
Abstract
Background
Conserving migratory birds is challenging due to their reliance on multiple distant sites at different stages of their annual life cycle. The concept of “flyway”, which refers to all areas covered by the breeding, nonbreeding, and migrating of birds, provides a framework for international cooperation for conservation. In the same flyway, however, the migratory activities of the same species can differ substantially between seasons and populations. Clarifying the seasonal and population differences in migration is helpful for understanding migration ecology and for identifying conservation gaps.
Methods
Using satellite-tracking we tracked the migration of Whimbrels (Numenius phaeopus variegatus) from nonbreeding sites at Moreton Bay (MB) and Roebuck Bay (RB) in Australia in the East Asian–Australasian Flyway. Mantel tests were used to analyze the strength of migration connectivity between the nonbreeding and breeding sites of MB and RB populations. Welch’s t test was used to compare the migration activities between the two populations and between northward and southward migration.
Results
During northward migration, migration distance and duration were longer for the MB population than for the RB population. The distance and duration of the first leg flight during northward migration were longer for the MB population than for the RB population, suggesting that MB individuals deposited more fuel before departing from nonbreeding sites to support their longer nonstop flight. The RB population exhibited weaker migration connectivity (breeding sites dispersing over a range of 60 longitudes) than the MB population (breeding sites concentrating in a range of 5 longitudes in Far Eastern Russia). Compared with MB population, RB population was more dependent on the stopover sites in the Yellow Sea and the coastal regions in China, where tidal habitat has suffered dramatic loss. However, RB population increased while MB population decreased over the past decades, suggesting that loss of tidal habitat at stopover sites had less impact on the Whimbrel populations, which can use diverse habitat types. Different trends between the populations might be due to the different degrees of hunting pressure in their breeding grounds.
Conclusions
This study highlights that conservation measures can be improved by understanding the full annual life cycle of movements of multiple populations of Whimbrels and probably other migratory birds.
Similar content being viewed by others
Background
The annual life cycle of migratory birds depends on distant breeding, nonbreeding, and stopover sites as well as the interlinkage shaped by these sites (Newton 2008). The concept of “flyway”, which refers to all of the areas covered by breeding, nonbreeding, and migrating of birds, provides a framework for international cooperation in the conservation of migratory birds and their key areas (Boere and Stroud 2006). Over the past decades, field surveys have identified many key areas used by migratory birds, which has facilitated the designation of conservation hotspots along a flyway (e.g., Bamford et al. 2008). However, the temporal and spatial patterns of migration can be quite different between seasons and between populations of the same species in the same flyway (e.g., Battley et al. 2012; Hewson et al. 2016). Understanding the seasonal and population difference in migration activities can be helpful for detecting the factors linked with population dynamics, for finding conservation gaps, and for developing targeted conservation measures.
Geographical distribution greatly affects the temporal and spatial patterns of migratory populations. In general, the higher the latitude of the breeding site, the later the suitable time to initiate breeding (especially for those species breeding in the Arctic which must wait for snow melt), and thus the later the birds depart from nonbreeding sites and arrive at breeding sites (Conklin et al. 2013; Briedis et al. 2016). With an increase in nonstop flight distance, birds require a longer time to deposit more fuel to support migratory flight before departure (Conklin et al. 2013; Zhao et al. 2017). Moreover, migratory routes are closely linked with the availability and quality of stopover sites. The quality of stopover sites can affect the population dynamics of migratory birds. For example, a decline in the numbers of the Common Cuckoo (Cuculus canorus) that breed in the UK is related to the use of the western migration route, which provides low quality habitat (Hewson et al. 2016). In the East Asian–Australasian Flyway (EAAF), the rapid population decline of many migratory shorebirds is related to the dramatic habitat loss and degradation at their critical stopover sites in the Yellow Sea (Hua et al. 2015; Piersma et al. 2016; Studds et al. 2017).
Migration routes and stopover sites can also differ between the northward and southward migration of the same population. Weather conditions, especially wind conditions, influence the seasonal difference in migration activities (Alerstam et al. 2003). Because flying with a tailwind reduces fuel consumption and increases migration speed, birds prefer to use migration routes that provide tailwind assistance, and seasonal differences in wind direction and strength may therefore greatly affect migration routes and stopover decisions (e.g., Shamoun-Baranes et al. 2017; Carneiro et al. 2019). Migration can also be affected by the activities after arrival at the migration destination. Because breeding activities are time-constrained, migratory birds face great time pressure during migration to their breeding grounds. As a consequence, for boreal breeding species, birds generally exhibit shorter stopover durations, and thus faster migration speeds, during northward than southward migration (Battley et al. 2012; Nilsson et al. 2013; Zhao et al. 2017).
In recent years, many migratory shorebirds in the EAAF have suffered rapid population decline (Hua et al. 2015; Piersma et al. 2016; Studds et al. 2017). Clarifying the full annual life cycle of movements is the basis for identifying potential causes of these declines and conservation gaps. The Whimbrel (Numenius phaeopus) is a long-distance migratory shorebird with a worldwide distribution. In the EAAF, there are about 65,000 individual Whimbrels (Hansen et al. 2016), and these belong to the subspecies N. p. variegatus (Engelmoer and Roselaar 1998). They are mainly distributed in Australia and Southeast Asia during the nonbreeding period and in Far Eastern Russia during the breeding period. Roebuck Bay (RB) in Northwest Australia and Moreton Bay (MB) in East Australia are two major nonbreeding sites for the Whimbrels (Bamford et al. 2008; Rogers et al. 2011; Wilson et al. 2011). Since the 1990s, Whimbrels numbers have declined at MB (Wilson et al. 2011) but increased at RB (Rogers et al. 2019). The causes for these different population trends are unclear (Conklin et al. 2014).
