
Abstract
Curcuma longa L. extract (CLE) exerts various biological functions including antioxidant, anti-inflammation, anticancer, and antiallergenic effects. However, its immune-enhancing capacity remains unclear. Therefore, the immune-enhancing effect of CLE was investigated in RAW 264.7 cells and cyclophosphamide (CPP)-induced immunosuppression model. CLE upregulated nitric oxide (NO) and reactive oxygen species production and increased inducible nitric oxide synthase and cyclooxygenase-2 expression without affecting the RAW 264.7 cells viability. The results of quantitative real-time reverse transcription polymerase chain reaction and sandwich enzyme-linked immunosorbent assay showed that CLE increased the gene expression and protein levels of tumor necrosis factor-α, interleukin-6, and interleukin-1β in RAW 264.7 cells. Moreover, CLE upregulated p65, I kappa B kinase α/β, and I kappa B α (IκBα) phosphorylation and downregulated IκBα expression in RAW 264.7 cells. CLE also increased p65 translocation from the cytoplasmic to the nucleus in RAW 264.7 cells. The oral administration of CLE increased organ indexes (including the spleen and thymus) and NO production in peritoneal macrophages and improved natural killer cell activity in CPP-induced immunosuppression BALB/c mice. Overall, CLE could be a useful health functional food material that can improve innate immunity via macrophage activation.
Similar content being viewed by others


Introduction
The coronavirus disease 2019 pandemic resulted in increased levels of hospitalizations and deaths worldwide [1, 2]. Thus, there is a need to enhance immunity, which is a defense mechanism against pathogens including viruses, microbes, and particulate matter [3]. Immune-enhancing nutraceuticals are considered as a promising strategy to protect the body from environmental pathogens. Therefore, the food industry is attempting to develop products with immune-enhancing functional materials [4]. Although various researchers are trying to develop new materials for enhancing immunity, research on the development of immune-enhancing nutraceutical materials that protect the host body from pathogen infiltration is still required.
The innate immune system immediately responds to infectious pathogens and acts as the primary defense mechanism before the occurrence of adaptive immunity. However, it is less accurate and specific to antigens than the adaptive immune system. Macrophages are one of the most important early response immune cells in the innate immunity [5]. Macrophage activation plays a critical role in host defense to harmful foreign substances by regulating inflammation, killing pathogens, and repairing tissues [6]. If macrophages do not function normally, they can cause severe health problems including immunodeficiency diseases. Considering that the number or function of macrophages can be weakened in people with reduced immunity (e.g., advanced age or other factors), improving the function of macrophages can contribute to immunity enhancement.
Nuclear factor-kappa B (NF-κB), a representative transcription factor, controls developmental processes, cell growth, apoptosis, and inflammatory and immune responses; it is composed of a heterodimeric catalytic subunit p65 and regulatory subunit p50 [7]. Once the pathogen-associated molecular patterns of foreign pathogens are recognized by the pattern recognition receptors of macrophages, the NF-κB signaling pathways are activated [8]. As an immune enhancement mechanism, NF-κB promotes the transcription of its target genes, including tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and interleukin-1β (IL-1β), and increases the generation of reactive oxygen species (ROS) and nitric oxide (NO) [9, 10]. The cytokines provoke the maturation and proliferation of innate and adaptive immune cells, while ROS and NO directly remove the pathogens; as a result, immune homeostasis is sustained [8, 11].
Curcuma longa L. (CL) is a plant in the Zingiberaceae (ginger) family, the rhizomes of which are used. It is the most cultivated species in the genus Curcuma and mainly grows in temperate climates such as Korea and Southeast Asia [12, 13]. CL contains curcumin, a natural and major physiologically active compound [13]. CL is known to effectively prevent and improve various diseases [4]. It has a wide range of pharmacological properties including anti-memory loss; anticancer; anti-inflammation; and diabetes, liver function, and digestive function improvement [4, 13].
This study showed that CL is a potential immune-enhancing nutraceutical that can increase the gene expression of immune-mediated transporter cytokines (e.g., TNF-α, IL-6, and IL-1β) and production of ROS and NO via upregulation of the NF-κB signaling pathway. Moreover, oral administration of CL extract (CLE) exhibited immune-enhancing effect by improving the spleen index, nitrite production, and natural killer (NK) cell activity in cyclophosphamide (CPP)-induced immunosuppression mice.
Materials and methods
Materials
Dulbecco’s Modified Eagle Medium (DMEM), RPMI-1640, penicillin/streptomycin solution, and fetal bovine serum (FBS) were purchased from Thermo Scientific (Logan, UT, USA). Primary antibodies against p-p65 (#3033), p65 (#8242), I kappa B kinase α (IKKα; #61294), p-IKKα/β (#2697), I kappa B α (IκBα; #4814), p-IκBα (#2859), inducible nitric oxide synthase (iNOS; #13120), cyclooxygenase-2 (COX-2; #12282), and α/β-tubulin (#2148) were purchased from Cell Signaling Technologies (Danvers, MA, USA). Primary antibody against β-actin (sc-47778) was purchased from Santa Cruz Biotech (CA, USA). Antibody against Lamin B1 (ab16048) was purchased from Abcam (Cambridge, Cambridgeshire, UK).
