
Abstract
As one of the world’s earliest pottery origins in Eurasia, the role of pottery during Neolithic period in South China remains unclear. Here, we present a new understanding on the utilization pattern of early pottery from the Neolithic sites of Dayan, Dingsishan and Zengpiyan in Guangxi province, China. Compound-specific stable isotope analysis of pottery and bone lipids show that a persistent use of terrestrial C3 and C4 resources throughout the pottery utilization history highlight the culinary practice and terrestrial exploitation during Neolithic period. Contrary to the traditional view that the primary use of these pottery was closely linked with aquatic resource processing, no aquatic biomarkers were found in the potsherds, yet these compounds were detected in the cooking experiments in this study, indicating that aquatic resources in these sites were probably processed or consumed in other ways. Moreover, a remarkable diachronic change from extensive use of C3 and C4 resources to the intensive use of certain kinds of C3 resources in different Neolithic stages, further indicating that an ecological tradition rooted in the subtropical and tropical environments as well as the Neolithic social evolution exerted great influence upon the initial production and the subsequent utilization of early pottery in prehistoric South China.
Similar content being viewed by others

Introduction
The phenomenon of pottery emergence towards the end of the Late Pleistocene made East Eurasia the recognized area of pottery origin. A series of early pottery vessel traditions that appeared simultaneously in South China, the Japanese Islands and the Russian Far East between approximately 20000–10000 BP [1,2,3,4], launched the ongoing discussion on the pottery use and the trajectories of prehistoric social evolution associated with them [5, 6]. In contrast to the traditional view that pottery use was linked with agricultural communities [7,8,9], new advances in archaeological research have been successful in linking pottery use with the adaptation to changing ecological and social structures even before farming within Eurasia [10, 11] and demonstrating that the use of early pottery and resource processing had a strong association with the newly established ecological conditions in the Japanese archipelago and the Russian Far East during the transition to Holocene [12, 13].
As one of the oldest regions of pottery origin in Eurasia, the invention and utilization of pottery in South China have always attracted much attention [1, 5, 10, 11]. The earliest pottery appeared in several sites in South China at the end of the terminal Pleistocene, such as pottery containers excavated from Yuchanyan cave in Hunan province, Xianrendong cave in Jiangxi province and Dayan and Zengpiyan caves in Guangxi province [1, 4, 14], and gradually adopted in the subsequent millennia with further social evolution. It has been suggested that the initial function of these early pottery had more connection with resource processing, particularly for aquatic resources [14,15,16,17]. Significantly, the most prominent feature concerning the pottery-bearing sites across the area is the apparent increase in the intensity of shellfish, which was not seen in quantities before the early Neolithic periods [18, 19]. Accordingly, extensive aquatic resource exploitation during the local Neolithic has been demonstrated through stable carbon and nitrogen isotope analysis of human and animal bone collagen [20, 21].
However, recent molecular analysis of lipid residues in early pottery from Neolithic shell mounds in South China did not detect reliable evidence of aquatic biomarkers, such as isoprenoid fatty acids (e.g., 4,8,12-TMTD, phytanic and pristanic acids) and ⍵-(o-alkylphenyl) alkanoic acids (APAAs), and their presence is indicative of cooking aquatic foods [22]. Conversely, a series of terrestrial biomarkers, such as phytosterols, terpenes, alkanes and animal cholesterols, predominate in these early potsherds [23, 24], suggesting that the vessels were primarily used for processing terrestrial plants and animals. Indeed, it has long been argued that the use of early pottery vessels was affected by the ecological and social traditions across different regions [5, 10, 25, 26]. This is particularly true in South China, where abundant subtropical and tropical resources remain relatively stable through millions of years [27]. Similarly, the archaeological records of lithic assemblages and quantities of the terrestrial remains co-existed with the pottery sherds in the study sites may suggest a deep relationship between pottery use and the adaption to the ecological and social traditions in South China [5, 10]. Although archaeological evidence indicates that resource processing was a crucial factor for pottery use in prehistoric South China, detailed pottery residue analysis remains limited.
This study aims to investigate the function of early pottery and diachronic change of resource utilization patterns throughout the Neolithic times in South China by analyzing lipid residues in pottery and contemporaneous animal bones from the Neolithic sites (ca. 12,000–6000 cal BP) of Dayan, Dingsishan and Zengpiyan in Guangxi province, China. While it has long been assumed that these early pottery vessels were closely linked with the processing of aquatic resources, little evidence of aquatic exploitation was found in previous studies [23, 24]. Cooking experiments were also conducted in this study to determine the formation of the aquatic biomarkers through different cooking conditions and further improve our understanding of the characteristic of aquatic lipids in pottery vessels after cooking.
Principles of pottery residue analysis
Organic residues in archaeology include DNA, proteins, lipids and other biomolecular from archaeological deposits. Of which, lipids are defined by their small atomic mass units (> 200 but < 1000 amu) and hydrophobicity, this property forms the basis for their preservation and extraction from archaeological materials [28]. Biomarkers, as “chemical fingerprints” of lipids, are the compounds that originated from specific biological and biochemical components of natural products [29]. These compounds are usually preserved in archaeological pottery, via surface adsorption or trapping within molecular-sized pores of the inside wall. Based on the concept of archaeological biomarkers, lipid residues extracted from pottery could provide molecular evidence for the foodstuffs (such as aquatic resources, terrestrial animals, edible plants), and various products (such as resins, waxes, tars), which were likely to have been exploited in the past [29, 30]. On the other hand, plants have distinct δ13C values depending on their photosynthetic pathways, such as C3, C4 or CAM plants, the δ13C values of animal and plant tissues generally reflect the isotopic level of local environments [31]. Moreover, the δ13C values of food and fluid ingested by animals also have a strong impact on the isotopic compositions of the tissues they synthesize [32]. Thus, the application of compound-specific stable isotope analysis of the fatty acids that composed of lipid could contribute to further distinguishing the animal and plant type, such as ruminants, non-ruminants and dairy fats [30]. To date, the use of organic residue analysis has shown great potential in revealing the diversified resource utilization in ancient pottery among different regions, such as the earliest use of aquatic resources in the Japanese archipelago and North Africa [33, 34]; terrestrial plant processing in China, Southeast Asia and North America [35,36,37,38]; and animal domestication and beeswax utilization in Europe [39,40,41].
Archaeological background
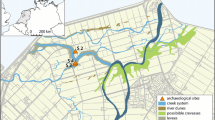
The archaeological sites of Dayan, Zengpiyan and Dingsishan are located in Guangxi province of China with Dayan and Zengpiyan sites lying in the karst plain along the Li Jiang River in Northern Guangxi, while the Dingsishan site is situated on the banks of Yong Jiang River in Southern Guangxi (Fig. 1). These sites were systematically excavated during 1997 and 2001 by the Institute of Archaeology, Chinese Academy of Social Sciences and Archaeological Team of Guangxi Zhuang Autonomous Region [42,43,44], and are representative of the Neolithic material culture of South China.