Field surveys have revealed seasonal differences in numbers of Whimbrels at stopover sites along the EAAF. Many birds stopover in the Yellow Sea region during northward migration (Barter 2002; Bamford et al. 2008) but mainly stopover on the coasts of the Kamchatka Peninsula (Gerasimov et al. 2018), south China (Carey et al. 2001; Bai et al. 2015), Indonesia (Eaton et al. 2016) and Papua New Guinea (Bishop 2006) during southward migration (Conklin et al. 2014). However, few surveys were conducted on southward migration (Choi et al. 2016). Shorebird banding and flag resightings (Minton et al. 2006) indicate that the migration route used by the Whimbrels may be more eastward during southward than northward migration, i.e., more birds migrating south use the Kamchatka Peninsula rather than the Yellow Sea region as a stopover site (Minton et al. 2006). However, not many Whimbrels were flagged and these flagged birds mainly came from nonbreeding sites in East Australia. Moreover, few flagged birds were recovered due to lack of people looking for flags. The annual life cycle of Whimbrels in the EAAF therefore remains unclear.
The rapid development of bio-tracking technology facilitates the identification of the migration routes, key stopover sites, and migration connectivity of migratory birds. In this study, we tracked the nonbreeding Whimbrel populations at MB and RB to identify their breeding sites and stopover sites, and compare their seasonal and population differences in migration activities. We compare their migration with those of Whimbrels in the East Atlantic Flyway and the Americas Flyway, and we also discuss the potential causes of the differences in migratory activities.
Methods
Bird capture and tagging
Whimbrels were captured using cannon nets at high tide in MB and RB by the Queensland Wader Study Group (QWSG)/the Australasian Wader Studies Group (AWSG) respectively in the nonbreeding seasons of 2017‒2018. These were part of the banding programs of the Australasian Wader Studies Group (RB) and the Queensland Wader Study Group (MB). At MB, five birds were tagged with 5-g solar Platform Terminal Transmitters (PTTs, Microwave Telemetry, Inc., Columbia, Maryland, US) in November 2017. In February 2018 at RB, five birds were tagged with 5-g PTTs and 11 birds were tagged with 7-g solar Global Positioning System–Global System for Mobile Communication tags (GPS–GSMs, Hunan Global Messenger Technology Co., Ltd., Hunan, China). Tags were attached to the lower back of birds with leg-looped harnesses (Kuang et al. 2019). All of the tagged individuals were adults based on plumage characters (Prater et al. 1977). Relative to the body mass of the individual captured Whimbrels, the tag weight ranged from 1.1 to 2.0%.
The PTT duty cycle was set to 10 h on/48 h off. GPS–GSM tags were programed to make one record of location (fix) at 2–6 h intervals. Only the fixes with high positioning accuracy (error < 500 m for PTTs and < 100 m for GPS–GSM tags) were included in this study.
Data processing
All of the tagged Whimbrels initially remained near the capture site after release. We assumed that the birds had initiated their northward migration when they flew northward more than 50 km from the capture sites and did not return. We assumed that birds had arrived at the breeding site when they no longer moved northward and when two successive fixes were less than 50 km apart. Similarly, we assumed that birds had initiated their southward migration when they flew southward more than 50 km from the breeding sites and did not return. During migration, sites where birds remained within 0.5 degree of latitude for more than 48 h and where two successive fixes were less than 50 km apart were considered to be stopover sites. The geographical location of the nonbreeding site, stopover site, and breeding site for each individual was the arithmetic mean of all of the geographical coordinates at each site (Giunchi et al. 2019). Migration distance was calculated as the accumulation of the great circle distance between the nonbreeding, successive stopover, and breeding sites.
Departure time from and arrival time at the nonbreeding, stopover, and breeding sites was estimated for each individual bird. If the time interval between two consecutive fixes before and after departure from the site did not exceed 24 h, the last record at the site was used as the departure time. If the time interval between two consecutive fixes before and after arrival at the site did not exceed 24 h, the first record at the site was used as the arrival time. If the time interval between two consecutive fixes of departure from or arrival at the site exceeded 24 h, the departure time was calculated as the average time between the last fix at the site and the next fix after departure from the site, and the arrival time was calculated as the average time between the first fix at the site and the previous fix before arrival at the site (Giunchi et al. 2019).
Migration duration was calculated as the period from start to finish of the migratory flight. Stopover duration was the period between arrival at and departure from the stopover site. Overall stopover duration for each bird was the sum of all stopover durations during migration. Travel duration was calculated as the difference between the migration duration and the overall stopover duration. Migration speed was calculated as the migration distance divided by the corresponding migration duration. Travel speed was calculated as the migration distance divided by the travel duration.
Birds do not generally fly along the shortest path between the nonbreeding and breeding site. We calculated the detour index of migration route by dividing the great circle distance between nonbreeding and breeding site by the actual migration distance. Long-distance migratory birds deposit some amount of fuel before the first leg of the migratory flight (Zhao et al. 2017). Generally, birds flying a longer distance require to store more fuel prior to initiate migratory flight (Lindström 2020). To detect the contribution of the pre-migration fuel store to first migratory flight, we calculated the distance and duration of the first leg flight after departure from nonbreeding and breeding sites.
Data analysis
Of the 21 tagged Whimbrels, eight completed both the northward and southward migration (three from MB and five from RB), and another two from RB completed the northward but not the southward migration. Individuals failed to track were due to equipment failure or death of birds. For example, four birds with tags were resighted in the field but no data received, and one bird was killed in Yakut, Russia by local hunter, and two birds were lost when encountering typhoon in the East China Sea. We analyzed migration data from birds that completed at least a single trip, including 10 birds (7 with PTTs and 3 with GPS–GSMs) that completed northward migration and 8 birds (7 with PTTs and 1 with GPS–GSMs) that completed southward migration. Mantel tests were used to analyze the strength of migration connectivity between the nonbreeding and breeding sites of the two populations (Ambrosini et al. 2009). The pairwise distance matrix for each of the two populations was established based on the geographical location of the nonbreeding and breeding sites for each individual (Ambrosini et al. 2009). The distance matrix included all individuals that arrived at the breeding site during northward migration (seven individuals from RB and three individuals from MB). Distance between individuals was calculated as the great circle distance. The “ADE4” package in R was used to calculate the Mantel correlation coefficient (rM) for each population (Dray and Dufour 2007). The larger the coefficient, the stronger the migration connectivity.