Plant collection and extraction
Curcuma longa L. (CL) was purchased from Jindo Nonghyup (Jindo, Republic of Korea). CL was mixed with deionized water at a 1:30 ratio at 80 °C and then extracted in a constant temperature shaking VS-1205SW1 water bath (Vision Scientific, Bucheon, Korea) for 4 h at 80 °C. Next, the extract was centrifugated for 30 min at 8000 rpm (CR22N, Eppendorf Himac Technologies Co., Ltd., Tokyo, Japan). The supernatant was concentrated at 40 °C using an RKTS-22060-SNN concentrator (Genevac Ltd., Philadelphia, PA, USA) and then lyophilized with an FDU-7024 freeze-dryer (Operon co., Gimpo, Korea). The yield of the water extraction process was 20.66%. The obtained CL extracts (CLE) powder was dissolved in sterile distilled water and used as a sample (25, 50, and 100 μg/mL).
Animals
Six-week-old female BALB/c mice were supplied by Samtaco Bio Korea (Gyeonggi-do, Korea). Mice were housed in 25 °C and 50% humidity-controlled quarters with a 12 h light/dark cycle. All mouse received humane care, and the study protocol (KNU-2022-0386) was approved and carried out d according to Kyungpook National University animal use and care guidelines. Animals were stabilized with access freely to food and water for 1 week prior to oral administration.
Cell culture
Murine RAW 264.7 macrophages were obtained from the Korean Cell Line Research Foundation (Seoul, Korea). Cells cultured using DMEM containing 10% FBS and 1% 100 U/mL penicillin and 100 μg/mL streptomycin solution in CO2 incubator.
MTT assay
To evaluate the toxicity of RAW 264.7 cells to CLE, colorimetric assay with Thiazolyl blue tetrazolium bromide (MTT) was conducted [14]. RAW 264.7 cells were distributed into a 96-well plate (3 × 105 cells/mL) and cultured in CO2 incubator. Treated cells with CLE (25, 50, and 100 μg/mL) cultured for 24 h. The cultured cells were incubated for 4 h by dividing MTT reagent (5 mg/mL; Sigma-Aldrich, St. Louis, MO, USA) into the 96-well plate by 10 μL. After that, about 80 μL of the supernatant of the cell cultured medium was removed, and 100 μL of dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO, USA) was divided. The 96-well plate was wrapped in silver foil and shaken at 15 min and 100 rpm to dissolve. After that, SpectraMax iD3 (Molecular Devices, San Jose, CA, USA) measured absorbance at 595 nm.
Nitrite assay
To evaluate the nitrite (N\({\text{O}}_{2}^{ - }\)) generation of RAW 264.7 cells to CLE, colorimetric assay with Griess reagents was conducted [14]. RAW 264.7 cells were distributed into a 96-well plate (3 × 105 cells/mL) and cultured in CO2 incubator. Treated cells with CLE (25, 50, and 100 μg/mL) cultured for 24 h. Similar amounts of Griess A reagent (0.2% N-(1-naphthyl) ethylenediamine dihydrochloride) and Griess B reagent (1% sulfanilamide and 5% phosphoric acid) were added to 100 μL of cell supernatant at each group for 15 min at 100 rpm to dissolve. After that, SpectraMax iD3 measured absorbance at 550 nm. Sodium nitrite was dissolved by concentration and used as a standard curve of NO.
Measurement of ROS
DCF-DA was used as fluorescent probe to measure intracellular ROS production in RAW 264.7 cells [14]. RAW 264.7 cells were divided into a 96-well plate (3 × 105 cells/mL) and cultured in CO2 incubator. Treated cells with CLE (25, 50, and 100 μg/mL) cultured for 24 h. After washing twice with warm phosphate-buffered saline (PBS), DCFH-DA was diluted to 20 μM in DMEM without phenol red and FBS in a state where light was blocked. Thereafter, 200 μL of DCF-DA dilution medium was added to the cells and reacted for 30 min. After washing twice with warm PBS, the absorbance was measured using SpectraMax iD3.
ELISA assay
Sandwich enzyme-linked immunosorbent assay (ELISA) was conduted to measure the production of cytokines (TNF-α, IL-6, and IL-1β) in RAW 264.7 cells. Cells were divided into 48-well plates (1.5 × 105 cells/mL) and cultured in CO2 incubator. Treated cells with CLE (25, 50, and 100 μg/mL) cultured for 24 h. Cytokines were detected using Mouse cytokine uncoated ELISA kits (Invitrogen, Waltham, MA, USA) according to the manufacturer instructions. After that, SpectraMax iD3 measured at 450 nm and 570 nm.