Map showing the location of the study sites within South China
Archaeological deposits in these sites can be divided into four major phases: terminal Paleolithic (15000–12000 BP), Early Neolithic (12000–9000 BP), Middle Neolithic (9000–7000 BP) and Late Neolithic (7000–6000 BP), through radiocarbon dating and material cultural associations [42, 43]. The chronology and cultural frameworks based on archaeological assemblages in these sites suggested a typical local culture techno-complex characterized by flexed burials, flake lithic assemblages and other stone, bone and shell tools, as well as an abundance of aquatic and terrestrial animal and plant remains [14, 19, 45]. Notably, early pottery vessels (Fig. 2) were found in these sites, with the earliest pottery container (mainly round-bottomed ‘fu’) dated to approximately 12000 BP at Dayan and 10000 BP at Dingsishan, and are comparable to contemporary pottery from the sites of Zengpiyan cave in Guangxi province and Yuchanyan cave in Hunan province, China [42].

Early pottery samples from Dayan and Dingsishan sites: a–d the earliest pottery sherd from Dayan site (a pottery sample in this study, b, c general view of the earliest pottery sherd, d restoration image of the earliest pottery with the sample position, pictured from Xinrong Jiang), e–l pottery sherds from Dingsishan site (e, f early Neolithic, g–j middle Neolithic, k, l late Neolithic)
There are several shared features between the early pottery (dated to approximately 12000–6000 BP) in these sites, such as small quantity but large size, simple cord-marked and thick walls (about 2.9 cm thick for earliest potsherd from Dayan cave, Fig. 2c, d). These pottery vessels are tempered with large particles of calcite or quartzite, indicating low firing temperature and pristine technology in Early Neolithic [43]. From the middle Neolithic, an increase in the quantity, quality and pottery types, particularly, a significant shift towards smaller pottery sizes and thin walls, suggests subsequent development in pottery technology [42, 43]. Besides the manufacturing technology, the functional need for resource preparation is also determined by the reconstruction of life histories of pot vessels through absorbed lipids [23, 24], but the role of these early pottery within a changing social context in prehistoric South China remains unclear.
Materials and methods
Materials
A total of 62 archaeological samples, including 52 potsherds (examples of the potsherds are represented in Fig. 2) and 10 animal bones which include Water buffalo (n = 3), Deer (n = 2), Small carnivore (n = 1), Wild boar (n = 1) and Freshwater fish (n = 3), from Neolithic sites of Dayan and Dingsishan and Zengpiyan in Guangxi, South China; combined with 36 modern replica potsherds and foodstuffs from cooking experiments (Additional file 1: Figure S1) are selected for lipid residue analysis through Gas Chromatography-Mass Spectrometry (GC–MS) and Gas Chromatography-Combustion-Isotope Ratio Mass Spectrometry (GC-C-IRMS).
Cooking experiments
Modern replica potsherds (n = 30) and food products (n = 6) in cooking experiments were used to generate comparative data for further understand the characteristics of aquatic lipids in pottery through different time and temperature.
For the cooking experiments, modern replica pottery vessels used in this study were wheel-thrown and manufactured in Jingdezhen, Jiangxi Province, China. The mineral composition of these pots mainly includes local Pot Clay and sand, and the firing temperatures were relatively high (around 1000 ℃) to avoid fire-crack. The food products (Additional file 1: Table S1), including shellfish (Viviparus), freshwater fish (Cyprinus carpio), meat (lamb), taro, Chinese yam, and leafy vegetables, were placed into these replica pots over the charcoal fire stoves.
To determine the temperature and time required for the formation of aquatic lipids in pottery vessels, particularly for aquatic biomarkers (such as, ω-(o-alkylphenyl)-alkanoic acids (APAAs), phytanic, pristanic acids and 4,8,12-TMTD), series of temperatures on 230 °C, 250 °C and 280 °C adjustment from previous studies [46, 47], were set as 5 h and 10 h (1 × 10 times) for three groups respectively, each group used the same portions of food ingredients (Additional file 1: Table S1). In addition, a small amount of roasting and raw meat from shellfish were also prepared for comparison (Additional file 1: Table S1). The experiments were conducted at the field workstation of the Institute of Archaeology, Chinese Academy of Social Science. The food products used in this study were bought from supermarket in Beijing and Guangxi province, China.
Lipid extraction
1–2 mm of the inside surface of the potsherd were first mechanically removed using a hand-drill in order to eliminate exogenous residue and possible contamination. About 1 g of potsherd was then ground to powder and placed into a glass tube. 4 mL methanol was added and ultrasonicated for 15 min. 800 μL of concentrated sulfuric acid was added and heated for 4 h at 70 ℃, after centrifuged for 5 min, the supernatant was further extracted with hexane (4 × 2 × 2 mL) and collected via a prepared Pasteur pipette three times. Finally, the extracts were dried under a gentle stream of nitrogen. An internal standard of 10 μL n-tetratriacontane (C34:0) or n-hexatriacontane (C36:0) was added to each sample before analysis by GC–MS.
The archaeological animal bones were used as isotopic baseline for pottery lipids in this study. These bone samples were taken from the same one where bone collagen and hydroxyapatite were well preserved [21, 48]. While given the preservation of the bone lipids, a two-step extraction [49] was conducted to obtain the endogenous lipid in bones. About 1 g of bone fragments (limb) were mechanically cleaned and ground to powder using a pestle and mortar. The bone powder was first washed with solvent extracted with dichloromethane/methanol (2:1 vol/vol, 4 × 2 × 2 mL); after being ultrasonicated for 15 min and centrifuged for 5 min, the supernatant was removed and repeated three times and then dried completely under a gentle stream of nitrogen. The second step was using acidified methanol procedure to the remaining bone powder exactly as above.
Analytical methods
All the lipid extracts were then analysed by GC–MS and GC-C-IRMS. Since the lipid composition of the archaeological potsherds (n = 52) has already been investigated in our previous work [23], GC–MS analysis were conducted on the archaeological bones (n = 10) and modern materials (n = 36) from experimental cooking.
GC–MS analysis of the methylated extracts was carried out using an Agilent 7890A gas chromatograph attached to a 5975C mass detector (Agilent Technologies) in electron ionization mode (70 eV) at the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences. A capillary column DB-5HT (15 m × 0.32 mm × 0.1 μm) was used to separate samples. 1 μL of the sample was injected in a split-less mode. The temperature program was as follows: initial temperature 50 °C for 1 min, increased to 100 °C at 15 °C/min, then up to 375 °C at 10 °C/min, with 10 min isothermal. Helium was used as the carrier gas in a constant flow rate of 2.0 mL/min. The MS ion source temperature was 230 °C; the interface temperature was 300 °C; the MS quadrupole temperature was 150 °C. The spectrometer is run in full-scan mode and in selected ion monitoring (SIM) mode in order to monitor aquatic biomarkers. The group of ions at m/z 262, 290, 318, 346 were selected to assess APAAs of carbon chain lengths C16–C22, and the fragment ion of the base peak m/z 105; ions of m/z 74, 87, 213, 270 corresponding 4,8,12-trimethyltridecanoic acid (TMTD) fragmentation; m/z 74, 88, 101, 312 corresponding to pristanic acid; m/z 74, 101, 171, 326 corresponding to phytanic acid, respectively [22, 50]. Identification of common lipids was performed by comparison with the NIST mass spectral database or mass spectra from the published literature.