A two-tailed Welch t-test was used to compare the migration activities of the RB and MB population in the same season. We calculated the logarithm of the seasonal ratio of migration speed, overall stopover duration, and travel speed for each individual in order to compare the seasonal difference in migration activities (Nilsson et al. 2013; Schmaljohann 2018). Linear mixed models (LMMs) were used to analyze the effect of travel speed and overall stopover duration on the seasonal difference in migration speed, with individual as a random variable. All statistical analyses were conducted with R software (R Core Team 2018), and results are reported as mean ± SD.
Results
Migration routes and stopover sites
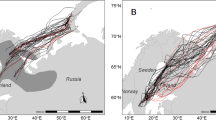
During their northward migration, Whimbrels from the same nonbreeding site followed a similar migration route for their first leg of flights after departure (Fig. 1a, b). The seven tagged birds from RB (Northwest Australia) flew over Southeast Asia to stopover sites on the south China coast, and then moved northward along the coast to the Yellow Sea region (Fig. 1a). The three tagged birds from MB (East Australia) flew over the Western Pacific to stopover sites on the Japan or Yellow Sea coast (Fig. 1b). The first leg flight from RB was 4791 ± 222 km, which represented 44% of the total northward migration distance (11,018 km). The first leg flight from MB was 7121 ± 412 km, which represented 55% of the total northward migration distance (12,991 km). Both the flight distance and duration were significantly longer (t = 9.23, df = 2.52 and p = 0.006 for flight distance; t = 2.38, df = 3.65 and p = 0.04 for duration) from MB than from RB (Table 1). The seven birds from RB flew different migration routes after departure from the Yellow Sea region, and their breeding sites were dispersed over a large region in Far Eastern Russia, i.e., from 105.1° to 173.3° E (Fig. 1a). In contrast, the three birds from MB flew a similar migration route and their breeding sites were concentrated in the east of Far Eastern Russia, from 167.7°–172.3° E (Fig. 1b). The migration connectivity (rM) was 0.39 and 0.99 for birds from RB and MB, respectively.

Nonbreeding sites (diamonds in Australia), northward (a, b) and southward (c, d) migration routes, stopover sites (filled in symbols), and breeding sites (pentagrams) of Whimbrels captured at Roebuck Bay (a, c) and Moreton Bay (b, d). Solid lines show the complete one-way migration routes, and dotted lines show incomplete one-way migration routes of individual birds
During southward migration, six of the seven birds from RB followed migration routes that were similar to their northward migration routes and stayed at one or more stopover sites in mainland China; in contrast, one of the seven RB birds, which had a breeding ground at the east of Far Eastern Russia, flew across the Western Pacific to Bismarck Archipelago and then returned to RB (Fig. 1c). Of the three birds from MB, one stopped over in the Yellow Sea, and two flew across the Western Pacific from the Kamchatka Peninsula to Bismarck Archipelago. All three flew back to MB after a stopover at Papua New Guinea (Fig. 1d). The first leg flight of the southward migration was 1292 and 2182 km for birds from MB and RB, respectively; these distances represented about 10% of the total migration distance for each population (Table 1).
None of the tagged birds paused their migration in Southeast Asia during their northward migration. The main stopover sites were located along the coast of China and Northeast Asia. Nine of the 10 tagged birds stopped over in the Yellow Sea region and the other one stopped over in Japan (Fig. 1a, b). During southward migration, most birds from the RB stopped over in the coastal region of China including the Yellow Sea region and Southeast Asia, except for the one RB bird that bred in the east of Far Eastern Russia (Fig. 1c). All three birds from MB, however, stopped over in Kamchatka and eastern Papua New Guinea/Bismarck Archipelago, but only one used the Yellow Sea and none used Southeast Asia as a stopover site (Fig. 1d). Birds stopped over one more time during southward (4.0 ± 1.2 times) than the northward (2.7 ± 1.3 times) migration (two populations combined).
Migration routes of the Whimbrels breeding in the East of Far Eastern Russia were further east and shorter during southward than the northward migration. All four birds used stopover sites in the Yellow Sea and adjacent region (Japan) during their northward migration, but three of the four birds took a shorter migration route, flying across the Western Pacific without a stopover in Asia during their southward migration.
A total of 67 stopover sites used by Whimbrels were identified in this study, including 29 sites during northward migration and 38 sites during southward migration. During northward migration, stopover sites mainly concentrated in the Yellow Sea and the South China while during south migration, stopover sites were dispersed along migration routes including southeast Asian where birds did not stopover during northward migration (Fig. 1, Additional file 1: Table S1).
Migration timing and stopover duration
During northward migration, there was no significant difference between the two populations in the departure date from nonbreeding sites (t = 1.01, df = 2.81, p = 0.39) or arrival date at breeding sites (t = 0.90, df = 6.78, p = 0.40). The migration distance was nearly 2000 km longer for birds from MB than for birds from RB (t = 2.51, df = 7.97, p = 0.04), but migration duration did not significantly differ between the two populations (t = 1.08, df = 7.014, p = 0.32). The detour index was slightly lower (t = 1.94, df = 7.68, p = 0.05) for birds from MB than for birds from RB. The overall migration speed was similar (t = 0.07, df = 5.34, p = 0.94) between the two populations (Table 1).
During southward migration, the two populations did not significantly differ in the departure date from the breeding sites (t = 0.78, df = 7.83 and p = 0.46) or the arrival date at the nonbreeding sites (t = 0.18, df = 4.67 and p = 0.87) (Table 1). Migration distance was about 1000 km longer for birds from MB than from RB, but migration duration did not significantly differ (t = 0.57, df = 4.48, p = 0.60) between the two populations. The detour index and overall migration speed were similar (t = 1.51, df = 5.07, p = 0.19) for the two populations (Table 1).
During northward migration, the overall stopover duration was longer (t = 2.84, df = 7.37, p = 0.01) for birds from MB than for birds from RB. Two of three birds from MB and six of seven birds from RB stopped over in the Yellow Sea region. Both stopover duration (t = 0.32, df = 2.78, p = 0.77) and percentage of the overall stopover duration (t = 0.06, df = 3.08, p = 0.95) in the Yellow Sea region were similar between the MB and RB birds (Fig. 1, Table 1).
