
Abstract
Background
Commercial diets are frequently formulated to meet or exceed nutrient levels including those of limiting essential amino acids (AA) covering potential individual variations within the herd. However, the provision of dietary excess of AA, such as Lys, may lead to reduced appetite and growth in pigs. The mechanisms modulating these responses have not been extensively investigated. This study evaluated the effect of Lys dietary excesses on performance and satiety biomarkers in post weaning pigs.
Methods
Twenty-four pigs aged 21 d and weighing 6.81 ± 0.12 kg (mean ± SEM) were individually housed and offered 1 of 4 dietary treatments for 3 weeks: a diet containing a standardized ileal digestible Lys reaching 100% (T0), 120% (T1), 150% (T2) or 200% (T3) of the NRC (2012) requirements. At the end of the experiment, blood samples from the cephalic vein of the T0 and T3 groups were obtained for AA analysis. In addition, primary intestinal cultures from T0 pigs were used, following their humane killing, to evaluate the effect of Lys on gut hormone secretion and AA sensors gene expression under ex vivo conditions.
Results
Feed intake was linearly reduced (P < 0.001) and the weight gain to feed ratio reduced (P < 0.10) with increased dietary levels of Lys during the third- and first-week post weaning, respectively. Cholecystokinin concentration (P < 0.05) and the metabotropic glutamate receptor 1 and the solute carrier family 7 member 2 (P < 0.10) gene expression was enhanced in proximal jejunum tissues incubated with Lys at 20 mmol/L when compared to the control (Lys 0 mmol/L). Plasma Lys and Glu (P < 0.05) concentration increased in the T3 compared to T0 pigs. In contrast, plasma levels of His, Val, Thr, Leu (P < 0.05) and Gln (P < 0.10) were lower in T3 than T0 pigs.
Conclusion
The present results confirm that excess dietary Lys inhibits hunger in pigs. Moreover, the results provide evidence of pre- and post-absorptive mechanisms modulating these responses. Lys dietary excesses should be narrowed, when possible, to avoid negative effects of the AA on appetite in pigs.
Similar content being viewed by others

Background
Commercial pig feeds are frequently formulated to meet or exceed the requirements of key nutrients, such as limiting essential amino acids (AA). Nutritionists commonly apply margins above the minimum levels to account for potential practical issues (e.g., poor mixing) and potential individual variations on the nutritional requirements within the herd [1]. However, the excessive intake of some AA has been associated with reduced appetite in pigs [2]. Lys, the most limiting essential AA, can trigger satiation (feeling of fullness that leads to the termination of a meal) and satiety (feeling of fullness that prevents the initiation of a new meal) when provided in excess in pigs, such as through an oral gavage [3]. The negative effect of Lys on feed intake has been partly associated with the release of gut hormones with anorexigenic properties, such as cholecystokinin (CCK) [4]. While the release of CCK has been linked with the activation of enteroendocrine cells in the duodenum, previous studies in our group and others indicated that the effect of Lys on the release of CCK may occur mainly in the jejunum or other more distal segments of the gastrointestinal tract [2, 5]. In addition, the short half-life of CCK combined with the long-term effects of the AA on feed intake indicate that post absorptive mechanisms may also be involved in the satiating effect of Lys in pigs [3].
Following intestinal absorption into the portal vein and potential hepatic uptake, AA that reach the central nervous system (CNS) may function as appetite regulators, signalling the brain about the body metabolic status [6,7,8]. Reduced feed intake has been observed following the intake of dietary excesses of Leu in pigs [9]. The satiating effect triggered by the ingestion of high doses of Leu has been attributed for the most part to the detection of branched-chain amino acid (BCAA) imbalances in blood reaching the CNS [10, 11]. Changes in the circulating concentrations of essential amino acids (EAA), including BCAA, and non-essential amino acids (NEAA) have been noted following the ingestion of Lys excess diets in pigs [12, 13]. However, the blood kinetic of dietary Lys and the relationship with feed intake has not been extensively studied. A better understanding of the role of Lys on appetite modulation could help optimize feed formulations and enhance performance during the post weaning period, where low feed intake and body weight gain are frequently monitored in pigs.
The aim of the present study was to investigate the effect of dietary excess of Lys on performance and satiety related biomarkers including gut hormone secretion (CCK and glucagon-like peptide 1 (GLP-1)) and AA sensor gene expression in the gastrointestinal tract, as well as the blood kinetic of AA in post weaning pigs. It was hypothesized that dietary excess of Lys will reduce feed intake and increase the secretion of CCK in the proximal jejunum of post weaning pigs.
