
Abstract
Background
Studies on the spatial ecology of invasive species provide critical information for conservation managers such as habitat preferences and identification of native species at risk of predation. To understand the spatial ecology of non-native Burmese pythons (Python molurus bivittatus), now well-established in Everglades National Park and much of South Florida USA, we radio-tracked 19 wild-caught adult pythons, 16 with VHF tags during 2006 through 2009 and 3 by GPS tags between 2010 and 2011. Our goal was to identify individual core-use areas and quantify home ranges, as well as to explore correlations of python movements with environmental parameters such as the presence of surface water.
Results
Radio-tracking periods ranged from 87 to 697 days for snakes with VHF tags, with a total of 5,119 tracking days (mean ± 1 SD = 319.9 ± 184.3 days); GPS tracking periods ranged from 12 to 93 days, with a total of 146 tracking days (mean ± 1 SD = 48.7 ± 40.7 days). We observed mean individual radio-tracked python home ranges of 22.5 km2 (2250 ha) with overall low site fidelity; all home ranges were within the park boundary. Python core-use areas included slough and coastal habitat types, and we delineated 18 common-use areas (that is, areas where individual core-use areas spatially overlapped). Tree islands were a principal feature of common-use areas, even if they were not the predominant habitat type. Multiple common-use areas were in proximity to roads. The longest movements of individual pythons correlated well with presence of surface water, and occurred during both wet and dry seasons.
Conclusions
High-use areas determined from python habitat-use and movement data may be optimal locations for targeted control efforts and further studies on impacts to native fauna.
Similar content being viewed by others

Background
Understanding habitat-use patterns is a fundamental aspect of animal ecology. Discerning the scope of these patterns can aid in designing appropriately timed and scaled management strategies for habitats of concern, including threatened, endangered, and invasive species. Competition with and predation by invasive species are continuing threats to native biodiversity worldwide [1-3], and invasive species can also cause harm to other ecological resources [4]. A comprehensive understanding of these threats requires information on movement and habitat-use patterns of invasives, or ‘exotics’ , in their new environments. It is estimated that nearly half of the imperiled species in the US are threatened by exotic species [2], possibly because a growing number of exotics are successfully invading and establishing viable populations.
The US state of Florida is home to a large variety of endangered species taxa [5] and more exotic animals than any other state. Snakes in particular have been shown to pose particularly high risks as potentially invasive species [6,7], and the recent establishment of Burmese pythons (Python molorus bivittatus) in south Florida poses a major threat to both the sensitive Everglades ecosystem and native species of conservation concern [8]. Dorcas et al. [9] recently described severe declines in mammal populations within the Everglades; the temporal and spatial patterns of python proliferation indicate that Burmese pythons may be one major cause of these declines.
Burmese pythons are long-lived, large-bodied constricting snakes native to Southeast Asia. Habitat and dietary generalists, these ambush predators can reach lengths greater than 6 m [10,11] and produce large clutches of eggs (8 to 107 eggs [12,13]). Despite considerable upkeep requirements to maintain and sustain giant snakes in captivity, Burmese pythons are often imported into the US where they are bred and sold as exotic pets. Although the route of entry into the wild is unclear, pythons were first observed in South Florida’s Everglades National Park (ENP) in 1979, and although not recognized as established until the year 2000 [14], the snakes were likely established by at least the mid 1980s [15]. Pythons have spread throughout ENP [16,17], and impacts from pythons on some populations of native fauna in South Florida appear to be significant [9].
Despite likely being established in ENP for several decades [18], their impacts are not well understood. A single report of a translocated, radio-tagged python from their native range is available [19]; results from that study indicated a home range of 12.3 ha over a 24-day period. However, previous ecological studies on other species of snakes, including pythons, have shown that movements and habitat use can vary in relation to a number of factors such as season [20-24], sex [25,26], body size [24,27], and prey [28]. Additionally, studies focusing on large constrictors have found seasonal shifts in habitat use [29-31] and movements related to changes in weather [32]; the home range size of pythons may therefore differ in new versus native habitats. Research on large constrictors in Australia has also shown that radio-tagged snakes displayed high site fidelity, even after translocation [33]. Thus, it is reasonable to expect that pythons may display site fidelity to particularly suitable habitats, yet it is not known what those habitats may be in South Florida.
In recent years, increases in the numbers of pythons removed from ENP [34] have prompted investigations into the impacts of pythons on native prey populations [9,35], possible establishment scenarios [15], risk assessments [36], and projections of potential habitat invasion in the southeastern US. [37-40]. Additionally, because little was known about the biology of this non-native species, researchers have tested the tolerance of wild-caught P. m. bivittatus to cold winter temperatures outside of Florida, where none of the snakes survived [18] and to estuarine and marine salinities in a laboratory setting, where hatchling pythons were able to sustain prolonged periods in saline water [41]. A severe winter cold snap in south Florida showed that adult pythons had limited ability to withstand exposure to prolonged cold, wet weather [42]. However, despite progress in understanding the biology of this invasive species in the Everglades, python movements, home range, and general habitat-use patterns are largely unknown.
