
Abstract
Background
Reduced protein diet manifested potential to mitigate heat production based on the concept of ideal amino acid profile. The hypothesis of this study was that lactating sows fed a low crude protein (LCP) diet with supplemental amino acid produce less heat compared to those fed a high crude protein (HCP) diet under both thermal neutral (TN) and heat stress (HS) conditions.
Methods
Thirty-two lactating sows were allotted to HCP (193 g CP/kg) and LCP (140 g CP/kg) diets under thermal neutral (TN, 21 ± 1.5 °C) or cycling heat stress (HS, 32 ± 1.5 °C daytime and 24 ± 1.5 °C nighttime) conditions. Diets contained 0.90% SID lysine and 10.8 MJ/kg net energy. Positive pressure indirect calorimeters were used to measure gas exchange in individual sows with litters, and individual piglets on days 4, 8, 14 and 18. Sow and litter weights were recorded on days 1, 10 and 21.
Results
Sow total heat production (THP) was calculated by subtracting litter THP from sow + litter THP based on BW0.75. Sow BW and body protein (BP) loss was greater for LCP diet compared to HCP diet in peak lactation (P < 0.05 and P < 0.01, respectively) and throughout the entire lactation period (P < 0.05 and P = 0.056, respectively) under HS conditions. Heat-stressed sows fed HCP diet had higher (P < 0.05) rectal temperature at 13:00 (P < 0.05) and 19:00 (P < 0.01), and higher respiration rate at 07:00 (P < 0.05), 13:00 (P < 0.05) and 19:00 (P < 0.05) compared to TN sows fed HCP diet. In sows fed LCP diet, those under HS tended to have higher (P = 0.098) rectal temperature at 13:00 and had higher (P < 0.05) respiration rate at 07:00, 13:00 and 19:00 compared to TN sows. The relationship between daily THP and days in lactation of sows fed LCP diet was quadratic (P < 0.05), with an ascending trend until day 14 and a descending trend from days 14 to 18. Sows fed LCP diet had lower daily THP at day 18 (P < 0.001) compared to those fed the HCP diet under HS conditions.
Conclusion
Reduction in THP in sows fed LCP diet was largely associated with THP on day 18 of lactation under HS conditions. Feeding LCP diets alleviated the increased body temperature in sows under HS conditions throughout lactation, which was accompanied by a reduction in respiration rate. Total heat production is associated with days in lactation, in particular under HS conditions with THP appearing to peak between days 14 and 18.
Similar content being viewed by others

Explore related subjects
Find the latest articles, discoveries, and news in related topics.Background
Despite various cooling strategies, swine production systems are suboptimal in the summer [1]. Heat stress (HS) causes a series of adaptive behavioral and metabolic changes [2], including reduced voluntary feed intake [3, 4] and milk production in sows [5], elevated respiration rate (RR) and body temperature [6], and increased lipid tissue deposition in growing pigs [7, 8]. Swine are naturally HS sensitive due to a lack of functional sweat glands [9] and the existence of a substantial subcutaneous fat layer [8]. Newer genetic lines for greater lean yield have also contributed to an increase in metabolic heat production [7, 10]. In 2003, heat stress contributed to $360 million in annual economic losses to the United States swine industry [1]. This figure increased to $900 million in 2010 [11].
Greater metabolic rate during lactation due to the intense demand for milk production and litter-rearing [12] increases heat sensitivity [5] and HS risk to a larger extent than other production stages [4]. Therefore, reducing heat production in lactating sows exposed to high environmental temperatures may improve production efficiency and welfare. Reducing dietary protein decreases metabolic heat production in growing-finishing pigs [13, 14]. It was estimated [15] that heat production at peak lactation decreased from 288.7 to 154.8 MJ/(d·BW0.75) in sows housed under thermal neutral (TN) conditions by lowering dietary CP from 187 to 138 g/kg. The study objective was to use indirect calorimetry to measure heat production of individual lactating sows fed a diet containing either 184 g CP/kg or a NIAA diet containing 136 g CP/kg and housed under TN or HS environments. It was hypothesized that feeding a reduced-CP diet formulated to contain a near ideal amino acid (NIAA) profile decreases total metabolic heat production in lactating sows under TN and HS conditions compared to feeding a non-reduced CP diet formulated to meet SID lysine requirement with feed ingredients as sole sources of lysine.
Methods
Animals, feeding and experimental design
The experiment was conducted at the USDA-ARS Livestock Behavior Research Unit (West Lafayette, IN) in four consecutive blocks. Thirty-two multiparous (parity 3.25 ± 0.54) lactating Yorkshire × Landrace sows were used, with 8 sows randomly assigned to 1 of 2 dietary treatments per block. In each block, sows were individually housed in farrowing stalls, with 6 located in chambers [12], and 2 for backup substitutes outside of chambers. Sows were exposed to a TN environment (21.0 ± 1.5 °C and 41.8% ± 6.5% relative humidity) in blocks 2 and 4, or cycling HS environment (24.0 ± 1.7 °C and 32.0 ± 1.3 °C during nighttime and daytime, respectively, and 47.3% ± 5.4% relative humidity) in blocks 1 and 3, described in further details below. All sows were acclimated to diets (2.2 kg/d) and ambient temperature 6 days prior to farrowing. After farrowing, HS sows in blocks 1 and 3 were provided ad libitum access to feed. Feed allowance of TN sows (i.e., blocks 2 and 4) was calculated based on feed intake of HS sows within the respective dietary treatments from the preceding block including the backup substitute sows. Feed was provided 3 times daily, and orts were weighed and discarded every other day to avoid interfering with calorimetry day and maintain protocol consistency. No creep feed was provided to piglets and all animals had free access to water. Tail docking, ear notching, teeth clipping, iron injection, and castration were performed according to farm protocol 24 h post birth. Sows were housed in farrowing crates, and litters were standardized to 11.5 ± 0.9 piglets within the first 24 h of birth. Sow and litter weights were recorded, and sow backfat was measured with a backfat scanner (Lean-meater®, series 12, Renco Corp., Golden Valley, MN, USA) on days 1, 10, and at weaning. Milk samples were obtained from all sows on days 6 and 16 to represent early and peak lactation, respectively. Piglets were weaned on day 18 ± 1.7 due to farrowing schedule and constraints of the breeding program. Two sows were weaned on days 15 and 16 and their performance data (feed intake, litter weight gain, piglet ADG) from day 10 to weaning were excluded from the analyses.
