
Abstract
The tailbeat frequency (TBF) together with tailbeat amplitude (TBA) of fish are tightly correlated with swimming speed. In addition, these parameters can be used as indicators of metabolic rate and general activity level, provided that appropriate calibration studies have been performed in the laboratory. If an implantable bio-logger could measure TBF and TBA, it would, therefore, have great potential as a tool to monitor swimming behaviours and bioenergetics over extended periods of time in free roaming fish within natural or farm environments. The purpose of this study was, therefore, to establish a method for deriving accurate TBF and variations in TBA from activity tags that log high-resolution acceleration data. We used 6 tagged Atlantic salmon (Salmo salar) of ≈1 kg and subjected them to two types of swim trials in a large swim tunnel system. Test speeds were either incrementally increased in 20-min intervals until steady swimming ceased, or constant speed of 60 cm s−1 was given in a 4-h sustained test. The TBFs were visually observed by camera and compared with computed values from the activity tags. In the incremental trials the TBF increased linearly with swimming speed, while it remained constant during the 4 h of sustained swimming. The TBFs measured by activity tags were within ± 0.1 beat s−1 of the visual measurements across the swim speeds tested between 30 to 80 cm s−1. Furthermore, TBF and its corresponding relative swim speed were consistent between trial type. The relative TBA increased with swimming speed as a power function, showing that the fish relies on changes in both amplitude and frequency of tail movements when swimming at higher speeds, while adjustments of amplitude only play a minor part at lower speeds. These results demonstrate that TBFs can be measured accurately via activity tags, and thus be used to infer swimming activities and bioenergetics of free roaming fish. Furthermore, it is also possible to estimate changes in TBA via activity tags which allows for more nuanced assessments of swimming patterns in free roaming fish.
Similar content being viewed by others

Introduction
Biotelemetry and biosensing devices have been used extensively in fish biological research for decades to obtain otherwise unobtainable individual-based knowledge of fish species in their natural environments [30, 45]. Various types of tags have been developed for a range of purposes that may be externally attached or implanted internally, and function either as data transmitters or data storage tags [11, 13]. The purpose of tagging studies has typically been to gain fundamental knowledge of the spatial and temporal distribution of wild fish, including foraging behaviours, migration patterns, and movements during reproduction events [2, 14, 25, 56]. Such work also has broader applications in conservation efforts and population managements by documenting the environmental preferences as well as the potential vulnerabilities of fish species when subjected to anthropogenic interferences [12, 13, 48, 55]. More recently, tags have also gained popularity in aquaculture research as an approach to assess fish welfare in sea cage environments [17, 46, 51].
Parameters typically measured in tagging studies are movements over extended periods of time [30, 45] or activity levels [16, 18, 41], often in combination with data of the physical environment, such as depth, temperature, and oxygen saturation [56, 59, 74]. In recent years, along with miniaturization and increased storage capacity of computer chips, commercially available tags have been developed to measure more sophisticated physiological parameters such as heart rate in fish in addition to other parameters [6, 9].
Biosensors or telemetry devices can provide a valuable link between highly controlled laboratory experiments and the actual conditions experienced by free swimming fish in their ambient environment. For instance, in the laboratory heart rate and acceleration of tagged fish can systematically be measured in resting conditions, at increasing activity levels, and in response to acute stress [9, 36, 37, 76]. Tag data can then be correlated with other parameters of interest, particularly the swimming speed and the metabolic rate of the fish [37, 76]. Once such calibration studies have been performed, it is then possible to infer additional information in free roaming tagged fish species subjected to less well-defined contexts. Specifically, obtaining accurate estimates of swimming speeds and metabolic rates indirectly via tagging devices would have tremendous potential in many areas of fish biological research, as they can describe the bioenergetics and swimming patterns at a high temporal resolution over prolonged timescales of fish in their ambient environment [24, 75].
Heart rate may not be the most precise indicator of swimming speed or metabolic rate, at least in salmonid species, because changes in the stroke volume (blood pumped per heartbeat) play a greater part in controlling cardiac output [61, 63]. As such, while heart rate still increases between inactivity and high swimming intensity, it becomes a crude predictor of swimming speeds and metabolic rates when assessing the entire range of activity found in fish, particularly at intermediate levels. Furthermore, heart rate can be drastically elevated owing to various stressors, while the fish is mostly inactive, and in those cases heart rate does not reflect swimming speeds at all, e.g., [37]. Tags that measure acceleration have also been investigated for their applicability to estimate swimming speeds and bioenergetics in fish. Similar to heart rate, activity proxies derived from acceleration data have been shown to somewhat correlate with swimming speeds and metabolic rates, at least when assessing very low and very high activity levels, e.g., [9]. However, these parameters also suffer from limitations in how accurately they may predict swimming speeds and metabolic rates over the full range of activity levels in fish [9, 22, 75, 76].
A potentially more precise candidate for inferring swimming speeds and metabolic rates in free roaming fish is the tailbeat frequency (TBF). It is well-documented that TBFs are tightly and linearly correlated with swimming speeds in various fish species tested in the laboratory, and that metabolic rate is sensitive to changes in swimming speeds. TBFs can thus function as proxies for metabolic rates at different activity levels [35, 37, 60]. The TBF can be derived from acceleration measurements, provided they are logged at a sufficiently high frequency (i.e., at least 2 × the expected maximum tailbeat rate). However, there are few studies, where electronic tags have been used to calculate TBF, and these are to our knowledge limited to older efforts on rainbow trout (Oncorhynchus mykiss) and Japanese flounder (Paralichthys olivaceus) [41, 42]. Interestingly, analogous work has been done in birds, where wing beat frequency has been estimated via accelerometer tags [21, 65].
In addition to the TBF, the tail beat amplitude (TBA) is required to obtain a full assessment of how the fish generates thrust at various activity levels. Estimates of TBA are generally more complicated to obtain and we are unaware of any previous efforts to compute a TBA from acceleration tags. We theorize that absolute values of TBA will be difficult to obtain based on acceleration, but that it will be possible to acquire a measure of changes in TBA over time by comparing the measured amplitudes in acceleration. Hence, it may be possible to derive relative changes in TBA from high frequency acceleration tags together with TBF.
To be sure that novel tag derived parameters can provide relevant information about free roaming fish, they first need to undergo proof of concept studies in controlled laboratory settings, where data collected from tags are validated [7, 27, 62, 76]. As such, tag derived TBFs, therefore, require confirmation with visually observed values at different swimming speeds before they can be used in the field.