Materials and methods
Animals, housing, and diets
Twenty-four newly weaned (24 days of age) pigs (Landrace × Large White; body weight (mean ± SEM) = 6.81 ± 0.12 kg) were assigned to 4 dietary experimental groups with homogeneous body weight and balanced gender, and individually housed in slatted floor pens (1.7 m × 1.2 m) at the Herston Medical Research Centre at The University of Queensland (Herston Campus, Brisbane, Australia). Pigs were kept with 12 h of light (intensity of 40–60 lx) and the room temperature was maintained at 29 °C during the first week and then progressively decreased by 1 °C weekly. Pigs had ad libitum access to feed and water throughout the experiment. The 4 experimental diets were: a standard starter diet meeting 100% the requirements set by the NRC (1.20 standardised ileal digestible (SID) Lys; T0), or 3 dietary treatments with an excess SID Lys reaching 120% (T1), 150% (T2) or 200% (T3) of the NRC recommendations [14]. To differentiate the Lys effect on appetite from an increase in dietary nitrogen content, Ala (L-isoform, food grade) (Bulk Supplements, Henderson, USA) was supplemented to T0, T1 and T2 to make all diets isonitrogenous. Alanine was chosen over other potential AA based on its previously reported lack of impact on appetite and gut peptide secretion in pigs [2, 5]. The composition of experimental diets is shown in Table 1. Feed intake was measured daily and body weight weekly (d 7, 14 and 21).
Blood and tissue sample collection
On d 22, blood samples (2 mL) were collected from the right cephalic vein of T0 and T3 (control and highest Lys level, respectively) pigs. To facilitate the procedure (and minimize stress) pigs were moved to a separate room, anaesthetized with isoflurane (2%–3%) via mask ventilation and placed in the dorsal recumbency position on top of a surgical table with the right forelimb extended backward and slightly outward (to expose the cephalic vein). Upon collection, blood samples were immediately transferred into 2 mL EDTA vacutainers (Becton, Dickinson and Company, Franklin Lakes, USA) and placed on ice before further processing (i.e., centrifugation). Blood samples from the lowest and highest Lys treatments were chosen for collection to distinctly identify the AA blood profile related to the consumption of dietary Lys excesses in weaning pigs. Prior to blood collection, all animals had access to a morning meal for 30 min. All feeders were removed 1 h before blood sampling. The time of blood collection was chosen based on previous reports on AA blood kinetics in pigs [15].
Following the blood sampling, pigs were humanely killed using Lethabarb® (intravenous administration, 162.5 mg/kg) (Virbac, Milperra, Australia). The small intestine was then removed from the carcass, and the length measured from the pylorus to the ileocecal valve. Proximal jejunum (approx. 100–120 cm distal from the pylorus) and ileum (10 cm proximal from the ileocecal valve) from control pigs (T0 group) were immediately removed and placed in ice cold Krebs-Ringer Bicarbonate (KRB)/HEPES buffer adjusted to pH 7.4 bubbled with O2/CO2 (95%/5%) and promptly transported to the laboratory (The University of Queensland, St Lucia Campus, Brisbane, Australia) for the preparation of primary intestinal cell cultures and analysis of CCK and GLP-1 secretion. Magnesium chloride (0.0468 g/L), potassium chloride (0.34 g/L), sodium chloride (7 g/L), sodium phosphate dibasic (0.1 g/L), sodium phosphate monobasic (0.18 g/L), D-glucose (1.8 g/L) and HEPES (5.9 g/L) were used to prepare the KRB/HEPES buffer (all chemicals (reagent grade) were purchased from Sigma-Aldrich (Castle Hill, Australia)). The locations for sampling were selected based on previously published data on the small intestine morphometry of young pigs [16].
Primary intestinal cell culture
Proximal jejunum and ileum samples were collected for primary intestinal cultures following the procedure previously described by Voortman et al. [17] to study the effect of free Lys on gut peptide secretion and the expression of AA sensors. In brief, upon arrival at the laboratory, intestinal segments were cleaned of debris using KRB/HEPES buffer, cut open longitudinally and stripped of their outer muscle layers before equally sized round samples (1.13 cm2) were excised using a biopsy punch (Acuderm Inc., Fort Lauderdale, USA) and placed in 24-well plates (ThermoFisher Scientific, Waltham, USA) filled with ice-cold KRB/HEPES buffer. After 30 min at room temperature, plates were moved into a humidified incubator (NU5510, Nuaire, Plymouth, USA) for 1 h at 37 °C and 5% v/v CO2, after which the content of each well was replaced with a new solution of prewarmed KRB/HEPES buffer containing Lys (analytical grade) (Sigma-Aldrich) at 0, 10, 20 or 30 mmol/L and no D-glucose. Plates were placed back in the incubator for an additional 1 h and then the supernatant collected, mixed with phenylmethanesulfonyl fluoride at 100 mmol/L (to prevent enzymatic degradation of gut peptides) (Sigma-Aldrich) and stored in Eppendorf tubes at −80 °C for hormone analysis. Tissue samples were placed in RNAlater (Sigma-Aldrich) at room temperature for 24 h before frozen at −80 °C for later mRNA analysis. Lactate dehydrogenase (LDH) activity was measured with Roche LDH reagent kit PLUS (4744926001 Sigma-Aldrich) in tissue samples (by evaluating the leakage of intracellular LDH into supernatant stored at 4 °C) and compared to a positive control incubated with 1% Triton-X 100 (Sigma-Aldrich), to determine the viability of the intestinal culture.
