
Abstract
Background
The effects of selective pulmonary vascular tone alterations on cardiac preload have not been previously examined. Therefore, we evaluated whether changing pulmonary vascular tone either by hypoxia or the inhalation of aerosolized prostacyclin (PGI2) altered intrathoracic or pulmonary blood volume (ITBV, PBV, respectively), both as surrogate for left ventricular preload. Additionally, the mean systemic filling pressure analogue (Pmsa) and pressure for venous return (Pvr) were calculated as surrogate of right ventricular preload.
Methods
In a randomized controlled animal study in 6 spontaneously breathing dogs, pulmonary vascular tone was increased by controlled moderate hypoxia (FiO2 about 0.10) and decreased by aerosolized PGI2. Also, inhalation of PGI2 was instituted to induce pulmonary vasodilation during normoxia and hypoxia. PBV, ITBV and circulating blood volume (Vdcirc) were measured using transpulmonary thermo-dye dilution. Pmsa and Pvr were calculated post hoc. Either the Wilcoxon-signed rank test or Friedman ANOVA test was performed.
Results
During hypoxia, mean pulmonary artery pressure (PAP) increased from median [IQR] 12 [8–15] to 19 [17–25] mmHg (p < 0.05). ITBV, PBV and their ratio with Vdcirc remained unaltered, which was also true for Pmsa, Pvr and cardiac output. PGI2 co-inhalation during hypoxia normalized mean PAP to 13 (12–16) mmHg (p < 0.05), but left cardiac preload surrogates unaltered. PGI2 inhalation during normoxia further decreased mean PAP to 10 (9–13) mmHg (p < 0.05) without changing any of the other investigated hemodynamic variables.
Conclusions
In spontaneously breathing dogs, changes in pulmonary vascular tone altered PAP but had no effect on cardiac output, central blood volumes or their relation to circulating blood volume, nor on Pmsa and Pvr. These observations suggest that cardiac preload is preserved despite substantial alterations in right ventricular afterload.
Similar content being viewed by others

Background
Pulmonary vascular tone (PVT) plays a central role in the regulation of cardiac preload and afterload, and may influence cardiac output (CO) by influencing venous return (VR) to the left ventricle (LV) and hereby determines LV preload. Simultaneously, PVT influences afterload of the right ventricle (RV). While VR was recognized as an important determinant of cardiac output (CO) decades ago by relating right atrial pressure (RAP) to LV output [1,2,3,4], it is important to consider that the RV receives blood from the systemic circulation, whereas the LV receives blood from the pulmonary circulation. The pulmonary blood volume compartment may therefore be regarded a surrogate measure of LV preload, which can be estimated reliably [5] by transpulmonary dilution as intrathoracic or pulmonary blood volume (ITBV and PBV, respectively, Fig. 1).The pulmonary circulation is a very compliant and low-pressure vascular bed [6, 7]. PVT can be altered artificially in both directions, e.g., by hypoxia or aerosolized prostacyclin (PGI2). Hypoxia increases PVT through hypoxic pulmonary vasoconstriction (HPV) [6], increasing pulmonary artery pressure (PAP). PGI2 administration instead, decreases PVT through relaxing of the vascular smooth muscle [8]. Interestingly though, the effects of selective alterations in PVT on the central blood volume compartment remains unknown to a large extent, as previous studies used isolated organ models only or solely focused on pressure-effects [9,10,11,12].

Schematic overview of the central blood volume compartment as determined by thermo-dye dilution. After injection of ice-cold indocyanine green in the right atrium, the dye dilution curves that are derived from the fiberoptic catheters placed in the pulmonary artery and ascending aorta, allow the calculation of PBV (green) and ITBV (blue and green combined. Modified from [5]
In addition, not only the pulmonary blood volume compartment itself, but also its distribution with the peripheral blood volume compartment might be influenced by alterations in PVT [13]. Therefore, the cumulative effect on the central blood compartment in terms of surrogate measures of (biventricular) cardiac preload, afterload and CO, remains elusive too.