We designed a multi-year radio-tracking study of 16 wild-caught Burmese pythons to quantify their home ranges, characterize movement patterns across the landscape, and classify habitat use by area over time. We tested the hypothesis that pythons displayed site fidelity to the location of their initial capture. Over a shorter time period, we also tested the ability of custom GPS tags to provide multiple high-quality daily locations for an additional three pythons. Our results have implications for designing appropriately-scaled management strategies to control pythons and for understanding the potential impact of this exotic reptile on native species, many of which are of high conservation concern.
Results
Radio- and GPS tracking
We successfully radio-tracked 16 pythons (12 female, 4 male); individuals ranged in size from 263.0 to 472.0 cm total length (TL), with a mean size of 352.7 cm (SD 65.4 cm; Table 1). Radio-tracking periods ranged from 87 to 697 days, with a total of 5,119 python days across all pythons and a mean of 319.9 days (SD = 184.3 days; Table 1). For those days on which we obtained locations for radio-tracked pythons from both aerial and ground-based telemetry (n = 33 days), the straight-line distance between aerial and walk-in locations was on average 160.9 m (SD = 158.1 m); this distance is representative for eight pythons (11, 12, 14, 17, 18, 23, 25, and 27) and was calculated to estimate accuracy of aerial locations, not for use in analysis.
Three additional female pythons were also GPS-tracked; these individuals ranged in size from 466.0 to 537.0 cm TL, with a mean size of 503.7 cm (SD 35.7 cm; Table 1). GPS tracking periods ranged from 13 to 93 days, with a total of 146 days and a mean of 48.7 days (SD = 40.7 days). We recorded a total of 679 GPS locations for all three snakes, with the number of locations/day ranging from 0 to 21, with a mean of 4.7/day (SD 3.9/day; Table 1). Python locations were recorded during both daytime (391 points; 58%) and nighttime (288 points; 42%) periods.
Radio-tracked python movements
On average, pythons moved a mean of 31.2 km from the initial capture location to the final locations recorded, with a range of 6.8 to 80.1 km; average daily distances moved were 0.04 to 0.18 km (Table 1). Maximum values for mean daily distances traveled ranged from 0.11 to 1.40 km (Table 1), and these movements occurred during both wet and dry seasons. Four pythons moved their greatest mean daily distances in December, three in October, three in June, and two in May; other pythons moved their greatest mean daily distance in July, August, September, and November (Table 1).
Minimum convex polygon (MCP) home ranges for radio-tracked pythons varied in size from 1.7 to 87.4 km2, with a mean of 22.5 km2 (SD = 21.3 km2) or 2,250 ha (Table 2). The size of MCPs was not influenced by tracking duration or gender (t = 0.49, P = 0.630). The mean size of MCPs for females (n = 12) was 24.1 km2 (SD = 23.9 km2), whereas the mean size for males (n = 4) was 17.8 km2 (SD = 12.1 km2).
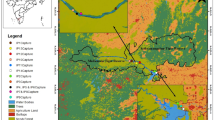
After overlaying all python MCPs to determine common-use areas, we identified 18 individual polygons of areas that were used by multiple radio- and GPS-tracked pythons (Figure 1, Table 3). A given common-use area was used by between two and five pythons, and most commonly (that is, mode) by two pythons. The mean size of common-use areas was 4.8 km2 (SD = 6.1 km2) and up to 22.4 km2.

Common MCP areas of radio-tracked Burmese pythons ( Python molurus bivittatus ) in Everglades National Park, South Florida, USA (a) and zoom-in map of python common-use areas (b).
Of 16 radio-tracked pythons, we had sufficient number of daily locations for 13 pythons to create kernel density estimates (KDEs), of which least squares cross-validation (LSCV) was minimized only for five pythons, likely due to mostly linear movements of pythons. Thus, since LSCV was not minimized, we only attempted KDEs for these five pythons. The KDE home ranges of these five pythons are widely spaced, and there was no common area (Figure 2). Mean home ranges (95% KDE) and core areas (50% KDEs) across all five pythons were 7.3 km2 (SD = 5.8 km2) and 1.5 km2 (SD = 1.4 km2), respectively, smaller than the MCP estimates.

Kernel density estimates for the five radio-tracked Burmese pythons ( Python molurus bivittatus ) in Everglades National Park, South Florida, USA.
MCP home ranges for pythons occurred primarily within slough habitat for 9/16 (56%) radio-tracked pythons and within coastal habitat for 6/16 (38%) pythons (Table 2). Mean distance of the centroid to the nearest road for each MCP common-use areas (n = 18) was 0.5 km (SD = 0.7 km; Table 3). Further, mean elevation of the common-use areas ranged from 17.0 to 50.3 cm (Table 3), and habitat types for common-use areas were predominantly slough and coastal, included tree islands, with some variation across the landscape (Table 4). Up to 12 tree islands were present in one common-use area, with individual tree island areas up to 0.4 km2 in size (Table 4). Radio-tracked pythons that crossed a road (n = 11) did so a 2 to 23 times, with a mean of 6.9 crossings (SD = 7.2 crossings) throughout their tracking periods.