Amino acid and hormone analysis
Blood samples were centrifuged (3,000 r/min) at 4 °C for 10 min within 1 h from collection and the plasma aliquoted into Eppendorf tubes before frozen at −80 °C until AA analysis. To prepare the plasma for the AA evaluation, samples were first centrifuged at 14,000 × g at room temperature for 30 min using Amicon® Ultra 0.5 mL filters (3 kDa MWCO) (Merck Millipore, Burlington, USA). The filtrate was then aliquoted into Eppendorf tubes and mixed, in the same proportion (1:1), with an internal standard solution containing 2-aminobutanoic acid and sarcosine. AA within the processed samples were then derivatised following the methodology described by Valgepea et al. [18], and the concentration analysed by using RP-HPLC. Supernatant samples were analysed for total CCK and GLP-1 using a Porcine Cholecystokinin ELISA kit from MyBioSource (MBS264395, San Diego, USA) and a glucagon-like peptide-1 (Total) ELISA kit from Merck Millipore (EZGLPT1-36 K, Burlington, USA), respectively. Intra- and inter-assay coefficients of variation for the CCK kit were 6.0% and 9.3%, respectively, whereas GLP-1 intra- and inter-assay coefficients of variation for the GLP-1 kit were 1.9% and 2.2%, respectively. Optical density recordings of the ELISA plates were performed in BMG LABTECH FLUOstar OPTIMA (BMG Labtech, Mornington, Australia).
RNA extraction and RT-qPCR analysis
The extraction and isolation of RNA from proximal jejunum was performed using first Trizol® Reagent (15596026) and PureLink® RNA Mini kit (12183018A) (Invitrogen, Carlsbad, USA), respectively, following the manufacturer’s instruction. Sample’s RNA quality and concentration were determined using Nano Drop spectrophotometer (NanoDrop 8000, ThermoFisher Scientific, Waltham, USA). QuantiTect® Reverse Transcription kit (205313, QIAGEN, Hilden, Germany) was used for cDNA synthesis. Primers for the AA sensors and the reference genes used in this study have been previously published [5, 19, 20] and are listed in Table 2. The real-time qPCR reaction contained 5 μL of SYBR Green master mix solution, 3 μL of RNAs free water, 1 μL of cDNA sample, 0.5 μL forward and reverse PCR primers and 0.05 μL of ROX reference dye solution. The RT-qPCR program used for the analysis has been previously published [2]. In short, following initial denaturation at 95 °C for 2 min, the PCR cyclin condition was 40 cycles of 95 °C for 15 s and 60 °C for 60 s using a real time PCR system (QuantStudio 6, ThermoFisher Scientific, Waltham, USA). The housekeeping gene GAPDH was used as an internal control to normalize the expression of target genes. The relative expression of genes was calculated following the Pfaffl method [21].
Statistical analysis
Statistical analysis was performed using R software (RStudio, Inc., Boston, Massachusetts, USA). All data are expressed as the mean ± SEM. Performance parameters [average daily feed intake (ADFI), average daily gain (ADG), and gain to feed ratio (G:F)] were analysed using a linear mixed model with “sex”, “weight”, “Lys dietary dose”, “week”, and the interaction of the last 2 as fixed effects, and “pig” as random effect. Plasma AA concentration from T0 and T3 pigs were analysed with a linear mixed model with “Lys dietary dose”, “sex” and “weight” as fixed effect and “pig” as random effect. Ex vivo data (gut hormone secretion and AA sensors gene expression) were analysed with a linear mixed model considering “Lys mmol/L treatment” as fixed effect and “pig” as random effect. Comparison of the treatments vs. the control were performed with a Dunnett test. The number of samples (n) refers to the number of pigs used. Results were considered statistically significant at P ≤ 0.05 and trends when 0.05 ≤ P ≤ 0.10.