Compound-specific carbon stable isotope analysis of archaeological samples was carried out on a Thermo Trace Gas Chromatography Ultra coupled with an Isotope Ratio Mass Spectrometer (Thermo Fisher Scientific MAT253) via a combustion reactor held at 1000 °C, at the Institute of Atmospheric Physics, Chinese Academy of Sciences. All samples were diluted with hexane and 1 μL of each sample was injected into a DB-5 MS fused silica capillary column (30 m × 0.25 mm, 0.25 μm film thickness). The GC oven temperature was set for 50 °C (2 min) to 120 °C at 15 °C/min, then from 120 to 300 °C at 5 °C/min, with a final isotherm at 300 °C for 16 min. High-purity grade helium was used as a carrier gas maintained at a constant flow rate of 2.0 mL/min. Compounds were combusted to CO2 and ionised in a mass spectrometer. The ion intensities of m/z 44, 45 and 46 were monitored so as to automatically compute the 13C/12C ratio for each sample peak by comparison with a calibrated CO2 reference gas of known isotopic composition through Isodat 3.0 Gas Isotope Ratio MS Software (version 3.0; Thermo Fisher). The analysis results are reported as per mil (‰) relative to internationally defined standards for carbon (Vienna Pee Dee Belemnite, VPDB). Isotopic values of each sample were monitored by use of an International standard mixture of fatty acid methyl esters with known δ13C values (Indiana F8-3 standard; obtained from Biogeochemical Laboratories, Indiana University), the measurement error of δ13C values of each compound based on duplicate analysis was less than ± 0.4‰. Moreover, the δ13C values were corrected for the potential carbon atoms that occurred during methylation using the measured carbon isotopic value of the methanol used in pretreatments.
Results
Aquatic biomarkers from cooking experiments
The results of the cooking experiments are listed in Additional file 1: Table S1. In total, the lipid extracts from 36 cooking materials, including replica potsherds and foodstuffs resulted in a considerable abundance of endogenous saturated and unsaturated fatty acids (C11:0–C25:0, C16:3–C24:2), long-chain hydroxy fatty acids, DPA (7,10,13,16,19-docosapentaenoic, C22:5), DHA (4,7,10,13,16,19-docosahexaenoic acid, C22:6), EPA (5,8,11,14,17-eicosapentaenoic acid, C20:5), plant sterols and animal cholesterols. The aquatic biomarkers of ω-(o-alkylphenyl) alkanoic acids (APAAs-C16–C22), and a trilogy of isoprenoid fatty acids, including 3,7,11,15-tetramethylhexadecanoic acid (phytanic acid), 2,6,10,14-tetramethylpentadecanoic acid (pristanic acid) and 4,8,12-trimethyltridecanoic acid (4,8,12-TMTD), were also detected in both potsherds and foodstuffs (Additional file 1: Table S1).
Notably, our results show that while the heating conditions at 250 ℃ for 5 to 10 h can produce partial APAAs with carbon length C18 and C20, abundant long-chain APAAs (carbon length of C16–C22) are generated at a higher temperature of 280℃ for 5 h (Additional file 1: Table S1). Moreover, contrary to a previous study [47], APAAs were not detected at a lower temperature of 230 ℃ for either 5 or 10 h (Fig. 3 and Additional file 1: Table S1), suggesting an increased temperature at ≥ 250 ℃ should be the prerequisite for the formation of APAAs. In addition, the sealed condition is seemingly not necessary for the production of APAAs, as abundant APAAs were detected in fired meat of shellfish under direct open fire at 280 ℃ for only 30 min, which further suggests that high temperature is needed to form these compounds (Additional file 1: Table S1).

Total ion current (TIC) and m/z 105 of the cooking shellfish. Bottom: fatty acids distributions without APAAs heated at 230 ℃ for 5 h. Top: Mass chromatograms of base peak (m/z 105) with molecular ions of C18–C22 APAAs heated at 280 ℃ for 5 h
APAAs with different carbon lengths and low concentrations of phytanic acid were detected either in the separate vessels containing aquatic and plant foods or in a vessel containing both of them (Additional file 1: Table S1). While 4,8,12-TMTD were only detected in the vessels containing aquatic foods (Fig. 4 and Additional file 1: Table S1), which aligns with previous studies that such a compound occur stably in aquatic environments owing to the fat catabolism of aquatic organisms [22, 51]. Moreover, as 4,8,12-TMTD was also detected in the raw shellfish as well as vessels containing aquatic foods cooked at a lower temperature of 230 ℃ for 5 h (Additional file 1: Table S1) in this study, it seems to suggest that the presence of this compound may not require high temperature or protracted heating, but rather reflects a typical characteristic of aquatic lipids.

TIC chromatograms from cooking shellfish (top) and plants (bottom). Top: APAAs based on ions m/z 105, 262, 290, 318 and 346 with carbon length C16–C22. 4,8,12-TMTD based on ions m/z 87 and 270. Phytanic acid was monitored with ions based on m/z 101. Bottom: APAAs with carbon length C18 and phytanic acid were also detected after cooking plants
Thus, based on the cooking experiments in this study, we suggest that despite the formation of APAAs in pottery needs high heating conditions, 4,8,12-TMTD may not. Instead, it suggests that if the aquatic foods were ever processed in pottery even at a lower heating condition, certain amount of 4,8,12-TMTD could be detected. However, the post-depositional survival of this compound under different environmental conditions needs further investigation.
Lipid molecular signature in archaeological bones
The preservation of bone lipids is an important prerequisite for lipid residue analysis. For the archaeological bones in this study, the total lipid concentrations decreased from 1128.8 to 750.5 μg g−1 (Additional file 1: Table S2) by using the modified procedure of two-step extraction [49], probably due to the loss of both endogenous lipids and exogenous contaminants, such as phthalates from plasticizers, during the repeat of solvent washes in the first step. An obvious removal of several phthalates in deer bone confirmed the efficiency of this method (Additional file 1: Figure S2). Furthermore, the lipid distributions in animal bones after the solvent washed were similar between species, mainly dominated by saturated fatty acids of C12:0–C30:0, branched fatty acids of C14:0 and C16:0 and monounsaturated fatty acids of C18:1 and C22:1. Cholesterol derivatives were also present in trace amounts (Additional file 1: Figure S2). These characteristics indicate the endogenous lipids had been preserved in bone fats and provide an essential assurance for the compound-specific stable isotope analysis.
Stable isotope composition of archaeological bones and pottery lipids
Ten archaeological animal bones, including Water buffalo (n = 3), Deer (n = 2), Small carnivore (n = 1), Wild boar (n = 1) and Freshwater fish (n = 3), from Neolithic sites of Zengpiyan and Dayan were selected as the isotopic baseline for compound-specific δ13C values of pottery residues. The δ13C values of palmitic (C16:0) and stearic acids (C18:0) extracted from bone lipids (presented in Additional file 1: Table S2) ranged from −30.0 to −26.6‰ for C16:0 and −30.0 to −28.3‰ for C18:0 (n = 10), indicating a widely C3 based lipid origins. However, a more complicated isotopic fractionation related to lipid synthesis and different dietary metabolism among animal species should be considered when compared to isotopic data of lipid, collagen and apatite from the same bone [21, 48].