Movements of some radio-tracked pythons were correlated to air temperature: movements of python 14 were significantly correlated with mean and min daily air temperature; python 23 movements were significantly correlated with min and max daily air temperatures; and movements of python 27 were significantly (negatively) correlated with all three factors (mean, min, and max daily air temperatures; Table 5). Moreover, for all radio-tracked pythons, mean daily distances moved were lower when water was present (75.4 m, n = 451, SD = 116.0 m) versus absent (143.6 m, n = 260, SD = 220.2 m; Satterthwaite two-tailed t-test; P < 0.0001, t = −5.74).
Site fidelity was tested across the entire tracking periods for all radio-tracked pythons, and only one male (python 15) showed site fidelity throughout the tracking duration (that is, movements were more constrained than a random path). However, we observed seasonal site fidelity: pythons 15, 16, and 17 displayed site fidelity during the dry season, whereas pythons 15, 17, and 22 displayed site fidelity during the wet season. There were too few observations for pythons 11 and 28 in the dry season and for pythons 14 and 31 during the wet season to assess seasonal site fidelity.
GPS-tracked python movements
On average, GPS-tracked pythons moved a mean of 7.2 km from initial capture location to final location, with a range of 1.0 to 18.4 km; average daily distances moved were 0.1 to 0.2 km (Table 1). Maximum values for mean daily distances traveled ranged from 0.2 to 2.4 km (Table 1). Pythons moved on average 32.2 m during daytime and 18.0 m during night time hours, with a range of (0 to 621.0 m) and (0 to 566.0 m), respectively. MCP home ranges varied in size from 0.1 to 23.1 km2, with a mean of 7.9 km2 (SD = 13.0 km2). Two pythons shared a common area with size of 372 m2. LSCV was minimized only for one python; the resulting KDE home range was 0.02 km2 (95% KDE) with a core area of 0.002 km2 (50% KDE).
GPS-tracked pythons were found in a mosaic of habitats during their tracking time, including upland forest pineland, slough, tree islands, coastal, lowland forest, disturbed, and marsh prairie habitats (Table 2). Within their home ranges, pythons were detected in slough and marsh prairie habitat types most frequently, although there was some use of coastal habitat, tree islands, and upland forest pinelands. Our MCP analysis showed that 1/3 (33%) of GPS-tracked pythons had >50% of their home ranges within slough habitat, whereas 2/3 (67%) of GPS-tracked pythons had >50% of their home range within marsh prairie habitat (Table 2). None of the GPS-tracked pythons crossed a road during their time in the field.
Discussion
Our study presents the first home range estimates for wild Burmese pythons in their introduced range. Home range size for Burmese pythons in ENP (MCP 22.5 km2 or 2,250 ha) were far greater than those reported in their native range, albeit for a single report from the Lantau Island, Hong Kong, showing a home range size of 12.3 ha over 24 days [19]. Our home range estimate was also much greater than that estimated for most other ambush snake predators that have previously been radio-tracked [43]. Large-bodied carpet pythons (Morelia spilota) had home ranges of 17.6 ha [30] and 22.5 ha [33], and diamond pythons (Morelia spilota spilota) had home ranges of 52 ha for males and 27 ha for females [29]. Further, daily movements of Burmese pythons in ENP (range 0.11 to 1.40 km) were larger than that reported for diamond pythons (100 m per week; [33]), and we observed only seasonal site fidelity patterns, indicating that most pythons did not consistently stay in a limited home range area throughout the year. ENP pythons have the ability to move relatively large distances and occupy relatively large home ranges, a capability that must be considered when population-spread scenarios are examined and risk assessments are performed. Further, [44] provided evidence that Burmese pythons are capable of homing long distances (21 to 36 km), in a relatively short period of time (94 to 296 days), after displacement at a scale previously undocumented for any snake species.
In addition to our python geo-referenced presence and absence data, this new home range information can help further our understanding of which prey species are at risk in ENP, including those that may be threatened, endangered, or rare. For example, in ENP, wood storks (Mycteria americana), Florida panthers (Puma concolor coryi), and Cape Sable seaside sparrows (Ammodramus maritimus mirabilis) are species of conservation concern that have home ranges proximal to the common-use areas of radio-tracked pythons [45-47]. Because pythons eat a variety of prey in ENP [48-50], it is possible that these species of conservation concern may be at risk of predation. Therefore, a logical next step is to determine the impact of pythons on native species near these core and common-use areas in the Everglades. Whether pythons can slow the pace of native population restoration for other top predators in ENP such as alligators and iconic wading birds is already a serious management concern, but empirical evidence is lacking.
Pythons displayed significant movements when surface water was absent, indicating dispersal may occur under locally ‘dry’ conditions. Perhaps Everglades’ pythons need to move greater distances from one relatively wet habitat patch to another to maintain access to adequate prey resources. Our evidence of site fidelity by season for some individuals during wet and dry seasons may support this hypothesis. Future fine-scale tracking coupled with very fine-scale activity loggers could complement and refine these initial radio-tracking results to illuminate dispersal strategies used by pythons in ENP.
Roads can serve as either a barrier or a corridor for wildlife [51], we found that 67% of radio-tracked pythons crossed a road with no record of road-caused mortality during the study period. Although home ranges for many snakes included roads, this may be biased because road-cruising was a capture method used in this study. Regardless, the ability of pythons to utilize roads with possibly relatively lower rates of mortality than other species (see [51]) further highlights the inconspicuous and cryptic nature of pythons in ENP, where overall detectability is low.