In this study, we tested if a commercially available activity tag with a high frequency data logging capacity could measure the TBF and TBA in Atlantic salmon (Salmo salar) subjected to two types of swim tunnel trials. In the first trial, we performed a critical swim speed inspired test with incremental increases in flow speeds in 20-min intervals to assess a range of swimming activities. In the second trial, we used a sustained swimming test, where fish were subjected to a constant flow speed for 4 h to evaluate the consistency of TBF and TBA over time at the same activity level. Our primary aims were to investigate how well values computed from tag measurements correlated with visual measurements for TBF (this was not possible for TBA as amplitude is difficult to gauge visually) and correlate computed TBF and TBA values with swimming speeds.
Methods
Animal husbandry
Atlantic salmon post smolts (Aquagen) were kept in large indoor holding tanks (diameter: 3 m; volume: 5.3 m3) at the Matre Research Station, Institute of Marine Research, Norway. Tanks were supplied with 12 °C filtered seawater (34 ppt) via a flowthrough system (120 l min−1). A simulated natural photoperiod was provided, and fish were fed commercial feed (4.5 mm pellet size; Skretting) in excess each day via automated feeders. Fish were acclimated to these conditions for a minimum of 1 month before the experimental trials began. The experimental trials were performed in October and November 2020, and the use of animals was approved by the Norwegian Food Safety Authorities under ethics permit identification number 24444.
High frequency acceleration tags
Commercially available accelerometer Data Storage Tags (Technosmart axy-5 s activity tags, dimensions 22 × 13 × 10 mm, weight 4.5 g, www.technosmart.eu) were used to obtain the acceleration data. This data was then used to estimate TBF values and relative changes in TBA. In addition, the TBF estimates could be compared to visual TBF measurements. The tags were programmed in TechnoSmart X MANAGER software (v. 1.8.3) to log tri-axial acceleration (X, Y and Z) with a sampling frequency of 25 Hz within a range of G (± 4 G). Tag outputs were set to 8-bit values, mapping measured acceleration values within this range. The accelerometer tags were pre-calibrated by the manufacturer. In cases, where physical data in absolute units are of interest re-calibrations may be considered, e.g., [21]. However, in the present study we only report frequencies and relative changes.
Tagging of fish
Atlantic salmon weighing 1068 ± 26 g (Table 1) were tagged after anesthetizing with 150 mg L−1 Finquel for approximately 4 min and then placed inverted on a holding tray. During tag implantation, 75 mg L−1 Finquel was continuously flowing over the gills via a tube to keep the fish sedated. A Technosmart axy-5 s tag was then inserted into the abdominal cavity through an incision made aft of the pectoral fin. The incision was closed with 2 to 3 stitches, gently dried and then a thin layer of antibacterial Histoacryl® was applied to further seal the wound. The tagging procedure for each fish was completed in less than 5 min. A total of 8 fish were tagged for this study.
Swim tunnel system
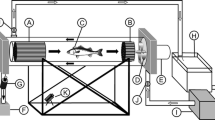
To test the efficacy of measuring swimming performance with activity tags, fish were swum individually in a 1905 l Brett-type swim tunnel setup as described previously by Remen et al. [54]. In brief, the cylindrical swim section was 248 cm long with an internal diameter of 36 cm. Controlled water currents within the tunnel was generated with a motor driven propeller (Flygt 4630, 11° propeller blade, Xylem Water Solutions Norge AS, Norway). The swim tunnel was connected via a large hose to the same header tank as used to supply water to the fish tanks, allowing for a continuous flow through the setup which maintained a 12 °C test temperature and ensured optimal oxygen levels at all times. A camera was deployed downstream behind the rear grid to record the trials and for visual estimates of TBF without disturbing the fish. At the rear end, a removable top lid allowed easy access into the tunnel for adding and removing fish.
Swim trials
The day before a swim trial, one tagged fish was transferred to the swim tunnel for acclimation overnight at a water flow rate of 20 cm s−1. This flow rate is generally too low to initiate continuous swimming efforts in Atlantic salmon of the size tested here and instead they typically balance on the floor of the tunnel. An overnight acclimation period is standard procedure in Atlantic salmon swim trials to allow recovery of acute handling stress and acclimation to a novel environment before the onset of swim tests, e.g., [37].
Three tagged fish (A–C, Table 1.) were swum individually on separate days following a typical critical swim speed protocol, e.g., [5] with the exception that the fish were removed from the tunnel before fatigue occurred, as inferred from the transition between steady swimming to the onset of burst and glide swimming behaviour. Starting from the initial overnight resting current speed of 20 cm s−1, for each fish on the day of the swim trial at 10:00 h, the water flow in the swim tunnel was increased by 10 cm s−1 every 20 min until the fish no longer swam steadily. Once the fish stopped swimming steadily and began exhibiting burst and glide behaviour, signifying transition to partially anaerobically fuelled swimming and imminent fatigue, e.g., [72], it was removed from the tunnel. The fish was then immediately euthanized with an anaesthetic overdose of Finquel, whereafter the weight and fork length were recorded, and the activity tag recovered.
While fish were swimming at each different speed interval, TBF (time taken for 100 tail beats) were measured manually using video camera footage and a stopwatch. Visual measurement of TBFs is a simple procedure when the fish is readily observed on camera. In addition, the swim trials were recorded, so that visual measurements could be confirmed later on.
Three other tagged fish (D–F, Table 1) were similarly tested individually on separate days. However, here a sustained swimming speed test was used following a similar procedure to that of Hvas and Oppedal [31]. Starting from the initial overnight resting current speed of 20 cm s−1 at 09:54 h, the waterflow was increased by 10 cm s−1 every 2 min, so that by 10:00 h the waterflow speed was 60 cm s−1. A sustained speed of 60 cm s−1 was chosen here to represent an intermediate challenge slightly above the expected optimal cruising speed for minimum cost of transport, but still well within the aerobic limit of Atlantic salmon [31, 38]. Each fish was then swum at this flow rate for the next 4 h. During this time manual measurements of the TBF were made every 30 min using the camera footage and a stopwatch similar to the incremental swim trials. After 4 h of swimming, the fish was removed from the tunnel and euthanised with an anaesthetic overdose of Finquel, fish weight and fork length were recorded, and the activity tag was recovered.
The remaining two of the tagged fish did not cope adequately in the tunnel environment as they were reluctant to swim consistently for the full trial period. These were, therefore, discarded from further analyses, providing 3 replicates for each trial type.
Tag data extraction and post processing analysis
Raw data time series were downloaded and converted from 8-bit binary values into acceleration values in G (± 4 G) in X MANAGER and stored as data sets in.csv file format.