During southward migration, the overall stopover duration was similar (t = 0.63, df = 4.58, p = 0.56) for birds from MB and RB. Four of the seven birds from RB stopped over in the Yellow Sea region for 35.5 ± 16.1 days, which accounted for 52.6 ± 29.3% of the overall stopover duration. The other three birds did not stopover in the Yellow Sea region. One of the three birds from MB stopped over in the Yellow Sea region; that bird remained at stopover sites for 33.5 days, which accounted for 53.3% of its overall stopover duration (Fig. 1, Table 1). None of the three MB birds stopped over in the coastal region of China while the RB birds stopped over for 28.8 ± 18.0 days, accounting for 48.2 ± 34.4% of the overall stopover duration.
Seasonal difference in migration activity
The overall migration speed of birds was faster (t = 9.51, df = 14.78, p < 0.001) during northward migration than during southward migration (Table 1). This was mainly due to the shorter migration duration and stopover duration during northward migration than during southward migration (Additional file 1: Table S2). There was no significant difference in migration distance (t = 0.37, df = 14.26, p = 0.72) or detour index (t = 0.55, df = 12.51, p = 0.59) between the northward migration and the southward migration (Table 1).
Discussion
This study provided new information on the migration of Whimbrels from two nonbreeding sites (RB and MB) in the EAAF. The dates for the start and end of the migration were similar for the two populations in both seasons, perhaps because the latitudes of breeding sites are similar for the two populations. Migration routes, stopover sites, and stopover duration, however, differed between the populations and between seasons. Moreover, the migration connectivity between nonbreeding and breeding sites was strong for the MB population (all birds nested in a narrow longitudinal band in Far East Siberia) but weak for the RB population (birds nested across a broad band spanning ~ 60 degrees of longitude). Seasonal and population differences in migration highlight the importance of identifying the full annual movements of multiple populations of the same species in the same flyway.
Migration of Whimbrels in the EAAF
Using band recovery and flag resighting data, Minton et al. (2006) suggested that, unlike most shorebirds in the EAAF that pass through the coasts of the Asian mainland during their northward migration, Whimbrels take an eastward migration route and their main stopover sites are located in the Kamchatka region of Far Eastern Russia and Japan. Field surveys also have indicated the importance of Kamchatka for Whimbrels on both northward (Lappo et al. 2012) and southward (Gerasimov et al. 2018) migration. This is consistent with our tracking results for Whimbrels from MB (located in East Australia) but inconsistent with our tracking results for Whimbrels from RB (located in Northwest Australia). It is not surprising because most of the banded and flagged Whimbrels were from East Australia (Minton et al. 2006). In the current study, we found that unlike the Whimbrels from MB, Whimbrels from RB depend on the stopover sites along the coasts of the Asian mainland during both their northward and southward migration. Actually, of the two recovered birds that originate from RB, one was recovered in the south Yellow Sea (Minton et al. 2006). As a consequence, the inconsistence might be due to relatively few birds being flagged in RB and lack of people looking for flagged birds along the coasts of the Asian mainland at the study period (Ma et al. 2013a). Results from our tracking study support speculation by Minton et al. (2006) that “there is only one report from the Yellow Sea in China but… this under-reflects the true importance of the Yellow Sea as a migratory stopover for this species”. Moreover, Minton et al. (2006) exhibited that all the breeding region of Whimbrels in Australia is related to Anadyr region in Far Eastern Russia. Our tracking study identified two new breeding regions (central-northern Sakha and eastern Krasnoyarskiy Kray) for Whimbrels in RB. These breeding regions fit well with the breeding distribution shown by Lappo et al. (2012).
We found that, during their northward migration, the Whimbrels that breed in the east of Far Eastern Russia tend to veer or “detour” westward to the Yellow Sea region (Fig. 1b). This has been found in several species that spend the non-breeding season in eastern Australia or New Zealand, such as the Grey-tailed Tattler (Tringa brevipes, Coleman et al. 2018), the Ruddy Turnstone (Arenaria interpres, Minton et al. 2011), and the Bar-tailed Godwit (Limosa lapponica baueri, Battley et al. 2012). The “detours” may be related to the use of stopover sites in the Yellow Sea region for fuel deposition. Suitable breeding period is short for shorebirds breeding at high latitudes; arriving at breeding sites in good condition is therefore important so that breeding can be quickly initiated. Many studies have indicated that shorebirds in the Yellow Sea region during their northward migration deposit huge amounts of fuel that can be used for both the migratory flight and activities at breeding sites (e.g., Hua et al. 2013; Ma et al. 2013b). During their southward migration, in contrast, birds can restore their body condition after arriving at nonbreeding sites.
Whimbrels from Australia are believed to breed mainly in Far Eastern Russia (Barter 2002; Bamford et al. 2008). However, the migration connectivity between nonbreeding and breeding populations is still unclear. Although sample sizes were small, this study showed that the Whimbrels from MB breed in a relatively small region in the southeast of Chukchi, while the breeding areas of the Whimbrels from RB are more widely distributed in Far Eastern Russia. This indicates that the migration connectivity is stronger for Whimbrels from MB than from RB. All eight individuals that were successfully tracked for their round-trip migration returned to the nonbreeding sites in the previous year, suggesting that Whimbrels exhibit high fidelity to their nonbreeding sites. Further analysis of the genetic traits of birds at various breeding and nonbreeding sites should help clarify the migration connectivity and gene flow in the flyway.
N. p. variegatus was considered to be the only subspecies in the EAAF with breeding at the east of Far Eastern Russia (Bamford et al. 2008; Conklin et al. 2014). In this study, the breeding grounds of most tracked birds match the breeding range of variegatus but new breeding regions (central-northern Sakha and eastern Krasnoyarskiy Kray) were identified for RB birds. These regions have been suggested to be the overlap zone of subspecies phaeopus and variegatus (Engelmoer and Roselaar 1998) and the nature of intergradation is unclear. Although our tracking results cannot identify whether there is intergradation between the two subspecies and whether phaeopus and variegatus might even be different species (Livezey 2010), we found RB Whimbrels have a widespread breeding region in Far Eastern Russia. Combining with migratory tracking, comparison on the morphological and genetic traits of birds at both breeding and nonbreeding sites will help to clarify the issue in taxonomy.
