistochemical and electrophysiological study. Biol Reprod. 2000;63(5):1518–24.
Abdel-Ghani MA, El-Sherry TM, Abdelhafeez HH. Effect of growth differentiation factor-9 (GDF-9) on the progression of buffalo follicles in vitrified-warmed ovarian tissues. Reprod Domest Anim. 2016;51(5):795–803.
Juengel JL, Hudson NL, Berg M, Hamel K, Smith P, Lawrence SB, Whiting L, McNatty KP. Effects of active immunization against growth differentiation factor 9 and/or bone morphogenetic protein 15 on ovarian function in cattle. Reproduction. 2009;138(1):107–14.
Authors’ contributions
PL conceived the experiments and writed the initial draft of the manuscript; JJ and QH performed the experiments; JM and ZZ managed data collection; JY and LL designed the experiment and conducted data analysis. All authors read and approved the final manuscript.
Authors’ information
PF Li and ZW Zhu are doctors, associate professor of college of life science, Shanxi Agricultural University; JZ Meng is master, lecturer of Wujiang college, Tongren university; JJ Jing is doctoral candidate of College of Animal Science and Technology, Shanxi Agricultural University; QL Hao is graduate students of college of life science, Shanxi Agricultural University; JB Yao is doctor, professor of Division of Animal and Nutritional Sciences, West Virginia University, and the 100-Talent programme of Shanxi Agricultural University; LH Lyu is doctor, professor of College of Animal Science and Technology, Shanxi Agricultural University.
Acknowledgements
Authors are grateful of Prof. George W Smith and Prof. James Richard Pursley in Michigan State University in the USA for their directions of research designs and manuscript preparation.
Competing interests
We confirm that there are no known competing interests associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
Availability of data and materials
Availability of data and materials are included in the manuscript, figures and table.
Consent for publication
All authors read and approved the final manuscript, and consented for publication.
Ethics approval and consent to participate
We confirm that this study did not involve relevant clause of the Ethics Committee, and all animal procedures were performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
Funding
This study was supported by Shanxi Scholarship Council of China Grant No. 2014-key 5, Shanxi Sci-technological Collaboration Grant No. 201603D421006, Shanxi Talent Introduction and Sanjin Talent Program, Shanxi Provincial Talent Introduction and SXAU (Shanxi Agricultural University) Major Research Achievement Cultivation Grant No. zdpy 201403/201503 to Lyu; Shanxi Key Research and Development Plan (general) Agriculture Project Grant No. 201703D221020-1, SXAU Introduction of Doctor Research Startup Fund Grant No. 2014ZZ04 to Li; Chinese Natural Science Foundation Grant No. 31402156, SXAU Program for the Top Young Innovative Talents Grant No. TYIT201403 to Zhu.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Li, P., Meng, J., Jing, J. et al. Study on the relationship between expression patterns of cocaine-and amphetamine regulated transcript and hormones secretion in porcine ovarian follicles. Biol Res 51, 6 (2018). https://doi.org/10.1186/s40659-018-0154-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40659-018-0154-y