Results
The ADFI, ADG and G:F conversion ratio of post weaning pigs for the 4 dietary treatments are shown in Table 3. No significant differences were observed during the first 2 weeks post weaning. ADFI was linearly reduced (P < 0.001) with the increase of dietary Lys (T0 > T1 > T2 > T3) during the third week post weaning. The decrease in ADFI during the third week accounted for approximately 1.5 g for each percentage (as nutrient content) of SID Lys above the level of the control diet (up to approximately 150 g reduced feed intake at 200% of SID Lys requirement). When looking at the overall (0 to 21 d post weaning) effect of dietary Lys content on ADFI, higher levels of Lys in the feed tended (P < 0.10) to reduce feed intake in post weaning pigs by approximately 0.8 g for each percentage (as nutrient content) of SID Lys added above the level of the control diet (up to approximately 80 g of reduced feed intake at 200% of SID Lys requirement). However, no significant linear response was observed for ADG during the first, second, third week or the overall period. A linear response of Lys levels to reduce G:F ratio was found as a trend (P < 0.10) during the first week. Nonetheless, no differences were observed during the second week, third week or the overall period. The quadratic effect of excess dietary Lys content on performance parameters was not significant (thus, results were not included).
The circulating levels of plasma EAA and NEAA in T0 or T3 fed pigs are shown in Fig. 1 and 2, respectively. Plasma Lys concentrations increased by more than 10-fold (P < 0.001) in the T3 compared to T0. In contrast, plasma levels of His and Val (P < 0.001) as well as Thr and Leu (P < 0.05) were significantly lower in T3 than T0 fed pigs. Of the NEAA, plasma Glu increased (P < 0.05) while Gln tended (P = 0.07) to decrease in T3 compared to T0 pigs. Non-significant differences were observed for other AA plasma concentrations between the dietary treatments. Overall, total EAA levels in plasma, but not NEAA, were significantly higher in T3 when compared to the T0 group (P < 0.001).

Blood levels of essential amino acids (EAA) in pigs fed Lys dietary excesses. Effect of dietary Lys levels (100% of requirements (T0) vs. 200% of requirements (T3)) on EAA (Lys (A), His (B), Ile (C), Leu (D), Met (E), Phev(F), Thr (G), Trp (H) and Val (I)) plasma levels in young pigs. Data are presented as the mean ± SEM (n = 6). ***P ≤ 0.001, **P ≤ 0.01, *P ≤ 0.05

Blood levels of non-essential amino acids (NEAA) in pigs fed Lys dietary excesses. Effect of dietary Lys levels (100% of requirements (T0) vs. 200% of requirements (T3)) on NEAA (Ala (A), Arg (B), Asn (C), Asp (D), Cys (E), Gln (F), Glu (G), Gly (H), Pro (I), Ser (J) and Tyr (K)) plasma levels in young pigs. Data are presented as the mean ± SEM (n = 6). *P ≤ 0.05, tendency = 0.05 ≤ P ≤ 0.10
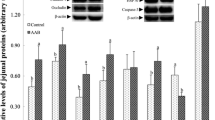
Levels of LDH were 6.77% ± 2.15% and 5.50% ± 1.93% for proximal jejunum and ileum, respectively, when compared to the positive control. Thus, based on the LDH results, Lys at 10, 20 or 30 mmol/L had no significant impact on the integrity of the intestinal tissues (below 10% of the positive control denotes good viability). Gut hormone concentration following Lys exposure are shown in Fig. 3. CCK secretion was enhanced in proximal jejunum tissues incubated with 20 mmol/L (P < 0.05) of Lys compared to the control (KRB/HEPES buffer free of Lys). In contrast, GLP-1 levels did not significantly change in the ileum following exposure to 10, 20 or 30 mmol/L Lys when compared to the control. The effect of Lys at 20 mmol/L (dose chosen for analysis based on gut hormone results) on the mRNA expression levels of AA sensors and receptors in proximal jejunum are illustrated in Fig. 4. The metabotropic glutamate receptor 1 (mGluR1) and the cationic AA transporter solute carrier family 7 member 2 (SLC7A2) gene expression level tended (P = 0.07) to increase in 20 mmol/L Lys treated tissues compared to the control group. Besides these 2 AA sensors, Lys at 20 mmol/L did not enhance the gene expression of other AA receptors and transporters tested in this study.

Cholecystokinin (CCK) and glucagon-like peptide 1 (GLP-1) concentration in porcine intestinal cultures exposed to Lys. CCK and GLP-1 secretion using an ex vivo jejunum (A) or ileum (B) culture from young pigs. Tissue cultures were incubated for 1 h with a KRB/HEPES buffer containing 0 (control), 10, 20 and 30 mmol/L Lys. Data are presented as the mean ± SEM of 3 biological replicates per pig (n = 5). *P ≤ 0.05

Gene expression of AA sensors in porcine jejunum exposed to Lys. Gene expression (mRNA abundance) of AA receptors and transporters (T1R1 (A), T1R3 (B), CaSR (C), mGluR1 (D), mGluR4 (E), SLC7A1 (F) and SLC7A2 (G)) in the proximal jejunum of young pigs following 1 h incubation with a KRB/HEPES buffer containing 0 (control) or 20 mmol/L Lys. Data are presented as mean ± SEM (n = 6). P values indicate a statistical trend: 0.05 ≤ P ≤ 0.10
Discussion
This study illustrated the effect of excess dietary Lys on performance and satiety biomarkers in post weaning pigs. It was hypothesized that Lys dietary excess would significantly reduce feed intake, stimulate the secretion of CCK in the gastrointestinal tract and alter the concentration of EAA and NEAA in blood. In general, the hypothesis was supported by the results.