Dietary treatments
Ingredients and calculated nutrient composition of the diets are presented in Table 1. Analyzed total (hydrolysate) and free AA concentrations are presented in Table 2. The NRC model [16] was used to estimate requirements for AA, net energy (NE), calcium (Ca) and phosphorus (P). The requirements were predicted based on the following parameters: sow BW of 210 kg, parity number of 2 and above, and daily intake of 6 kg/d, litter size of 10, and piglet ADG of 280 g/d over a 21-day lactation period. The model predicted a minimum sow BW loss of 7.5 kg and the protein:lipid in the model was adjusted to the minimum allowable value of near zero. All diets were formulated to contain the same SID lysine (0.90%) and NE (10.8 MJ/kg) concentrations.
The control diet was formulated using corn and soybean meal as the only sources of lysine to meet NRC [16] SID lysine requirement (0.90%) and consequently contained 187.5 g CP/kg. Valine SID concentration was 0.77% which was near SID requirement of 0.79% [16]. All other indispensable amino acid (IDAA) SID concentrations were in excess relative to NRC [16]. This diet is referred to as the high crude protein (HCP) throughout the remainder of this manuscript.
A second diet balanced to reach a near ideal AA (NIAA) profile was formulated as described in Zhang et al. [17]. Briefly, the NIAA diet was designed by reducing soybean meal relative to corn to meet the minimum SID leucine requirement 1.03%, which corresponded to a CP concentration of 137.5 g/kg. Then, supplemental crystalline source of l-histidine, l-isoleucine, l-lysine, dl-methionine, L-histidine, L-isoleucine, L-lysine, DL-methionine, L-phenylalanine, L-threonine, L-tryptophan and L-valine were added to meet the minimum SID requirement for those AA. Crystalline dl-methionine was added to meet the requirement of methionine + cysteine. This diet is referred to as the low crude protein (LCP) diet throughout the remainder of the manuscript.
Environmental control and physiological monitoring
Under TN conditions, ambient temperature was kept constant at 21.0 ± 1.5 °C, beginning 6 days prior to expected farrowing through weaning. Under HS conditions, a cycling HS approach was used to simulate natural temperature variations over a 24-h period during the summer season. Sows were progressively adapted to increasing ambient temperature over a 6-day period prior to the expected farrowing date, with the basal room temperature of 21.0 °C increased by 1.8 °C per day to a maximum of 32 °C by day 7, which corresponded to day 114 of gestation. The nighttime temperature under HS conditions was maintained at 24 °C. By day 2, the temperature exceeded 24 °C, therefore it was gradually decreased beginning at 15:00 to reach 24 °C by 19:00. During lactation, the room temperature was gradually increased every day from 24.0 °C beginning at 07:00 to 32.0 °C at 11:00, and thereafter the ambient temperature was maintained at 32.0 °C until 15:00. The temperature was gradually decreased beginning at 15:00 to reach 24.0 °C by 19:00.
Physiological indicators of HS included body temperature (vaginal and rectal temperature) and RR. Vaginal temperature was recorded in 10 min intervals, 24 h per day starting at day 3 of lactation until weaning using vaginal implants (iButton, accuracy ±0.1 °C; Dallas Semi-conductor, Maxim, Irving, TX) as previously described [18, 19]. Rectal temperature and RR were recorded daily at 07:00, 13:00, and 19:00 starting at lactation day 1 until day of weaning. Respiration rate (breaths/min) was measured by counting flank movement for 15 s and multiplying by 4 as previously described [19]. Lights were automatically turned off and on at 21:00 and 06:00, respectively.
Indirect Calorimetry
In each block, six sows and their litters were housed in indirect calorimetry chambers and THP was determined on days 4–5, 8, 14–15 and 16–19 of lactation (corresponding to days 4, 8, 14 and 18, respectively, in the remainder of the manuscript). One sow (LCP, block 2, TN) farrowed later than her expected due date and therefore did not participate in the last calorimetry measurement day (i.e., day 18) due to constraints of the breeding schedule. Another sow (LCP, block 1, HS) completed half of her last calorimetry day on day 16 also due to her late farrowing date relative to her expected day. These 2 sows were weaned on days 15 and 16, respectively. Calorimetry was conducted in accordance with methods described in detail by Johnson et al. [12]. Briefly, within each indirect calorimetry testing day, THP was determined from 19:00–07:00 (overnight), 07:00 (pre-feeding), 08:00, 09:00, 10:00, 11:00, 13:00 (pre-feeding), 15:00 and 19:00 (pre-feeding). Indirect calorimetry was also conducted on sentinel piglets to measure their THP on days 4, 8, 14 and 18, and detailed in Johnson et al. [12]. The sentinel litter data were then used as a correction factor to estimate THP of the individual test sows as previously described [12].
Nutrient analysis for diet and Milk
Feed was subsampled and submitted to the Agricultural Experiment Station Chemical Laboratories (University of Missouri-Columbia, Columbia, MO, USA) for AA analysis as previously described [17] to verify accuracy of feed mixing. Milk samples were submitted to the Michigan Dairy Herd Improvement Association (NorthStar Cooperative, Lansing, MI, USA) for analyses of fat, true protein, lactose, total solids and milk urea N (MUN) using infrared spectroscopy.
Calculations
Milk N concentration
Milk N concentration was calculated [16] based on milk true protein and milk MUN concentrations as follows (Eq. 1):
Milk energy concentration
The milk energy content was calculated [20] as follows (Eq. 2):
Heat production
Heat production was calculated [21] as follows (Eq. 3):
Urinary N excretion accounts for only 0.24–0.64% of the THP in pigs [22], therefore it was not included in the calculation.
Sow metabolic CO2 (Eq. 4), O2 (Eq. 5) and THP (Eq. 6) was calculated by subtracting litter THP from sow + litter THP based on BW0.75 of sow and litter, respectively.