Therefore, in this experimental study in spontaneously breathing dogs, we altered PVT in both directions by hypoxia and aerosolized prostacyclin, in order to gain more insight in the cumulative effects on ITBV and PBV as surrogate measures of LV preload. Since it is most likely that RV preload will be influenced as well, we calculated the pressure gradient between the right atrium and the more “upstream” side of the systemic circulation (mean systemic filling pressure (Pmsf) [2, 14,15,16,17], as a RV preload surrogate, in addition to mean PAP—resembling RV afterload.
Finally, to gain more insights in compensatory mechanisms in an intact circulation, the distribution between the PBV, ITBV and the (systemic) circulating blood compartment (Vd circ) has been studied as well.
Methods
Animals and instrumentation
Experiments were performed at the animal laboratory of the Department of Experimental Anesthesiology of the University of Düsseldorf, Germany. Reporting of the study was set up in accordance with ARRIVE 2.0 guidelines [18]. After approval from the local district governmental animal investigation committee (North-Rhine Westphalia in Dusseldorf, Germany; registered as Ref. 23.05-2303-84/96), six adult Foxhound dogs (3 males, 3 females, median age 38 months) were treated according to the principles of the National Institute of Health guidelines for animal care (NIH publication nr 86–23, revised 1985), the same dogs on which we previously reported [5].
The dogs were raised and housed in the Animal Research Laboratory of the Heinrich-Heine University (Düsseldorf, Germany). Relevant surgical procedures for preparation—performed several weeks before the experiments—have been described extensively before [5] and involved implanting an ultrasound transit-time flow probe (16–20 mm S-series with silicone shielded U-reflector, Transonic Systems, NY, USA) around the pulmonary artery for continuous recording pulmonary blood flow and cardiac output (COTransonic). Two catheters were placed in the ascending aorta for blood sampling and arterial blood pressure measurement, and a fiberoptic thermistor probe (4F, Pulsiokath PV2024, Pulsion Medical Systems, Munich, Germany) was inserted to measure indocyanine green (ICG) plasma concentration and blood temperature. In addition, a 7F fiberoptic thermodilution catheter (Arrow International, Reading, MA) was introduced into a pulmonary artery via a dog’s hindlimb under fluoroscopy prior to each experiment. After completion of the experiments, the dogs were kept in the research facility until they died of natural causes.
Measurements
Mean arterial blood pressure (MAP), mean PAP and central venous pressure (CVP) were measured at the level of the heart and simultaneously recorded on an eight-channel polygraph (model RS 3800, Gould, Cleveland, OH, USA) and a cassette data recorder (model XR-5000, TEAC®, Tokyo, Japan). Pulmonary capillary wedge pressure (PCWP) was determined intermittently. Partial arterial oxygen pressure (PaO2) and oxygen saturation (SaO2) were obtained by blood gas analysis (ABL3, Radiometer, Copenhagen, Denmark). COtransonic was calibrated in vivo using the direct Fick principle using O2 uptake (Deltatrac II Metabolic Monitor), and the arterial to mixed venous oxygen content difference as measured by a galvanic cell (Lex-O2-CON-TL)—as described previously [19].
The determination of Vd circ, PBV and ITBV has been described in detail previously [5]. In short, blood volumes were determined using transpulmonary thermo-dye dilution by bolus injection of ice-cold ICG (< 5 °C, 0.2 mg kg−1). Here, ITBV represents the volume of blood between the injection site (right atrium) and recording site (ascending aorta), while PBV represents the volume of blood between the right atrium and aortic valve (Fig. 1). ITBV was calculated as: COtransonic * mean transit time (mtt) of the ice-cold bolus between the respective injection and recording site. PBV was calculated similarly, yet the mtt was derived by deconvolution of the dye dilution curve that was based on a pulmonary transport function [20]. Vdcirc was calculated as: COtransonic * mtt of the overall circulation in a 30 min time period, as fitted by an aortic dye dilution curve over a recirculation model. The ratios between central and systemic circulating blood volumes (ITBV/Vd circ and PBV/Vd circ) were calculated afterwards.