As anticipated, the dietary inclusion of Ala, particularly in the control diet, did not have a negative effect on feed intake and growth in pigs, which allowed us to differentiate the satiating effect of increasing dietary Lys compared to nitrogen contents. A linear effect of dietary Lys levels on ADFI was observed indicating that the higher the Lys excess the more feed intake was inhibited in post weaning pigs. These results are consistent with the study of Edmonds and Baker [22] in which a depression in feed intake and weight gain was observed in pigs fed dietary increments of Lys between 1.15% and 4.6% (as nutrient content). In the present study, the excess of Lys did not result in changes in growth but marginally decreased the G:F ratio reflecting that the catabolism of excess Lys (nitrogen clearance from the system) could have reduced the energy available for growth (protein accretion) as the liver and kidneys, which are major sites of amino acid degradation, will require more energy to metabolize the high amounts of circulating Lys [23]. However, the same may not have occurred in the pigs supplemented with higher quantities of Ala (control diet) as a significant portion of this NEAA is generally oxidised and used as metabolic fuel by enterocytes before reaching the blood stream in pigs [24]. The absence of statistical significance observed for some of the performance parameters with differences above 10% among treatments (e.g., ADG and G:F ratio) may be related to the relative low number of pigs used (n = 6/treatment).
Consistent with the anorexigenic effect observed in the animals, a 20 mmol/L Lys solution increased CCK secretion from the proximal jejunum. In pigs, AA concentrations above 10 mmol/L have been described in the ileum following consumption of protein meals [25]. This indicates that the doses of Lys tested in the present study, and their impact on CCK, are physiologically relevant. Moreover, these results agree with previous studies indicating an effect of Lys on CCK plasma levels in pigs after an oral gavage [4]. The ability of Lys to stimulate CCK secretion in the jejunum, but not the duodenum, may be related to the higher expression of AA sensors in this segment of the small intestine since it is the principal site of AA absorption [2, 20]. Compatible with this observation, it was observed that the expression of mGluR1 and SLC7A2 tended to increase in Lys treated jejunal tissues. Both mGluRs and AA transporters are expressed in enteroendocrine cells [26, 27]. However, their potential involvement in CCK secretion is speculative since gut hormone release and AA sensing has not been described in pigs to date [28, 29].
In contrast to CCK, Lys did not trigger the release of GLP-1 in the ileum at even supraphysiological doses (30 mmol/L) suggesting that the Lys anorexigenic effect may be independent of enteroendocrine L-cell activation in pigs. It is important to mention as well that, in this study, it is unlikely that high doses of free Lys could have reach the distal ileum and triggered the release of GLP-1 as crystalline AA true ileal digestibility has been reported to be close to 100% in pigs (their absorption is more proximal to that of protein bound AA) [30].
Beside gut peptides, post absorptive signals related to changes in the concentration of AA in blood may also be interpreted as key peripheral signals relevant to satiation and satiety [6, 7, 31]. For instance, intravascular injection of Lys reduced feed intake to a similar extent to an oral gavage in mice, suggesting a direct effect of the AA in the CNS [8]. In our study, the provision of excess Lys in the feed altered the AA blood profile of pigs in 4 distinct patterns. Firstly, Lys supplementation significantly increased the concentration of circulating Lys (pattern 1). These results agree with previous studies in finishing and growing pigs illustrating a linear increase in Lys plasma concentrations with increasing dietary Lys doses [12, 13, 32]. This linear increase has been attributed to the low catabolism and oxidative properties of Lys, which may contribute to a substantial amount of the AA reaching the blood stream intact from the gastrointestinal tract [33]. Once in the blood, Lys may interact with the hypothalamus to modulate appetite in pigs. The interaction of blood AA with the CNS may be partially mediated by tanycytes, radial glial-like cells that line the third ventricle (funnel-shaped structure that is in contact with the hypothalamus) and have access to circulating plasma due to their proximity to fenestrated capillaries [34]. Lys, in particular, has been identified as a robust tanycyte stimulant in rodents [35]. Thus, Lys may interact with the hypothalamus inducing satiety through the stimulation of tanycytes in pigs as well. Future research should investigate the effect of intravenous injections of Lys on feed intake in pigs to corroborate a satiating effect independent of gut hormone secretion.