We found an overlap in MCP home ranges for individuals; however, it should be noted that based on how we acquired our study animals (that is, sometimes opportunistically obtaining pythons that were co-located with other radio-tagged pythons or close to roads), one might expect an overlap in some habitat-use areas. Overlapping home ranges may offer adult pythons increased encounter rates with other adults during the springtime mating period. Moreover, common-use areas may contain especially suitable habitat patches where factors such as prey availability and environmental conditions are favorable. In our study, the predominant habitat class within python home ranges was slough, and many pythons frequented areas with tree islands; tree islands are patches of woody vegetation within a freshwater wetland matrix dominated by non-woody species [52]. Thus, in addition to providing a hospitable shaded, structured habitat, tree islands are also utilized by many small mammals and birds. Some tree islands consist of rough terrain with nearly impenetrable vegetation, making surveys for secretive animals in such habitat extremely difficult. To date, no organized searches for pythons on tree islands in ENP have been initiated, but resource managers may need to give consideration to python search efforts in these habitats as data from radio-tagged pythons indicates that they spend long periods of time there. Our results suggest that evaluation of python habitat use on tree islands occurring in close proximity to python home range polygons is warranted.
Knowledge of the ‘detectability’ of an invasive species is important for planning control and eradication programs [53]. Mazzotti et al. [42] reported that pythons in South Florida were less detectable by humans in natural areas than in artificial habitats, potentially a result of the combination of habitat accessibility and python visibility. A field test of traps in the wild further revealed low overall capture rates and detections of pythons in surveyed areas [54]. Burmese pythons are extremely cryptic and well camouflaged in the Everglades, thus making them relatively difficult to detect for any given observer. In addition, the use of newer technology like our custom GPS tags can provide much more fine-scale location data for interpreting python habitat use and movements during day and night time hours.
Surveys and control measures such as trapping and direct, repeated visual searches in likely python habitats at specific times of the year (that is, winter) and day may increase removal rates of pythons. Currently, the extent that such removals actually reduce an apparently dense population of secretive snakes in a largely inaccessible landscape is unknown and could be low. Regardless, our results shed light on the area in which a given adult python’s daily activities are focused, and as such, inform planning of searches (that is, location and areal extent) intended to detect pythons in the wild.
Conclusions
The persistence of the non-native Burmese python population in the Everglades necessitates a range of scientific tools to understand the biology of this species in its new environment. Our results provide a new understanding of python movement rates, home range size, and habitat preferences. In addition, the use of GPS tracking technology can provide detailed daily movement data which we have not had to date, information that is useful for refining estimates of the impact these invaders are having on native prey populations. Sites we have identified as common- and core-use areas may be suitable for monitoring for python presence (for example, skin sheds) and targeted for python removal efforts. Moreover, these sites may be useful as study areas for detecting changes in species composition of prey resources available to pythons.
Methods
Ethics statement
All python radio-tracking was permitted under University of Florida animal care protocols F162 and 009-08-FTL, Florida Fish and Wildlife Conservation Commission permit ESC 08–02 and National Park Service (Everglades) permits EVER-2007-SCI-001 and EVER-2009-SCI-001.
Study site
We conducted python tracking from October 2007 through April 2012 within ENP (Figure 3). Everglades National Park is 0.9 million ha of wilderness bordered on the west and north by Big Cypress National Preserve and Tamiami Trail (US 41), extending into the mangrove-lined west coast of Florida and Florida Bay [12]. Habitat types in ENP are varied and include cypress swamps, hardwood hammocks, and wet prairies characterized by sawgrass (Cladium jamaicense). We conducted tracking in several locations throughout ENP, as well as in the remote coastal mangroves.

Study site where we conducted radio-tracking of Burmese pythons ( Python molurus bivittatus ) in Everglades National Park, South Florida, USA. Points represent initial capture locations for radio- and GPS-tracked pythons.
Python captures and implantation of radio and GPS transmitters
We caught all pythons (radio-tracked and GPS-tracked) by conducting road-cruising surveys of ENP roadways and levees during morning and evening hours. We obtained additional pythons by finding new individuals in proximity to already radio-tagged pythons. We hand-captured all pythons and temporarily restrained each individual in a snake bag within a large plastic storage container. We recorded capture locations using a handheld GPS unit. We measured each python, taking mass in kilograms and standard lengths (snout-vent length (SVL), total length (TL), and girth, in centimeters). Each python was implanted intraperitoneally (in the body cavity) with two radio transmitters [55-59]; the second transmitter was a ‘backup’ in the event that the first one failed. We used two transmitter sizes, Holohil models SI-2 (11 g) and AI-2 (25 g) (Holohil Systems, Ltd., Carp, ON, Canada). Three pythons received the additional GPS tag (Quantum 4000E Medium Backpack; Telemetry Solutions, Concord, CA, USA); transmitter weights did not exceed 0.1% of each snake’s body mass. The expected battery life for each VHF transmitter was between 9 and 12 months. GPS tags lasted between 2 and 5 months, depending on the frequency (number of times/day) with which the tags were programmed to attempt to acquire locations and how long each location took to complete. Tags were programmed to record locations every hour. Points from the tags were downloaded after the transmitter was recovered. We anesthetized each snake before implantation using isoflurane administered with an anesthesia machine; the surgery lasted between 30 to 60 min for each snake. After surgery, we observed each python for at least 24 h before release. Prior to release, we visually inspected each python to assess its health and alertness, as well as any problems related to implantation. None of the snakes used in our analyses showed any problems related to transmitter implantation. Snakes were tracked to the extent the battery life would allow, until they needed to be re-implanted with new transmitters or in the case of four snakes, until they were found deceased.