Pmsf was estimated mathematically using a Pmsf analogue (Pmsa) [15, 21] offline using a validated algorithm [15] incorporating MAP, COTransonic and CPV. Pmsa was calculated using the formula:
\({\text{Pmsa }} = \, a \, *{\text{ CVP }} + \, b \, *{\text{ MAP }} + \, c \, *{\text{ CO}}\). Here, a + b = 1 (a = 0.96 and b = 0.04, reflecting the contribution of venous and arterial compartments), c reflects an assessment of resistance and is determined on age, weight and height and was determined according to previous studies in dogs [22]. Pressure for venous return (Pvr) was calculated as: \({\text{Pvr }} = {\text{ Pmsa }}{-}{\text{ CVP}}\), heart efficiency (Eh) as: Eh = Pvr/Pmsa, and resistance for venous return (RVR) as: RVR = Pvr/CO.
Experimental program
Anesthesia was performed using pentobarbital (20 mg kg−1 injection, followed by a continuous infusion of 4 mg kg−1 h−1). The dogs were breathing room air spontaneously.
We increased PVT by lowering inspiratory oxygen concentration by adding nitrogen (AGA Linde, Medical Gases) to a plastic hood fixed above the dog’s head and upper trunk. The flow rate was adjusted to reduce FiO2 to about 0.1. FiO2 was measured continuously via a gas probe fixed in the middle of the hood above the dog’s head (Capnomancy® Ultima SV, Datex-Engstrom, Helsinki, Finland). PVT was decreased by adding aerosolized prostacyclin (PGI2) to the inspiratory gases: 10 μg/ml crystalline PGI2 (Flolan®, GlaxoWellcome, Hamburg, Germany) solution was instilled into the chamber of an ultrasound nebulizer (Siemens Elema, Solna, Sweden) by an infusion pump (Perfusor, Braun, Melsungen, Germany), which was connected to the orifice of a tube, inserted into the trachea prior to this intervention to minimize absorption of PGI2 at the upper airways. Aerosolized PGI2 was inhaled during normoxia (FiO2 0.21) or during hypoxia (FiO2 0.1). The nebulizer gas flow (pressurized air and nitrogen) was adjusted (range 6–10 L min−1) to achieve and maintain a reduction in mean pulmonary arterial pressure of 10% below baseline. The mean dose of aerosolized PGI2-solution was approximately 7.5 µg kg−1 min−1. To assure hemodynamic stability, the dogs were observed for 30 min after introduction of the catheters. The intervention sequence started with PGI2 administration, followed by hypoxia and ended with PGI2 co-administration (n = 10). In three sequences, hypoxia was administered only, without PGI2 (co-) administration due to logistic reasons. Before each intervention, a separate baseline measurement was performed to serve as baseline for that particular intervention (Fig. 2).

Schematic overview of the order of intervention and the number of interventions performed
Interventions lasted approximately 30 min each. Measurements were performed at the end of each intervention during steady state. There were no relevant data available to estimate effect sizes of interventions, so an a priori sample size calculation was not performed.
Statistics
Statistical analysis was performed using SigmaPlot 13.0 (Systat Software, San Jose, USA). COTransonic values were normalized for body weight. Continuous data were assumed non-normally distributed. Analyses were performed pairwise: data from interventions were compared with preceding baseline measurements to account for within-subject variability. The Wilcoxon-signed rank test was used for pairwise comparisons. In case of three consecutive measurements, the Friedman ANOVA was used, and the Dunn’s test was applied for post hoc testing. Statistical significance was set at P-values < 0.05.
Results
A total of 33 interventions were performed in 6 dogs on 13 different study days. PVT was increased by the administration of a hypoxic gas mixture, with FiO2 ranging from 0.08 to 0.12, which was temporarily combined with the inhalation of aerosolized PGI2 in ten of these interventions. In 10 interventions, PVT was solely reduced using inhaled aerosolized PGI2, without simultaneous administration of a hypoxic gas mixture. No adverse events occurred during any of the interventions.