Litter weight (LW) could not be recorded on calorimetry days (days 4, 8, 14 and 18), therefore LW was estimated by assuming linear growth rate from days 1 to d 10 and from days 10 to wean day (Eq. 7, 8, 9, 10).
Statistical analysis
Data were analyzed by ANOVA using the Mixed model procedures of SAS 9.4 (SAS Inst. Inc., Cary, NC). For the analysis of performance (Table 3), body composition (Table 4) and milk composition (Table 5) data, the following model was used:
Response = diet + environment + stage + blockenvironment + sowdiet × block + diet × environment + diet × stage + environment × stage + e.
The response of sow depended on the fixed effects of diet (HCP vs. LCP), environment (TN vs. HS), and lactation stage (early vs. peak lactation, if applicable). The random effects included block nested within the environment (TN and HS), individual sow nested within diet and block. The interactive effects of diet × environment, diet × stage, and environment × stage were also included.
For the analysis of physiological data, rectal temperature and RR was first averaged over the lactation period for each sow at each measurement time (07:00, 13:00 and 19:00). (Table 6) and the following model was used:
Response = diet + environment + time + blockenvironment + sowdiet × block + diet × environment + diet × time + environment × time + e.
The response of sow depended on the fixed effects of diet (HCP vs. LCP), environment (TN vs. HS), and repeated measurements of time for body temperature and RR (07:00, 13:00 and 19:00). The random effect included block nested within the environment (TN and HS), individual sow nested within diet and block. The interactive effect of diet × environment, diet × time, and environment × time were also included.
For the analysis of vaginal temperature (Fig. 1), the following model was used:

Vaginal temperature of sows fed high crude protein (HCP; 192.4 g/kg crude protein) and low crude protein (LCP; 140.0 g/kg crude protein) diets and exposed to thermal neutral (TN) and heat stress (HS) environments. Within the same environment (TN or HS), diets (LCP vs. HCP) differed (P < 0.01). Within the same diet (HCP or LCP), environments (HS vs. TN) differed (P < 0.01). Standard error of the mean, SEM = 0.183
Response = diet + environment + day + blockenvironment + sowdiet × block + diet × environment + diet × day + environment × day + e.
The vaginal temperature (i.e., response) was averaged daily, and depended on the fixed effects of diet (HCP vs. LCP), environment (TN vs. HS), and repeated measurement of day of lactation. The random effects included block nested within the environment (TN and HS), individual sow nested within diet and block. The interactive effects of diet × environment, diet × day, and environment × day were also included.
The THP on days 4, 8, 14 and 18 of lactation was analyzed to compare dietary effect (HCP vs. LCP) within each environment (HS or TN) (Table 7). Under HS, ME intake (MEI) between diets varied, thus the MEI was included as a covariable in the model as follows:
Response = MEI + diet + day + block + sowdiet × block + diet × day + e.
The response of sow corrected for MEI depended on the fixed effects of diet (HCP vs. LCP) and repeated measurements of each calorimetry day (days 4, 8, 14 and 18). The random effects included block, individual sow nested within diet and block. The interactive effect of diet × day was also included. Under TN, sows were pair-fed to HS counterparts, and therefore MEI was fixed. The MEI was not an independent and random variable, thus the model was the same as under HS except that the covariable MEI was not included.
The THP at different daytime points on days 4, 8, 14 and 18 of lactation was analyzed to compare dietary effect (HCP vs. LCP) within each environment (HS or TN) via double repeated measurements (day and sampling time) (Table 8). Under HS, MEI was included as a covariable in the model as follows:
Response = MEI + diet + day + sampling time + block + sowdiet × block + diet × day + diet × sampling + day × sampling + e.
The response of sow was corrected by MEI and depended on the fixed effect of diet (HCP vs. LCP), and double repeated measurements of calorimetry day (days 4, 8, 14 and 18) and sampling time (07:00, 08:00, 09:00, 10:00, 11:00, 13:00, 15:00 and 19:00) of CO2 and O2. The random effect included block, individual sow nested within diet and block. The interactive effect of diet × day, diet × sampling time, and day × sampling time were also included. Under TN, the model was the same as under HS, except that the covariable was not included.
Effects were declared significant at P ≤ 0.05 and tendency were declared at 0.05 < P ≤ 0.10.
Results
Experimental diets
Diet composition and nutrient concentrations are presented in Table 1 and IDAA concentrations are presented in Table 2.
Performance
Sow and litter performances are presented in Table 3.
LCP vs. HCP
Under TN and HS, daily feed intake and backfat loss, and litter weight gain did not differ between sows fed LCP and HCP diets at any stages of lactation. Under TN, BW loss did not differ between diets. Under HS, BW loss was greater for sows fed the LCP diet compared to the HCP diet in peak and entire lactation (P < 0.05) period.
HS vs. TN
For the HCP diet, daily feed intake, backfat loss, and litter weight gain did not differ between HS and TN exposed sows at any lactation stage. For the HCP diet, sows under HS conditions lost less BW (P < 0.05) in peak lactation, when compared to TN conditions. For the LCP diet, average daily feed intake, backfat loss, and litter weight gain did not differ between sows housed under HS or TN conditions at any lactation stage. For the LCP diet, sows under HS conditions tended to lose more BW compared to TN conditions (P = 0.052) in peak lactation.
Body lipid and protein mobilization
Body lipid and protein mobilization data are presented in Table 4 and illustrated in Supplementary Figure 1.
LCP vs. HCP
Under TN conditions, body lipid (tissue) and body protein (tissue) mobilization did not differ between sows fed LCP and HCP diets at any lactation stages. Under HS conditions, body lipid (tissue) mobilization did not differ between sows fed LCP and HCP diets at any lactation stage. Under HS conditions, when compared to the HCP diet, sows fed the LCP diet mobilized and tended to mobilize more body protein (tissue) in peak (P < 0.01) and throughout the entire lactation phase (P = 0.056), respectively.
HS vs. TN
For the HCP diet, body lipid (tissue) mobilization did not differ between HS and TN conditions at any lactation stage. Sows fed the HCP diet under HS conditions mobilized less (P < 0.05) body protein (tissue) during peak lactation when compared to TN exposed sows. Body lipid (tissue) mobilization did not differ between HS and TN conditions for sows fed the LCP diet at any lactation stage. Sows fed the LCP diet under HS conditions mobilized more (P < 0.05) protein (tissue) in peak lactation and tended to lose more (P = 0.072) protein (tissue) in the entire lactation period compared to sows fed the LCP diet under TN conditions.