The δ13C values of bone components from different species are shown in Fig. 5. The general depletion patterns of 13C in the C16:0 and C18:0 fatty acids relative to collagen and apatite in terrestrial mammals are consistent with previous studies due to discrimination against 13C during the metabolic pathway of lipid synthesis which resulted in more depleted δ13C values than other tissues, e.g., protein, carbohydrate and cholesterol [52, 53]. For most of the terrestrial mammalian bones in Fig. 5, including deer, wild boar and small carnivores, the δ13C values of fatty acid, collagen and apatite indicate a C3 lipids origin with dietary digestion and biosynthetic process. Notably, subtle variation of δ13C in apatite (−11.3‰) relative to fatty acids (−29.8‰ for C16:0 and −28.3‰ for C18:0) and collagen (−21.9 ± 0.1‰) from wild boar may relate to a small intake of C4 foods. This difference is particularly clear in the bone composition of water buffalo (Fig. 5 and Additional file 1: Table S2), in which collagen (−12.8 ± 3.7‰) and apatite (−8.3‰) indicated a more C4 diet (enriched in 13C), while the fatty acids (−27.0 ± 0.5‰ for C16:0 and −28.9 ± 0.3‰ for C18:0) show a depleted in 13C by 14.2‰ (Δδ13C = C16:0FA-COLL) and 18.7‰ (Δδ13C = C16:0FA-APAT). This may be mainly reflected by variations of isotopic fractionation during carbon metabolic and biosynthesis in the organism [54, 55].

The offsets of δ13C values between bone components of different animals
Controlled animal feeding experiments have found significant differences (−3.3‰ to + 12.2‰) in δ13C values of tissues from animals fed on a diet of constant isotopic composition [53, 56] and pointed out that the fractionation of 13C from diet to tissue is not identical on different diets, such as the proportion of C3 vs C4, or aquatic vs terrestrial in diet. Considering the habitat environments and foraging behavior of water buffalo, its food resource may contain terrestrial and aquatic C3 and C4 plants. This mixed diet could lead to a larger isotopic difference in these bone components, although sex, age and physical condition may also have secondary effects [53].
Carbon sources are complex in the freshwater ecosystem and can give rise to a wide range of isotopic ratios in freshwater environments. The δ13C values of bone components plotted in Fig. 5 shows a pattern that fatty acids are more depleted than their corresponding collagen and apatite values for freshwater fish, suggesting a typical lipid origin similar to the terrestrial C3 pathway. As observed for most of the terrestrial mammals in Fig. 5, their δ13C values provide a terrestrial isotopic baseline of C3 dominates.
Stable isotopic data of 52 pottery lipids, which included 11 early Neolithic, 20 middle Neolithic and 21 late Neolithic samples, from Dayan and Dingsishan sites with the reference isotopic baseline of animal bone fatty acids (n = 10) from Zengpiyan and Dayan sites are plotted in Fig. 6a. In general, the overall isotope results of pottery lipids are quite variable with about 70% (37 of 52) of the samples yielding plant biomarkers [23]. These data tended to have a broader range of δ13C values from −33.4 to −17.5‰ for C16:0 and −35.5 to −22.9‰ for C18:0 (n = 37). Few samples show an increase of C4 signals relative to the reference animals, indicating lipids ranging from predominantly C3 resources with some C4 resources.

Plot of δ13C values (a) and the offset values (Δ13C, b) of C16:0 and C18:0 in archaeological bone and pottery lipids from Zengpiyan, Dayan and Dingsishan sites
Samples without plant biomarkers have the molecular composition characterized by higher abundance of C18:0 and lower abundance of C16:0 [23]. These samples showing a relatively narrow distribution with δ13C values ranging from −31.6 to −26.4‰ for C16:0 and −31.9 to −28.0‰ for C18:0 (n = 15), and therefore reflect that lipids were derived entirely from C3 animals. Except for a few extreme data pointing to an increased C4 resource, the δ13C values of most samples fall into the isotopic ranges between terrestrial C3 and aquatic resources in Fig. 6a. However, aquatic lipid origin cannot be verified solely from isotopic data, due to some overlapping in the ẟ13C values of aquatic lipids and terrestrial C3 lipids, especially aquatic biomarkers were not detected in these pottery samples [23]. Based on the results of cooking experiments shown in Fig. 4 and Additional file 1: Table S1, potsherds that have been processed for aquatic stuffs under high heating conditions yielded aquatic biomarkers including isomers of APAAs (C16–C22) and at least one isoprenoid fatty acids. Other potsherds that underwent low temperature conditions as well as raw aquatic materials had partial aquatic biomarkers, especially 4,8,12-TMTD. Thus, we suggest that if pottery had been used to process aquatic resources, partial aquatic biomarkers at least should be detected. Therefore, a predominantly terrestrial resource origin is convincing based on the current results.
This result is further supported in Fig. 6b by plotting Δ13C (δ13C18:0–δ13C16:0) values against δ13C16:0 values. The offset (Δ13C) reflected the different routing of C18:0 and C16:0 during lipid synthesis in ruminant dairy fats and adipose fats, emphasizing the difference in biosynthetic process between non-ruminant and ruminant fat types [57]. From the data shown in Fig. 6b, the classification of ruminant dairy fats from adipose fats and non-ruminant adipose fats can be separated through Δ13C values of animal bone fatty acids by displaying the offsets lower than 0.0‰ and −3.3‰ [58], and provide the reference proportion that approximately 40% (22/52) of the ruminant resources and 60% (30/52) of non-ruminant resources, including terrestrial plants, were being processed in these early pots, which is consistent with archaeobotanical and zooarchaeological studies in the site area [19, 44, 59].
One sample fell within the reference range of ruminant dairy fats with a clear C4 signal which was beyond our expectation. However, no clear lipid components of dairy fats, such as triacylglycerols, were identified. Low abundance of short-chain (C8:0, C12:0–C14:0) fatty acids detected in this sample [23], were probably from the degradation of free fatty acids through thermal cracking, or by bacterial action [60]. Given that only one sample falls into this range and no archaeological evidence related to dairy products or activities have been found during the Neolithic times in the site area, we suggest that dairying was unlikely to take place during the early stage of the Neolithic in this region, although occasional utilization cannot be ruled out.
Overall, isotope results combined with molecular information demonstrate that a mixture of terrestrial C3 and C4 resources, including plants, ruminants and non-ruminants, dominate the utilization history of the early pottery vessels in both sites. However, the evidence of frequent processing of aquatic products within these pots remain less clear. The findings suggest that despite the abundant edible aquatic resources in the study area, the Neolithic people had a clear preference for processing terrestrial resources via pottery vessels, and indicated that the culinary practice was homogeneous in this region and was persistent throughout Neolithic times.
Discussion
A new understanding on the resource utilization pattern through pottery during Neolithic times in South China
The emergence and utilization of pottery in South China have been widely debated, mainly focusing on the function behind its invention [5, 10, 61]. A persistent use of terrestrial C3 and C4 resources through our systematically stable isotope analysis of pottery lipids together with plenty of terrestrial biomarkers (such as a series of diterpenes, triterpenes, sterols, cholesterols and long-chain ketones) in previous studies, suggests these Neolithic vessels have mainly been used to process terrestrial plants and animals, rather than aquatic resource, which is in contrast with the traditional view. Moreover, our new results also different from that in Japan and the Eastern Russia, where most of the early pots yielded aquatic signals, suggesting the extensive exploitation of aquatic resources [12, 13]. The pottery use pattern in this study represents a unique phenomenon among prehistoric shell mounds, providing new insight into the local traits of ecological tradition and social evolution during Neolithic times in South China.