Because the tags measured acceleration, some considerations were made before extracting TBF and TBA. When implanting tags in fish there will be an offset between the accelerometer's sensing axes and the BODY coordinate system, i.e., the (imagined) coordinate system fixed to the fish's centre of gravity (Fig. 1). The offset between the longitudinal axes (i.e., X and X,b) will be negligible compared with the offset between the other axes if the tag is implanted, such that it rests against the abdomen of the fish as is the common practice, e.g., [10].

Illustration of how the accelerometer measurement axes of the tag (X, Y and Z) relate to orientation axes of the fish (X,b, Y,b and Z,b). Assuming that X is aligned with X,b, pitch (Θ) will define the angle between X and the horizontal plane, while the roll (ɸ) and yaw (ψ) angles will define used to describe the deviation between the Y and Y,b and Z and Z,b, respectively
By assuming that the tag remains stationary inside the fish and that the raw data means for the entire duration of the measurements represent the stationary acceleration offsets, the angular offsets for roll (ɸ) and pitch (θ) and yaw (ψ) can be calculated using
and
where AX, AY and AZ are the measured accelerations along the different axes. The measured accelerations can then be rotated to the BODY coordinate system using
where s and c denote sine and cosine, respectively [20]. Following rotation, gravity was subtracted from the Z-axis and data were filtered using a 4th order Butterworth bandpass filter with low and high frequency limits of 0.8 and 5 Hz, respectively, to suppress signal noise outside the expected TBF frequency band. An example of the effect of these processing steps is given in Fig. 2.

Effect of processing steps, exemplified using data from Fish C. The leftmost column shows the raw measurements, the middle column data after rotation using Eq. 4, while the rightmost column shows the resulting data after subtracting gravity and smoothing through filtering
The rotated and filtered acceleration time series were represented in the time domain. However, in this case the frequency components were of particular interest, thus it was desirable to present the data also in the frequency domain to evaluate the signal's frequency constituents and amplitude in G (1G = 9.81 m/s2). To do this, the data must first be transformed using specific signal processing methods. One such transform is the 'Fast Fourier Transform' (FFT) which returns the frequency components for the entire signal duration to which it is applied. While FFT is a proven method for determining the frequency composition of complex signals, it analyses the whole period as one, and thus cannot capture changes in the frequency composition that occur during the signal period. To acquire this ability, a modified FFT approach called the 'Short Time Fast Fourier Transform' (STFFT) was applied instead. The STFFT algorithm applies a rolling time window to the input signal, applies the FFT to the data within this window and then shifts by a pre-set amount to overlap with itself before re-applying the FFT to the data within the new window. This is repeated for the entire duration of the signal. By choosing the appropriate window length and overlap, the frequency and amplitude content of the input signal can be analysed with respect to time.
For the rotated acceleration data, the acceleration component AY,b is aligned with the expected direction of the tail beat (i.e., laterally outwards from the fish body), hence any distinct tail beat frequency would be expected to appear in the frequency spectrum of AY,b. STFFT using a 20 s window and 75% overlap was, therefore, applied to this axis. For each (overlapping) window, the highest frequency spectrum peak was considered to represent both TBF (placement on the frequency axis) and TBA (peak height) resulting in one TBF and TBA result per 5 s (Fig. 3).

Example frequency spectrum from Fish C at 30 cm s−1. The horizontal axis denotes frequency, while the vertical axis denotes the proportion of the total measured acceleration that is calculated for the different frequency components. For a fish swimming at a steady speed, the former of these will correspond to the TBF, while the second will relate to the TBA
Unlike for TBF, the value obtained through STFFT for TBA will not represent the absolute value of the TBA as acquiring this parameter from accelerations entails integrating the acceleration time series twice, which tends to amplify measurement errors and inaccuracies. Moreover, since the tags were placed intraperitoneally, the conversion would also require translating and transforming accelerations measured in the abdomen to those exhibited by the tail of the fish, which would in turn depend strongly on the fish morphology and other unknown factors. However, provided that the tag position within the fish is fixed after implantation, variations in amplitude should also be expressed through variations in the height of the spectral peaks acquired through the STFFT. All analyses based on TBA were, therefore, made using the relative TBA, i.e., a measure of how much the TBA changes between different speeds.
To summarize, this tag data extraction and post processing analysis results in a processing flow consisting of five main steps as illustrated in Fig. 4.

Flow chart showing the processing procedure for acquiring tailbeat frequency (TBF) and tailbeat amplitude (TBA) data from the accelerometer tag measurements
Statistical analysis
For the incremental and sustained swim trials, correlations between observed TBFs taken via the camera footage and tag estimated TBFs, were tested using Pearson’s correlation coefficient. For the sustained swim trial, linear regression using the mean hourly TBF estimates from tag measured data was used to test for any change in TBF over the 4-h period. To compare between swim trials, the ratio of TBF measured via the tags to BL s−1 between swim trial type at the flow speed of 60 cm s−1 was tested using a one-way ANOVA. For all tests, the significant difference level was set at p < 0.05. All analyses were conducted using SPSS 26 statistical software package. Data are reported as mean ± standard error of the mean unless specified otherwise.
Results
Tailbeat frequency
In the incremental swim speed tests the TBF increased linearly with swimming speed from 30 to 80 cm s−1 (Fig. 5A–C). Above 80 cm s−1, steady swimming ceased for fish B and C as they began to show initial signs of fatigue, so the swim trial for both these fish ended, while for fish A, erratic changes in swimming pattern did not occur until after 90 cm s−1 (Fig. 5A–C). For the 4-h sustained swim speed tests at 60 cm s−1, the fish exhibited consistent swimming behaviour for the full duration of the test (Fig. 5D–F).

Incremental (A–C) and sustained (D–F) swim trials of tagged fish. For all graphs, black line represents tag estimated mean tailbeat frequency (TBF) of 10 s subset measurements with 50% overlap between subsets. For fish A–C, red line indicates tag estimated mean TBF for each 20-min interval at a specific current speed. Grey circles with a grey line indicate mean frequency of 3 visual estimates taken from camera recordings within each of those respective intervals. Error bars represents mean ± s.e.m. For fish D–F, vertical dashed lines indicate each hour from the start and end point of the 4-h sustained swim speed tests at 60 cm sec−1. Here, red line indicates tag estimated mean tail beat frequency for the complete trial, and grey circles with a grey line indicate visual point estimates of tailbeat frequency in 30-min intervals
In the incremental swim speed tests, the mean TBF measured from camera observations ranged from 1.53 beats s−1 ± 0.08 at 30 cm s−1 to 2.65 beats s−1 ± 0.06 at 80 cm s−1, and the TBF derived from acceleration measurements ranged from 1.40 beats s−1 ± 0.05 at 30 cm s−1 to 2.67 beats s−1 ± 0.06 at 80 cm s−1 (Fig. 6A). There was a strong linear correlation between the camera observed and tag estimated TBFs (Pearson’s r2 = 0.99, P < 0.001 (Fig. 6B). In the sustained swim trials, the overall mean TBF observed via the camera were 2.17 ± 0.07 beats s−1 and for the tag, 2.19 ± 0.03 beats s−1 (Fig. 6C). There was a strong linear correlation between the mean hourly camera observed and tag estimated TBFs (Pearson’s r2 = 0.88, P < 0.001) (Fig. 6D). Using tag estimated TBF only, there was no significant change in the mean hourly TBF over the 4-h period showing that the TBF remained constant at constant swimming efforts over time (F1,10 = 4.5, P = 0.06).