Milk yield and composition
Milk composition data are presented in Table 5.
LCP vs. HCP
Under TN conditions, milk yield, milk true protein, lactose, fat and energy concentrations did not differ between sows fed the LCP and HCP diets at any lactation stage. Sows fed the LCP diet under TN conditions had lower MUN in both early (P < 0.01) and peak (P < 0.01) lactation and tended to have lower milk N concentration (P = 0.098) when compared to sows fed the HCP diet under TN conditions. Under HS conditions, milk yield, milk true protein and lactose did not differ between sows fed LCP and HCP diets at any lactation stage. Under HS conditions, when compared to the HCP diet, sows fed the LCP diet had lower MUN in both early (P < 0.01) and peak (P < 0.01) lactation, and higher milk energy (P < 0.05), fat (P < 0.05), and tendency for lower milk N concentration (P = 0.063) in early lactation. Under HS conditions, milk energy, fat, lactose and N concentrations did not differ between LCP and HCP fed sows in peak lactation.
HS vs. TN
No HS versus TN environmental differences were detected for milk production, milk true protein, MUN, N, energy, lactose and fat concentrations for sows fed either HCP or LCP diets.
Physiological response to ambient temperature
The rectal temperature and RR data are presented in Table 6. Vaginal temperature data are depicted in Fig. 1.
HS vs. TN
Sows fed the HCP diet under HS conditions had higher rectal temperatures at 13:00 (P < 0.05) and 19:00 (P < 0.01), and higher RR at 07:00 (P < 0.01), 13:00 (P < 0.01) and 19:00 (P < 0.01) compared to sows fed the HCP diet under TN conditions. Under HS conditions, sows fed the LCP diet tended to have higher (P = 0.098) rectal temperatures at 13:00, and greater RR at 07:00 (P < 0.01), 13:00 (P < 0.01) and 19:00 (P < 0.01) when compared to sows fed the LCP diet under TN conditions. Overall, sows under HS conditions had higher (P < 0.01) vaginal temperatures throughout the entire lactation period when compared to TN exposed sows, regardless of dietary treatment.
LCP vs. HCP
Under TN, sow rectal temperature and RR did not differ between LCP and HCP diets at 07:00, 13:00 and 19:00. Under HS, compared to HCP diet, sows fed LCP diet had lower rectal temperature (P < 0.05) at 19:00, lower RR at 07:00 (P < 0.05) and tended to have lower RR at 19:00 (P = 0.085). Overall, sows fed the LCP diet had lower (P < 0.01) vaginal temperatures over the entire lactation period when compared to sows fed the HCP diet, regardless of environmental exposure.
Heat production
Total heat production data are presented in Tables 7 and 8.
Nighttime (19:00–07:00)
Under HS conditions, sows fed the LCP diet tended to have lower THP at day 4 (P = 0.092), and lower THP at day 18 (P < 0.05) when compared to sows fed the HCP diet. No other THP differences were detected during nighttime for any comparison.
Daytime (07:00–19:00)
Under TN conditions, sows fed the LCP diet tended to have lower THP (P = 0.093) at day 14 when compared to sows fed the HCP diet. Under HS conditions, sows fed the LCP diet had lower THP (P < 0.01) at day 18 when compared to sows fed the HCP diet. No other THP differences were detected in daytime for any comparison.
24-h period
Under HS conditions, sows fed the LCP diet had lower THP on day 18 (P < 0.001) when compared to sows fed the HCP diet.
Entire lactation period
The relationship between daily (overall 24 h) THP of sows fed LCP diet as lactation progressed was quadratic (P < 0.05) under HS, showing an ascending trend until day 14 and a descending trend from days 14 to 18. This relationship was also observed for sows fed LCP diet under HS environment in daytime (07:00–19:00) (P < 0.05) and nighttime (19:00–07:00) (P < 0.05). For sows fed HCP diet, this relationship was quadratic under TN during daytime (07:00–19:00) (P < 0.05). There was no relationship between THP and days in lactation for sows fed HCP in nighttime under TN.
Entire lactation period (daytime)
Under TN conditions, sows fed the LCP diet had lower (P < 0.05) THP at 07:00 on day 14 when compared to sows fed the HCP diet. Under HS conditions, sows fed the LCP diet tended to have lower THP at 07:00 (P = 0.061) and 15:00 (P = 0.062) on day 8 when compared to sows fed the HCP diet. On day 14, THP tended to be lower in sows fed LCP diet than HCP diet at 10:00 (P = 0.076) and did not differ at other time points. On day 18, THP was lower at 08:00 (P < 0.01), 09:00 (P = 0.08), 10:00 (P < 0.01), 11:00 (P < 0.01), 13:00 (P < 0.01) and 15:00 (P < 0.01), and did not differ at 19:00 (Table 8).
Discussion
Daily metabolic O2 consumption and CO2 production values (supplementary Tables 1 and 2) were similar to those reported in growing pigs [23], ranging from 31.93 to 34.21 L/(d·BW0.75) and 30.99 to 32.42 L/(d·BW0.75) for metabolic CO2 production and O2 consumption, respectively. In addition, daily THP values were similar those reported by Jakobsen et al. [24] who estimated an average THP of 686 kJ/(d·BW0.75) for individual lactating sows fed diets containing 188 g/kg CP by indirect calorimetry and double labeled water technique. Cabezón et al. [25] reported a model predicted-value of 745 kJ/(d·BW0.75) for parity 3–5 sows and assuming a BW of 250 kg. These findings are in line with results of the current study. Earlier on, Bond et al. [26] measured THP of lactating sows, including their litters at 385 kJ/(d·BW0.75) using indirect calorimetry, reflecting lower lactation demand relative to the current study and others. Brown-Brandl et al. [10] and Stinn and Xin [27] reported THP values ranging from 808 to 1418 kJ/(d·BW0.75) and from 1188 to 1695 kJ·/(d·BW0.75), respectively. In both of these studies, calorimetry was conducted at the facility level, hence the THP values include sows with their litters which are expected to be higher than for individual sows. In the current study, results of daily THP including sows and litters (Supplementary Table 7) were also higher than those of sows alone (Table 7).