In the ecological context of South China, the subtropical and tropical environments, compared to other regions in East Eurasia, promote the rich and unique endemic flora and fauna locally [62]. The forests landscape reshaped from the onset of the Pleistocene and the hunting tradition of the large mammals which documented by the paleolithic phases in the study sites can be viewed as the ecological tradition for prehistoric people on one hand [59, 62], these settings in turn provide the environmental background for pottery production during the late Pleistocene in south China on the other hand. From this point of view, though aquatic resources were included in the dietary spectrum of the Neolithic people, the continuous use of the terrestrial resource might be explained by the adaptation to the ecological traditions that inherited by generations of the local Paleolithic people. Besides, plant resources, particularly those with higher sugar contents, have been reported as the potential factor resulting in human oral diseases during Paleolithic times [63]. This also raises another issue that related to the need for cooked food. Compared to animals, some plants can be processed to remove toxicity and became easily digestible [64]. Almost 70% (37 of 52) of the pottery samples that yielded plant biomarkers in the study sites, particular in Dingsishan site, together with the population size which reflected by total number of the human skeleton remains in the Dingsishan site (no less than 300 individuals [65]), suggests some kinds of terrestrial resources probably became part of the staple foods with social development and population growth during Neolithic times.
Furthermore, pottery itself, as a symbol of cultural evolution, represents a materialized form of public activities embedded in prehistoric social life and constantly linked with the transformation of prehistoric social structures, such as farming [5]. Although the transition to agriculture in South China is less clear than in North China and West Asia, where archaeological evidence made the hunter-gather to farmer connection much easier to trace [66, 67]. We suggest that such a case can also be reflected by pottery residue analysis. As shown in Fig. 7, our isotopic data combined with the published data from the Zengpiyan site (Additional file 1: Table S4), provide a remarkable diachronic change from extensive to the intensive tendency of resources used during different Neolithic stages, particularly within the early and middle Neolithic stages. Broader distribution of δ13C values demonstrates the resources derived from a wide range of C3 and C4 ecotone. This was further supported by the evidence of zooarchaeology and stable isotope analysis of human and animal bone apatite [19, 68], which showed a variety of niches around the study areas, including dense tropical forests, open C3 woodland with patches of C4 grassland [68], and such ecosystem may expand the hunting scope and point to relatively high or seasonal mobility for hunting and gathering food resources during these two stages.

Diachronic change of δ13C values of C16:0 and C18:0 in Neolithic South China
By contrast, the intensive use of C3 resources (Fig. 7) during the late Neolithic may closely map onto the plant and animal management or a more sedentary lifestyle. The scale of human selection on plant species in this stage can be seen in the number of which are common in the subtropical and tropical regions, including root plants (e.g. Dioscorea, Araceae, Taccaceae, Alismataceae), Coix and Triticeae Dumort [69, 70]. Despite domesticated animals being fewer in species [19], the feeding strategy of other animals may also occur, such as deer, water buffalo and wild boar which were found in large quantities in the study sites [19, 59]. Furthermore, an increased number of processing tools, such as smaller pottery vessels and grinding and digging tools, on a regional scale during the late Neolithic [71], also indicates that intentional food production or cultivation could arise in this stage [71, 72]. As illustrated in Fig. 7, compared to a mixture of C3 and C4 resources in the previously two stages, the intensive use of certain C3 plants and animals in late Neolithic, implied that the early mobile groups were possibly replaced by a new social structure of sedentary groups that build on food production during this time, laying the foundation for the subsequent agricultural development and social evolution in prehistoric South China.
In summary, resource processing is undoubtedly a major driving force for pottery invention and usage in South China. The ecological tradition and the prehistoric social evolution path hidden implied by the resource utilization pattern are worthy of further investigation. Perhaps it is owing to the unique ecological environments in South China, where the resource utilization patterns of early pottery and the subsequent social development behind pottery production are different from those of other regions in East Eurasia during Neolithic times.
The possibility of lacking the aquatic lipids in early pottery
Aquatic biomarkers including ω-(o-alkylphenyl) alkanoic acids (APAAs-C16–C22), isoprenoid fatty acids (phytanic acid, pristanic acid, 4,8,12-TMTD), and middle to long-chain dihydroxy fatty acids, are the diagnostic evidence for the identification of aquatic lipids [22, 41, 46, 73,74,75,76,77]. These biomarkers were frequently found in archaeological pots, and were identified in some of the earliest pottery [12, 13, 22, 33], providing the direct evidence for processing aquatic resources.
The absence of aquatic biomarkers in these pottery samples was somewhat surprising, since evidence from faunal and stable isotope of human and animal bone collagen shows clear consumption of aquatic foods. A similar phenomenon was also found in the previous work [24]. Both findings seemingly contradict the traditional assumption that the pottery in these sites was primarily used for processing aquatic resources [16, 18]. For this issue, we suggest the following possibilities.
Firstly, although the samples we selected in this study represent the typical pottery container for cooking [18, 43], the small sample size or type may have clouded the detection of aquatic lipids. Secondly, lipids were extracted in every potsherd, and their concentrations were all above the minimum amount required for interpretation (> 5 μg g−1), with the highest concentration up to 6666.7 μg g−1, indicative of a relatively good lipid preservation [23]. Even considering the lipid degradation during the long-term burial, it is still difficult to explain the presence of a series of terrestrial lipids but only absence of the aquatic ones. In fact, these compounds remain relative stable after burial, and were commonly found in a wide range of archaeological pottery [13, 22, 50, 73,74,75,76,77], though sometimes in trace amounts [22]. Thus, another possible explanation may be related to the cooking conditions. The cooking experiments in this study suggested that certain kinds of aquatic biomarkers could be detected in pottery at lower firing conditions. However, if these products were cooked in a shorter time, such as gently boiled in a few minutes, the firing conditions may not enough to form various aquatic biomarkers in pottery. Indeed, a large collection of well-preserved mollusk shell remains in the study sites may indicate that they have not undergone intense burning.
The last possibility may be related to dietary habits or cooking tools for aquatic resources during Neolithic period. Eating raw food has been viewed as a dietary habit in ancient and modern societies in South China, thus the possibility of eating certain kinds of aquatic food without processing, such as fish, should not be completely excluded. In addition, prehistoric people may choose other vessels to cook aquatic foods. Forest resources, such as bamboo and wood, are widespread in South China and adjacent Southeast Asia, and they are still common tools for the daily cooking of aquatic food in this vast area today [78, 79]. Although it is presently unclear how aquatic resources were processed and cooked in South China during the Neolithic times, the lack of evidence for aquatic products in our pottery samples suggests that they were probably prepared in other ways. In future, organic residue analysis on larger archaeological assemblages including pottery vessels, stone tools and other possible artifacts may help understand the culinary practices then.
