
Abstract
Etridiazole (EDZ), a thiadiazole-containing toxic chemical, is widely used as a fungicide. Regular usage of EDZ may reach and contaminate water bodies, but its adverse effects on aquatic vertebrates have not been well studied. Therefore, the present study aimed to evaluate the harmful effects of EDZ using zebrafish (ZF) (Danio rerio) embryos. ZF embryos were treated with 3.75, 7.5, 15, 30, and 60 mg/L of EDZ. Subsequently, mortality and developmental toxicities were quantified at 24, 48, 72, and 96 h post fertilization (hpf). The results showed that embryo mortality was concentration- and time-dependent. The median lethal concentration (LC50) of EDZ at 96-h was 25.58 ± 1.49 mg/L. Besides, EDZ induced a series of morphological deformities, including abnormal somite formation, abnormal eye pigmentation, abnormal tail morphology, tail kinks, skeletal malformations (lordosis, kyphosis, and scoliosis), and yolk sac edema in a concentration-dependent manner. Among the deformities, the most significant were reduced heartbeat and increased incidence of pericardial edema. The median effective concentration (EC50) of EDZ at 96-h was 17.93 ± 2.22 mg/L and the 96-h teratogenic index (TI) value was 1.52. Taken together, these results indicate that EDZ is a teratogen, and primarily affects the cardiovascular system of ZF.
Similar content being viewed by others
Explore related subjects
Find the latest articles, discoveries, and news in related topics.Introduction
The use of thiadiazoles and thiadiazole derivatives as pesticides has been rising worldwide. Etridiazole (EDZ), 5-ethoxy-3-trichloromethyl-1,2,4-thiadiazole, has been widely used as a fungicide for the control of Pythium and Phytophthora species on agricultural and horticultural crops [1]. In Korea, the amount of EDZ imported in 2016 was about 16 tons [2]. In addition to its agricultural usage, EDZ is widely applied to the golf course turf at a relatively high level (1 g/0.93 m2 for every 10–14 days), raising the concern that the golf course runoff may reach surface water, posing a serious threat to non-target aquatic organisms. With an estimated Henry’s Law constant of 3.0 × 10−5 atm-m3/mole, vapor pressure of 0.011 mm Hg, and water solubility of 117 mg/L, volatilization from water surfaces is expected to be an important environmental fate process of EDZ [3]. However, volatilized chemicals can be redeposited by rain and reach the water bodies. Etridiazole is stable for hydrolysis, aqueous photolysis, and not readily biodegradable [1]; hence, it may remain in the aquatic environment for a significant period. The bioconcentration of EDZ in aquatic organisms is very high, and the measured bioconcentration factors of EDZ were 93–323 [3].
Etridiazole is a toxic chemical and its acute exposure to Swiss albino mice inhibits the hepatic drug-metabolizing enzyme system, proving to be hepatotoxic [4]. Etridiazole has been observed to adversely affect body weight gain, viability, and skeletal formation in rabbit fetuses [1]. Moreover, EDZ is known to affect the liver-to-body weight ratio in both rats and dogs [5]. Trichloro acetonitrile (TCAN), a precursor of EDZ, is known to induce embryonic lethality, fetal weight reduction, and cardiovascular anomalies in rats [6]. Studies on Salmonella typhimurium and Chinese hamster ovary cells after EDZ exposure have shown genotoxic reactions; hence, EDZ is considered mutagenic [1]. EDZ has been listed as hazardous to the aquatic environment-category 1 and is highly toxic to non-target aquatic plants [1].
Recently, zebrafish (ZF) has been suggested as the best model for fish acute toxicity studies and is widely used for evaluating the environmental risk of pesticides and other chemicals [7, 8]. ZF is considered an excellent alternative to animal models because of its small size (2–4 cm), high fecundity, large brood size, and short life cycle [7]. Embryonic stages, developmental processes, and organogenesis are well characterized in ZF. Furthermore, ZF embryos develop rapidly outside of the mother and are transparent. Thus, a series of morphological and behavioral adverse effects of the chemical can be scored clearly and used to predict the toxic effect in the ZF model of toxicity [9].