Western blot analysis
RAW 264.7 cells were distributed into a 6-well plate (3 × 105 cells/mL) and cultured in CO2 incubator for Western blot analysis. Treated cells with CLE (25, 50, and 100 μg/mL) cultured several times. After the reaction time, cells washed twice with cold PBS, and dissolved with lysis buffer (Cell Signaling Technologies, Danvers, MA, USA) coupled with a phosphatase inhibitor cocktail and protease (Thermo Fisher Scientific Inc., Waltham, UT, USA). Cells were recovered into 1.5 mL tubes using a scraper. After voltexing and centrifuging, supernatant was measured using a detergent-compatible protein assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Mixture of protein and dye was loaded onto a 10% sodium dodecyl sulfate–polyacrylamide gel by electrophoresis. Then, it was transferred to a polyvinylidene difluoride membrane (Immobilon®-FL transfer membrane, Millipore, MA, USA). After that, membranes were blocked and were kept primary antibodies overnight at 4 °C. Subsequently, the secondary antibody was reacted for 1 h and treated with EzWestLumi plus to measure protein expression and phosphorylation [15]. GeneGnome XRQ-NPC Imager (Syngene, Cambridge, UK), respectively quantified and visualized protein bands.
Cytoplasmic and nuclear proteins fraction
RAW 264.7 cells distributed into 6 cm dishes (4 × 105 cells/mL) and cultured in CO2 incubator. Treated cells with CLE (25, 50, and 100 μg/mL) for 30 min washed twice with cold PBS, and cells were separated into cytoplasmic and nuclear proteins using Thermo Scientific™ NE-PER extraction kit (Thermo Fisher Scientific Inc., Logan, UT, USA) followed the manufacturer instructions. Lysates were analyzed by Western blot analysis.
qRT-PCR analysis
RAW 264.7 cells were divided into a 6-well plate (3 × 105 cells/mL) and cultured in CO2 incubator. Treated cells with CLE (25, 50, and 100 μg/mL) cultured for 24 h. Cells treated with RNA isolation buffer (TaKaRa, Kyoto, Japan) followed manufacturer instructions. To synthesize cDNA, the extracted RNA was reverse transcribed using ReverTra Ace™ qPCR RT Master Mix (TOYOBO, Osaka, Japan) [16]. Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) was conducted using the SYBR Green Real-Time PCR Master Mix (TOYOBO, Osaka, Japan), and Table 1 displayed the primer sequences. The relative intensity of gene expression was determined on using a qRT-PCR detection system (Bio-Rad Inc., Hercules, CA, USA), and values for each gene were normalized to the GAPDH expression level using comparative ΔΔCq method.
Immunofluorescence
RAW 264.7 cells were divided into chambered coverslip with eight wells for cell culture (Ibidi, Gräfelfing, Germany, 1 × 105 cells/mL) and cultured in CO2 incubator. Treated cells with CLE (50 and 100 μg/mL) for 30 min were fixed with 4% formaldehyde, permeabilized with cold 100% MeOH, blocked with blocking buffers comprising 5% FBS and 0.3% Triton X-100, and incubated with p65. Cells were treated with goat anti-rabbit IgG H&L conjugated to Alexa Fluor® 488-conjugated antibody for visualization. For cytoplasmic p65 staining in red and nuclear p65 staining in blue, cells were stained with VECTASHIELD Antifade Mounting Medium containing 4′,6-diamidino-2-phenylindole (Vector Laboratories, Burlingame, CA, USA) [15]. The translocation of p65 was observed through a fluorescence microscope, and LAS X (Leica Microsystems, Wetzlar, Hessen, Germany) recorded images.
Immunosuppression model and sample administration
Mice were randomly divided into four groups (n = 8 per group): control group administered with DW, CPP-injected group, and CPP-injected and CLE (50 mg/kg/day)-administered group, and CPP-injected and CLE (100 mg/kg/day)-administered group. DW or CLE was administered orally once daily for 19 consecutive days. Immunosuppression was induced in BALB/C mice by intraperitoneal injection of CPP (Sigma-Aldrich, MO, USA) 80 mg/kg on day 17, 18, and 19.
Immune organ index
Animals were sacrificed on day 20. Their spleen and thymus were isolated and weighed. The indexes of spleen and thymus were calculated using the following formula:
Preparation of splenocytes
Spleens were excised from the sacrificed animals and placed in RPMI-1640 (HyClone, Logan, USA) containing 10% FBS. Afterward, the spleens were crushed and passed through a 40-μm pore mesh (SPL Life Sciences, Gyeonggi-do, Korea) to obtain splenocytes. Samples were centrifugated for 5 min at 1300 rpm and 4 °C. To remove red blood cells, splenocytes were resuspended in cold Red Blood Cell Lysis Buffer (Sigma-Aldrich, St. Louis, MO, USA) for 5 min and washed with RPMI-1640. Then, after spinning down, pellet was suspended in medium.