Secondly, Lys supplementation also affected the circulating levels of five additional EAA and two NEAA. While Glu plasma levels increased (pattern 2), Leu, Val, Thr, His (pattern 3), and Gln levels (pattern 4) decreased in the Lys supplemented pigs compared to the control group. Increases in Glu plasma levels have been associated with low feed intake and satiety in rodents [8]. Interestingly, in previous studies, pigs fed excess Lys showed higher Glu circulating blood levels [13]. The primary site of Lys catabolism is in the liver, producing Glu as one of the main metabolites [33, 36]. Thus, dietary Lys excess seems to result in an increase of liver catabolic pathways raising the circulating levels of Glu. Hence, the CNS sensing of increased Glu plasma levels may contribute to the anorexigenic effect of dietary Lys excess in pigs.
On another note, Lys supplementation was associated with decreased circulating levels of Thr, His and Val. These results are consistent with observations previously reported in growing and finishing pigs [13, 32]. In contrast, Lys supplementation did not affect Leu levels in our experiment. The low blood levels of Leu observed following the increase in Lys intake may be explained by an increase in the activation of intestinal AA antiporters that facilitate the uptake of extracellular cationic AA in exchange for intracellular Leu [37]. The effect of excess dietary Lys on Leu plasma levels is of particular interest considering the strong appetite modulatory properties of Leu in pigs [10, 11]. High circulating levels of Leu may reduce feed intake by interacting with the key hypothalamic (mediobasal and paraventricular hypothalamus) and brainstem structures (nucleus of the solitary tract) as shown in mice [38, 39]. The opposite effect would be expected if plasma levels of Leu were kept low linked to Lys excess levels. The gastrointestinal interaction between Leu and Lys is a subject that merits further investigation.
In contrast to Leu, a direct effect of plasma Val and His on appetite modulatory structures within the hypothalamus has not been established. However, previous studies reported lower performance following the provision of Val and His deficient diets in pigs [40,41,42]. We speculate that the reduction in plasma levels of His and other AA may have been sensed by the brain contributing to the reduced feed intake observed in Lys over supplemented pigs [43]. Potentially, the decrease in plasma levels of EAA are a consequence of a rise in protein synthesis in the whole body related to the increased availability of Lys [13].
Conclusions
Excess dietary Lys reduced feed intake by enhancing satiety in post weaning pigs. Increasing Lys levels in starter diets raised plasma Lys and Glu while lowering circulating levels of Leu, His, Val, and Thr. In addition, Lys increased CCK secretion in the proximal jejunum. Dietary excesses of Lys should be narrowed, when possible, to avoid any potential negative effects on appetite in post weaning pigs.
Availability of data and materials
All data generated or analysed during this study are available from the corresponding author upon reasonable request.
Abbreviations
- AA:
-
Amino acid
- ADFI:
-
Average daily feed intake
- ADG:
-
Average daily gain
- BCAA:
-
Branched-chain amino acid
- CCK:
-
Cholecystokinin
- CNS:
-
Central nervous system
- EAA:
-
Essential amino acid
- G:F:
-
Gain to feed ratio
- GLP-1:
-
Glucagon-like peptide 1
- LDH:
-
Lactate dehydrogenase
- mGluR1:
-
Metabotropic glutamate receptor 1
- NEAA:
-
Non-essential amino acid
- SID:
-
Standard ileal digestible
- SLC7A2:
-
Solute carrier family 7 member 2
References
Presto Åkerfeldt M, Lindberg JE, Göransson L, Andersson K. Effects of reducing dietary content of crude protein and indispensable amino acids on performance and carcass traits of single-phase- and 2-phase-fed growing-finishing pigs. Livest Sci. 2019;224:96–101.
Müller M, Van Liefferinge E, Navarro M, Garcia-Puig E, Tilbrook A, van Barneveld R, et al. CCK and GLP-1 release in response to proteinogenic amino acids using a small intestine ex vivo model in pigs. J Anim Sci. 2022;100:skac093.
Müller M, Xu C, Navarro M, Masiques NE, Tilbrook A, van Barneveld R, et al. An oral gavage of lysine elicited early satiation while gavages of lysine, leucine, or isoleucine prolonged satiety in pigs. J Anim Sci. 2022;100:skac361.
Müller M, Xu C, Navarro M, Masiques NE, Tilbrook A, van Barneveld R, et al. Leucine (and lysine) increased plasma levels of the satiety hormone cholecystokinin (CCK), and phenylalanine of the incretin glucagon-like peptide 1 (GLP-1) after oral gavages in pigs. J Anim Sci. 2023;101:skad175.