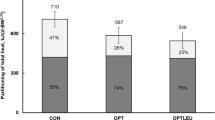
The lower MUN concentration for sows fed LCP diets under both TN and HS conditions resulted from less oxidation of excessive dietary AA and urea synthesis [14, 17, 28], implying lower heat associated with LCP diet. In fact, sows fed the LCP diet produced less daily metabolic heat than those fed the HCP diet throughout lactation, in particular on day 18 under HS conditions. The alleviation of body temperature without an increase in RR under HS conditions indicates that body temperature reduction was diet-induced rather than a result of improved thermoregulation. The estimated THP values of lactating sows based on energy balance [15] were 711 and 586 kJ/(d·BW0.75) with decreasing dietary CP from 187 to 138 g/kg, respectively. In the present study, THP generated from indirect calorimetry decreased from 649 to 582 kJ/(d·BW0.75) under TN conditions, and from 657 to 590 kJ/(d·BW0.75) under HS conditions by feeding the same diets. Thus, this study validates the estimated values by Zhang et al. [15]. In growing-finishing pigs, Kerr et al. [14] reported that decreasing dietary protein from 160 to 120 g/kg reduced THP from 690 to 669 kJ/(d·BW0.75) under TN conditions, respectively, and from 615 to 569 kJ/(d·BW0.75) under HS conditions, respectively. Le Bellego et al. [13] reported a reduction in THP from 1494 to 1393 kJ/(d·BW0.65) in response to decreasing dietary CP from 190 to 120 g/kg, respectively. Herein, a reduction of THP in response to decreasing dietary CP were 12.4 and 13.5 kJ/g CP under TN and HS, respectively. Such values for growing-finishing pigs were up to 7.5 and 20.5 kJ/g CP reduction under TN and HS conditions, respectively [13, 14, 29]. In the study by Kerr et al. [14], pigs under HS had a lower feed intake than those under TN because they were not pair-fed. Thus, it is possible that the difference in feed intake contributed to a larger reduction in heat (20.5 kJ/g CP) compared to values reported herein (13.5 kJ/g CP). Under either TN or HS conditions, both average daily feed intake and milk production did not differ between HCP and LCP diets. Therefore the lower THP in sows fed the LCP diet compared to the HCP diet on lactation day 18 was likely attributed to reduced oxidation of excessive dietary AA and urea synthesis [14, 17, 28]. The theoretical heat reduction associated with less AA intake was 1439 kJ/d [28]. This value was estimated using the NE model for the growing-finishing pig, and thus excluded heat associated with mammary metabolism.
The relationship between THP and days in lactation in this study was previously reported by others [10, 12, 27], and followed a similar trend to that of milk production, piglet growth and nutrient demand [22]. Toner et al. [30] described the milk production curve, composed of the colostral, ascending, plateau and descending phases, with duration of the ascending phase varying from day 14 to 28 of lactation, depending on breed, nutrition, parity, and other factors [31, 32]. Hansen et al. [33] reported a mean time to peak lactation of 18.7 days from a meta-analysis study. Increasing THP with progression of lactation, followed by a descending trend, reflects THP associated with lactation demand. The respiratory quotient (RQ = CO2 output: O2 input) values in this study (supplementary Tables 3 and 6) remained close to 1 throughout lactation, indicating that dietary carbohydrates were serving as primary oxidative substrate [34], and that sows were not in severe negative energy balance. A RQ close to 1 was also previously reported at fed state in growing pigs [10, 23, 35, 36], gestating sows [27, 37] and lactating sows [12, 24, 27].
The lower THP during nighttime compared to daytime, regardless of environmental conditions, was expected and similar to previous findings [10, 27]. This response was likely due to lower feed intake and activity level, as previously described [38], and also due to circadian rhythm differences between daytime and nighttime [10]. Reduced THP between daytime and nighttime corresponded to a 19% and 16% decrease under TN and HS conditions, respectively. Stinn and Xin [27] reported a daytime to nighttime THP reduction of 27% and 6% during late gestation and lactation, respectively, in sows housed at 20 °C.
In addition to less AA oxidation, the lower thermic effect of feeding supplemental AA versus intact protein may possibly contribute to lower THP in LCP diet [16]. To our knowledge, heat increment measurement in lactating sows has not previously been reported. In this study, THP measured at different time points during the day was not affected by the feeding schedule (07:00, 13:00 and 19:00), which was likely attributed to short duration of time between feedings. The longest time between air sampling measurements was 12 h, between the last evening feeding at 19:00 and the morning feeding at 07:00. In growing-finishing pigs, THP was reported to differ between pre- and post-feeding under feed restriction exceeding 30 h [35, 36]. In these studies, the RQ decreased to 0.8, suggesting oxidation of body protein and adipose tissues [34] and pointing to a fasted state [39]. Note that in this study, the RQ before the morning feeding was approximately 1 (see supplementary Table 6), suggesting the major substrate for oxidation was glucose with blood glucose level maintained by hepatic glycogen availability [40]. Therefore, 12 h fasting overnight was not sufficient to elicit a fasting state, despite the high metabolic demands of lactation.
Animals under high ambient temperature reduce their metabolic heat production and improve heat losses by latent and sensible pathways [5]. Thus, reduced feed intake, milk production or growth rate have been considered as adaptive mechanisms to high ambient temperature through mitigation of metabolic heat [5]. It was traditionally recognized that maintenance cost increases under HS in ruminants [41], rodents [42] and swine [43], as a result of greater energy associated with heat dissipation, such as panting. Conversely, using a comparative slaughter technique, Johnson et al. [44] estimated that pigs exposed to HS require 2.5 MJ/d less ME for maintenance than pigs raised under TN conditions. Yunianto et al. [45] also reported lower heat production and reduced plasma triiodothyronine (T3), thyroxine (T4) in broiler chickens under HS than TN. Lower THP under HS was also found in growing pigs [14, 46, 47]. Heat reduction under HS may be related to lower visceral mass [48] or feed intake [46]. In lactating sows, in addition to milk nutrient synthesis, the main contributor to THP is heat increment of feeding [16, 49]. Herein, it was initially planned to pair-feed TN sows to preceding HS sows in order to compare THP between TN and HS. However, feed intake between diets varied within either TN or HS environment, thus MEI was included as a covariable under HS to adjust THP. The MEI under TN was fixed due to pair feeding, and was not an independent and random variable, thus the covariable MEI was not included under TN. Thus, the THP under TN and HS was not compared since THP was analyzed with a different model (i.e., TN without covariable MEI and HS with covariable MEI).