Preparation of peritoneal macrophages
The skin above the peritoneum was removed and visualized to obtain peritoneal fluid. Then, 10 mL of PBS was injected intraperitoneally and the abdomen was massaged to dispense the fluid. Peritoneal lavage fluid was recovered from the peritoneal cavity using a 20-mL syringe and centrifugated for 10 min at 1000 rpm and 4 °C. Pellet was suspended in DMEM, and the number of cells/mL was measured using a hemocytometer. Peritoneal cells were divided into a 96-well plate (2 × 106 cells/mL) and cultured in CO2 incubator. Plate was washed twice with warm PBS and cultured in DMEM with concanavalin A (5 μg/mL) for 24 h. Nitric oxide in peritoneal macrophages was measured using nitrite assay.
Measurement of splenocyte proliferation
Splenocytes were divided using RPMI-1640 and divided into a U-bottom 96-well plate (5 × 106 cells/mL). Then, cells were cultured for 48 h in CO2 incubator. Splenocyte proliferation determined via MTT assay. SpectraMax iD3 measured absorbance at 570 nm.
NK cell activity
YAC-1 cells (target) were divided into a U-bottom 96-well plate (5 × 105 cells/mL) and cocultured with splenocytes (effector) at an E/T ratio of 1:20. The cocultured cells were incubated for 24 h with RPMI-1640 in CO2 incubator. NK cell activity determined via MTT assay. SpectraMax iD3 measured absorbance at 570 nm.
Statistical analysis
All experimental results were represented as mean ± standard deviation. Statistical analyses were organized via GraphPad Prism 9 software (San Diego, CA, USA). The comparison between the control group and the CLE experimental group was analyzed by one-way analysis of variance. Statistical significance was indicated at p < 0.05 compared with control group.
Results
CLE improved NO and ROS production and iNOS and COX-2 expression in RAW 264.7 cells
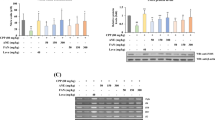
NO is one of the defense compounds that respond to invading pathogens in our body [17]. Therefore, the effect of CLE on NO production was evaluated in RAW 264.7 cells, and 1 μg/mL of lipopolysaccharide (LPS) was adopted as a positive control [18]. The results showed that 100 μg/mL of CLE significantly increased NO production by 312.95% and that CLE (25, 50, and 100 μg/mL) did not affect the viability of RAW 264.7 cells (Fig. 1A, B). ROS could be a criterion for immune capacity because it can directly destroy pathogens [19]. By using dichlorodihydrofluorescein diacetate, a fluorescence probe that responds to ROS, CLE was found to significantly increase ROS production by 209.41% (Fig. 1C, D). Whether CLE affects NO production via the regulation of iNOS and COX-2 expression, an indicator of immune enhancement, was further evaluated [20]. The results showed that CLE significantly increased iNOS and COX-2 expression in RAW 264.7 cells (Fig. 1E, F).

CLE improved NO and ROS production and iNOS and COX-2 expression in RAW 264.7 cells. RAW 264.7 macrophage cells were treated with CLE at 25, 50, and 100 μg/mL for 24 h. A NO was measured using Griess reagents, and CLE increased NO production. B, C ROS was measured using DCF-DA staining, and CLE increased the production of ROS. D CLE significantly improved expression of iNOS and COX-2. E Expression of iNOS, COX-2, and β-actin was detected by Western Blot and quantified by β-actin. F CLE showed no cytotoxicity in RAW 264.7 cells. Mean ± standard deviation from three independent experiments is shown. **p < 0.01 and ***p < 0.001 indicate significant differences compared to control group
CLE enhanced the levels of TNF-α, IL-6, and IL-1β in RAW 264.7 cells
Because cytokines, which are signaling molecules, activate immune cells including complement, phagocytes, and NK cells [21], the effect of CLE on cytokine expression was evaluated. qRT-PCR assay clearly showed that CLE significantly upregulated TNF-α, IL-6, and IL-1β mRNA expression in RAW 264.7 cells (Fig. 2A–C). Additionally, CLE significantly increased TNF-α, IL-6, and IL-1β expression in RAW 264.7 cells (Fig. 2D–F).