We detected obvious seasonal difference in the use of stopover sites. During northward migration, birds took a long-distance, nonstop flight after departing from nonbreeding sites, and their stopover sites were therefore concentrated in the second half of their migratory journey, i.e., along the coasts of China, Japan, and the Yellow Sea region. During southward migration, stopover sites were widely distributed on the Kamchatka Peninsula, the Yellow Sea region, and the coasts of China, southeast Asia, and Papua New Guinea and the Bismarck Archipelago (Fig. 1c, d). This is consistent with the results from field surveys (Barter 2002; Hadden 2004; Bishop 2006; Bamford et al. 2008; Bai et al. 2015) and reported in McClure (1974). Our tracking results confirm field observations by Gerasimov et al. (2018), for Whimbrels that breed in East Siberia, the Kamchatka Peninsula is an important gathering site during southward migration before their cross-sea southward flight. Our tracking results also indicated that the Kamchatka Peninsula was less used during the northern migration. Field surveys have reported that about 51% (Gerasimov et al. 2018) to 91% (Schuckard et al. 2006) of the Whimbrels in the EAAF stopped over on the Kamchatka Peninsula during southward migration, which is much more than birds recorded during northward migration, although a large number of birds also can be recorded on northward migration (Lappo et al. 2012).
Most of stopover sites the Whimbrels used in this study have been identified as the sites of international importance (Bamford et al. 2008; Conklin et al. 2014) but some new recognized sites have not yet (Additional file 1: Table S1). During northward migration, the newly recognized sites were concentrated in the south coast and the northeast of China, where RB birds did stopover at. During southward migration, the newly recognized sites were concentrated in the southwest coast of Kamchatka, Papua New Guinea and Bismarck Archipelago, south coast of China, and Sulawesi in Indonesia (Additional file 1: Table S1). Although large flocks of Whimbrels have been reported in the southeast Asia on southward migration and nonbreeding period (Conklin et al. 2016), the detailed sites recorded by our tracking are different from former records. Field surveys in these regions are required to clarify the importance of these sites for the Whimbrels and other shorebirds.
The first leg of the migratory flight during northward migration is far for both MB and RB populations, i.e., the leg covers about half of the total migration distance. This is also the case for some other shorebirds that spend the nonbreeding period in Oceania and that breed in the Arctic and subarctic (Additional file 1: Table S3). Fuel deposition rate of shorebirds is relatively low at low latitudes where there is less food (macrobenthos on tideland) than at high latitudes (Piersma et al. 2004). To improve their fuel deposition rate, shorebirds can take a long-distance, nonstop flight to “jump over” the low latitudes (Piersma et al. 2004; Aharon-Rotman et al. 2016). This requires high-quality habitats with abundant food to support rapid fuel deposition pre-migration at nonbreeding sites. During their northward migration, birds from MB exhibited longer first leg flights than birds from RB, suggesting that birds from MB deposit more fuel than birds from RB before the onset of migration. This inference is supported by the body mass data at the two sites: there was no significant difference in body mass between the two populations from October to December; while at pre-migration period in March, body mass of the Whimbrels at MB was significantly higher than that at RB (Additional file 2: Figure S1). We therefore suggest that high quality habitats at nonbreeding sites are important for long-distance migration.
We found RB population intensively used China’s coast including the Yellow Sea during both northward and southward migration, while MB population less used during southward migration. This is similar to the migration of Bar-tailed Godwits in the EAAF: menzbieri Godwits (mainly stay in Northwest Australia in nonbreeding season) use the Yellow Sea on both migrations while baueri Godwits (mainly stay in New Zealand and East Australia in nonbreeding season) use the Yellow Sea only on northward migration (Battley et al. 2012). Menzbieri population suffered more rapid decline than baueri population; this is likely due to their different dependence on the stopover sites in the Yellow Sea, where the critical tidal habitats suffered dramatic loss over the past several decades (Conklin et al. 2016). However, Whimbrel populations have declined at MB (Wilson et al. 2011) but increased at RB in the recent few decades (Rogers et al. 2019). This is the opposite of the situation of Godwits. This may be because the Whimbrels can use a variety of habitat types (including widely distributed farmland and shrub) during migration (Kuang et al. 2019), thus tidal habitat loss along China’s coast had limited impact on the Whimbrels. Recent study indicated that huge numbers of Whimbrels were killed by local hunters in Kamchatka (37,000 birds per year, Khlokov et al. 2020), suggesting MB birds that breed in the east of Far Eastern Russia suffered serious hunting pressure. Further studies on the survival rates at different periods and sites in the full annual life cycle (e.g., Piersma et al. 2016) are important to clarify the different trends between the two populations.
Migration of Whimbrels in different flyways
Recent studies have indicated that Whimbrels that breed in North America and Iceland use only a limited number of stopover sites during migration. They generally make one stopover during their northward migration, and Icelandic Whimbrels can fly directly from the breeding to nonbreeding site during their southward migration (Alves et al. 2016; Johnson et al. 2016; Carneiro et al. 2019, Additional file 1: Table S4). Whimbrels in the EAAF, in contrast, stopover for an average of three and four times during their northward and southward migration, respectively. This might be related to the longer migration distances of the Whimbrels in the EAAF (11,000 and 13,000 km for birds from MB and RB, respectively) than in the other two flyways (9800 and 6700 km for birds in North America and Iceland, respectively, Additional file 1: Table S4). Wind conditions (tailwinds or headwinds) in different regions can also affect stopover decisions during migration (Alerstam et al. 2003; Ma et al. 2011; Carneiro et al. 2019). Whimbrels that breed in Iceland and North America are likely to obtain wind assistance (Alves et al. 2016; Johnson et al. 2016; Carneiro et al. 2019) while birds in the EAAF might encounter unfavorable tailwinds near the equator (Wang et al. 2020) and in the south hemisphere during southward migration (Gill et al. 2014). The adverse wind condition can increase the possibility of stopover of birds during migratory flight (Ma et al. 2011).