Sows fed the LCP diet lost more BW than those fed the HCP diet under HS conditions, which may be attributed to greater body protein mobilization. Increased partitioning of AA towards the mammary gland at the expense of maternal body reserves in sows fed an LCP diet has been suggested by Huber et al. [50]. Long term exposure to HS environment may further aggravate skeletal muscle catabolism [51,52,53]. The loss of BW and protein reserve is of potential concern for subsequent reproductive cycle [54] and therefore additional research is needed to evaluate the feasibility of feeding an LCP diet over several parities. Similar findings have been reported under TN conditions [17, 50, 55]. On the other hand, others reported that sow BW loss did not differ between HCP and LCP diets under HS conditions [56, 57]. Of note, sows fed HCP diets lost less BW under HS compared to the pair fed TN (PFTN) counterparts in peak lactation, with similar results observed in gilts fed 175 g/kg CP [52]. Thus, PFTN animals may be under greater physiological stress compared to their HS counterparts due to nutrient restriction [52]. Although not observed in this study due to lack of power, the lower THP under HS conditions [5] may lead to more dietary energy partitioning towards maintaining maternal BW compared to PFTN. In the latter, the greater BW loss of PFTN counterparts fed 175 g/kg CP was also due to body protein loss. Conversely, when fed the LCP diet, sows in the current study tended to lose more BW and body protein under HS compared to their PFTN counterparts, suggesting an interaction between diet and environment. It is possible that the LCP diet was limiting in certain AA under HS condition. For instance, AA oxidation increases due to greater maintenance cost under HS conditions [43]. In addition, lactating sows exposed to HS conditions were reported to have reduced milk concentration of arginine, valine and proline [3]. These observations [3, 43] suggest that HS may increase oxidation of certain AA and as a result, may lead to AA imbalance.
Conclusion
Feeding reduced CP diets with a NIAA profile alleviated the increased body temperature of sows under HS environment which was accompanied by a reduction in respiration rate. Feeding LCP diets reduced daily THP by 10.3% over the lactation period, and this reduction was associated with the THP response on day 18 of lactation. Sows fed LCP diet had 73% average reduction in MUN and maintained similar feed intake and lactation performance compared to sows fed HCP, suggesting that reduction of THP in sows fed LCP was attributed to less oxidation of excessive dietary AA and reduced urea synthesis. Total heat production is associated with days in lactation, in particular under HS conditions with THP appearing to peak between days 14 and 18.
Results suggest that sows under HS environment and fed reduced dietary CP with a NIAA balance demonstrated less physiological stress to heat. The reduction of THP also implies an increased dietary energy utilization efficiency for lactation during the later stage of lactation. Zhang et al. [15] also indicated the efficiency of energy utilization based on energy balance data and estimated heat production was greater in the peak stage of lactation in sows fed a NIAA profile diet. These results shed additional light on the potential benefits of feeding low protein diets on a larger scale, including maximizing production efficiency, improving welfare of lactating sows under global warming and potentially mitigating the carbon footprint. Amino acid requirements of lactating sows exposed to HS will need to be re-evaluated in order to formulate diets with NIAA profile that maintain maternal body protein retention in order to implement such nutritional strategy over multiple parities.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on request.
Abbreviations
- AA:
-
Amino acid
- BP:
-
Body protein
- BW:
-
Body weight
- CP:
-
Crude protein
- DM:
-
Dry matter
- HCP:
-
High crude protein
- HS:
-
Heat stress
- IDAA:
-
Indispensable amino acid
- LCP:
-
Low crude protein
- LW:
-
Litter weight
- MEI:
-
Metabolizable energy intake
- MUN:
-
Milk urea nitrogen
- NE:
-
Net energy
- NIAA:
-
Near ideal amino acid profile
- RQ:
-
Respiratory quotient
- RR:
-
Respiration rate
- SID:
-
Standardized ileal digestibility coefficient
- THP:
-
Total heat production
- TN:
-
Thermal neutral
References
St-Pierre NR, Cobanov B, Schnitkey G. Economic losses from heat stress by US livestock industries. J Dairy Sci. 2003;86:E52–77.
Bernabucci U, Lacetera N, Baumgard LH, Rhoads RP, Ronchi B, Nardone A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal. 2010;4:7.
Pérez Laspiur J, Trottier NL. Effect of dietary arginine supplementation and environmental temperature on sow lactation performance. Livst Prod Sci. 2001;70:159–65.
Williams AM, Safranski TJ, Spiers DE, Eichen PA, Coate EA, Lucy MC. Effects of a controlled heat stress during late gestation, lactation, and after weaning on thermoregulation, metabolism, and reproduction of primiparous sows. J Anim Sci. 2013;91:2700–14.
Renaudeau D, Collin A, Yahav S, de Basilio V, Gourdine JL, Collier RJ. Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal. 2012;6:5.
Johnson JS, Boddicker RL, Sanz-Fernandez MV, Ross JW, Selsby JT, Lucy MC, et al. Effects of mammalian in utero heat stress on adolescent body temperature. Int J Hyperth. 2013;29:696–702.
Brown-Brandl TM, Nienaber JA, Xin H, Gates RS. A literature review of swine heat production. Trans ASAE. 2004;47:259–70.
Qu H, Yan H, Lu H, Donkin SS, Ajuwon KM. Heat stress in pigs is accompanied by adipose tissue–specific responses that favor increased triglyceride storage. J Anim Sci. 2016;94:1884–96.
Curtis S. Environmental Management in Animal Agriculture. Iowa: Iowa State University Press. Ames; 1983. p. 6–96.
Brown-Brandl TM, Hayes MD, Xin H, Nienaber JA, Li H. Heat and moisture production of modern swine. ASHRAE Trans. 2014;120:469–89.