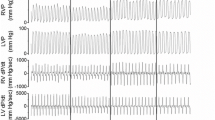
A typical example is shown in Fig. 3.

Original registration in an anesthetized spontaneously breathing dog. After 2 heart cycles, recording speed was reduced and tracings of COTransonic and pulmonary arterial pressure (PAP) were switched to mean values. At the end of each intervention, blood volumes were determined in duplicate. During hypoxia, PAP increased markedly, whereas pulmonary capillary wedge pressure (PCWP), and pulmonary blood volume (PBV) remained unchanged. *: PCWP was determined intermittently at three moments. **: a zero-adjustment was performed at three moments
Hypoxia
During hypoxia mean PAP increased from a median (interquartile range) value of 12 (8–15) mmHg at normoxia to 19 (17–25) mmHg (p < 0.05; Table 1) during hypoxia. MAP increased from 85 (82–91) mmHg during normoxia, to 93 (90–97) mmHg during hypoxia (p < 0.05). SV decreased from 27 (25–33) mL during normoxia to 23 (19–27) mL during hypoxia (p < 0.05). There was no change in CO. heart rate (HR) increased from 90 (82–97) bpm during normoxia to 115 (102–130) bpm during hypoxia (p < 0.05). Neither CVP nor PCWP changed during hypoxia (Table 1).
There was no change in both PBV and ITBV, neither during normoxia, nor during hypoxia (Fig. 4 and Table 1). Moreover, both Vdcirc, and the ratio between Vdcirc and PBV, and ITBV, respectively, remained unaltered as well as Pmsa and derived variables (Table 1).

A Boxplot in which the absolute values of ITBV (green) and PBV (blue) are shown at baseline (n = 13), during hypoxia (n = 13), and during inhalation of aerosolized PGI2 (n = 10) while the dogs were still exposed to hypoxia. B Boxplot in which the absolute values of PBV (blue) and ITBV (green) are shown at baseline (n = 10) and during subsequent administration of PGI2 (n = 10)
Inhaled aerosolized PGI2
Aerosolized PGI2 inhalation under normoxia decreased mean PAP from 13 (11–14) mmHg to 10 (9–13) mmHg, p < 0.05. HR decreased from 89 (83–97) bpm to 84 (78–93) bpm (p < 0.05). Other conventional hemodynamic variables remained unchanged after administering PGI2, as shown in Table 1.
There were no changes in PBV, ITBV, Vdcirc (Table 1, Fig. 4B), nor was there a change in VD circ or its ratio with PBV or ITBV. This was also true for Pmsa and derived variables (Table 1).
Upon co-administration of aerosolized PGI2 during hypoxia, the increase in mean PAP was reversed (i.e., from a median value of 19 mmHg back to 13 mmHg, p < 0.05). MAP and HR increased to 95 (92–106) mmHg and 105 bpm, respectively, compared to baseline values (p < 0.05). Other conventional hemodynamic variables remained unaltered (Table 1).
There were again no changes in any of the measured blood volumes, neither did the ratio between either PBV or ITBV with Vd circ change (Fig. 4A, Table 1). This was also true for the calculated values of Pmsa and its derived variables, except for a slight increase in Pvr.
Discussion
In this experimental study in spontaneously breathing dogs, hypoxia substantially increased PVT. SV decreased, which was compensated by an increase in HR so that ultimately CO remained constant. Preload was maintained too, both for the LV (PBV, ITBV) and for the RV (Pvr) and the distribution of central and circulating blood volume remained unaltered. Therefore, it appears that an increase in PAP as a surrogate of RV afterload does not impair CO in healthy dogs, has no measurable effect on left and right cardiac preload, and does not alter the distribution of blood within the intrathoracic and systemic circulation. In addition, reducing PVT by endobronchial PGI2, neither changed CO, nor changed any of the surrogate measures of preload, and finally neither altered the distribution of blood between the central blood volume compartment and circulating blood volume compartment.