Radiotelemetry
We released each python within a few hundred meters of its original capture location (Figure 3). Once a week, we radio-located each radio-tracked python with a VHF tag from a fixed-wing aircraft (Cessna 182), flying at speeds between 100 and140 km/h and altitudes between 152 and 610 m. We used a Habit Osprey VHF telemetry receiver (Habit Research Ltd., Victoria, British Columbia, Canada) and H antennas mounted to the wings of the airplane or handheld from the ground to monitor telemetry signals. We recorded GPS locations for all pythons located during walk-ins; however, walk-in data was only used to confirm aerial locations and ground-based GPS locations, not in analysis of python movements or home range, as the number of visual confirmations was not consistent across all pythons. Pythons with GPS transmitters were also tracked aerially and visually located weekly. Locations were manually recorded not only on a handheld GPS by the observer during the flight as for all other pythons but also on-board the implanted GPS tag.
Statistical and spatial data analysis
Python movements
To visualize radio-tracked python movements over time, we plotted aerial telemetry locations for all pythons by date using a geographic information system [60] and created tracks by connecting successive points, assuming a straight-line movement. For a subset of radio-tracked pythons for which we had walk-in locations and aerial telemetry locations on the same day, we calculated the distance between aerial and ground locations to test the accuracy of aerial locations.
To calculate the mean daily and average mean daily distance traveled, we connected successive points, assuming a straight-line movement. We used aerial locations for radio-tracked pythons and chose one GPS location per day for each GPS-tracked snake. If there were multiple locations per day, we chose the point calculated using the maximum number of satellites (range 3 to 10). If there were multiple locations that shared a maximum number of satellites, we chose the point with highest maximum satellite signal strength (range 32 to 45) used to calculate the location. To separate out movements during day and night time hours, we calculated the distance between successive locations during those times. Since data collected between radio-tracked and GPS-tracked pythons were collected on different scales (weekly versus multiple per day), and with different technology, data for GPS pythons was analyzed separately.
Home ranges
For all radio-tracked pythons, we calculated the MCP (n = 16) estimates and, when possible (at least 20 daily locations), KDE (n = 5) from aerial tracking data to define home ranges. For GPS-tracked pythons, we used all GPS locations to calculate MCPs (n = 3) and KDEs (n = 1).
Home range metrics, by definition, describe the area traversed by an animal during normal daily activities, excluding migrations or erratic movements [61]. Estimates of MCP identify home ranges as the area within a polygon formed by joining outermost re-sighting positions of an animal [62-65]; MCP has been commonly applied in other snake tracking studies [28,29,31]. In addition to calculating MCPs, we assessed the overlap of individual python MCPs to determine ‘hotspots’ or common-use areas and calculated the centroid for each MCP. Similarly, kernel density is a non-parametric method used to identify one or more areas of disproportionately heavy use (that is, core areas) within a home-range boundary [66,67], with appropriate weighting of outlying observations. As [67] suggested, for KDEs, we used the 95% contour to represent home ranges and the 50% to depict ‘core’ areas for each animal following [68,69]. We used the Home Range Tools extension [70] with ArcGIS 9.3 [60] and a fixed kernel LSCV smoothing factor (h cv ; [68,71]) for each individual KDE; we estimated KDEs only for pythons that had ≥20 mean daily locations.
Habitat use and environmental variables
We overlaid the ENP boundary on all resulting maps of python home ranges and core-use areas. We reclassified a land cover and land use map [72], using a modified Davis classification for habitat types [73]; modifications included compiling similar habitat types into fewer, broader categories of habitat classes. The reclassified map includes upland forest/pineland, slough, tree islands, coastal, lowland forest, disturbed, marsh prairie, and other habitat types. Additionally, because many previous python observations have been logged near roads (NPS and USGS, unpublished data), which can act as barriers to other animal movements [51], we determined the number of individual python movement paths that crossed a road.
To assess how python movements differed seasonally, we calculated movement distances across seasons; we defined seasons in ENP as ‘wet’ during May through October and ‘dry’ during November through April [74]. To explore whether radio-tracked python movements were correlated with environmental variables, we assessed Pearson’s correlation coefficients with mean, min, and max air temperature recorded at a local weather station (that is, JBTS owned by South Florida Water Management District), downloaded from their DBHYDRO database [75]. We also assessed movements in relation to water presence/absence and elevation using a spatially explicit daily surface water depth model developed by Everglades Depth Estimation Network [76,77]. We conducted all statistical tests in R [78] and used an α level of 0.05 for all analysis to assess statistical significance.