Literature on the toxic effects of EDZ on aquatic organisms, particularly on fish, are scarce. Thus, the present study aimed to evaluate the adverse effects of EDZ using ZF embryos. Organization for Economic Co-operation and Development (OECD) guideline 236 was adopted to establish a concentration-related response of ZF embryos at the early developmental stages: 24, 48, 72, and 96 h post fertilization (hpf) to EDZ. Specifically, embryo mortality and developmental deformities were scored after EDZ exposure. These data demonstrate that EDZ exposure induces cardiac and skeletal deformities in a concentration-dependent manner at a concentration significantly lower than the lethal concentration.
Materials and methods
Chemicals and reagents
Etridiazole (98.4% purity) and 2,4-dichloroaniline (98% purity) were obtained from Sigma-Aldrich (St. Louis, MO, USA). The highest test concentration of EDZ (60 mg/L) was prepared by adding an EDZ solution to the E3 medium, which contained 29 mg of sodium chloride (NaCl), 0.83 mg of potassium chloride (KCl), 4.8 mg of calcium chloride (CaCl2), 8.15 mg of magnesium chloride (MgCl2), and 10 μL of 1% methylene blue per 100 mL of deionized water at pH 7.2. Other test concentrations were obtained by serial dilutions. All chemicals used in this study were obtained from Sigma-Aldrich, unless otherwise stated.
Zebrafish maintenance and embryo collection
Male and female fish were maintained separately in about a 50-L glass aquarium filled with dechlorinated tap water. The aeration and filtration of water were continuously maintained. The fish batches were maintained under a 14 h light/10 h dark photoperiod and a constant temperature of 25 ± 1 °C. The ZF were fed 2–3 times a day with dry flake food (Top meal, Tabia, Korea), live brine shrimp (INVE aquaculture, Dendermonde, Belgium), and blood worms (Hikari Bio-pure, Himeji, Japan).
Mass spawning was performed for egg collection. The required numbers of male and female fish were collected from the main aquariums and were transferred separately into plastic aquariums (25 × 20 × 12 cm, Daehan Biolink, Korea). Fish were fed ad libitum with larvae of Artemia spec. or blood worms. Meanwhile, a plastic aquarium (40 × 25 × 18 cm, Daehan Biolink, Korea) filled with 10 L of dechlorinated tap water and aerated for approximately 1 h to achieve more than 80% oxygen saturation. A nylon mesh (30 × 20 × 13 cm) was installed in the tank to prevent cannibalism by the adult ZF. Five females and five males were transferred to the nylon mesh, and the plastic aquarium was covered with a transparent lid. The culture was kept at a constant temperature of 26.5–27.0 °C and a 14 h light/10 h dark regime. On the next day, the fertilized eggs were collected 30 min after the light was initiated and washed 5 times in E3 medium.
Toxicity assay
The assay was conducted five times in accordance with the OECD guideline 236 (Fish Embryo Acute Toxicity Test) [10]. Briefly, the required quantities of ZF fertilized embryos at around 1.5 hpf were transferred to a Petri dish (100 mm) containing different concentrations (3.75, 7.5, 15, 30, and 60 mg/L) of EDZ. The toxicity assay was conducted in 24 well plates (SPL Life Sciences, Korea). Five plates for five test concentrations of EDZ, one for the positive control (2,4-dichloroaniline at a concentration of 4 mg/L), and one for the control (E3 medium) were used. Twenty wells of each plate were filled with 2 mL of the test solution and the remaining four wells were filled with 2 mL of E3 medium (internal controls). Within 3 hpf, the embryos (one per well) were transferred into plates containing the respective test chemicals. The plates were transferred to an incubator to maintain a temperature of 26 ± 1 °C under dark conditions. Each well was replenished with approximately 1.5 mL of fresh test solution every 24 h.