Yin J, Han H, Li Y, Liu Z, Zhao Y, Fang R, et al. Lysine restriction affects feed intake and amino acid metabolism via gut microbiome in piglets. Cell Physiol Biochem. 2017;44:1749–61.
Mellinkoff SM, Frankland M, Boyle D, Greipel M. Relationship between serum amino acid concentration and fluctuations in appetite. J Appl Physiol. 1956;8:535–8.
Harper AE, Benevenga NJ, Wohlhueter RM. Effects of ingestion of disproportionate amounts of amino acids. Physiol Rev. 1970;50:428–558.
Jordi J, Herzog B, Camargo SM, Boyle CN, Lutz TA, Verrey F. Specific amino acids inhibit food intake via the area postrema or vagal afferents. J Physiol. 2013;591:5611–21.
Wiltafsky MK, Pfaffl MW, Roth FX. The effects of branched-chain amino acid interactions on growth performance, blood metabolites, enzyme kinetics and transcriptomics in weaned pigs. Br J Nutr. 2010;103:964–76.
Wessels AG, Kluge H, Hirche F, Kiowski A, Schutkowski A, Corrent E, et al. High leucine diets stimulate cerebral branched-chain amino acid degradation and modify serotonin and ketone body concentrations in a pig model. PLoS One. 2016;11:e0150376.
Kwon WB, Touchette KJ, Simongiovanni A, Syriopoulos K, Wessels A, Stein HH. Excess dietary leucine in diets for growing pigs reduces growth performance, biological value of protein, protein retention, and serotonin synthesis1. J Anim Sci. 2019;97:4282–92.
Zeng PL, Yan HC, Wang XQ, Zhang CM, Zhu C, Shu G, et al. Effects of dietary lysine levels on apparent nutrient digestibility and serum amino acid absorption mode in growing pigs. Asian-australas J Anim Sci. 2013;26:1003–11.
Regmi N, Wang T, Crenshaw MA, Rude BJ, Wu G, Liao SF. Effects of dietary lysine levels on plasma free amino acid profile in late-stage finishing pigs. SpringerPlus. 2016;5:888.
National Research Council. Nutrient requirements of swine. 11th ed. Washington, DC: The National Academic Press; 2012.
Li T, Huang S, Li J, Liu H, Wang W, Li N, et al. Dynamic changes of postprandial plasma metabolites after intake of corn-soybean meal or casein-starch diets in growing pigs. J Animal Sci Biotechnol. 2019;10:48.
Adeola O, King DE. Developmental changes in morphometry of the small intestine and jejunal sucrase activity during the first nine weeks of postnatal growth in pigs. J Anim Sci. 2006;84(1):112–8.
Voortman T, Hendriks HF, Witkamp RF, Wortelboer HM. Effects of long- and short-chain fatty acids on the release of gastrointestinal hormones using an ex vivo porcine intestinal tissue model. J Agric Food Chem. 2012;60(36):9035–42.
Valgepea K, Loi KQ, Behrendorff JB, Lemgruber RSP, Plan M, Hodson MP, et al. Arginine deiminase pathway provides ATP and boosts growth of the gas-fermenting acetogen Clostridium autoethanogenum. Metab Eng. 2017;41:202–11.
da Silva EC, de Jager N, Burgos-Paz W, Reverter A, Perez-Enciso M, Roura E. Characterization of the porcine nutrient and taste receptor gene repertoire in domestic and wild populations across the globe. BMC Genomics. 2014;15:1057.
Tian M, Heng J, Song H, Zhang Y, Chen F, Guan W, et al. Branched chain amino acids stimulate gut satiety hormone cholecystokinin secretion through activation of the umami taste receptor T1R1/T1R3 using an in vitro porcine jejunum model. Food Funct. 2019;10:3356–67.
Pfaffl MW. A new mathematical model for relative quantification in real-time rt-pcr. Nucleic Acids Res. 2001;29(9):e45.
Edmonds MS, Baker DH. Failure of excess dietary lysine to antagonize arginine in young pigs. J Nutr. 1987;117(8):1396–401.
Chen HY, Lewis AJ, Miller PS, Yen JT. The effect of excess protein on growth performance and protein metabolism of finishing barrows and gilts. J Anim Sci. 1999;77:3238–47.
He W, Wu G. Oxidation of amino acids, glucose, and fatty acids as metabolic fuels in enterocytes of developing pigs. Amino Acids. 2022;54:1025–39.
Cho CY, Bayley HS. Amino acid composition of digesta taken from swine receiving diets containing soybean or rapeseed meals as sole source of protein. Can J Physiol Pharmacol. 1972;50:513–22.
Reimann F, Williams L, da Silva Xavier G, Rutter GA, Gribble FM. Glutamine potently stimulates glucagon-like peptide-1 secretion from glutag cells. Diabetologia. 2004;47(9):1592–601.