Observed and tag computed tailbeat frequencies (TBF) in the incremental swim speed trial (A) and the sustained swim trial (C), where error bars are mean ± s.e.m. Followed by correlation plots between tag estimated and observed TBF in their corresponding trials on the left panels (B and D)
Using tag measured TBFs as a proxy for swimming speed in the sustained swim trial at the flow rate of 60 cm s−1 the average tag TBF of 2.19 ± 0.03 beats s−1 equated to a swim speed of 1.28 ± 0.01 BL s−1 (mean fork length of 46.8 ± 0.2 cm). This was consistent with the incremental swim trial during the 20 min period when the flow rate was also 60 cm s−1 as the average tag measured TBF of 2.16 ± 0.07 beats s−1 equated to a swim speed of 1.30 ± 0.02 BL s−1 (mean fork length 46.1 ± 0.8 cm). As such, when comparing the mean ratio of TBF to BL s−1 at this specific flow speed between the trial types, there was no difference (TBF: BL. s−1: Incremental = 1.66 ± 0.07; Sustained = 1.70 ± 0.03; F1,4 = 0.5, P = 0.5).
Changes in tailbeat amplitude and thrust with increasing swimming speed
There was a distinct relationship between the changes in relative TBA and the swimming speed of the fish (Fig. 7A), with the amplitude appearing to stay constant for the lower speeds (30–40 cm−1) and then increasing substantially across the higher speeds (50–80 cm−1). This relationship was best described with a power function providing an R2 of 0.998. Furthermore, the changes in TBF with increasing swimming speed were well-described by a linear regression, where R2 was 0.998 (Fig. 7B). The product of TBF and TBA across swimming speed could then provide a visualization of the relative increase in thrust generated by the fish, and this relationship followed a power function (R2 of 0.999) (Fig. 7C).

Relative tail beat amplitudes at increasing current speeds as compared to 30 cm s−1 fitted with a power function (A), and tailbeat frequency at increasing current speeds fitted with a linear function (B). In C, the changes in thrust generated relative to 30 cm s−1 is shown and fitted with a power function, where thrust is expressed as the product of the mean tailbeat amplitude and frequency at each current speed. Data in A and B are mean ± s.e.m
Discussion
In this study, we investigated whether commercially available acceleration tags with a high frequency logging capacity could be used to accurately compute TBFs in Atlantic salmon across a range of swimming speeds as well as providing consistent estimates over time at a constant intermediate swimming speed. Since estimates of TBFs from tags were within ± 0.1 beats s−1 of visual counts across the assessed swimming speeds and trial types, we conclude that acceleration tags effectively can be used to measure TBFs with enough accuracy to be used as a proxy for swimming speeds and bioenergetics in free roaming fish. In addition, we were able to compute changes in relative tail beat amplitude (TBA) and found that this parameter increases with swimming speed in a pattern resembling a power function. Combined, TBF and TBA derived from acceleration tags can thus provide additional assessments of how free roaming fish generate thrust in different scenarios.
Tailbeat frequency as a proxy for swimming speed and metabolic rate
Acceleration tags have been used to infer swimming speeds and metabolic rates in fish on several occasions (e.g., [22, 57, 73, 75]. However, acceleration data has previously mostly been sampled at lower frequencies, typically 1 Hz [3, 4, 76]. In consequence, categorization of swimming activity and associated energetic costs in such studies tends to be limited to crude categorizations, such as resting, routine or burst swimming behaviours [1, 9]. By calculating the TBF from high resolution acceleration tag data as in the present study, it would be possible to define a more fine-grained scale of swimming activity levels that would give greater precision in categorising the movements of free roaming fish. This will be particularly useful at intermediate activity levels, which are presumably the energetic states that fish occupy most of the time, for instance during foraging or migration which are associated with optimal cruising speeds at minimum transport costs [15, 71].
TBF tag data can also be expected to provide more accurate swimming speed estimates than other measurable parameters, such as heart rate and instantaneous acceleration, since it is directly linked with the undulatory locomotion. In the present study, TBF increased linearly from approximately 1.4 to 2.6 beats s−1 with increasing swimming speeds from 30 to 80 cm s−1. Similar strong relationships between TBFs and swimming speeds are well-documented in various fish species [35, 37, 60], and highlights the potential for using previously established regression lines to estimate swimming speeds from TBF data in free-swimming tagged fish.
The metabolic rate is highly sensitive to changes in activity, and the energetic requirements at increasing swimming speeds follow a power function [50]. This relationship reflects the hydrodynamics of swimming as the drag force increases in proportion to the square of the speed, meaning that the power required to overcome drag increases with the cube of the speed [66, 70], making it energetically more and more expensive to increase swimming efforts further. For instance, in Atlantic salmon the increase in aerobic metabolic rate between low and high swimming speeds approximates a power function and increases with a factor of 3 to 6, depending on the acclimation temperature [32]. Since the metabolic rate response to increasing swimming speeds is well-understood in fish and, moreover, is readily measured with swim tunnel respirometry in laboratory experiments, robust calibration curves can be gathered, so that TBF tag data also can be used as a proxy of metabolic rate.
Our estimates of relative TBA as a function of swimming were best described by a power function, similar to the metabolic requirement at increasing swimming speeds. Meanwhile, the TBF increased linearly with swimming speed. However, the drag forces increase in proportion to the square of the speed, as discussed above. To explain the observed propulsion of the fish, the amplitude, therefore, needed to increase relatively more at higher speeds, as shown here. Furthermore, the product of TBF and TBA can be used to estimate the relative thrust generated by the fish. Owing to the patterns of TBF and TBA with increasing swimming speed, thrust was also best described as a power function. This reflects the above-mentioned hydrodynamics of swimming and corresponding metabolic requirements to overcome increasing drag forces [66, 70]. Changes in TBA can, therefore, provide additional information on how fish generates thrust and contribute to making accelerometer tag data even more accurate in predicting swimming speeds in ambient environments.