Effects of hypoxia on the pulmonary circulation
HPV substantially increased mean PAP. HPV predominantly entails precapillary vascular smooth muscle contraction, and is a well-recognized phenomenon in mammals that helps to match regional perfusion and ventilation [7]. While being a protective native reflex, HPV may be particularly dangerous in subjects who cannot tolerate the associated (sudden) increase in PAP, e.g., those with reduced (right) ventricular function or an intracardiac shunt. E.g., pulmonary atelectasis with associated regional hypoxia—inducing HPV—may preserve systemic oxygenation at the cost of an increase in RV afterload [23].
We observed that the increase in mean PAP during hypoxia was associated with an increase in both HR and MAP. Likely, this observation can be attributed to an increased sympathetic tone [24]. The increase in HR allowed CO maintenance during hypoxia, despite a decrease in SV. It remains to be elucidated whether SV decreased secondary to the increase in afterload or hypoxia-induced myocardial ischemia (albeit no electrocardiographic signs of ischemia were noted). Most importantly, the increase in mean PAP following hypoxia was not accompanied by a change in PBV and ITBV, indicating that LV preload remained preserved—a subject that has not been investigated in an intact circulation before. Previous studies that were performed in isolated animal lung preparations were inconclusive as central blood volume was either reduced [11], preserved [12], or increased [25]—in response to HPV.
Secondarily, Pmsa and Pvr remained constant as well during hypoxia, suggesting that the increase in RV afterload did not affect RV preload. As Pmsa (resembling the net balance between systemic vascular tone and volume) remained constant, we conclude that hypoxia did not change systemic vascular tone and volume, and consequently did not change RV preload, as resembled by a constant Pvr. The unaltered heart efficiency (EH; Pvr divided by Pmsa) additionally shows that CO was maintained and could “overcome” VR in spite of the increase in RV afterload, with a maintained pressure gradient between the (right) heart and the returning venous blood. Hence, the resistance to venous return (RVR), in which CVP acts as an opposing pressure for generating CO, remained unaltered. In other words: despite the sudden hypoxia-induced increase in RV afterload, the heart was able to maintain RAP (or CVP) low, in order to preserve VR to the right heart.
It must be stressed that PBV and ITBV were considered as surrogate measures of cardiac preload and we did not record functional measures of LV preload, e.g., stroke volume variation. Still, we looked for changes in these blood volumes, and the net conclusion from the above-mentioned mechanisms predominantly support the hypothesis that RV and LV preload remained unaltered.
Finally, as we simultaneously investigated the influence of changes in PVT on Vdcirc, we could demonstrate absence of impact on the distribution between PBV, ITBV and Vdcirc. We speculate that increases in PVT can be “easily” countered by the high compliance of the pulmonary circulation.
The effects of PGI2
PGI2 is a potent vasodilator that can be applied systemically (i.v.) and endobronchially (aerosolized PGI2) [26, 27]. After endobronchial administration, aerosolized PGI2 produces selective pulmonary vasodilation, i.e., PAP is selectively lowered without lowering systemic arterial pressure [28]. To the best of our knowledge, the effects of selective pulmonary vasodilation (e.g., by PGI2 administration) on blood volumes have not been studied before in an in vivo model with an intact circulation. In this study, we observed a minor effect of PGI2 inhalation on PAP and MAP without any effect on measures of central blood volume, Pmsa and Pvr. Interestingly however, PGI2 administration during hypoxia allowed mean PAP to return to pre-hypoxic values, suggesting that hypoxia “unmasks” the pulmonary vasodilatory effects of PGI2 and resolves HPV despite ongoing hypoxia: it may be argued that PGI2 has no further vasodilatory influence on PVT under normal conditions.