CLE enhanced the levels of TNF-α, IL-6, and IL-1β in RAW 264.7 cells. RAW 264.7 macrophage cells were treated with CLE at 25, 50, and 100 μg/mL for 24 h. A–C mRNA expression was analyzed via qRT-PCR, and CLE elevated mRNA expression of TNF-α, IL-6 and IL-1β. D–F Protein levels were measured by sandwich ELISA, and CLE significantly enhanced protein level of TNF-α, IL-6 and IL-1β in cell culture supernatants. Mean ± standard deviation from three independent experiments is shown. *p < 0.05 and ***p < 0.001 indicate significant differences compared to control group
CLE activated the NF-κB signaling pathway and its translocation from the cytosol to the nucleus in RAW 264.7 cells
NF-κB, a transcriptional factor with catalytic p65 and regulatory p50 dimers, plays a vital role in the regulation of the innate immune system [22]. Since CLE promoted the expression of NF-κB target genes including cytokines and iNOS, whether CLE is involved in regulating immune enhancement via the NF-κB signaling pathway was investigated. The results showed that CLE clearly increased p65 phosphorylation in RAW 264.7 cells (Fig. 3A, B). Additionally, CLE increased IKKα/β and IκBα phosphorylation and decreased IκBα degradation in RAW 264.7 cells (Fig. 3C–F). Once activated, NF-κB has to translocate from the cytosol to the nucleus to bind to its specific promoter and subsequently start transcription. The results of Western blot assay with fractioned cytosol and nucleus and immunofluorescence assay clearly showed that CLE increased the localization of p65 from the cytosol to the nucleus in RAW 264.7 cells (Fig. 4A–C).

CLE activated the NF-κB signaling pathway in RAW 264.7 cells. RAW 264.7 macrophage cells were treated with CLE at 25, 50, and 100 μg/mL for 30 min (A) CLE induced phosphorylation of p65 in RAW 264.7 macrophage cells. C CLE induced phosphorylation of IKKα/β and IκBα in RAW 264.7 macrophage cells. Additionally, CLE induced degradation of IκBα in RAW 264.7 macrophage cells. (B, D–F) Expression of NF-κB signaling pathway proteins was detected by Western Blot and quantified. Mean ± standard deviation from three independent experiments is shown. ***p < 0.001 indicates significant differences compared to control group

CLE increased p65 translocation from the cytosol to the nucleus in RAW 264.7 cells. RAW 264.7 macrophage cells were treated with CLE at 25, 50, and 100 μg/mL for 30 min. A, B Nuclear translocation of p65 was analyzed by western blot and quantified, and was elevated by CLE. Lamin B1 and α/β-tubulin were used as loading control in nucleic acid and cytoplasmic protein, respectively. C The localization of p65 was measured via immunofluorescence and CLE showed that p65 moved from the red-stained cytoplasm to the blue-stained nucleus. Mean ± standard deviation from three independent experiments is shown. ***p < 0.001 indicates significant differences compared to control group. C, cytosol fraction; N, nucleus fraction
Oral administration of CLE prevented the loss of spleen and thymus indexes in CPP-induced immunosuppression mice
To confirm whether the immune-enhancing effect of CLE in RAW 264.7 cells could be reflected in vivo, CPP-induced immunosuppression rodent study was performed. The specific experimental procedure is depicted in Fig. 5A. The effects of CLE on body weight and spleen and thymus indexes were evaluated in CPP-induced immunosuppression mice. The body weight of all groups was significantly reduced by CPP injection, except the control group (Fig. 5B). The spleen and thymus indexes were decreased by CPP injection, but oral administration of CLE significantly attenuated these indexes (Fig. 5C–E).

Oral administration of CLE prevented the loss of spleen and thymus indexes in CPP-induced immunosuppression mice. A The CLE at 50, 100 mg/kg/day was orally administrated in mice for 19 days. Also, the CPP at 80 mg/kg/day was injected on days 17, 18 and 19 to induce immunosuppression. B The body weight of each group was measured. C–E After sacrifice, the organ weight and size of spleen and thymus were measured. #p < 0.001 indicates significant differences compared between control group and CPP-injected group. *p < 0.05 indicates a significant differences compared to CPP-injected group
Oral administration of CLE regulated the innate immune system in CPP-induced immunosuppression mice
Since CLE alleviated the reduction of CPP-induced organ index, the effect of CLE on splenocyte proliferation, NO production, and NK cell activity was evaluated in CPP-induced immunosuppression mice. The results showed that CPP injection and oral administration of CLE did not significantly affect splenocyte proliferation (Fig. 6A). To evaluate the innate immune response by CLE, peritoneal macrophages were used to analyze the effect of CLE on NO production. The results showed that oral administration of CLE significantly increased CPP-induced NO production in peritoneal macrophages (Fig. 6B). Additionally, oral administration of CLE significantly increased CPP-induced NK cell activity in primary cultured splenocytes (Fig. 6C).