Site fidelity
Finally, to test the hypothesis that radio-tagged pythons displayed high site fidelity during their entire tracking periods or by season (wet and dry), we conducted Monte Carlo random walk simulations using Animal Movement Analysis Extension for ArcView 3.2 [79]; we compared observed movement paths with 1,000 randomly generated paths for all radio-tracked animal for both entire tracking periods and by season.
References
Flather CH, Knowles MS, Kendall IA. Threatened and endangered species geography. Bioscience. 1998;48:365–76.
Wilcove DS, Rothstein D, Dubow J, Phillips A, Losos E. Quantifying threats to imperiled species in the United States. Bioscience. 1998;48:607–15.
Gurevitch J, Padilla DK. Are invasive species a major cause of extinctions? Trends Ecol Evol. 2004;19:470–4.
Pimentel D, Zuniga R, Morrison D. Update on the environmental and economic costs associated with alien invasive species in the United States. Ecol Econ. 2005;52:273–88.
Dobson AP, Rodriguez JP, Roberts WM, Wilcove DS. Geographic distribution of endangered species in the United States. Science. 1997;275:550–3.
Reed RN. An ecological risk assessment of nonnative boas and pythons as potentially invasive species in the United States. Risk Anal. 2005;25:753–66.
Kraus FR. Alien reptiles and amphibians: a scientific compendium and analysis. New York: Springer; 2009.
Reed RN, Rodda GH. Giant constrictors: biological management profiles and an establishment risk assessment for nine large species of python, anacondas, and the boa constrictor. Open-file report 2009–1202. Reston, VA: U.S Geological Survey, Department of the Interior; 2009.
Dorcas ME, Willson JD, Reed RN, Snow RW, Rochford ME, Miller MA, et al. Severe mammal declines coincide with proliferation of invasive Burmese pythons in Everglades National Park. Proc Natl Acad Sci U S A. 2012;109:2418–22.
Wall F. Ophidia taprobanica or the snakes of Ceylon. Columbo: Govt. printer; 1921.
Pope CH. The giant snakes. New York: Knopf; 1961.
Lederer G. Fortpflanzungsbiologie unduntwicklung von Python molurus molurus (Linne) und Python molurus bivittatus (Kuhl). Die Aquarien- Und Terrarien Zeitschrift. 1956;9:243–8.
Branch WR, Erasmus H. Captive breeding of pythons in South Africa, including details of an interspecific hybrid (Python sebae natalensis x Python molurus bivittatus). J Herp As Afr. 1984;1984:1–10.
Meshaka Jr WE, Loftus W, Steiner T. The herpetofauna of Everglades National Park. Florida Sci. 2000;63:84–103.
Willson JD, Dorcas ME, Snow RW. Identifying plausible scenarios for the establishment of invasive Burmese pythons (Python molurus) in southern Florida. Biol Invasions. 2011;13:1493–504.
Snow RW, Johnson VM, Brien ML, Cherkiss MS, Mazzotti FJ. Python molurus bivittatus: nesting. Herp Rev. 2007;38:93.
Snow RW, Krysko KL, Enge KM, Oberhofer L, Warren-Bradley A, Wilkins L. Introduced populations of Boa constrictor (Boidae) and Python molurus bivittatus (Pythonidae) in southern Florida. In: Henderson RW, Powell R, editors. The biology of boas and pythons. Utah: Eagle Mountain Publishing; 2007. p. 416–38.
Dorcas ME, Willson JD, Gibbons JW. Can invasive Burmese pythons inhabit temperate regions of the southeastern United States? Biol Invasions. 2011;13:793–802.
Goodyear NC. Python molurus bivittatus (Burmese python): movements. Herp Rev. 1994;25:71–2.
Diffendorfer JE, Rochester C, Fisher RN, Brown TK. Movement and space use by Coastal Rosy Boas (Lichanura trivirgata roseofusca) in Coastal Southern California. J Herpetol. 2005;39:24–36.
Bell SLM, Herman TB, Wassersug RJ. Ecology of Thamnophis sauritis (Eastern Ribbon Snake) at the northern limit of its range. Northeast Nat. 2007;14:279–92.
Sperry JH, Taylor CA. Habitat use and seasonal activity patterns of the Great Plains ratsnake (Elaphe guttata emoryi) in central Texas. Southwest Nat. 2008;53:444–9.
Wisler C, Hofer U, Arlettaz R. Snakes and monocultures: habitat selection and movements of female grass snakes (Natrix natrix L.) in an agricultural landscape. J Herpetol. 2008;42:337–46.
Pizzatto L, Madsen T, Brown GP, Shine R. Spatial ecology of hatchling water pythons (Liasis fuscus) in tropical Australia. J Trop Ecol. 2009;25:181–91.
Moore JA, Gillingham JC. Spatial ecology and multi-scale habitat selection by a threatened rattlesnake: the Eastern Massasauga (Sistrurus catenatus catenatus). Copeia. 2006;4:742–51.
Kapfer JM, Coggins JR, Hay R. Spatial ecology and habitat selection of bullsnakes (Pituophis catenifer sayi) at the northern periphery of their geographic range. Copeia. 2008;4:815–26.