Additionally, we observed that the distribution between PBV, ITBV and Vdcirc were unaffected by aerosolized PGI2, which contrasts previous research where prostacyclines were administered systemically for malignant hypertension treatment in humans (n = 7) [28]. Although Pvr decreased subtly during PGI2 co-administration during hypoxia, which may suggest a very slight increase in RV preload, this observation is probably of negligible clinical significance. While SV was preserved, CO remained constant as well, although it may be speculated that if a larger sample size was obtained, the (trend-like) changes in CO following PGI2 inhalation may have turned statistically significant. Finally, it might be speculated that PGI2 inhalation directly causes a reduction in heart rate. To the best of our knowledge, there is no literature available to support this assumption.
Study limitations
We have previously demonstrated that the applied instrumentation permits a reliable assessment of ITBV, PBV and Vdcirc [5], meaning that the applied technique can be regarded sufficiently sensitive to detect small changes in the measured blood volumes. Yet, it is important to stress that Vdcirc systematically underestimates total blood volume by about 40% [29] mainly due to incomplete mixing of the dye in more slowly perfused tissues. However, changes in Vdcirc can still be regarded reliable [5, 20]. Also, previous investigations either focused solely at pressure-effects, or were performed on perfused isolated organs [9, 10, 12] and did not allow assessing the effects of selective, solitary alterations in PVT on an intact circulation which we considered indispensable since it is the systemic circulation that determines VR. This implies that the complex interplay between cardiac preload and afterload can only be reliably assessed in an intact circulation.
In contrast, the use of a fixed FiO2 (about 0.10) and standardized PGI2 doses may be considered a limitation of the study, as the hemodynamic effects may be ‘dose’-dependent. It may be argued that during more severe hypoxia and/or higher dosages of PGI2, compensatory physiologic mechanisms might fail and would reveal a more pronounced hemodynamic perturbance, although the current observations do not demonstrate such a trend.
Since the spleen and hepatosplanchnic circulation function as a blood reservoir in dogs in case of sympathetic activation [30], blood volumes may be altered by hypoxia and aerosolized PGI2 inhalation. However, given that Vdcirc remained unaltered during the study, this effect—if there were any—is most likely negligible.
Our experiments took place while dogs were breathing spontaneously and observations can therefore not be extrapolated directly to mechanically ventilated subjects. Mechanical ventilation is associated with a shift of blood from the intra- to the extrathoracic compartment [31], which would intrinsically influence the results. To account for the effects of endotracheal intubation, a control group served for evaluation of time-related and spontaneous variations in pulmonary and systemic variables. In this control group, the dogs breathed air with and without an endotracheal tube, and no hemodynamic differences were observed (data not shown).
Finally, in the primary outcome variables of this study—i.e., the data on measured blood volumes—no statistically significant differences were found between the various interventions. It should be stressed that a relatively limited number of subjects was investigated, in which a limited number of interventions was performed. Even though the observed effect sizes between interventions appear to be very small, it may be true that a larger sample size would have resulted in (subtle) statistically significant differences between the various interventions.
Conclusions
In this experimental study in spontaneously breathing dogs, hypoxia increased pulmonary vascular tone substantially. Cardiac output was maintained, owing to an increase in heart rate to compensate a decrease in stroke volume. Central blood volumes and their distribution with circulating blood volume, and the pressure for venous return remained unaltered.
PGI2 decreased pulmonary vascular tone substantially: CO, central blood volumes and their relation to circulating blood volume, as well as pressure for venous return remained unaltered.