Nakamura E, Hasumura M, Uneyama H, Torii K. Luminal amino acid-sensing cells in gastric mucosa. Digestion. 2011;83(Suppl 1):13–8.
Roura E, Fu M. Taste, nutrient sensing and feed intake in pigs (130 years of research: then, now and future). Anim Feed Sci Technol. 2017;233:3–12.
Roura E, Depoortere I, Navarro M. Review: Chemosensing of nutrients and non-nutrients in the human and porcine gastrointestinal tract. Animal. 2019;13(11):2714–26.
Chung TK, Baker DH. Apparent and true amino acid digestibility of a crystalline amino acid mixture and of casein: comparison of values obtained with ileal-cannulated pigs and cecectomized cockerels. J Anim Sci. 1992;70:3781–90.
Blouet C, Schwartz GJ. Brainstem nutrient sensing in the nucleus of the solitary tract inhibits feeding. Cell Metab. 2012;16(5):579–87.
Roy N, Lapierre H, Bernier JF. Whole-body protein metabolism and plasma profiles of amino acids and hormones in growing barrows fed diets adequate or deficient in lysine. Can J Anim Sci. 2000;80:585–95.
Liao SF, Wang T, Regmi N. Lysine nutrition in swine and the related monogastric animals: muscle protein biosynthesis and beyond. SpringerPlus. 2015;4:147.
Bolborea M, Dale N. Hypothalamic tanycytes: potential roles in the control of feeding and energy balance. Trends Neurosci. 2013;36:91–100.
Lazutkaite G, Soldà A, Lossow K, Meyerhof W, Dale N. Amino acid sensing in hypothalamic tanycytes via umami taste receptors. Mol Metab. 2017;6:1480–92.
Gatrell SK, Berg LE, Barnard JT, Grimmett JG, Barnes KM, Blemings KP. Tissue distribution of indices of lysine catabolism in growing swine. J Anim Sci. 2013;91:238–47.
Reiser S, Christiansen PA. Stimulation of basic amino acid uptake by certain neutral amino acids in isolated intestinal epithelial cells. Biochim Biophys Acta Biomembr. 1971;241:102–13.
Cota D, Proulx K, Smith KAB, Kozma SC, Thomas G, Woods SC, et al. Hypothalamic mTOR signaling regulates food intake. Science. 2006;312:927.
Blouet C, Jo Y-H, Li X, Schwartz GJ. Mediobasal hypothalamic leucine sensing regulates food intake through activation of a hypothalamus–brainstem circuit. J Neurosci. 2009;29:8302.
Gloaguen M, Le Floc’h N, Corrent E, Primot Y, van Milgen J. Providing a diet deficient in valine but with excess leucine results in a rapid decrease in feed intake and modifies the postprandial plasma amino acid and α-keto acid concentrations in pigs. J Amin Sci. 2012;90:3135–42.
Xu YT, Ma XK, Wang CL, Yuan MF, Piao XS. Effects of dietary valine:lysine ratio on the performance, amino acid composition of tissues and mRNA expression of genes involved in branched-chain amino acid metabolism of weaned piglets. Asian-australas J Anim Sci. 2018;31:106–15.
Cemin HS, Vier CM, Tokach MD, Dritz SS, Touchette KJ, Woodworth JC, et al. Effects of standardized ileal digestible histidine to lysine ratio on growth performance of 7- to 11-kg nursery pigs. J Anim Sci. 2018;96:4713–22.
Heeley N, Blouet C. Central amino acid sensing in the control of feeding behavior. Front Endocrinol. 2016;7:148.
Acknowledgements
The authors acknowledge Mr. Allan Lisle statistical counselling and the contribution of the Queensland node of Metabolomics Australia at The University of Queensland for the amino acid analyses of the blood samples collected.
Funding
This study was partially supported by The University of Queensland and Australian Pork Limited as part of the project APL 2016/053.
Author information
Authors and Affiliations
Contributions
MM: conceptualization, investigation, data curation, formal analysis, writing – original draft. EVL: investigation and writing – review and editing. AT: conceptualization, writing – review and editing. RB: conceptualization, writing – review and editing. ER: conceptualization, writing – review and editing, supervision, project administration, funding acquisition and resources. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All procedures were performed under veterinary supervision and approved by The University of Queensland Animal Ethics Committee (Animal Ethics Certificate: CNFS/568/16).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Müller, M., Van Liefferinge, E., Tilbrook, A. et al. Excess dietary Lys reduces feed intake, stimulates jejunal CCK secretion and alters essential and non-essential blood AA profile in pigs. J Animal Sci Biotechnol 15, 24 (2024). https://doi.org/10.1186/s40104-023-00971-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-023-00971-9