Roth TC, Greene BD. Movement patterns and home range use of the northern watersnake (Nerodia sipedon). Copeia. 2006;3:544–51.
Madsen T, Shine R. Seasonal migration of predators and prey: pythons and rats in tropical Australia. Ecology. 1996;77:149–56.
Slip DJ, Shine R. Habitat use, movements, and activity patterns of free-ranging diamond pythons, Morelia spilota spilota (Serpentes: Boidae): a radiotelemetric study. Aust Wildlife Res. 1988;15:515–31.
Shine R, Fitzgerald M. Large snakes in a mosaic rural landscape: the ecology of carpet pythons Morelia spilota (Serpentes: Pythonidae) in coastal eastern Australia. Biol Cons. 1996;76:113–22.
Heard GW, Black D, Robertson P. Habitat use by the inland carpet python (Morelia spilota metcalfei: Pythonidae): seasonal relationships with habitat structure and prey distribution in a rural landscape. Austral Ecol. 2004;29:446–60.
Brown GP, Shine R, Madsen T. Spatial ecology of slatey-grey snakes (Stegonotus cucullatus, Colubridae) on a tropical Australian floodplain. J Trop Ecol. 2005;21:605–12.
Pearson D, Shine R, Williams A. Spatial ecology of a threatened python (Morelia spilota imbricata) and the effects of anthropogenic habitat change. Austral Ecol. 2005;30:261–74.
National Park Service [http://www.nps.gov/ever/naturescience/burmesepythonremoval.htm]
Holbrook J, Chesnes T. An effect of Burmese pythons (Python molurus bivittatus) on mammal populations in southern Florida. Florida Sci. 2011;74:17–24.
Fujisaki I, Hart KM, Mazzotti FJ, Rice KG, Snow RW, Rochford MR. Risk assessment of potential invasiveness of exotic reptiles imported to South Florida. Biol Invasions. 2009;12:2585–96.
Pyron RA, Burbrink FT, Guiher TJ. Claims of potential expansion throughout the U.S. by invasive python species are contradicted by ecological niche models. PLoS One. 2008;3:e2931.
Rodda GH, Jarnevich CS, Reed RN. What parts of the US mainland are climatically suitable for invasive alien pythons spreading from Everglades National Park? Biol Invasions. 2009;11:241–52.
Van Wilgen NJ, Roura-Pascual N, Richardson DM. A quantitative climate-match score for risk-assessment screening of reptile and amphibian introductions. Environ Manage. 2009;44:590–607.
Rodda GH, Jarnevich CS, Reed RN. Challenges in identifying sites climatically matched to the native ranges of animal invaders. PLoS One. 2011;6:e14670.
Hart KM, Schofield PJ, Gregoire DR. Experimentally derived salinity tolerance of hatchling Burmese pythons (Python molurus bivittatus) from the Everglades, Florida (USA). J Exp Mar Biol Ecol. 2012;413:56–9.
Mazzotti FJ, Cherkiss MS, Hart KM, Snow RW, Rochford MR, Dorcas ME, et al. Cold-induced mortality of invasive Burmese Pythons in south Florida. Biol Invasions. 2010;13:143–51.
Macartney JM, Gregory PT, Larsen KW. A tabular survey of data on movements and home ranges of snakes. J Herpetol. 1988;22:62–73.
Pittman SE, Hart KM, Cherkiss MS, Snow RW, Fujisaki I, Smith BJ, et al. Homing of invasive Burmese pythons is South Florida: evidence for map and compass senses in snakes. Biol Lett. 2014;10:20140040.
Cox JJ, Maehr DS, Larkin JL. Florida panther habitat use: new approach to an old problem. J Wildl Manage. 2006;70:1778–85.
Cassey P, Lockwood JL, Fenn KH. Using long-term occupancy information to inform the management of Cape Sable seaside sparrows in the Everglades. Biol Cons. 2007;139:139–49.
Frederick PC, Meyer KD. Longevity and size of wood stork (Mycteria americana) colonies in Florida as guides for an effective monitoring strategy in the Southeastern United States. Waterbirds. 2008;31:12–8.
Snow RW, Brien ML, Cherkiss MS, Wilkins L, Mazzotti FJ. Dietary habits of the Burmese python, Python molurus bivittatus, from Everglades National Park. Florida Herp Bull. 2007;101:5–7.
Rochford MR, Krysko K, Nifong J, Wilkins L, Snow RW, Cherkiss MS. Python molurus bivittatus (Burmese python). Diet Herp Review. 2010;41:97.
Dove CJ, Snow RW, Rochford MR, Mazzotti FJ. Birds consumed by the invasive Burmese python (Python molurus bivittatus) in Everglades National Park, Florida. USA Wilson J Ornithol. 2011;123:126–31.
Shepard DB, Kuhns AR, Dreslik MJ, Phillips CA. Roads as barriers to animal movement in fragmented landscapes. Anim Conserv. 2008;11:288–96.
Wetzel PR, Sklar FH, Coronado CA, Troxler TG, Krupa SL, Sullivan PL, et al. Biogeochemical processes on tree islands in the greater everglades: initiating a new paradigm. Crit Rev Env Sci Tec. 2011;41:670–701.