Pollmann DS. Seasonal effects on sow herds: industry experience and management strategies. Des Moines: Midwest American Society of Animal Science; 2010.
Johnson JS, Zhang S, Morello GM, Maskal JM, Trottier NL. Technical note: development of an indirect calorimetry system to determine heat production in individual lactating sows. J Anim Sci. 2019;97:1609–18.
Le Bellego L, van Milgen J, Dubois S, Noblet J. Energy utilization of low-protein diets in growing pigs. J Anim Sci. 2001;79:1259–71.
Kerr BJ, Yen JT, Nienaber JA, Easter RA. Influences of dietary protein level, amino acid supplementation and environmental temperature on performance, body composition, organ weights and total heat production of growing pigs. J Anim Sci. 2003;81:1998–2007.
Zhang S, Johnson JS, Qiao M, Trottier NL. Reduced protein diet with near ideal amino acid profile improves energy efficiency and mitigate heat production associated with lactation in sows. J Anim Sci Biotechno. 2020;11:4.
NRC. Nutrient requirements of swine. 11th rev. Washington: Natl. Acad. Press; 2012.
Zhang S, Qiao M, Trottier NL. Feeding a reduced protein diet with a near ideal amino acid profile improves amino acid efficiency and nitrogen utilization for milk production in sows. J Anim Sci. 2019;97:3882–97.
Johnson JS, Shade KA. Characterizing body temperature and activity changes at the onset of estrus in replacement gilts. Livst Sci. 2017;199:22–4.
Kpodo KR, Duttlinger AW, Johnson JS. Effects of pen location on thermoregulation and growth performance in grow-finish pigs during late summer. Trans Anim Sci. 2019;3:1375–82.
Weast RC, Astle MJ, Beyer WH. CRC handbook of chemistry and physics. Boca Raton: CRC Press Inc.; 1984.
Brouwer E. Report of sub-committee on constant and factors. In: Blaxter KL, editor. Energy metabolism, vol. 11. Troon: European Association for Animal Production; 1965. p. 441–3.
Chamberlin DP. Impacts of reducing dietary crude protein with crystalline amino acid supplementation on lactating sow performance, nitrogen utilization and heat production. MS. Thesis. East Lansing: Michigan State niversity; 2017.
Jaworski NW, Liu DW, Li DF, Stein HH. Wheat bran reduces concentrations of digestible, metabolizable, and net energy in diets fed to pigs, but energy values in wheat bran determined by the difference procedure are not different from values estimated from a linear regression procedure. J Anim Sci. 2016;94:3012–21.
Jakobsen K, Theil PK, Jørgensen H. Methodological considerations as to quantify nutrient and energy metabolism in lactating sows. J Anim Feed Sci. 2005;14(Suppl. 1):31–47.
Cabezón FA, Schinckel AP, Richert BT, Peralta WA, Gandarillas M. Development and application of a model of heat production for lactating sows. Prof Anim Sci. 2017;33:357–62.
Bond TE, Kelly CF, Heitman H Jr. Hog house air conditioning and ventilation data. Trans ASAE. 1959;2:1–4.
Stinn JP, Xin H. Heat and moisture production rates of a modern U.S. swine breeding, gestation, and farrowing facility. Trans ASABE. 2014;57:1517–28.
Zhang S, Trottier NL. Dietary protein reduction improves the energetic and amino acid efficiency in lactating sows. Anim Prod Sci. 2019;59:1980–90.
Noblet J, Henry Y, Dubois S. Effect of amino acid balance on nutrient utilization and carcass composition of growing swine. J Anim Sci. 1987;65:717–26.
Toner MS, King RH, Dunshea FR, Dove H, Atwood CS. The effect of exogenous Somatotropin on lactation performance of first-litter sows. J Anim Sci. 1996;74:167–72.
Elsley FWH. Nutrition and lactation in the sow. In: Falconer IR, editor. Lactation. London: Butterworths; 1971. p. 393–411.
Harkins M, Boyd RD, Bauman DE. Effect of recombinant porcine somatotropin on lactational performance and metabolite patterns in sows and growth of nursing pigs. J Anim Sci. 1989;67:1997–2008.
Hansen AV, Strathe AB, Kebreab E, France J, Theil PK. Predicting milk yield and composition in lactating sows: a Bayesian approach. J Anim Sci. 2012;90:2285–98.
Nienaber JA, DeShazer JS, Xin H, Hillman PE, Yen JT. 2009. Measuring energetics of biological processes. In: DeShazer, J. S., editor. Livestock energetics and thermal environmental management. St. Joseph (MI):ASAE; p. 73–112.
Li Z, Li Y, Lv Z, Liu H, Zhao J, Noblet J, et al. Net energy of corn, soybean meal and rapeseed meal in growing pigs. J Anim Sci Biotechno. 2017;8:44.
Lyu Z, Li Y, Liu H, Li E, Li P, Zhang S, et al. Net energy content of rice bran, defatted rice bran, corn gluten feed, and corn germ meal fed to growing pigs using indirect calorimetry. J Anim Sci. 2018;96:1877–88.
Wang Z, Chen Y, Ding J, Liu H, Lyu Z, Dong W, et al. Net energy content of five fiber-rich ingredients fed to pregnant sows. Anim Sci J. 2019;00:1–9.
Pedersen S, Rom HB. 2000. Diurnal variation in heat production from pigs in relation to animal activity. European Agricultural Engineers.
Labussière E, Dubois S, van Milgen J, Bertrand G, Noblet J. Fasting heat production and energy cost of standing activity in veal calves. Br J Nutr. 2008;100:1315–24.
Berg JM, Tymoczko JL, Gatto GJ, Stryer L. Biochemistry. 8th ed. New York: W. H. Freeman and Company; 2015. p. 617–37.
Beede DK, Collier RJ. Potential nutritional strategies for intensively managed cattle during thermal stress. J Anim Sci. 1986;62:543–54.
Collins FG, Mitros FA, Skibba JL. Effect of palmitate on hepatic biosynthetic functions at hyperthermic temperatures. Metabolism. 1980;29:524–31.