Because the fuel deposition rate at stopover sites is much slower than the fuel consumption rate during flight, stopover duration is much longer than the duration of migratory flight, and thus migration speed is mainly determined by stopover duration (Kölzsch et al. 2016; Schmaljohann 2018). In the EAAF, Whimbrels have shorter stopover durations and thus faster migration speeds during their northward than their southward migration. This is consistent with results obtained for most migratory birds and is related to the time pressure for early arrival at breeding sties during northward migration (Newton 2008; Nilsson et al. 2013). Whimbrels that breed in Iceland and North America, however, have a shorter stopover duration and thus a faster migration speed during their southward than northward migration (Alves et al. 2016; Johnson et al. 2016; Carneiro et al. 2019, Additional file 1: Table S4). It is likely that Whimbrels that breed in Iceland and North America have a greater chance of obtaining tailwind assistance during their southward than northward migration. This benefits the migratory flights from breeding to nonbreeding sites (Alves et al. 2016; Johnson et al. 2016; Carneiro et al. 2019). The relatively short migration distance of Whimbrels breeding in Iceland also enables those birds to make a nonstop flight from the breeding to the nonbreeding site.
In response to the narrow breeding time window at high latitudes, some birds adopt a capital breeding strategy, i.e., they deposit nutrients that form the eggs before arriving at breeding sites (Hobson and Jehl 2010) and can therefore lay eggs soon after arriving at breeding sites. Capital breeders generally have a relatively long stopover duration en route (especially at final pre-breeding stopover sites), which enables them to deposit more fuel than required for the migratory flight (Hua et al. 2013). Whimbrels that breed in North America at least partially adopt a capital breeding strategy (Hobson and Jehl 2010). They spend 64 days at the breeding site and 34 days at stopover sites during their northward migration, and there is only an 11-day interval between arriving at the breeding site and laying eggs (Johnson et al. 2016). In contrast, Whimbrels that breed in Iceland tend to be income breeders (Carneiro et al. 2019). They have a short stopover duration during their northward migration (12 days), remain for a long period at the breeding site (95 days), and exhibit a long interval between arriving at the breeding site and laying eggs (22 days) (Carneiro et al. 2019). The current study indicates that Whimbrels in the EAAF have an average stopover duration of 26 days during northward migration and remain at the breeding site for 59 days (Table 1, Additional file 1: Table S4), which is similar to the Whimbrels in North America. As a consequence, we speculate that Whimbrels in the EAAF at least partially adopt a capital breeding strategy and that the conservation of key stopover sites that enable fuel deposition en route is critical for their population maintenance.
Conclusions
This study identified the migration routes and breeding areas as well as revealed the migration activities of two Whimbrel populations in the EAAF. We found there are obvious seasonal and population differences in migration activities within the EAAF and among flyways. This might be related to the different environmental conditions among populations, such as geographical distribution of breeding and nonbreeding sites, characteristics of migration routes and stopover sites, as well as weather conditions during migration. Environmental conditions shape the migration performance of migratory birds, which is also closely related to their physiological and ecological traits during migration as well as pre- and post-migration. We highlight that clarifying the full annual life cycle of movements of multiple populations is the basis for understanding the adaption to migratory life and for making conservation measures to migratory birds. In addition, many studies have highlighted the impact of habitat loss and degradation at stopover sites on migratory shorebirds along the EAAF (e.g., Melville et al. 2016; Piersma et al. 2016; Studds et al. 2017). Results of this study suggest that hunting pressure in breeding sites might be an important but neglected cause of population decline.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Aharon-Rotman Y, Gosbell K, Minton C, Klaassen M. Why fly the extra mile? Latitudinal trend in migratory fuel deposition rate as driver of trans-equatorial long-distance migration. Ecol Evol. 2016;6:6616–24.
Alerstam T, Hedenström A, Åkesson S. Long-distance migration: evolution and determinants. Oikos. 2003;103:247–60.
Alves JA, Dias MP, Méndez V, Katrínardóttir B, Gunnarsson TG. Very rapid long-distance sea crossing by a migratory bird. Sci Rep. 2016;6:38154.
Ambrosini R, Møller AP, Saino N. A quantitative measure of migratory connectivity. J Theor Biol. 2009;57:203–11.
Bai QQ, Chen JZ, Chen ZH, Dong GT, Dong JT, Dong WX, et al. Identification of coastal wetlands of international importance for waterbirds: a review of China Coastal Waterbird Surveys 2005‒2013. Avian Res. 2015;6:12.
Bamford M, Watkins D, Bancroft W, Tischler G, Wahl J. Migratory shorebirds of the East Asian—Australasian Flyway: population estimates and internationally important sites. Canberra: Wetlands International-Oceania; 2008.
Barter MA. Shorebirds of the Yellow Sea: importance, threats and conservation status. Canberra: Wetlands International-Oceania; 2002.
Battley PF, Warnock N, Tibbitts TL, Gill RE, Piersma T, Hassell CJ, et al. Contrasting extreme long-distance migration patterns in bar-tailed godwits Limosa lapponica. J Avian Biol. 2012;43:21–32.
Bishop KD. Shorebirds in New Guinea: their status, conservation and distribution. Stilt. 2006;50:103–34.
Boere GC, Stroud DA. The flyway concept: what it is and what it isn’t. In: Boere GC, Galbraith CA, Stroud DA, editors. Waterbirds around the World. Edinburgh: The Stationery Office; 2006. p. 40–7.
Briedis M, Hahn S, Gustafsson L, Henshaw I, Träff J, Král M, et al. Breeding latitude leads to different temporal but not spatial organization of the annual cycle in a long-distance migrant. J Avian Biol. 2016;47:743–8.
Carey GJ, Chalmers ML, Diskin DA, Kennerley PR, Leader PJ, Leven MR, et al. The avifauna of Hong Kong. Hong Kong: Hong Kong Bird Watching Society; 2001.
Carneiro C, Gunnarsson TG, Alves JA. Faster migration in autumn than in spring: seasonal migration patterns and non-breeding distribution of Icelandic Whimbrels Numenius phaeopus islandicus. J Avian Biol. 2019;50:e01938.