Oral administration of CLE regulated the innate immune system in CPP-induced immunosuppression mice. The CLE at 50, 100 mg/kg/day was orally administrated in mice for 19 days. Also, the CPP at 80 mg/kg/day was injected on days 17, 18 and 19 to induce immunosuppression. After sacrifice, splenocytes were isolated, and (A) splenocyte proliferation was measured using the MTT assay for 48 h. B The production of NO was measured in peritoneal macrophages with conA using Griess reagents. C NK cell activity targeting YAC-1 was measured using MTT assay. Mean ± standard deviation from three independent experiments is shown. #p < 0.05 indicates significant differences compared between control group and CPP-injected group. *p < 0.05, **p < 0.01 and ***p < 0.001 indicate a significant differences compared to CPP-injected group
Discussion
Innate immune system is the body’s first line of defense against invading pathogen’s infection. The innate immune response is not specific to antigens, but recognizes the characteristics of pathogens and is rapidly activated to help destroy the invader [23]. Therefore, innate immunity enhancement can reduce the risk of various diseases by protecting the body against pathogens. Macrophages are one of the important innate immune cells in the human body. Although the unusual hyperactivation of macrophages in response to environmental pathogens results in inflammation, in a normal state, macrophages play a role in maintaining homeostasis by phagocytosing foreign pathogens and activating other immune cells [5]. The present study aimed to develop an immune-enhancing nutraceutical with activation of the macrophage-mediated innate immune system.
Because NO produced by activated macrophages inhibits the proliferation of harmful bacteria and activates immune cells, the activity of several local food ingredients for NO production was screened. Among them, CLE was selected as one of the most effective materials. The results showed that CLE increased NO production by 3.13 times at 100 μg/mL compared with the control group. A previous study reported that 250 μg/mL of theabrownin isolated from Pu-erh tea increased NO production by 2.41 times compared with the control group [24]. Ren et al. showed that 100 μg/mL of polysaccharides from Cyrtomium macrophyllum increased NO production in RAW 264.7 cells by 3.34 times compared with the control group [25]. In comparing the results of previous studies and the present study, the NO production ability of CLE is similar or superior to that of other natural materials. Although abnormal ROS generation can break the body’s homeostasis by inducing hyperimmunity, ROS play an important role in innate immune response at proper concentrations. ROS not only exert direct antibacterial activity against pathogens such as bacteria, but also regulate immune signaling [26]. In the present study, 100 μg/mL of CLE significantly increased ROS production by 209.4% compared with the control group, which was lower than that of the LPS group, which increased by 570.9%. Therefore, CLE is a potential food ingredient that is capable of immune enhancement via macrophage-mediated NO and ROS production.
Cytokines are signaling substances that activate the immune system and play a major role in the interaction between innate immunity and adaptive immunity [27]. Among them, cytokines including TNF-α, IL-6, and IL-1β are secreted by macrophages as part of their response to infection and tissue damage [28]. Several studies showed that natural materials have immune-enhancing capacity via the upregulation of cytokines. Kim et al. showed that increased secretion of TNF-α, IL-6, and IL-1β from macrophages was shown as an indicator that Limosilactobacillus fermentum promotes immunity [29]. Polysaccharides from Polygonatum sibiricum Delar. ex Redoute increased TNF-α and IL-6 levels in macrophages and subsequently stimulated host immunity [30]. Our qRT-PCR results confirmed that CLE increased TNF-α, IL-6, and IL-1β gene expression and protein level in RAW 264.7 cells. Based on these results, this study verified whether CLE could improve the immune response to pathogens via the upregulation of cytokines including TNF-α, IL-6, and IL-1β.
NF-κB plays an vital role in the immune response of macrophages by promoting NO production through the expression of iNOS gene and the production of cytokines such as TNF-α, IL-6, and IL-1β [27]. To activate NF-κB transcription, NF-κB has to translocate to the nucleus and bind to a specific promoter sequence [27]. Based on the fractioned protein and immunofluorescence assay results, CLE promoted the translocation of p65 from the cytosol to the nucleus via IKKα/β, IκBα, and p65 phosphorylation. A previous study also showed that polysaccharide fraction of Lilium lancifolium Thunb. increases p65 phosphorylation, enhancing murine macrophage activation and exhibiting immune activity [31]. Therefore, it can be assumed that CLE activates NF-κB and promotes its penetration to the nucleus, thereby increasing the immune response through cytokine, NO, and ROS production. However, further studies are needed to determine which components of CLE contribute to this immune enhancement and which upstream molecule of NF-κB is directly affected.