Deformities scoring
At 24, 48, 72, and 96 hpf, mortality and morphological abnormalities of embryos were documented under a stereomicroscope (Stemi 508, Zeiss, Germany). The embryos with coagulation or the embryos that failed to develop normal were considered dead. At 24 hpf, defects in the somites and tail detachment (TD) were scored. At 48 hpf, abnormalities in eye pigmentation, tail morphology, and tail blood flow were scored. At 72 hpf, unhatched eggs, kink formation in the tail, yolk sac edema (YE), and pericardial edema (PE) were noted. At 96 hpf, unhatched eggs, fish that showed side-wise position (usually ZF “float up” at 96 hpf), swimming ability, response to touch, and spine deformities were scored.
All embryos were evaluated for deformities at all time points. For all deformities, scores were given in binominal way, normal (0) or abnormal (1). The percentages of embryo mortality at 24, 48, 72, and 96 hpf were calculated as the ratio of dead embryo or larvae at the time point of observation over the number of embryos at the start of the exposure (20 fertilized eggs). The percentage of deformity at 24 and 48 hpf was calculated as the ratio of embryos that showed deformity over the number of live embryos at the time point. The percentage of deformity at 72 and 96 hpf was calculated as the ratio of embryos that showed deformity over all embryos at the time point. The resulting percentage data from five independent experiments (n = 5), each with 20 replicates per concentration, were used to calculate the 48-h and 96-h lethal concentration, 50% (LC50) and median effective concentration (EC50 values). For the 48-h EC50 calculation, deformities such as tail detachment, abnormal somites, abnormal eye pigmentation, abnormal tail morphology, and abnormal tail blood flow were used. For the 96-h EC50 calculation, all scored deformities were used.
Heartbeat
After 48 h of EDZ treatment, the embryos were positioned to observe the heart. Heartbeats were counted for 20 s under a stereomicroscope. The obtained data were normalized to heartbeats per minute and presented. Heart beat survey was conducted three times (n = 3) and a total of 60 embryos from control, 57 embryos from 3.75 mg/L of EDZ, 59 embryos from 7.5 mg/L of EDZ, 49 embryos from 15 mg/L of EDZ, 35 embryos from 30 mg/L of EDZ, and 40 embryos from 60 mg/L of EDZ were assessed.
Body length
After 96 h of EDZ treatment, the fish were anesthetized with a final concentration of 0.1% ethyl 3-aminobenzoate methanesulfonate (Sigma-Aldrich, St. Louis, MO, USA) in 24-well plates. Unhatched embryos were gently pressed to break the chorion. Pictures were captured under a stereomicroscope, and the numerical data for the body length, from the mouth tip to the end of the tail fin, were obtained using OptiView 3.7 software (Korealabtech, Seongnam, Korea). The body length survey was conducted three times (n = 3). In each experiment, ten fish per dose of the EDZ and control were used.
Data analysis
The LC50 values, EC50 values, and other statistical analyses were performed using GraphPad Prism software version 5.0 (GraphPad Software, San Diego, CA, USA). Data are presented as mean ± standard error of the mean (SEM). Statistical significance was determined by one-way analysis of variance (ANOVA) to determine the effect of concentration. Statistical significance between the control and EDZ treatments was determined by an unpaired t test. The p values ≤ 0.05 were considered as statistically significant. The teratogenic index (TI) was calculated by dividing the LC50 by the EC50 (LC50/EC50).
Results
Acute toxicity of etridiazole
In this study, embryo mortality was assessed after exposure to various concentrations of EDZ. These data indicate that EDZ-induced toxicity is time- and concentration-dependent (Table 1). As suggested in the OECD 236, there was no mortality in the internal controls until 96 hpf. Furthermore, the mortality in the positive control (3,4-dichloroaniline-treated) was more than 85% even at 24 hpf, supporting the validity of the test. Etridiazole significantly increased the cumulative mortality of ZF embryos (p ≤ 0.0001). The cumulative mortality of the 3.75 mg/L EDZ-exposed embryos was similar to that of untreated embryos (Table 1). The percentage of cumulative mortality increased from 4.0% in the control group to 99.0% in the EDZ-treated groups. Cumulative embryonic death was elevated above the control values from a concentration of 7.5 mg/L of EDZ. Table 2 shows the LC50 values of EDZ at 48 and 96 hpf.