These cumulative observations suggest that cardiac preload was preserved despite substantial alterations in pulmonary vascular tone, i.e., right ventricular afterload.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- CO:
-
Cardiac output
- CVP:
-
Central venous pressure
- Eh:
-
Heart efficiency
- FiO2 :
-
Fraction of inspired oxygen (O2)
- HPV:
-
Hypoxic pulmonary vasoconstriction
- ITBV:
-
Intrathoracic blood volume
- LV:
-
Left ventricle
- MAP:
-
Mean arterial pressure
- PaO2 :
-
Arterial partial oxygen pressure
- PAP:
-
Pulmonary artery pressure
- PBV:
-
Pulmonary blood volume
- PCWP:
-
Pulmonary capillary wedge pressure
- PGI2 :
-
Aerosolized prostacyclin
- Pmsa:
-
Analogue of mean systemic filling pressure
- Pmsf:
-
Mean systemic filling pressure
- Pvr:
-
Pressure for venous return
- PVT:
-
Pulmonary vascular tone
- RAP:
-
Right atrial pressure
- RV:
-
Right ventricle
- RVR:
-
Resistance to venous return
- SaO2 :
-
Arterial oxygen saturation
- Vdcirc :
-
Circulating blood volume
- VR:
-
Venous return
References
Guyton AC (1968) Regulation of cardiac output. Anesthesiology 29:314–326
Magder S (2012) Bench-to-bedside review: an approach to hemodynamic monitoring—guyton at the bedside. Crit Care 16:236
Funk DJ, Jacobsohn E, Kumar A (2013) The role of venous return in critical illness and shock-part I: physiology. Crit Care Med 41:255–262
Maas JJ, Pinsky MR, de Wilde RB, de Jonge E, Jansen JR (2013) Cardiac output response to norepinephrine in postoperative cardiac surgery patients: interpretation with venous return and cardiac function curves. Crit Care Med 41:143–150
Vos JJ, Scheeren TW, Loer SA, Hoeft A, Wietasch JK (2016) Do intravascular hypo- and hypervolaemia result in changes in central blood volumes? Br J Anaesth 116:46–53
Lammers S, Scott D, Hunter K, Tan W, Shandas R, Stenmark KR (2012) Mechanics and function of the pulmonary vasculature: implications for pulmonary vascular disease and right ventricular function. Compr Physiol 2:295–319
Lumb AB, Slinger P (2015) Hypoxic pulmonary vasoconstriction: physiology and anesthetic implications. Anesthesiology 122:932–946
Mitchell JA, Ali F, Bailey L, Moreno L, Harrington LS (2008) Role of nitric oxide and prostacyclin as vasoactive hormones released by the endothelium. Exp Physiol 93:141–147
Ding ZP, Scheeren TW, Arndt JO (1999) Effects of pulmonary blood volume on vascular reactivity in the lung. Intensive Care Med 25:1413–1420
Nelin LD, Moshin J, Thomas CJ, Sasidharan P, Dawson CA (1994) The effect of inhaled nitric oxide on the pulmonary circulation of the neonatal pig. Pediatr Res 35:20–24
Dawson CA, Forrester TE, Hamilton LH (1975) Effects of hypoxia and histamine infusion on lung blood volume. J Appl Physiol 38:811–816
Welling KL, Sander M, Ravn JB, Larsen B, Abildgaard U, Amtorp O (1997) Effect of alveolar hypoxia on segmental pulmonary vascular resistance and lung fluid balance in dogs. Acta Physiol Scand 161:177–186
Isbister JP (1997) Physiology and pathophysiology of blood volume regulation. Transfus Sci 18:409–423
Vos JJ, Kalmar AF, Hendriks HG, Bakker J, Scheeren TW (2018) The effect of fluid resuscitation on the effective circulating volume in patients undergoing liver surgery: a post-hoc analysis of a randomized controlled trial. J Clin Monit Comput 32:73–80
Parkin G, Wright C, Bellomo R, Boyce N (1994) Use of a mean systemic filling pressure analogue during the closed-loop control of fluid replacement in continuous hemodiafiltration. J Crit Care 9:124–133
Gupta K, Sondergaard S, Parkin G, Leaning M, Aneman A (2015) Applying mean systemic filling pressure to assess the response to fluid boluses in cardiac post-surgical patients. Intensive Care Med 41:265–272
Vos JJ, Kalmar AF, Scheeren TW (2020) Bedside assessment and clinical utility of mean systemic filling pressure in acute care. J Emerg Crit Care Med 8:4–25
du Sert PN, Ahluwalia A, Alam S, Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U, Emerson M, Garner P, Holgate ST, Howells DW, Hurst V, Karp NA, Lazic SE, Lidster K, MacCallum CJ, Macleod M, Pearl EJ, Petersen OH, Rawle F, Reynolds P, Rooney K, Sena ES, Silberberg SD, Steckler T, Würbel H (2020) Reporting animal research: explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol 18:e3000411
Picker O, Schindler A, Scheeren TW (2000) Accuracy and reproducibility of long-term implanted transit-time ultrasound flow probes in dogs. Intensive Care Med 26:601–607
Hoeft A, Schorn B, Weyland A, Scholz M, Buhre W, Stepanek E, Allen SJ, Sonntag H (1994) Bedside assessment of intravascular volume status in patients undergoing coronary bypass surgery. Anesthesiology 81:76–86
Maas JJ, Pinsky MR, Geerts BF, de Wilde RB, Jansen JR (2012) Estimation of mean systemic filling pressure in postoperative cardiac surgery patients with three methods. Intensive Care Med 38:1452–1460
Lee JM, Ogundele O, Pike F, Pinsky MR (2013) Effect of acute endotoxemia on analog estimates of mean systemic pressure. J Crit Care 28:880.e9,880.15
Colebourn C, Newton J. Anonymous Acute and critical care echocardiography. 2017; 236–245.