CPP is widely used as a chemotherapy drug in the treatment of various types of cancer, autoimmune diseases, and organ transplants [32]. However, because of the immune disruption of CPP, CPP-induced immunosuppression mouse model is utilized for evaluating immune enhancement [33]. Since CPP has immunosuppressive activity by killing immune cells, the CPP-induced immunosuppression mouse model was considered to be suitable for demonstrating the immune-enhancing effect of CLE by evaluating whether CLE could alleviate the death of immune cells. Based on the innate immune-enhancing effect of CLE, this study verified whether this effect also occurs in CPP-induced immunosuppression mouse model. The results showed that CPP injection significantly decreased the body weight of mice, whereas oral administration did not. The spleen is one of the most important organs in our body that generates lymphocytes and destroys aging red blood cells. Thus, improving the proliferation capacity of spleen cells results in enhanced immune function [34]. CPP injection decreased the spleen and thymus indexes, whereas oral administration of CLE could reverse this effect. However, CLE could not recover the decreased splenocyte proliferation induced by CPP. Because the mechanism of action of CPP is to reduce the number of immune cells, oral administration of CLE was hypothesized to ameliorate the damage of these cells; however, it did not affect the proliferation of immune cells. Interestingly, oral administration of CLE attenuated the inhibition of NO production in peritoneal macrophages similar to the in vitro results. NK cells are leukocytes in the blood responsible for innate immunity [35], and they can recognize and kill cells infected by microorganisms or cancer cells directly [36]. Oral administration of CLE improved NK cell activity targeting YAC-1 cells, but CPP injection did not. These results showed that oral administration of CLE could alleviate impaired immune function through the prevention of decreased spleen and thymus indexes by blocking immune cell damage and improvement of NK cell activity and increasing NO production.
In conclusion, CLE improved NO and ROS production; TNF-α, IL-6, and IL-1β gene and protein expression; and NF-κB signaling pathway in RAW 264.7 cells. Additionally, oral administration of CLE ameliorated the decreased spleen and thymus indexes, NO production, and NK cell activity in CPP-induced immunosuppression mouse model. Taken together, these results suggest that CLE could be a promising nutraceutical for immune enhancement by improving innate immune responses. In addition, in order to apply the results of this study industrially as a health functional food material, follow-up research on the active compound contained in CLE will be needed. The compounds contained in CLE should be identified through UPLC-Q-TOF MS analysis, and the contents of compounds contained in CLE should be quantified through MRM analysis. After that, to specify compound with immune-enhancing activity among the compounds contained in CLE, screening should be performed by evaluating the effect of compounds on various immune-enhancing indicators including NO, ROS, and cytokines production.
Availability of data and materials
All data analysed during this study are included in this published article.
References
O’Driscoll M et al (2021) Age-specific mortality and immunity patterns of SARS-CoV-2. Nature 590(7844):140–145
Waxman JG et al (2022) Comparing COVID-19-related hospitalization rates among individuals with infection-induced and vaccine-induced immunity in Israel. Nat Commun 13(1):2202
Lee J et al (2021) Public Interest in Immunity and the Justification for Intervention in the early stages of the COVID-19 pandemic: analysis of google trends data. J Med Internet Res 23(6):e26368
Tripathy S et al (2021) Curcumin extraction, isolation, quantification and its application in functional foods: a review with a focus on immune enhancement activities and COVID-19. Front Nutr 8:747956
Hirayama D, Iida T, Nakase H (2017) The phagocytic function of macrophage-enforcing innate immunity and tissue homeostasis. Int J Mol Sci 19(1):92
Vasarri M et al (2020) Anti-inflammatory properties of the marine plant Posidonia oceanica (L.) Delile. J Ethnopharmacol 247:112252
Oeckinghaus A, Ghosh S (2009) The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb Perspect Biol 1(4):a000034
Wang J et al (2019) Immunomodulation of ADPs-1a and ADPs-3a on RAW264.7 cells through NF-κB/MAPK signaling pathway. Int J Biol Macromol 132:1024–1030
Liu T et al (2017) NF-κB signaling in inflammation. Signal Transduct Target Ther 2:17023
Lingappan K (2018) NF-κB in oxidative stress. Curr Opin Toxicol 7:81–86
Li C et al (2020) Structural characterization and immune enhancement activity of a novel polysaccharide from Moringa oleifera leaves. Carbohydr Polym 234:115897
Pan MH et al (2017) Effects of water extract of Curcuma longa (L.) roots on immunity and telomerase function. J Complement Integr Med 14(3):20150107
Akaberi M, Sahebkar A, Emami SA (2021) Turmeric and curcumin: from traditional to modern medicine. Adv Exp Med Biol 1291:15–39