Campos PH, Noblet J, Jaguelin-Peyraud Y, Gilbert H, Mormede P, de Oliveira Donzele RFM, et al. Thermoregulatory responses during thermal acclimation in pigs divergently selected for residual feed intake. Int J Biometeorol. 2014;58:1545–57.
Johnson JS, Sanz Fernandez MV, Gutierrez NA, Patience JF, Ross JW, Gabler NK, et al. Effects of in utero heat stress on postnatal body composition in pigs: I: growing phase. J Anim Sci. 2015;93:71–81.
Yunianto VD, Hayashit K, Kaiwda S, Ohtsuka A, Tomita Y. Effect of environmental temperature on muscle protein turnover and heat production in tube-fed broiler chickens. Br J Nutr. 1997;77:897–909.
Collin A, van Milgen J, Dubois S, Noblet J. Effect of high temperature on feeding behavior and heat production in group-housed young pigs. Br J Nutr. 2001;86:63–70.
Renaudeau D, Frances G, Dubois S, Gilbert H, Noblet J. Effect of thermal heat stress on energy utilization in two lines of pigs divergently selected for residual feed intake. J Anim Sci. 2013;91:1162–75.
Rinaldo D, Le Dividich J. Effects of warm exposure on adipose tissue and muscle metabolism in growing pigs. Comp Biochem Physiol. 1991;100A:995–1002.
Cabezón FA, Schinckel AP, Smith AJ, Marchant-Forde JN, Johnson JS, Stwalley RM. Technical note: initial evaluation of floor cooling on lactating sows under acute heat stress. Prof Anim Sci. 2017;33:254–60.
Huber L, de Lange CFM, Krogh U, Chamberlin D, Trottier NL. Impact of feeding reduced crude protein diets to lactating sows on nitrogen utilization. J Anim Sci. 2015;93:5254–64.
Wheelock JB, Rhoads RP, VanBaale MJ, Sanders SR, Baumgard LH. Effects of heat stress on energetic metabolism in lactating Holstein cows. J Dairy Sci. 2010;93:644–55.
Pearce SC, Gabler NK, Ross JW, Escobar J, Patience JF, Rhoads RP, et al. The effects of heat stress and plane of nutrition on metabolism in growing pigs. J Anim Sci. 2013;91:2108–18.
Rhoads RP, Baumgard LH, Suagee JK. Metabolic priorities during heat stress with an emphasis on skeletal muscle. J Anim Sci. 2013;91:2492–503.
Bergsma R, Kanis E, Verstegen MWA, van der Peet-Schwering CMC, Knol E. F. Lactation efficiency as a result of body composition dynamics and feedintake in sows. Livest Sci. 2009;125:208–22.
Chamberlin DP, Rozeboom DW, Erwin S, Trottier NL. Lactation performance in sows fed diets with graded levels of crystalline amino acids as substitute for crude protein at lysine requirement. J Anim Sci. 2015;93(E-Suppl. 2):69.
Chamberlin DP, Powers WJ, Rozeboom DW, Brown-Brandl TM, Erwin S, Walker C, et al. Impact of reduced dietary crude protein concentration with crystalline amino acid supplementation on lactation performance and ammonia emission of sows housed under thermo-neutral and thermal heat stress environments. J Anim Sci. 2015;93(E-Suppl. 2):67.
Johnston LJ. Ellis M, Libal GW, Mayrose VB, Weldon WC, NCR-89 committee on swine management. Effect of room temperature and dietary amino acid concentration on performance of lactating sows. J Anim Sci. 1999;77:1638–44.
Acknowledgements
The authors acknowledge Jim Liesman for the guidance on the statistical analyses, and Jacob Maskal, Torey Raber from USDA-ARS, and Katelin Ade from Purdue Swine Farm for their assistance with animal husbandry. Authors also wish to acknowledge the generous donation of crystalline amino acids by Ajinomoto Animal Nutrition North America, Inc.
Funding
This study was financially supported by the USDA-NIFA (award number 2014–67015-21832).
Author information
Authors and Affiliations
Contributions
The work was mainly conceived and designed by NLT, JSJ, and SZ. Experimental data were collected and analyzed by SZ. The manuscript was mainly written by SZ, and revised by NLT and JSJ. All the authors contributed to the writing, reading and approval of the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
Studies were conducted at the USDA-ARS Livestock Behavior Research Unit with the approval of Purdue Animal Care and Use Committee (protocol #1708001612).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing financial interest.
Supplementary information
Additional file 1:
Table S1. Metabolic oxygen (O2) consumption [L/(d·BW0.75)] of lactating sows fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S2. Metabolic carbon dioxide (CO2) production [L/(d·BW0.75)]of lactating sows fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S3. Respiratory quotient (RQ) of lactating sows fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S4. Metabolic oxygen (O2) consumption [L/(d·BW0.75)]during daytime of lactating sows fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S5. Metabolic carbon dioxide (CO2) production [L/(d·BW0.75)] during daytime of lactating sows fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S6. Respiratory quotient (RQ) during daytime of lactating sows fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S7. Metabolic total heat production [kJ/(d·BW0.75)] of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S8. Metabolic oxygen (O2) consumption [L/(d·BW0.75)] of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S9. Metabolic carbon dioxide (CO2) production [L/(d·BW0.75)] of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S10. Respiratory quotient (RQ) of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S11. Metabolic total heat production [kJ/(d·BW0.75)] during daytime of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S12. Metabolic oxygen (O2) consumption [L/(d·BW0.75)] during daytime of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S13. Metabolic carbon dioxide (CO2) production [L/(d·BW0.75)] during daytime of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S14. Respiratory quotient (RQ) during daytime of lactating sows with litters fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions. Table S15. Metabolic carbon dioxide (CO2) production, oxygen (O2) consumption, total heat production (THP) and respiratory quotient (RQ) of piglets from sows fed high crude protein (HCP) and low crude protein (LCP) diet and exposed to thermal neutral and heat stress conditions.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, S., Johnson, J.S. & Trottier, N.L. Effect of dietary near ideal amino acid profile on heat production of lactating sows exposed to thermal neutral and heat stress conditions. J Animal Sci Biotechnol 11, 75 (2020). https://doi.org/10.1186/s40104-020-00483-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-020-00483-w