Christy MT, Adams AAY, Rodda GH, Savidge JA, Tyrrell CL. Modelling detection probabilities to evaluate management and control tools for an invasive species. J Appl Ecol. 2010;47:106–13.
Reed RN, Hart KM, Rodda GH, Mazzotti FJ, Snow RW, Cherkiss MS, et al. A field test of attractant traps for invasive Burmese pythons (Python molurus bivittatus) in southern Florida. Wildlife Res. 2011;38:114–21.
Reinert HK, Cundall D. An improved surgical implantation method for radio-tracking snakes. Copeia. 1982;3:702–5.
Weatherhead PJ, Anderka FW. An improved radio transmitter and implantation technique for snakes. J Herpetol. 1984;18:264–9.
Hardy Sr DL, Greene HW. Surgery on rattlesnakes in the field for implantation of transmitters. Sonoran Herp. 1999;12:25–7.
Hardy Sr DL, Greene HW. Inhalation anesthesia of rattlesnakes in the field for processing and transmitter implantation. Sonoran Herp. 2000;13:110–4.
Dorcas ME, Willson JD. Innovative methods for studies of snake ecology and conservation. In: Mullin SJ, Seigel RA, editors. Snakes ecology and conservation. Ithaca: Comstock; 2009. p. 5–37.
Environmental Systems Research Institute (ESRI). ArcGIS 9.3. Redlands, CA: Geographic Information Systens (GIS); 2007.
Bailey JA. Principles of wildlife management. New York: Wiley-Liss; 1984.
Burt WH. Territoriality and home rage concepts as applied to mammals. J Mammal. 1943;24:346–52.
Jennrich RI, Turner FB. Measurement of non-circular home range. J Theoret Biol. 1969;22:227–37.
Worton BJ. A review of models of home range for animal movement. Ecol Model. 1987;38:277–98.
White GC, Garrott RA. Analysis of wildlife radiotracking data. New York: Academic; 1990.
Worton BJ. Kernal methods for estimating the utilization distribution in home range studies. Ecology. 1989;70:164–8.
Laver PN, Kelly MJ. A critical review of home range studies. J Wildl Manage. 2008;72:290–8.
Seaman DE, Powell RA. Identifying patterns and intensity of home range use. In A Selection of Papers from the Eighth International Conference on Bear Research and Management: February 1989; Victoria, British Columbia, Canada. International Association for Bear Research and Management; 1990:243–249.
Powell RA. Animal home ranges and territories. In: Boitani L, Fuller TK, editors. Research Techniques in animal ecology: controversies and consequences. New York: Columbia University Press; 2000. p. 65–110.
Rodgers AR, Carr AP, Smith L, Kie JG. HRT: home range tools for ArcGIS. Ontario, Canada: Ministry of Natural Resources, Centre for Northern Forest Ecosystems Research; 2005.
Worton BJ. Using Monte Carlo simulation to evaluate kernel-based home range estimators. J Wildl Manage. 1995;59:794–800.
South Florida Water Management District GIS Data Catalogue [http://www.sfwmd.gov/gisapps/sfwmdxwebdc/dataview.asp?]
Davis Jr JH. The natural features of southern Florida, especially the vegetation, and the Everglades. FL Dept Cons Geol Bull. 1943;25:311.
Myers RL, Ewel JJ. Ecosystems of Florida. Orlando, FL: University of Central Florida; 1990.
South Florida Water Management District DBHYDRO [http://my.sfwmd.gov/dbhydroplsql/show_dbkey_info.main_menu]
EDEN Everglades Depth Estimation Network [http://sofia.usgs.gov/eden]
Telis PA. The Everglades Depth Estimation Network (EDEN) for support of ecological and biological assessments: US Geological Survey Fact Sheet 2006–3087. Jacksonville, FL: U.S. Geological Survey, U.S. Department of the Interior; 2006.
R Development Core Team. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2011.
Hooge PN, Eichenlaub W, Hooge ER. Animal movement 2.5. Anchorage, AK: U.S. Geological Survey, Alaska Biological Science Center; 2001.
Acknowledgements
We thank M. Rochford, A. Wolf, T. Walters, T. Kieckhefer, T. Hill, M. Brien, J. Carrigan, M. Parry, B. Jeffery, J. Beauchamp, and B. Greeves for the assistance in the field. We are grateful to Y. Escribano for the assistance with GIS. H. Crowell assisted with collating literature cited and A. Sartain provided helpful suggestions on an earlier draft of the manuscript. Funding was provided by the National Park Service and the USGS Priority Ecosystem Science Program. Support for M.E. Dorcas was provided by Davidson College. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
KH, MC, BS, and IF performed the analyses and drafted the manuscript. MC and BS also performed the data collection. FM, RS, and MD helped to draft the manuscript. All authors read and approved the final manuscript.
Kristen M Hart, Michael S Cherkiss, Brian J Smith, Ikuko Fujisaki, and Xiaowei Wu contributed equally to this work.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Hart, K.M., Cherkiss, M.S., Smith, B.J. et al. Home range, habitat use, and movement patterns of non-native Burmese pythons in Everglades National Park, Florida, USA. Anim Biotelemetry 3, 8 (2015). https://doi.org/10.1186/s40317-015-0022-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40317-015-0022-2