Choi CY, Rogers KG, Gan XJ, Clemens RS, Bai QQ, Lilleyman A, et al. Phenology of southward migration of shorebirds in the East Asian–Australasian Flyway and inferences about stop-over strategies. Emu. 2016;116:178–89.
Coleman JT, Milton DA, Hitoshi A. The migratory movements of Grey-tailed Tattler Tringa brevipes from Moreton Bay. Stilt. 2018;72:2–8.
Conklin JR, Battley PF, Potter MA, Fox JW. Breeding latitude drives individual schedules in a trans-hemispheric migrant bird. Nat Commun. 2013;1:67.
Conklin JR, Verkuil YI, Smith BR. Prioritizing migratory shorebirds for conservation: action on the East Asian–Australasian Flyway. Hong Kong: WWF Hong Kong; 2014.
Conklin JR, Lok T, Melville DS, Riegen AC, Schuckard R, Piersma T, et al. Declining adult survival of New Zealand Bar-tailed Godwits during 2005–2012 despite apparent population stability. Emu. 2016;116:147–57.
Dray S, Dufour AB. The ade4 package: implementing the duality diagram for ecologists. J Stat Softw. 2007;22:1–20.
Eaton JA, van Balen B, Brickle NW, Rheindt FE. Birds of the Indonesian Archipelago. Barcelona: Lynx Edicions; 2016.
Engelmoer M, Roselaar CS. Geographic variation in Waders. Dordrecht: Kluwer Academic Publishers; 1998.
Gerasimov Y, Tiunov I, Matsyna A, Tomida H, Bukhalova A. Waders southward migration studies on west Kamchatka. Stilt. 2018;72:9–14.
Gill RE, Douglas DC, Handel CM, Tibbitts TL, Hufford G, Piersma T. Hemispheric-scale wind selection facilitates bar-tailed godwit circum-migration of the Pacific. Anim Behav. 2014;90:117–30.
Giunchi D, Baldaccini NE, Lenzoni A, Luschi P, Sorrenti M, Cerritelli G, et al. Spring migratory routes and stopover duration of satellite-tracked Eurasian Teals Anas crecca wintering in Italy. Ibis. 2019;161:117–30.
Hadden D. Birds and bird-lore of Bougainville and the North Solomons. Alderly: Dove Publications; 2004.
Hansen BD, Fuller RA, Watkins D, Rogers DI, Clemens RS, Newman M, et al. Revision of the East Asian-Australasian Flyway population estimates for 37 listed migratory shorebird species. Unpublished report for the department of the environment. Melbourne: Birdlife Australia; 2016.
Hewson CM, Thorup K, Pearce-Higgins JW, Atkinson PW. Population decline is linked to migration route in the common cuckoo. Nat Commun. 2016;7:12296.
Hobson KA, Jehl JR Jr. Arctic waders and the capital-income continuum: further tests using isotopic contrasts of egg components. J Avian Biol. 2010;41:565–72.
Hua N, Piersma T, Ma ZJ. Three-phase fuel deposition in a long-distance migrant, the red knot (Calidris canutus piersmai), before the flight to High Arctic breeding grounds. PLoS ONE. 2013;8:e62551.
Hua N, Tan K, Chen Y, Ma ZJ. Key research issues concerning the conservation of migratory shorebirds in the Yellow Sea region. Bird Conserv Int. 2015;25:38–52.
Johnson AS, Perz J, Nol E, Senner NR. Dichotomous strategies? The migration of Whimbrels breeding in the eastern Canadian sub-Arctic. J Field Ornithol. 2016;87:371–83.
Khlokov K, Gerasimov Y, Syroechkovskiy E. First attempt to evaluate hunting pressure on shorebirds in Kamchatka: progress report. Spoon-billed Sandpiper Task Force News Bull. 2020;22:31–4.
Kölzsch A, Müskens GJDM, Kruckenberg H, Glazov P, Weinzierl R, Nolet BA, et al. Towards a new understanding of migration timing: slower spring than autumn migration in geese reflects different decision rules for stopover use and departure. Oikos. 2016;125:496–507.
Kuang FL, Wu W, Ke WJ, Ma Q, Chen WP, Feng XS, et al. Habitat use by migrating Whimbrels (Numenius phaeopus) as determined by bio-tracking at a stopover site in the Yellow Sea. J Ornithol. 2019;160:1109–19.
Lappo EG, Tomkovich PP, Syroechkovski EE. Atlas of breeding waders in the Russian Arctic. Moscow: The Russian Geographical Society; 2012.
Lindström Å. Migration tracks of waders: avoiding the pitfalls of speed estimates and inferred strategies. Wader Study. 2020;127:2–3.
Livezey BC. Phylogenetics of modern shorebirds (Charadriiformes) based on phenotypic evidence: analysis and discussion. Zool J Linn Soc. 2010;160:567–618.
Ma ZJ, Hua N, Zhang X, Guo HQ, Zhao B, Ma Q, et al. Wind conditions affect stopover decisions and fuel stores of shorebirds migrating through the south Yellow Sea. Ibis. 2011;153:755–67.
Ma ZJ, Cheng YX, Wang JY, Fu XH. The rapid development of birdwatching in mainland China: a new force for bird study and conservation. Bird Conserv Int. 2013a;23:259–69.
Ma ZJ, Hua N, Peng HB, Choi CY, Battley PF, Zhou QY, et al. Differentiating between stopover and staging sites: functions of the southern and northern Yellow Sea for long-distance migratory shorebirds. J Avian Biol. 2013b;44:504–12.
McClure HE. Migration and survival of the birds of Asia. Bangkok: SEATO; 1974.
Melville DS, Chen Y, Ma ZJ. Shorebirds along the Yellow Sea coast of China face an uncertain future: a review of threats. Emu. 2016;116:100–10.
Minton C, Wahl J, Jessop R, Hassell C, Collins P, Gibbs H. Migration routes of waders which spend the non-breeding season in Australia. Stilt. 2006;50:135–57.
Minton C, Gosbell K, Johns P, Christie M, Klaassen M, Hassell C, et al. Geolocator studies on Ruddy Turnstones Arenaria interpres and Greater Sandplovers Charadrius leschenaultii in the East Asian-Australasia Flyway reveal widely different migration strategies. Wader Study Group Bull. 2011;118:87–96.