Chun A et al (2023) Physicochemical and functional properties of yeast-fermented cabbage. J Microbiol Biotechnol 33(10):1329–1336
So BR et al (2020) Kmeria duperreana (Pierre) dandy extract suppresses LPS-induced iNOS and NO via regulation of NF-κB pathways and p38 in murin macrophage RAW 2647 cells. Prev Nutr Food Sci 25(2):166–172
Kim JG et al (2020) Smilax guianensis vitman extract prevents LPS-induced inflammation by inhibiting the NF-κB pathway in RAW 2647 cells. J Microbiol Biotechnol 30(6):822–829
Schairer DO et al (2012) The potential of nitric oxide releasing therapies as antimicrobial agents. Virulence 3(3):271–279
Yeung YK et al (2021) Structural, antioxidant, prebiotic and anti-inflammatory properties of pectic oligosaccharides hydrolyzed from okra pectin by Fenton reaction. Food Hydrocolloids 118:106779
Martinvalet D, Walch M (2022) Editorial: the role of reactive oxygen species in protective immunity. Front Immunol 12:832946
Lee SM et al (2021) Enhanced immune response by vacuoles isolated from Saccharomyces cerevisiae in RAW 264.7 macrophages. Biosci Rep 41(9):BSR20211158
Kany S, Vollrath JT, Relja B (2019) Cytokines in inflammatory disease. Int J Mol Sci 20(23):6008
Hayden MS, Ghosh S (2011) NF-kappaB in immunobiology. Cell Res 21(2):223–244
Levy JA (2001) The importance of the innate immune system in controlling HIV infection and disease. Trends Immunol 22(6):312–316
Zhao L et al (2023) Theabrownin isolated from Pu-Erh tea enhances the innate immune and anti-inflammatory effects of RAW264.7 macrophages via the TLR2/4-mediated signaling pathway. Foods 12(7):1468
Ren Z et al (2014) Immune-enhancing activity of polysaccharides from Cyrtomium macrophyllum. Int J Biol Macromol 70:590–595
Herb M, Schramm M (2021) Functions of ROS in macrophages and antimicrobial immunity. Antioxidants (Basel) 10(2):313
Somensi N et al (2019) Carvacrol suppresses LPS-induced pro-inflammatory activation in RAW 264.7 macrophages through ERK1/2 and NF-kB pathway. Int Immunopharmacol 75:105743
Trinh TA et al (2020) Effect of herbal formulation on immune response enhancement in RAW 264.7 macrophages. Biomolecules 10(3):424
Kim S et al (2023) Immune-enhancing effects of Limosilactobacillus fermentum in BALB/c mice immunosuppressed by cyclophosphamide. Nutrients 15(4):1038
Zhang J et al (2019) Polysaccharides from Polygonatum sibiricum Delar. ex redoute induce an immune response in the RAW264.7 cell line via an NF-κB/MAPK pathway. RSC Adv 9(31):17988–17994
Pan G et al (2017) Immune-enhancing effects of polysaccharides extracted from Lilium lancifolium Thunb. Int Immunopharmacol 52:119–126
Yan H et al (2021) Prevention of cyclophosphamide-induced immunosuppression in mice with Traditional Chinese Medicine Xuanfei Baidu Decoction. Front Pharmacol 12:730567
Meng F et al (2017) Investigation on the immunomodulatory activities of Sarcodon imbricatus extracts in a cyclophosphamide (CTX)-induced immunosuppressanted mouse model. Saudi Pharmaceut J 25(4):460–463
Lewis SM, Williams A, Eisenbarth SC (2019) Structure and function of the immune system in the spleen. Sci Immunol 4(33):eaau6085
Souza-Fonseca-Guimaraes F, Adib-Conquy M, Cavaillon JM (2012) Natural killer (NK) cells in antibacterial innate immunity: angels or devils? Mol Med 18(1):270–285
Paul S, Lal G (2017) The molecular mechanism of natural killer cells function and its importance in cancer immunotherapy. Front Immunol 8:1124
Acknowledgements
This work was carried out with the support of “Development of K-Immunity Agricultural Food Technology Using Gyeongbuk Ingredients (Project No. LP0048882022)” Gyeongsangbuk-do Agricultural Research & Extension services and National Research Foundation of Korea Grant, funded by the Korean government (MEST) (NRF-2022R1A2C1010923).
Funding
This work was carried out with the support of “Development of K-Immunity Agricultural Food Technology Using Gyeongbuk Ingredients (Project No. LP0048882022)” Gyeongsangbuk-do Agricultural Research & Extension services and National Research Foundation of Korea Grant, funded by the Korean government (MEST) (NRF-2022R1A2C1010923).
Author information
Authors and Affiliations
Contributions
SJK: Data curation; Formal analysis; Software; Investigation; Writing—original draft; Writing—review and editing; Visualization. SK: Investigation; Methodology; Visualization; Formal analysis. SJ: Investigation; Methodology; Visualization. DHG: Investigation. JAR: Resources; Project administration. JMP: Resources; Project administration. SRY: Resources; Investigation; Methodology; Formal analysis. SKJ: Conceptualization; Validation; Writing—original draft; Writing—review and editing; Supervision; Project administration; Funding acquisition.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that there is no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, S.J., Kim, S., Jang, S. et al. Curcuma longa L. extract increased immune responses in RAW 264.7 cells and cyclophosphamide-induced BALB/c mice. Appl Biol Chem 67, 24 (2024). https://doi.org/10.1186/s13765-024-00865-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13765-024-00865-y