Conclusion
This study conducted a systematic analysis of archaeological pottery residues and bone lipids from Dayan, Dingsishan and Zengpiyan sites in Guangxi province of China, revealing the extensive processing of terrestrial C3 and C4 resources in pottery vessels throughout the Neolithic times in this area. The persistent use of terrestrial resources and the remarkable diachronic changes from extensive use of C3 and C4 resources to the intensive use of certain kinds of C3 resources in different Neolithic stages further suggest that the resource utilization patterns in early pottery may be closely related to the ecological tradition and the subsequent social evolution in prehistoric South China.
Notably, the divergence between the findings of zooarchaeology as well as stable isotope analysis in these sites, which suggest an extensive exploitation of aquatic resources, and the lipid residue evidence from the potsherds in our study, which strongly indicates the processing of terrestrial resources, offers a new insight into the pottery use and culinary practice during Neolithic times in South China. However, a larger investigation is needed to further understand this discrepancy.
In fact, the commonality of pottery utilization built on cumulative archaeological evidence not only reflects the dietary innovation at the onset of the Neolithic period across different regions, but also implies the potential difference of culinary traditions under diverse environmental contexts. A combined research of ecological, economic and social factors based on organic residue analysis is of significant importance for exploring the pottery invention and utilization in ancient China and beyond.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files.
References
Zhang C. The discovery of early pottery in China. Documenta Praehistorica. 2002;29:29–35.
Kobayashi T. Re-thinking the first pottery use and the concept of the Incipient Jomon in Japan. Recent Palaeolithic Studies in Japan: Proceedings for Tainted Evidence and the Restoration of Confidence in the Pleistocene Archaeology of the Japanese Archipelago. 2004. 61–7.
Lu TL. Early pottery in south China. Asian Perspect. 2010;49:1–42. https://doi.org/10.1353/asi.2010.0003.
Wu X, Zhang C, Goldberg P, Cohen D, Pan Y, Arpin T, et al. Early pottery at 20,000 years ago in Xianrendong Cave, China. Science. 2012;336(6089):1696–700.
Jordan P, Zvelebil M. Ceramics before farming: the dispersal of pottery among prehistoric Eurasian hunter-gatherers. Walnut Creek: Left Coast Press; 2009.
Keally CT, Taniguchi Y, Kuzmin YV. Understanding the beginnings of pottery technology in Japan and neighboring East Asia. Rev Archaeol. 2003;24(2):3–14.
Zvelebil M. On the transition to farming in Europe, or what was spreading with the Neolithic: a reply to Ammerman (1989). Antiquity. 1989;63(239):379–83.
Zvelebil M. Mesolithic prelude and neolithic revolution. In: Zvelebil M, editor. Hunters in transition: Mesolithic societies of temperate Eurasia and their transition to farming. Cambridge: Cambridge University Press; 1986. p. 5–16.
Gibbs K. Pottery invention and innovation in East Asia and the Near East. Camb Archaeol J. 2015;25(1):339–51.
Kuzmin YV. The origin of pottery in East Asia and its relationship to environmental changes in the Late Glacial. Radiocarbon. 2010;52(2):415–20.
Cohen DJ. The advent and spread of early pottery in East Asia: new dates and new considerations for the world’s earliest ceramic vessels. J Austronesian Stud. 2013;4(2):55–92.
Lucquin A, Robson HK, Eley Y, Shoda S, Veltcheva D, Gibbs K, et al. The impact of environmental change on the use of early pottery by East Asian hunter-gatherers. Proc Natl Acad Sci. 2018;115(31):7931–6.
Bondetti M, Lucquin A, Savelev NA, Weber AW, Craig OE, Jordan P. Resource processing, early pottery and the emergence of Kitoi culture in Cis-Baikal: Insights from lipid residue analysis of an Early Neolithic ceramic assemblage from the Gorelyi Les habitation site, Eastern Siberia. Archaeol Res Asia. 2020;24:100225.
Feng Y, Wang Y. The environmental and cultural contexts of early pottery in south China from the perspective of behavioral diversity in the terminal Pleistocene. Quatern Int. 2022;608–609:33–48. https://doi.org/10.1016/j.quaint.2020.10.005.
Prendergast ME, Yuan J, Bar-Yosef O. Resource intensification in the late upper paleolithic: a view from southern China. J Archaeol Sci. 2009;36(4):1027–37. https://doi.org/10.1016/j.jas.2008.12.002.
Zhang C, Hung H-C. Later hunter-gatherers in southern China, 18 000–3000 BC. Antiquity. 2012;86(331):11–29. https://doi.org/10.1017/S0003598X00062438.
Hung H-C, Chi Z, Matsumura H, Zhen L. Neolithic transition in Guangxi: a long development of hunting-gathering society in Southern China. In: Matsumura H, Hung H-C, Zhen L, Shinoda K, editors. Bio-anthropological studies of early Holocene hunter-gatherer sites at Huiyaotian and Liyupo in Guangxi, China. Tokyo: National Museum of Nature and Science Tokyo; 2017. p. 205–28.
Fu X. The early Neolithic cultures from the Lingnan region (in Chinese). Archaeol Res. 2012;9:37–48.
Lv P. Zooarchaeological study of the shell mounds in the Yong Valley of Guangxi (in Chinese). Beijing: Chinese Academy of Social Sciences; 2010.
Zhu S, Li F, Chen X, Fu X, Hu Y. Subsistence and health in Middle Neolithic (9000–7000 BP) southern China: new evidence from the Dingsishan site. Antiquity. 2020;95(379):13–26. https://doi.org/10.15184/aqy.2020.214.
Liu X, Wang R, Hu Y. Stable carbon and nitrogen isotope analyses of human and animal skeletal remains unearthed at Zengpiyan and Dayan sites in Guilin. Chin Archaeol. 2022;22(1):191–200. https://doi.org/10.1515/char-2022-0016.
Cramp L, Evershed R. Reconstructing aquatic resource exploitation in human prehistory using lipid biomarkers and stable isotopes. In: Holland H, Turekian K, editors. Treatise on geochemistry. 2nd ed. Oxford: Elsevier; 2014. p. 319–39.
Liu X, Hu Y, Wang S, Yang Y, Ren M, Fu Y. Organic residue analysis of the early pottery from Dayan and Dingsishan Sites in Guangxi, China (Accepted in Chinese). Archaeology. 2023;7:104–20.
Zhang M. Organic residue analysis of hunter-gatherer pottery from Zengpiyan, China, to investigate vessel use and diet. Bristol: University of Bristol; 2020.
Rice PM. On the origins of pottery. J Archaeol Method Theory. 1999;6(1):1–54. https://doi.org/10.1023/A:1022924709609.
Kuzmin YV. Origin of old world pottery as viewed from the early 2010s: when, where and why? World Archaeol. 2013;45(4):539–56. https://doi.org/10.1080/00438243.2013.821669.
Zheng Z, Ma T, Roberts P, Li Z, Yue Y, Peng H, et al. Anthropogenic impacts on Late Holocene land-cover change and floristic biodiversity loss in tropical southeastern Asia. Proc Natl Acad Sci. 2021;118(40):e2022210118. https://doi.org/10.1073/pnas.2022210118.
Cramp LJE, Bull ID, Casanova E, Dunne J, Roffet-Salque M, Whelton HL, et al. Lipids in archaeology. In: Pollard AM, Armitage RA, Makarewicz CA, editors., et al., Handbook of archaeological sciences. Hoboken: Wiley; 2023. p. 529–56.