Kahler RL, Goldblatt A, Braunwald E (1962) The effects of acute hypoxia on the systemic venous and arterial systems and on myocardial contractile force. J Clin Invest 41:1553–1563
Brower RG, Gottlieb J, Wise RA, Permutt S, Sylvester JT (1985) Locus of hypoxic vasoconstriction in isolated ferret lungs. J Appl Physiol 1987(63):58–65
Ivy DD (2010) Prostacyclin in the intensive care setting. Pediatr Crit Care Med 11:S41–S45
Scheeren T, Radermacher P (1997) Prostacyclin (PGI2): new aspects of an old substance in the treatment of critically ill patients. Intensive Care Med 23:146–158
Chaignon M, Tannieres-Ruffie ML, Lucsko M, Guedon J (1982) Acute hemodynamic effects of prostacyclin infusion in severe or malignant hypertension. Eur Heart J 3(Suppl C):39–43
Picker O, Wietasch G, Scheeren TW, Arndt JO (2001) Determination of total blood volume by indicator dilution: a comparison of mean transit time and mass conservation principle. Intensive Care Med 27:767–774
Carneiro JJ, Donald DE (1977) Blood reservoir function of dog spleen, liver, and intestine. Am J Physiol 232:H67-72
von Spiegel T, Giannaris S, Schorn B, Scholz M, Wietasch GJ, Hoeft A (2002) Effects of induction of anaesthesia with sufentanil and positive-pressure ventilation on the intra- to extrathoracic volume distribution. Eur J Anaesthesiol 19:428–435
Acknowledgements
The authors wish to thank Paul Soultis for his help in data analysis. Andreas Hoeft passed away, after approving the final version of this manuscript. This is one of his last works.
Funding
This work was supported by departmental and institutional funding only.
Author information
Authors and Affiliations
Contributions
JJV: data analysis and interpretation, manuscript writing. JKGW: study design, data analysis, collection and interpretation, manuscript writing. AH: data collection and interpretation, manuscript writing. TWLS: study design, data collection and interpretation, manuscript writing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Yes, by the district governmental animal investigation committee (North-Rhine Westphalia in Dusseldorf, Germany; registered as Ref. 23.05-2303-84/96.
Consent for publication
Not applicable.
Competing interests
JJV and JKGW report no competing interests. TWLS received research grants and honoraria from Edwards Lifesciences (Irvine, CA, USA) and Masimo Inc. (Irvine, CA, USA) for consulting and lecturing and from Pulsion Medical Systems SE (Feldkirchen, Germany) for lecturing. TWLS is editor-in-chief of the Journal of Clinical Monitoring and Computing.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vos, J.J., Wietasch, J.K.G., Hoeft, A. et al. Do alterations in pulmonary vascular tone result in changes in central blood volumes? An experimental study. ICMx 9, 59 (2021). https://doi.org/10.1186/s40635-021-00421-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40635-021-00421-8