Limitations of tailbeat frequency tags to infer swimming speeds and bioenergetics in fish
While the correlations between TBF, swimming speed, and associated energetic costs are well-established, they are also context dependent and influenced by various biological and environmental factors [35, 52, 60]. These must, therefore, be considered for proper interpretations of TBF tag data from fish in ambient environments and may limit the reliability of such tag data if parameters of unknown effects are involved. Thus, to fully exploit the possibility of TBF tags and to ensure the most accurate data interpretation, a nuanced suite of lab-based calibration studies is ideally required on a per species basis. In well-studied fish species, such as salmonids, it will be possible to amass correlation curves measured in many different contexts to strengthen data interpretations of TBF tags. However, TBF tags may have less value as a tool for energetic monitoring in less studied species, where swimming capacities and metabolic rates have not been measured systematically. Nevertheless, it will still be possible to describe the scope of TBFs used and the frequency distribution to quantify general swimming patterns through time when studying new fish species.
Apart from species differences in morphology and physiology, the size of the fish is presumably the most important factor to influence TBF and its correlation with swimming speed and metabolic rates. Specifically, smaller fish can attain a higher TBF and swim at higher relative swimming speeds (e.g., body lengths s−1), but larger fish are able to swim faster in absolute units (e.g., cm s−1) [35, 52]. The same TBF will, therefore, not correspond to the same swimming speed in fish of different size classes. It is thus important to consider size effects, especially if individuals remain tagged for prolonged periods and grow substantially in size. In addition, metabolic rates also scale with size in a somewhat predictable manner, where smaller individuals have a higher mass specific metabolic rate [43, 52].
Since the metabolism of ectothermic fish is greatly dependent on temperature this factor is also important to consider. Typically, metabolic rates increase with a factor of 2–3 per 10 °C, but thermal sensitivity depends on species, the specific portion of its thermal niche, and acclimation history [44]. Interestingly, the correlation between TBF and swimming speed is more consistent across temperatures than metabolic rate, except at thermal extremes which pose a limit to maximum swimming capacity [32, 35]. Hence, TBF as a predictor of swimming speed is less sensitive to fluctuations in temperature compared to metabolic rate.
Other factors worth considering include nutritional states and body condition [39, 47], life-stages and particularly sexual maturation [29, 68], impacts of diseases and parasites [33, 67], and possibly exposure to toxic substances in polluted environments [23, 49]. As such, when attempting to monitor the physiology and behaviour of free-swimming fish, there will always be unknown factors that could alter correlations between TBF, swimming speed and metabolic rate. Such uncertainties are an inherent challenge when studying animals outside the laboratory.
Applications and further development of tailbeat frequency tags
We envision several useful applications with TBF tags in future studies, particularly in aquaculture research with regard to fish welfare in salmon sea cages. Within these vast fish group sizes, the scope for detailed data acquisition from tagged sentinel fish is large [17, 69], and vital when moving towards the knowledge-based concept of precision fish farming [19]. In this, TBF should allow for recording behavioural modes and estimating the physical demand of swimming in fish groups driven by environmental features and social interactions (see [53] for established observation methods). For example, in response to high current speed, caged salmon change their group behaviour from circular schooling to a static group position directed against the inflowing water [40] which is likely to dampen current exposure within the fish school [34]. Moreover, if an algorithm like the one used in this study were to be implemented in acoustic tags carrying accelerometers, these could be set to transmit the computed tail beat rate in real time. This data could then be combined with measurements of absolute position [26] and movement speed [27, 28] in the cage, which would enable more detailed studies of the link between TBF and energetic expenditure and actual movement in sea cages.
Of fundamental interest, TBF can also be used to shed new light into questions such as individual fish need for rest and switch in behavioural mode connected with various photo-regimes and observed diel heart rate variation with night and day length [69], recovery from exhausting operational procedures, such as mechanical parasite treatment [18], and whether adverse conditions such as high temperature [32] or internal processes as sexual maturation or fasting will induce the urge for migration and thus change in swimming pace or pattern. Sentinel fish with TBF tags may also be important tools for documenting fish coping ability in novel aquaculture rearing technologies and systems. Possible applications in this area could include enabling continuous logging in harsh and remote ocean farming sites, serving as a component in early warning of negative buoyancy by increased swimming speed in submerged salmon cages [58], as well as providing documentation and tuning of water current exposure in closed containment systems [64].
Tags that measure TBF also have potential for providing new knowledge on fish species in the wild, for instance during periods of migration to feeding or spawning grounds, but also to generally obtain a more detailed understanding of natural activity patterns over extended periods of time. Here, some practical considerations are methods of tag retrieval as high-frequency acceleration tags presently only function as implantable data storage tags.
While the TBA parameter computed in this study provided a relative measure of the changes in TBA rather than the absolute value (as was the case for TBF), it was apparent that higher TBA-values correlated with higher swimming speeds. Furthermore, the patterns observed corresponded well with hydrodynamic predictions with regard to thrust needed to overcome drag at increasing speeds. Although this parameter may not be as easy to link directly with energetics, it can be combined with TBF to estimate the relative thrust generated by the fish. This provides a unitless measure of swimming output that can be considered analogous to the cardiac output which consists of heart rate multiplied by stroke volume. Similarly, stroke volume in fish is generally measured in relative changes, while heart rate implicitly is expressed as a frequency. Obtaining both TBA and TBF, therefore, allow for a more holistic assessment of swimming energetics.
Managing data from tailbeat frequency tags
The approach used in the present study to determine TBF and TBA is an adaptation of well-known signal processing techniques. Data were successfully rotated into the BODY coordinate system, thus allowing direct comparison of AY,b data based on the assumption that the mean of all recorded data represents the acceleration offset. This assumption is only valid if the tags do not shift within the peritoneal cavity during data capture. Future trials can address this by suturing tags in place for improved implantation control and easier result comparison between individuals. Alternatively, the offsets can be periodically re-calculated, compared to previous values and updated accordingly.
Furthermore, care must be taken when calculating the offsets (Eqs. 1, 2 and 3). Accurate offset calculations preclude that [AX, AY, AZ]T≉0. If [AX, AY, AZ]T≈0, i.e., below the signal noise level, AX,b, AY,b and AZ,b should be initialized to 0. Furthermore, for θ = 90°, Eq. 4 is singular (i.e., the denominator becomes 0) and cannot be solved. In free-swimming fish, this may occur when the fish is free to manoeuvre in all three dimensions (although salmon are unlikely to moving at as steep angles as θ = 90°). Future implementations of this approach should, therefore, either include a singularity check or use a singularity-free representation of rotation (e.g., quaternions).