Evershed RP. Organic residue analysis in archaeology: the archaeological biomarker revolution. Archaeometry. 2008;50(6):895–924. https://doi.org/10.1111/j.1475-4754.2008.00446.x.
Evershed R, Harden S, Copley M, Mutherjee A. Identification of animal fats via compound specific δ13C values of individual fatty acids: assessments of results for reference fats and lipid extracts of archaeological pottery vessels. Documenta Praehistorica. 2002. https://doi.org/10.4312/dp.29.7.
Bocherens H, Drucker D. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol. 2003;13(1–2):46–53. https://doi.org/10.1002/oa.662.
Stott AW, Davies E, Evershed RP, Tuross N. Monitoring the routing of dietary and biosynthesised lipids through compound—specific stable isotope (δ13C) measurements at natural abundance. Naturwissenschaften. 1997;84(2):82–6. https://doi.org/10.1007/s001140050354.
Craig OE, Saul H, Lucquin A, Nishida Y, Taché K, Clarke L, et al. Earliest evidence for the use of pottery. Nature. 2013;496(7445):351–4. https://doi.org/10.1038/nature12109.
Dunne J, Mercuri AM, Evershed RP, Bruni S, di Lernia S. Earliest direct evidence of plant processing in prehistoric Saharan pottery. Nat Plants. 2016;3(1):16194. https://doi.org/10.1038/nplants.2016.194.
Han B, Sun Z, Chong J, Lyu N, Rao H, Yang Y. Lipid residue analysis of ceramic vessels from the Liujiawa site of the Rui State (early Iron Age, north China). J Quat Sci. 2022;37(1):114–22. https://doi.org/10.1002/jqs.3377.
Shoda S, Lucquin A, Sou CI, Nishida Y, Sun G, Kitano H, et al. Molecular and isotopic evidence for the processing of starchy plants in Early Neolithic pottery from China. Sci Rep. 2018;8(1):17044. https://doi.org/10.1038/s41598-018-35227-4.
Lampert CD, Glover I, Heron C, Stern B, Shoocongdej R, Thompson GB. Characterization and Radiocarbon Dating of Archaeological Resins from Southeast Asia. In: Jakes KA, editor. Archaeological Chemistry: Materials, Methods and Meaning. ACS Symposium Series. Washington D.C.: American Chemical Society; 2002. p. 84–109.
Reber EA, Kerr MT, Whelton HL, Evershed RP. Lipid residues from low-fired pottery. Archaeometry. 2019;61(1):131–44. https://doi.org/10.1111/arcm.12403.
Drieu L, Lucquin A, Cassard L, Sorin S, Craig OE, Binder D, et al. A Neolithic without dairy? Chemical evidence from the content of ceramics from the Pendimoun rock-shelter (Castellar, France, 5750–5150 BCE). J Archaeol Sci Rep. 2021;35:102682. https://doi.org/10.1016/j.jasrep.2020.102682.
Courel B, Robson HK, Lucquin A, Dolbunova E, Oras E, Adamczak K, et al. Organic residue analysis shows sub-regional patterns in the use of pottery by Northern European hunter-gatherers. Royal Soc Open Sci. 2020;7(4):192016. https://doi.org/10.1098/rsos.192016.
Cramp LJE, Ethier J, Urem-Kotsou D, Bonsall C, Borić D, Boroneanţ A, et al. Regional diversity in subsistence among early farmers in Southeast Europe revealed by archaeological organic residues. Proc Biol Sci. 1894;2019(286):20182347. https://doi.org/10.1098/rspb.2018.2347.
Fu X. The Dingsishan site and the prehistory of Guangxi, South China. Bull Indo-Pac Prehist Assoc. 2002;22:63–72.
Fu X. A preliminary study of prehistoric cultural development in Guangxi region (in Chinese). In: Deng C, Chen X, editors. Tao li cheng xi ji Essays in Honor of Ann Zhimin. Hong Kong: Center for Chinese Archaeology and Art, The Chinese University of Hong Kong; 2004. p. 194–204.
IA Cass (Institute of Archaeology Chinese Academy of Social Sciences). The Zengpiyan Site in Guilin (in Chinese). Beijing: Cultural Relics Press; 2003.
Hung H-C. Prosperity and complexity without farming: the South China Coast, c. 5000–3000 BC. Antiquity. 2019;93(368):325–41. https://doi.org/10.15184/aqy.2018.188.
Evershed RP, Copley MS, Dickson L, Hansel FA. Experimental evidence for the processing of marine animal products and other commodities containing polyunsaturated fatty acids in pottery vessels. Archaeometry. 2008;50(1):101–13. https://doi.org/10.1111/j.1475-4754.2007.00368.x.
Bondetti M, Scott E, Courel B, Lucquin A, Shoda S, Lundy J, et al. Investigating the formation and diagnostic value of ω-(o-alkylphenyl)alkanoic acids in ancient pottery. Archaeometry. 2021;63(3):594–608. https://doi.org/10.1111/arcm.12631.
Liu X, Hu Y. Diagenesis and mechanism of bone apatite from early and middle Neolithic animals in South China——Case study from Zengpiyan cave in Guangxi (in Chinese). Quaternary Sciences. 2021;41(1):189–200.
Colonese AC, Farrell T, Lucquin A, Firth D, Charlton S, Robson HK, et al. Archaeological bone lipids as palaeodietary markers. Rapid Commun Mass Spectrom. 2015;29(7):611–8. https://doi.org/10.1002/rcm.7144.
Shoda S, Lucquin A, Ahn J-h, Hwang C-j, Craig OE. Pottery use by early Holocene hunter-gatherers of the Korean peninsula closely linked with the exploitation of marine resources. Quat Sci Rev. 2017;170:164–73. https://doi.org/10.1016/j.quascirev.2017.06.032.
Ackman RG, Hooper SN. Examination of isoprenoid fatty acids as distinguishing characteristics of specific marine oils with particular reference to whale oils. Comp Biochem Physiol. 1968;24(2):549–65.
DeNiro MJ, Epstein S. Mechanism of carbon isotope fractionation associated with lipid synthesis. Science. 1977;197(4300):261–3. https://doi.org/10.1126/science.327543.
Jim S, Ambrose SH, Evershed RP. Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen and apatite: implications for their use in palaeodietary reconstruction. Geochim Cosmochim Acta. 2004;68(1):61–72. https://doi.org/10.1016/S0016-7037(03)00216-3.
DeNiro MJ, Epstein S. Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta. 1978;42(5):495–506. https://doi.org/10.1016/0016-7037(78)90199-0.
Tieszen LL, Boutton TW, Tesdahl KG, Slade NA. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia. 1983;57(1):32–7. https://doi.org/10.1007/BF00379558.
Tieszen LL, Fagre T. Effect of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. In: Lambert JB, Grupe G, editors. Prehistoric human bone: archaeology at the molecular level. Berlin: Springer Berlin Heidelberg; 1993. p. 121–55.
Evershed RP. Compound-specific stable isotopes in organic residue analysis in archaeology. In: Colombini MP, Modugno F, editors. Organic mass spectrometry in art and archaeology. Hoboken: Wiley; 2009. p. 389–432.