Newton I. The migration ecology of birds. London: Academic Press; 2008.
Nilsson C, Klaassen RHG, Alerstam T. Differences in speed and duration of bird migration between spring and autumn. Am Nat. 2013;181:837–45.
Piersma T, Rogers DI, González P, Zwarts L, Niles LJ, de Lima I, et al. Fuel storage rates in red knots worldwide: facing the severest ecological constraint in tropical intertidal environments? In: Marra PP, Greenberg R, editors. Birds of two worlds. Washington: Smithsonian Institution Press; 2004. p. 262–73.
Piersma T, Lok T, Chen Y, Hassell C, Yang HY, Boyle A, et al. Simultaneous declines in summer survival of three shorebird species signals a flyway at risk. J Appl Ecol. 2016;53:479–90.
Prater AJ, Marchant JH, Vuorinen J. Guide to the identifcation and ageing of holarctic waders. Tring: BTO; 1977.
R Core Team. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2018. https://www.R-project.org/.
Rogers DI, Hassell CJ, Boyle A, Gosbell K, Minton C, Rogers KG, et al. Shorebirds of the Kimberley Coast—populations, key sites, trends and threats. J R Soc West Aust. 2011;94:377–91.
Rogers DI, Scroggie MP, Hassell CJ. Long-term monitoring of shorebirds in north Western Australia. Arthur Rylah Institute for Environmental Research, Technical Report 313; 2019.
Schmaljohann H. Proximate mechanisms affecting seasonal differences in migration speed of avian species. Sci Rep. 2018;8:4106.
Schuckard R, Huettmann F, Gosbell K, Geale J, Kendal S, Gerasimov Y, et al. Shorebird and gull census at Moroshechnaya Estuary, Kamchatka, Far East Russia, during August 2004. Stilt. 2006;50:34–46.
Shamoun-Baranes J, Liechti F, Vansteelant WMG. Atmospheric conditions create freeways, detours and tailbacks for migrating birds. J Comp Physiol A. 2017;203:509–29.
Studds CE, Kendall BE, Murray NJ, Wilson HB, Rogers DI, Clemens RS, et al. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat Commun. 2017;8:14895.
Wang Y, Yu Y, Zhang Y, Zhang HR, Chai F. Distribution and variability of sea surface temperature fronts in the south China sea. Estuar Coast Shelf Sci. 2020;240:106793.
Wilson HB, Kendall BE, Fuller RA, Milton DA, Possingham HP. Analyzing variability and the rate of decline of migratory shorebirds in Moreton Bay, Australia. Conserv Biol. 2011;25:758–66.
Zhao M, Christie M, Coleman J, Hassell C, Gosbell K, Lisovski S, et al. Time versus energy minimization migration strategy varies with body size and season in long-distance migratory shorebirds. Mov Ecol. 2017;5:23.
Acknowledgements
This paper owes a great debt to the late Clive Minton for his great contribution to shorebird study and conservation. We thank the Australasian Wader Studies Group, Queensland Wader Study Group, and the Northwest Australia Expedition 2018 team for their support for fieldwork. We appreciate Roz Jessop, Michael Dawkins, Prue Wright, Robert Bush, Brad Woodworth, Chi-Yeung Choi, Bingrun Zhu and many other volunteers for their assistance in the fieldwork. AWSG acknowledges the Yawuru People via the offices of Nyamba Buru Yawuru Limited for permission to catch birds on the shores of Roebuck Bay, traditional lands of the Yawuru people. CJH thanks his funders, WWF Netherlands, Spinoza Premium of Netherlands Organisation Prize for Scientific Research to Theunis Piersma and MAVA (Foundation Pour La Nature). The authors also wish to acknowledge the support of the Australian Bird and Bat Banding Scheme (ABBBS) for the provision of Banding licenses and bands to the A-Class banders involved in this research. We thank two anonymous reviewers for their comments and suggestions on an earlier version of the manuscript.
Funding
This study was financially supported by the National Key Research and Development Program of China (2018YFC1406402), the National Natural Science Foundation of China (31830089 and 31772467) and World Wide Fund for Nature Beijing Office (10003881).
Author information
Authors and Affiliations
Contributions
ZM and FK designed the study. JTC, CJH, KSKL, and GM conducted the fieldwork and deployed tags on birds. FK and ZM analyzed the data and led the paper writing with contributions from all authors. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Catching, tagging, and banding were carried out under the license of the Australian Bird and Bat Banding Scheme (ABBBS). Catching birds on the shores of Roebuck Bay was conducted with the permission of the Yawuru People. The MB Whimbrels were tagged under license CVL1337 issued to JTC by the Department of National Parks, Sport and Racing, Queensland Government and Department of Primary Industries Animal Ethics license CA 2015-03-845. The RB Whimbrels were tagged under Regulation 17 Licence 08-00741-3 authorised to CJH from the Department of Biodiversity, Conservation and Attractions.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Supplementary information
Additional file 1: Table S1.
Stopover sites used by the Whimbrels from MB (Moreton Bay) and RB (Roebuck Bay) during northward and southward migration. Table S2. Results of linear mixed model testing for the effect of overall stopover duration (Rstopover duration) and travel speed (Rtravelled speed) on the overall migration speed (Rmigration speed) of Whimbrels. Table S3. Distance and duration of the first-leg flights during northward migration of shorebirds in Australia and New Zealand. Table S4. Migration parameters (means) of Whimbrels in the East Asian–Australasian Flyway (EAAF, nonbreeding at Moreton Bay (MB) and Roebuck Bay (RB) in Australia), the Eurasian-Africa Flyway (breeding in Iceland), and the America Flyway (breeding in Manitoba, Canada).
Additional file 2: Figure S1.
Changes in the body mass of the Whimbrels at Moreton Bay (red circles) and Roebuck Bay (blue circles) during the nonbreeding period.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kuang, F., Coleman, J.T., Hassell, C.J. et al. Seasonal and population differences in migration of Whimbrels in the East Asian–Australasian Flyway. Avian Res 11, 24 (2020). https://doi.org/10.1186/s40657-020-00210-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-020-00210-z