Selection of signal processing sliding window width (20 s) and overlap (75%) was a trade-off between calculation robustness, computational demand and (timely) resolution for the results. For the signals measured in this study using a shorter window introduced more noise compared to longer windows due to the FFT averaging effect. Longer windows did not produce improved results while increasing computational demand. The parameters used during processing are, therefore, considered a reasonable compromise.
The filtering bandpass parameters (0.8 and 5 Hz) were chosen to suppress the effect of low and high frequency contributions outside the range of the expected TBF for Atlantic salmon, e.g., [37]. These parameters are highly species and size class specific and must be adapted accordingly with an appropriate TBF range (or for the general case). Finally, future tests using accelerometers to extract TBF and TBA should use sensor settings to maximize the signal to noise ratio, i.e., the lowest possible sensing range and the highest possible bit resolution.
The continuous development of more sophisticated implantable bio-loggers makes for an exciting future in fish biological research. Moreover, novel methods within biotelemetry and biosensors will help close the gap further between controlled laboratory trials and the more varied and unpredictable environments experienced by fish when allowed to swim around freely [8].
Data availability
Raw data will be made available upon request.
References
Algera DA, Brownscombe JW, Gilmour KM, Lawrence MJ, Zolderdo AJ, Cooke SJ. Cortisol treatment affects locomotor activity and swimming behaviour of male smallmouth bass engaged in paternal care: a field study using acceleration bio loggers. Physiology & Behaviour. 2017;181:59–68.
Aarestrup K, Økland F, Hansen MM, Righton D, Gargan P, Castonguay M, Bernatchez L, Howey P, Sparholt H, Pedersen MI, McKinley RS. Oceanic spawning migration of the European eel (Anguilla anguilla). Science. 2009;325(5948):1660–1660.
Brownscombe JW, Lennox RJ, Danylchuk AJ, Cooke SJ. Estimating fish swimming metrics and metabolic rates with accelerometers: the influence of sampling frequency. J Fish Biol. 2018;93(2):207–14.
Brown DD, Kays R, Wikelski M, Wilson R, Klimley AP. Observing the unwatchable through acceleration logging of animal behaviour. Anim Biotelemetry. 2013;1(1):1–16.
Brett JR. The respiratory metabolism and swimming performance of young sockeye salmon. J Fish Res Board Can. 1964;21:1183–226.
Brijs J, Sandblom E, Axelsson M, Sundell K, Sundh H, Huyben D, Broström R, Kiessling A, Berg C, Gräns A. The final countdown: continuous physiological welfare evaluation of farmed fish during common aquaculture practices before and during harvest. Aquaculture. 2018;495:903–11.
Brijs J, Sandblom E, Rosengren M, Sundell K, Berg C, Axelsson M, Gräns A. Prospects and pitfalls of using heart rate bio-loggers to assess the welfare of rainbow trout (Oncorhynchus mykiss) in aquaculture. Aquaculture. 2019;509:188–97.
Brijs J, Føre M, Gräns A, Clark TD, Axelsson M, Johansen JL. Bio-sensing technologies in aquaculture: how remote monitoring can bring us closer to our farm animals. Philos Trans R Soc B. 2021;376(1830):20200218.
Clark TD, Sandblom E, Hinch SG, Patterson DA, Frappell PB, Farrell AP. Simultaneous biologging of heart rate and acceleration, and their relationships with energy expenditure in free-swimming sockeye salmon (Oncorhynchus nerka). J Comp Physiol B. 2010;180(5):673–84.
Cooke SJ, Thorstad EB, Hinch SG. Activity and energetics of free-swimming fish: insights from electromyogram telemetry. Fish Fish. 2004;5(1):21–52.
Cooke SJ, Woodley CM, Brad EM, Brown RS, Nielsen JL. Advancing the surgical implantation of electronic tags in fish: a gap analysis and research agenda based on a review of trends in intracoelomic tagging effects studies. Rev Fish Biol Fisheries. 2011;21:127–51.
Costa DP, Breed GA, Robinson PW. New insights into pelagic migrations: implications for ecology and conservation. Annu Rev Ecol Evol Syst. 2012;43(73):2012.
Crossin GT, Heupel MR, Holbrook CM, Hussey NE, Lowerre-Barbieri SK, Nguyen VM, Raby GD, Cooke SJ. Acoustic telemetry and fisheries management. Ecol Appl. 2017;27(4):1031–49.
Dean MJ, Hoffman WS, Zemeckis DR, Armstrong MP. Fine-scale diel and gender-based patterns in behaviour of Atlantic cod (Gadus morhua) on a spawning ground in the Western Gulf of Maine. ICES J Mar Sci. 2014;71(6):1474–89.
Drenner SM, Clark TD, Whitney CK, Martins EG, Cooke SJ, et al. A synthesis of tagging studies examining the behaviour and survival of anadromous salmonids in marine environments. PLoS ONE. 2012;7(3): e31311.
Føre M, Alfredsen JA, Gronningsater A. Development of two telemetry-based systems for monitoring the feeding behaviour of Atlantic salmon (Salmo salar L.) in aquaculture sea-cages. Comput Electron Agric. 2011;76(2):240–51.
Føre M, Frank K, Dempster T, Alfredsen JA, Høy E. Biomonitoring using tagged sentinel fish and acoustic telemetry in commercial salmon aquaculture: a feasibility study. Aquacult Eng. 2017;78:163–72.
Føre M, Svendsen E, Alfredsen JA, Uglem I, Bloecher N, Sveier H, Sunde LM, Frank K. Using acoustic telemetry to monitor the effects of crowding and delousing procedures on farmed Atlantic salmon (Salmo salar). Aquaculture. 2018;495:757–65.
Føre M, Frank K, Norton T, Svendsen E, Alfredsen JA, Dempster T, Eguiraun H, Watson W, Stahl A, Sunde LM, Schellewald C. Precision fish farming: a new framework to improve production in aquaculture. Biosyst Eng. 2018;173:176–93.
Fossen TI. Marine control systems–guidance. navigation, and control of ships, rigs and underwater vehicles. Marine Cybernetics, Trondheim, Norway, Org. Number NO 985 195 005 MVA, www.marinecybernetics.com, ISBN: 82 92356 00 2; 2002
Garde B, Wilson RP, Fell A, Cole N, Tatayah V, Holton MD, Rose KA, Metcalfe RS, Robotka H, Wikelski M, Tremblay F, Whelan S, Elliott KH, Shepard EL. Ecological inference using data from accelerometers needs careful protocols. Methods Ecol Evol. 2022;13:813–25.