Copley MS, Berstan R, Dudd SN, Docherty G, Mukherjee AJ, Straker V, et al. Direct chemical evidence for widespread dairying in prehistoric Britain. Proc Natl Acad Sci. 2003;100(4):1524–9. https://doi.org/10.1073/pnas.0335955100.
Yu C. Preliminary research of subsistence patterns from Neolithic to bronze age in Lingnan and surrounding Area South China (in Chinese). Cult Relics South China. 2018;2:180–7.
Evershed RP, Stott AW, Raven A, Dudd SN, Charters S, Leyden A. Formation of long-chain ketones in ancient pottery vessels by pyrolysis of acyl lipids. Tetrahedron Lett. 1995;36(48):8875–8. https://doi.org/10.1016/0040-4039(95)01844-8.
Pearson R. The social context of early pottery in the Lingnan region of south China. Antiquity. 2005;79(306):819–28.
Zhao Y. Vegetation and climate reconstructions on different time scales in China: a review of Chinese palynological research. Veg Hist Archaeobotany. 2018;27(2):381–92. https://doi.org/10.1007/s00334-017-0655-6.
Wu X, Jin C, Cai Y, Zhang Y, Wang Y, Qin D, et al. Dental caries and mandibular anomalies of earlier modern humans from the Zhirendong site, Chongzuo, Guangxi (in Chinese). Acta Anthropologica Sinica. 2013;32(3):293.
Butterworth P, Ellis P, Wollstonecroft M. Why protein is not enough: the roles of plants and plant processing in delivering the dietary requirements of modern and early Homo. In: Hardy K, Kubiak-Martens L, editors. Wild harvest: plants in the hominin and pre-agrarian human worlds. Barnsley: Oxbow Books; 2016. p. 31–54.
Zhang P, Li F, Wang M. Analysis of dental caries and diet of the Neolithic population at the Dingsishan site, South China (in Chinese). Acta Anthropologica Sinica. 2018;37(3):393–405.
Fuller D, Kingwell-Banham E, Lucas L, Murphy C, Stevens C. Comparing pathways to agriculture. Archaeol Int. 2015;18:61–6. https://doi.org/10.5334/ai.1808.
Zhang C, Hung H-C. The emergence of agriculture in southern China. Antiquity. 2010;84(323):11–25. https://doi.org/10.1017/S0003598X00099737.
Liu X. Transition on hunting-fishing and gathering economy in Neolithic South China——Bioarchaeological study from Zengpiyan, Dayan, Dingsishan and Lingwu sites (in Chinese). Beijing: University of Chinese Academy of Sciences; 2021.
Zhang X, Huang C, Zhou Z, Olsen JW, Huang Q, Guan Y. Plant consumption by Early-Middle Neolithic peoples in Guangxi, South China: archaeobotanical evidence from the Dingsishan Site. Front Earth Sci. 2022. https://doi.org/10.3389/feart.2022.879908.
Li Z. Comparison of plant subsistence strategies at different geomorphic environments in Holocene, Lingnan region, China (in Chinese). Beijing: University of Chinese Academy of Sciences; 2016.
Liu L, Chen X. The archaeology of China: from the late paleolithic to the early bronze age. Cambridge world archaeology. Cambridge: Cambridge University Press; 2012.
Jiao T. Toward an alternative perspective on the foraging and low-level food production on the coast of China. Quatern Int. 2016;419:54–61. https://doi.org/10.1016/j.quaint.2015.06.060.
Copley MS, Hansel FA, Sadr K, Evershed RP. Organic residue evidence for the processing of marine animal products in pottery vessels from the pre-colonial archaeological site of Kasteelberg D east, South Africa: research article. S Afr J Sci. 2004;100(5):279–83. https://doi.org/10.10520/EJC96255.
Hansel FA, Copley MS, Madureira LAS, Evershed RP. Thermally produced ω-(o-alkylphenyl)alkanoic acids provide evidence for the processing of marine products in archaeological pottery vessels. Tetrahedron Lett. 2004;45(14):2999–3002. https://doi.org/10.1016/j.tetlet.2004.01.111.
Hansel FA, Evershed RP. Formation of dihydroxy acids from Z-monounsaturated alkenoic acids and their use as biomarkers for the processing of marine commodities in archaeological pottery vessels. Tetrahedron Lett. 2009;50(40):5562–4. https://doi.org/10.1016/j.tetlet.2009.06.114.
Lucquin A, Colonese AC, Farrell TFG, Craig OE. Utilising phytanic acid diastereomers for the characterisation of archaeological lipid residues in pottery samples. Tetrahedron Lett. 2016;57(6):703–7. https://doi.org/10.1016/j.tetlet.2016.01.011.
Lucquin A, Gibbs K, Uchiyama J, Saul H, Ajimoto M, Eley Y, et al. Ancient lipids document continuity in the use of early hunter–gatherer pottery through 9,000 years of Japanese prehistory. Proc Natl Acad Sci. 2016;113(15):3991–6. https://doi.org/10.1073/pnas.1522908113.
Bar-Yosef O, Eren MI, Yuan J, Cohen DJ, Li Y. Were bamboo tools made in prehistoric Southeast Asia? An experimental view from South China. Quatern Int. 2012;269:9–21. https://doi.org/10.1016/j.quaint.2011.03.026.
Weisskopf A. Elusive wild foods in South East Asian subsistence: modern ethnography and archaeological phytoliths. Quatern Int. 2018;489:80–90. https://doi.org/10.1016/j.quaint.2016.09.028.
Acknowledgements
We are grateful for the Archaeological Team of South China, Institute of Archaeology, Chinese Academy of Social Science (IA-CASS) support during sample selection and fieldwork. We would like to thank Research Fellow Xianguo Fu for systematic understanding of archaeological excavation in Guangxi province. Our work in the field was also facilitated by National Archaeological Museum of Zengpiyan Guilin, Guangxi province. We are also grateful to the Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences during sample pretreatment and analysis. Extending special thanks to Prof. Yunping Huang and Dr. Ying Zhang at Peking University for faunal analysis. We also thank Dr. Marvin Demicoli at University of Liverpool for English language revising.
Funding
This study was supported by the National Key R&D Program of China (No. 2022YFF0903500) and The Chinese Academy of Social Sciences innovation project (No. 2021KGYJ020). National Natural Science Foundation of China (No. 42072217). This fund was provided by the corresponding author Yimin Yang.
Author information
Authors and Affiliations
Contributions
XL and YY designed the research; YF provided the archaeological samples and backgrounds; XL performed the analysis; XL and MR analyzed the data and wrote the main manuscript text; SW, YY and YH revised the manuscript. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors consent.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Modern replica pottery vessels and cooking experiments. Figure S2. Comparison of lipid distributions in deer bone (a) direct acidified methanol procedure (b) two-step procedures. Table S1. The lipid results from cooking experiments. Table S2. δ13C values of bone components among different animals. Table S3. Compound-specific stable isotope data of potsherds from Dayan and Dingsishan sites. Table S4. Compound-specific stable isotope data of pottsherds from the Zengpiyan site.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, X., Ren, M., Fu, Y. et al. New insights into the use of Neolithic pottery in Guangxi of South China: organic residue analysis of experimental and archaeological pottery. Herit Sci 11, 201 (2023). https://doi.org/10.1186/s40494-023-01045-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40494-023-01045-9