Developmental toxicity of etridiazole
Table 1 shows the summarized data on the several deformities observed at 24, 48, 72, and 96 hpf after EDZ treatment. At a concentration of 3.75 mg/L of EDZ, the ZF embryos were similar to those of the controls (Table 1). However, ZF embryos exposed to EDZ from a concentration of 7.5 mg/L to 60 mg/L showed deformities in a concentration-dependent manner. The most commonly observed deformities were cardiac- and spine-related. Table 2 shows the cumulative EC50 values of EDZ at 48 and 96 hpf. The ratio of cumulative LC50 values to cumulative EC50 values was greater than 1 (Table 2), indicating that EDZ is a teratogen to ZF. Spine bending was the most sensitive end point tested with an EC50 of 12.2 mg/L, followed by PE (EC50: 13.0 mg/L), abnormal tail blood flow (EC50: 13.2 mg/L), abnormal eye pigmentation (EC50: 13.9 mg/L), abnormal somites (EC50: 16.5 mg/L), AT (EC50: 18.1 mg/L), KT (EC50: 21.6 mg/L), embryo mortality at 96 hpf (EC50: 25.6 mg/L), side wise position (EC50: 27.8 mg/L), unable to swim (EC50: 36.8 mg/L), YE (EC50: 38.2 mg/L), and TD (EC50: 52.8 mg/L) (Fig. 1).

Schematic diagram showing the comparison between the cumulative (96-h) LC50 (red text), cumulative (96-h) EC50 (red text), and the corresponding EC50 values of designated phenotype. The corresponding EC50 values for different endpoints investigated in this study (mean ± SEM (n = 3) for spine bending and mean ± SEM (n = 5) for all other endpoints) were plotted as vertical black lines on horizontal grayscale bars ranging from 0 to 60 mg/L of EDZ. The 96-h EC50 (mean ± SEM) indicates the cumulative value (n = 5) of all different endpoints at 96 hpf and 96-h LC50 (mean ± SEM) indicates the cumulative mortality (n = 5) at 96 hpf
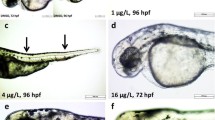
Images of frequently observed abnormal phenotypes and descriptions are shown in Fig. 2. Etridiazole interfered with somite formation in the ZF (p ≤ 0.0001). At 24 hpf, the control and 3.75 mg/L EDZ-treated embryos showed normal somites (Table 1). However, the EDZ-treated embryos from a concentration of 7.5 mg/L showed irregularly placed somite boundaries or no somites (Table 1 and Fig. 2a). The abnormal somite phenotype was greatly enhanced at 15 mg/L of EDZ and reached around 90% at a concentration of 60 mg/L EDZ. Some of the EDZ-treated embryo tails were not detached (Table 1 and Fig. 2a). However, there was no significant difference between the EDZ-treated and control groups.

Representative images showing EDZ-induced deformities. a 24 hpf (upper panel) and 48 hpf (lower panel). b 72 hpf (left side panel), and 96 hpf (right side panel). Controls showed normal morphology until 96 hpf. Yellow dotted line, shape of the tail; Red dotted circle, somites; white dotted circle, eye region; TD, tail detachment; AF, abnormal pectoral fin; PE, pericardial edema; YE, yolk sac edema; KT, kink in the tail; AT, abnormal tail; SB, spine bending; Yellow dotted circle, eyes. Scale 0.5 mm
Abnormalities in the eye pigmentation (retina) was observed after EDZ exposure (p ≤ 0.0001). At 48 hpf, the 3.75 mg/L EDZ-treated embryos showed eye pigmentation similar to that of the controls (Table 1 and Fig. 2a). However, from a concentration of 7.5 mg/L of EDZ, embryos showed abnormal eye pigmentation in a concentration-dependent manner. In addition, EDZ-treated animals exhibited smaller eyes from a concentration of 15 mg/L when compared to controls (Fig. 2b). Abnormal tail morphology (AT) was evident in the EDZ-treated embryos (p ≤ 0.001). AT was elevated from a concentration of 7.5 mg/L EDZ in a concentration-dependent manner (Table 1 and Fig. 2).