Gleiss AC, Dale JJ, Holland KN, Wilson RP. Accelerating estimates of activity-specific metabolic rate in fishes: testing the applicability of acceleration data-loggers. J Exp Mar Biol Ecol. 2010;385(1–2):85–91.
Grosell M, Pasparakis C. Physiological responses of fish to oil spills. Ann Rev Mar Sci. 2021;13:137–60. https://doi.org/10.1146/annurev-marine-040120-094802.
Halsey LG, Shepard ELC, Quintana F, Laich AG, Green JA, Wilson RP. The relationship between oxygen consumption and body acceleration in a range of species. Comp Biochem Physiol A: Mol Integr Physiol. 2009;152(2):197–202.
Harrison PM, Gutowsky LFG, Martins EG, Patterson DA, Leake A, Cooke SJ, Power M. Diel vertical migration of adult burbot: a dynamic trade-off among feeding opportunity, predation avoidance, and bioenergetic gain. Can J Fish Aquat Sci. 2013;70(12):1765–74.
Hassan W, Urke HA, Kristensen T, Ulvund JB, Alfredsen JA. System for real-time positioning and monitoring of fish in commercial marine farms based on acoustic telemetry and internet of fish (iof). In: The 29th International Ocean and Polar Engineering Conference. OnePetro; 2019
Hassan W, Føre M, Pedersen MO, Alfredsen JA. A new method for measuring free-ranging fish swimming speed in commercial marine farms using Doppler principle. IEEE Sensors J. 2020;20(17):10220–7.
Hassan W, Føre M, Urke HA, Ulvund JB, Bendiksen E, Alfredsen JA. New concept for measuring swimming speed of free-ranging fish using acoustic telemetry and Doppler analysis. Biosys Eng. 2022;220:103–13.
Hinch SG, Standen EM, Healey MC, Farrell AP. Swimming patterns and behaviour of upriver-migrating adult pink (Oncorhynchus gorbuscha) and sockeye (O. nerka) salmon as assessed by EMG telemetry in the Fraser River, British Columbia. Canada Hydrobiologia. 2002;483:147–60.
Hussey NE, Kessel ST, Aarestrup K, Cooke SJ, Cowley PD, Fisk AT, Harcourt RG, Holland KN, Iverson SJ, Kocik JF, Mills Flemming JE. Aquatic animal telemetry: a panoramic window into the underwater world. Science. 2015;48(6240):1255642.
Hvas M, Oppedal F. Sustained swimming capacity of Atlantic salmon. Aquac Environ Interact. 2017;9:361–9.
Hvas M, Folkedal O, Imsland A, Oppedal F. The effect of thermal acclimation on aerobic scope and critical swimming speed in Atlantic salmon, Salmo salar. J Exp Biol. 2017;220(15):2757–64.
Hvas M, Karlsbakk E, Mæhle S, Wright DW, Oppedal F. The gill parasite Paramoeba perurans compromises aerobic scope, swimming capacity and ion balance in Atlantic salmon. Conserv Physiol. 2017;5(1):cox066.
Hvas M, Folkedal O, Solstorm D, Vågseth T, Fosse JO, Gansel LC, Oppedal F. Assessing swimming capacity and schooling behaviour in farmed Atlantic salmon Salmo salar with experimental push-cages. Aquaculture. 2017;473:423–9.
Hvas M, Folkedal O, Imsland A, Oppedal F. Metabolic rates, swimming capabilities, thermal niche and stress response of the lumpfish, Cyclopterus lumpus. Biol Open. 2018;7:036079.
Hvas M, Folkedal O, Oppedal F. Heart rate bio-loggers as welfare indicators in Atlantic salmon (Salmo salar) aquaculture. Aquaculture. 2020;529: 735630.
Hvas M, Folkedal O, Oppedal F. Heart rates of Atlantic salmon Salmo salar during a critical swim speed test and subsequent recovery. J Fish Biol. 2021;98(1):102–11.
Hvas M, Folkedal O, Oppedal F. What is the limit of sustained swimming in Atlantic salmon post smolts? Aquac Environ Interact. 2021;13:189–98.
Hvas M. Swimming energetics of Atlantic salmon in relation to extended fasting at different temperatures. Conserv Physiol. 2022;10(1):coac037.
Johansson D, Laursen F, Fernö A, Fosseidengen JE, Klebert P, Stien LH, Vågseth T, Oppedal F. The interaction between water currents and salmon swimming behaviour in sea cages. PLoS ONE. 2014;9(5): e97635.
Kawabe R, Kawano T, Nakano N, Yamashita N, Hiraishi T, Naito Y. Simultaneous measurement of swimming speed and tail beat activity of free-swimming rainbow trout Oncorhynchus mykiss using an acceleration data-logger. Fish Sci. 2003;69(5):959–65.
Kawabe R, Naito Y, Sato K, Miyashita K, Yamashita N. Direct measurement of the swimming speed, tailbeat, and body angle of Japanese flounder (Paralichthys olivaceus). ICES J Mar Sci. 2004;61(7):1080–7.
Killen SS, Costa I, Brown JA, Gamperl AK. Little left in the tank: metabolic scaling in marine teleosts and its implications for aerobic scope. Proc R Soc B. 2007;274:431–8.
Lefevre S. Are global warming and ocean acidification conspiring against marine ecthotherms? A meta-analysis of the respiratory effects of elevated temperature, high CO2 and their interaction. Conserv Physiol. 2016;4:cow009.
Lucas MC, Baras E. Methods for studying spatial behaviour of freshwater fishes in the natural environment. Fish Fish. 2000;1:283–316.
Macaulay G, Warren-Myers F, Barrett LT, Oppedal F, Føre M, Dempster T. Tag use to monitor fish behaviour in aquaculture: a review of benefits, problems and solutions. Rev Aquac. 2021;13(3):1565–82.
Martínez M, Guderley H, Dutil J-D, Winger PD, Walsh SJ. Condition, prolonged swimming performance and muscle metabolic capacities of cod (Gadus morhua). J Exp Biol. 2003;206:503–11.
Metcalfe JD, Le Quesne WJ, Cheung WW, Righton DA. Conservation physiology for applied management of marine fish: an overview with perspectives on the role and value of telemetry. Philos Trans R Soc London Ser B. 2012;367:1746–56.
Morgan IJ, McDonald DG, Wood CM. The cost of living for freshwater fish in a warmer, more polluted world. Glob Chang Biol. 2001;7:345–55.
Norin T, Clark TD. Measurement and relevance of maximum metabolic rate in fishes. J Fish Biol. 2016;88:122–51.