At 72 hpf, there was an incidence of YE in EDZ-treated larvae (p ≤ 0.0001). The YEs were evident from a concentration of 15 mg/L EDZ (Table 1 and Fig. 2b). Some of the EDZ-treated embryos were not hatched (p ≤ 0.005 for 72 hpf, p ≤ 0.0001 for 96 hpf). At 72 and 96 hpf, the hatching of the 3.75–30 mg/L EDZ-treated embryos was similar to that of controls (Table 1). However, the 60 mg/L of EDZ-treated embryos were mostly not hatched. In addition, the EDZ-treated fish displayed a kink in the tails (KT) (p ≤ 0.05). The tail morphology of the 3.75 mg/L EDZ-treated ZF was similar to that of the control (Table 1 and Fig. 2b). However, KT was observed from a concentration of 7.5 mg/L of EDZ.
Etridiazole induced several deformities in the spine of ZF larvae (p ≤ 0.0001). Whereas the control fish presented a normal spine shape, EDZ-treated fish from a concentration of 7.5 mg/L of EDZ exhibited body axis curvature (Table 1). Spine deformities include lordosis (inward curvature of the spine), kyphosis (outward curvature of the spine), and scoliosis (3D rotation or sidewise curvature of the spine). The lordosis was more evident at lower concentrations of EDZ (7.5–15 mg/L) and accounted for ~ 90% of spinal deformities. Kyphosis was less frequent (~ 5% of spinal deformities). At higher concentrations of 30 and 60 mg/L EDZ, most of the fish displayed scoliosis.
Effect of etridiazole on heart development and cardiovascular function
After 48 h of EDZ treatment, subjective comparisons of the edema formation, regional blood flow, and heartbeats were assessed in the control and EDZ-treated groups as indicators of cardiovascular development and function [11]. Cardiovascular deformities were found to be increased in a concentration-dependent manner (Table 1 and Fig. 3). Etridiazole induced PE in a concentration-dependent manner (p ≤ 0.0001). There was an increased incidence of PE in all the tested EDZ concentrations (72 hpf; Table 1 and Fig. 3a). Furthermore, EDZ-treated groups showed elongation of the hearts (Fig. 3a), suggesting that EDZ may have a potential impact on cardiogenesis.

a Representative images of ZF embryos showing pericardial edema at the indicated concentration of EDZ. The dotted red line indicates the morphology of the heart. Scale 0.5 mm. b Represents the average heartbeats per minute at designated concentrations of EDZ counted at 48 hpf. The results are mean ± SEM (n = 3). Statistical difference of the EDZ treatment over the control was analyzed by unpaired t-test. **(p ≤ 0.005), ***(p ≤ 0.001)****(p ≤ 0.0001). c Representative images showing hyperemia (arrowhead) at indicated concentrations. Scale 0.5 mm
To assess the effect of EDZ on cardiovascular function, the heartbeats of the ZF embryos in the control and EDZ treatment groups were monitored. Figure 3b shows the average heartbeats of the ZF embryos treated with various concentrations of EDZ for 48 h. Etridiazole exposure led to a reduction of the heartbeats in a concentration-dependent manner (p ≤ 0.0001), significantly affecting the heartbeats from a concentration of 3.75 mg/L. In the control group, the average heartbeats per minute were noted as 158.10, whereas the heartbeats per minute were noted in the range of 149.60–87.29 in the EDZ-exposed groups. These results indicated that EDZ affects cardiac function.
The blood flow in the dorsal aorta region of the EDZ-treated embryos was abnormal (p ≤ 0.0001), whereas the blood flow of the embryos treated with 3.75 mg/L EDZ was similar to that of the controls (Table 1). However, the blood flow of the embryos that were exposed to higher concentrations of EDZ was either reduced or stopped, further supporting the notion that EDZ affects cardiac function. A considerable number of embryos showed severely stagnant blood cells (hyperemia) from a concentration of 7.5 mg/L EDZ (Fig. 3c), suggesting circulatory defects in EDZ-treated fish.