Nilsson J, Folkedal O, Fosseidengen JE, Stien LH, Oppedal F. PIT tagged individual Atlantic salmon registered at static depth positions in a sea cage: vertical size stratification and implications for fish sampling. Aquacult Eng. 2013;55:32–6.
Oldham T, Nowak B, Hvas M, Oppedal F. Metabolic and functional impacts of hypoxia vary with size in Atlantic salmon. Comp Biochem Physiol A: Mol Integr Physiol. 2019;231:30–8.
Oppedal F, Dempster T, Stien LH. Environmental drivers of Atlantic salmon behaviour in sea-cages: a review. Aquaculture. 2011;311:1–18.
Remen M, Solstorm F, Bui S, Klebert P, Vågseth T, Solstorm D, Hvas M, Oppedal F. Critical swimming speed in groups of Atlantic salmon Salmo salar. Aquac Environ Interact. 2016;8:659–64.
Rechisky EL, Welch DW, Porter AD, Jacobs-Scott MC, Winchell PM. Influence of multiple dam passage on survival of juvenile Chinook salmon in the Columbia River estuary and coastal ocean. Proc Natl Acad Sci USA. 2013;110:6883–8.
Rikardsen AH, Righton D, Strøm JF, et al. Redefining the oceanic distribution of Atlantic salmon. Sci Rep. 2021;11:12266.
Rutz C, Hays GC. New frontiers in biologging science. Biol Lett. 2009;5(3):289–92.
Sievers M, Korsøen Ø, Warren-Myers F, Oppedal F, Macaulay G, Folkedal O, Dempster T. Submerged cage aquaculture of marine fish: a review of the biological challenges and opportunities. Rev Aquac. 2022;14(1):106–19.
Stehfest KM, Carter CG, McAllister JD, Ross JD, Semmens JM. Response of Atlantic salmon Salmo salar to temperature and dissolved oxygen extremes established using animal borne environmental sensors. Sci Rep. 2017;7:4545.
Steinhausen MF, Steffensen JF, Andersen NG. Tail beat frequency as a predictor of swimming speed and oxygen consumption of saithe (Pollachius virens) and whiting (Merlangius merlangus) during forced swimming. Mar Biol. 2005;148(1):197–204.
Stevens ED, Randall DJ. Changes in blood pressure, heart rate and breathing rate during moderate swimming activity in rainbow trout. J Exp Biol. 1967;46:307–15.
Svendsen E, Føre M, Økland F, Gräns A, Hedger RD, Alfredsen JA, Uglem I, Rosten CM, Frank K, Erikson U, Finstad B. Heart rate and swimming activity as stress indicators for Atlantic salmon (Salmo salar). Aquaculture. 2021;531:735804.
Thorarensen H, Gallaugher PE, Farrell AP. The limitations of heart rate as a predictor of metabolic rate in fish. J Fish Biol. 1996;49:226–36.
Timmerhaus G, Lazado CC, Cabillon NAR, Reiten BKM, Johansen LH. The optimum velocity for Atlantic salmon post-smolts in RAS is a compromise between muscle growth and fish welfare. Aquaculture. 2021;532: 736076.
Van Walsum TA, Perna A, Bishop CM, Murn CP, Collins PM, Wilson RP, Halsey LG. Exploring the relationship between flapping behaviour and accelerometer signal during ascending flight, and a new approach to calibration. International journal of avian science. 2020;162(1):13–26.
Videler JJ. Fish swimming. 1st ed. London: Chapman & Hall; 1993.
Wagner GN, McKinley RS, Bjørn PA, Finstad B. Physiological impact of sea lice on swimming performance of Atlantic salmon. J Fish Biol. 2003;62:1000–9. https://doi.org/10.1046/j.10958649.2003.00091.x.
Wagner GN, Kuchel LJ, Lotto A, Patterson DA, Shrimpton JM, Hinch SG, et al. Routine and active metabolic rates of migrating adult wild sockeye salmon (Oncorhynchus nerka Walbaum) in seawater and freshwater. Physiol Biochem Zool. 2006;79:100–8. https://doi.org/10.1086/498186.
Warren-Myers F, Hvas M, Vågseth T, Dempster T, Oppedal F. Sentinels in Salmon aquaculture: heart rates across seasons and during crowding events. Front Physiol. 2021. https://doi.org/10.3389/fphys.2021.755659.
Webb PW. Hydrodynamics and energetics of fish propulsion. Bull Fish Res Board Can. 1975;190:1–158.
Weihs D. Optimal cruising speed for migrating fish. Nature. 1973;245:48–50.
Wilson RW, Egginton S. Assessment of maximum sustainable swimming performance in rainbow trout (Oncorhynchus mykiss). J Exp Biol. 1994;192:299–305.
Wilson SM, Hinch SG, Eliason EJ, Farrell AP, Cooke SJ. Calibrating acoustic acceleration transmitters for estimating energy use by wild adult Pacific salmon. Comp Biochem Phys A. 2013;164(3):491–8.
Wright DW, Stien LH, Dempster T, Oppedal F. Differential effects of internal tagging depending on depth treatment in Atlantic salmon: a cautionary tale for aquatic animal tag use. Curr Zool. 2019;65:665–73.
Wright S, Metcalfe JD, Hetherington S, Wilson R. Estimating activity-specific energy expenditure in a teleost fish, using accelerometer loggers. Mar Ecol Prog Ser. 2014;496:19–32.
Zrini ZA, Gamperl AK. Validating Star-Oddi heart rate and acceleration data storage tags for use in Atlantic salmon (Salmo salar). Anim Biotelemetry. 2021;9:12.
Acknowledgements
The technical staff at the Matre Research station for providing excellent animal husbandry.
Funding
This research was funded through the Research Council of Norway, EXPOSED Aquaculture Research Centre, grant number 237790. Martin Føre acknowledges the financial support from Salmar ASA/Salmar Aker Ocean AS.
Author information
Authors and Affiliations
Contributions
This study was conceived and designed by all authors. Experimental trials were performed by MH, analyses of tag data were done by ES and MF. First draft was written by FW-M with all co-authors providing thorough feedback before approving the final version. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The use of animals was approved by the Norwegian Food Safety Authorities under ethics permit identification number 24444.
Consent for publication
All authors have agreed to publish this work as presented here.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Warren-Myers, F., Svendsen, E., Føre, M. et al. Novel tag-based method for measuring tailbeat frequency and variations in amplitude in fish. Anim Biotelemetry 11, 12 (2023). https://doi.org/10.1186/s40317-023-00324-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40317-023-00324-3