Effect of etridiazole on growth and behavior
To assess the effect of EDZ on overall growth, the body lengths of the control and EDZ-treated groups were measured at 96 hpf. Figure 4 shows representative images and the average body length of ZF treated with different concentrations of EDZ. The average body length of the controls was noted as 3.99 mm. The EDZ-treated groups showed reduced body length in a concentration-dependent manner (p ≤ 0.001). The EDZ-treated fish showed 3.68 (3.75 mg/L), 3.73 (7.5 mg/L), 3.73 (15 mg/L), 3.30 (30 mg/L), and 2.32 (60 mg/L) mm of body lengths, respectively. At a concentration of 60 mg/L EDZ, the ZF displayed approximately 42% less body length than the controls.

a Representative images of ZF control and treated with designated concentrations of EDZ. b Graph showing the average body length of 96 hpf ZF at indicated concentrations of EDZ. The results are mean ± SEM (n = 3). Statistical difference of the EDZ treatment over the control was analyzed by unpaired t-test. *(p ≤ 0.05), **(p ≤ 0.005)
The swim bladders of the ZF at 3.75 mg/L of EDZ treatment were similar to that of control ZF (data not shown). However, more than 50% of EDZ-treated ZF from a concentration of 7.5 mg/L exhibited either malformed swim bladders or uninflated swim bladders (Additional file 1: Fig. S1). Intriguingly, most of the fish in the EDZ groups showed a ‘side-wise position’ (p ≤ 0.0001) and were unable to swim in response to touch stimuli (p ≤ 0.0001). The control, 3.75, and 7.5 mg/L EDZ-treated fish displayed normal swimming behavior and quickly escaped in response to touch (Table 1). However, the fish that were treated with 15 mg/L of EDZ showed swimming disabilities. Abnormal swimming was further enhanced at 30 mg/L of EDZ. At 30 mg/L of EDZ, the fish showed a mild response to touch and were unable to swim. Taken together, these results suggested that EDZ influences the swimming behavior of the ZF.
Discussion
The present study investigated the adverse effects of EDZ on ZF embryos by scoring multiple phenotypical endpoints after 24, 48, 72, and 96 h of exposure to EDZ. The results obtained in this study demonstrate that EDZ could cause several developmental deformities at a concentration significantly lower than the concentration that induces mortality.
The 96-h LC50 value of EDZ for the ZF embryos was 26 mg/L (Table 2). A similar result (23 mg/L) was observed when the sheepshed minnow was exposed to an EDZ metabolite (3-DCMT) [12]. Contrary to these findings, EDZ showed a much lower 96-h LC50 (2.4 mg/L) for the rainbow trout (Pesticide Properties DataBase (PPDB)) [13]. There were, however, diverging opinions on the relationship between ZF embryo toxicity and juvenile toxicity. For instance, a correlation study conducted between ZF embryos and routinely used juveniles (sheepshed minnow, bluegill sunfish, and rainbow trout) showed a poor correlation when all the pesticides were considered as a single group [14]. However, the authors found a better correlation between ZF embryo toxicity and juvenile toxicity when the pesticides were segregated based on their mode of action [14]. Moreover, the difference in the route of exposure and absorption of the chemical between developmental stages may influence the effective toxicity. The rainbow trout toxicity study showed that embryos were relatively resistant to the toxic effects of chemicals, and susceptibility increased during embryonic development and reached its maximum at an early fry stage [15]. The relative resistance of the embryos is partly attributed to the presence of chorion [16], which may protect the embryos by slowing down the entry of toxicants. The temperature at which the assay is performed might be another important parameter that influences the LC50 values [17].
Along with the series of deformities, EDZ-exposed fish displayed reduced body size (Fig. 4). Previous reports suggest that protein synthesis plays a significant role in the normal growth and development of ZF [18, 19]. The knockdown of ribosomal proteins, which are the key players in protein synthesis, leads to growth retardation and several other deformities in ZF. Moreover, Suzuki et al. [20] reported that 1,2,4-thiadiazole derivatives modulate muscarinic acetylcholine receptors (mAChRs), which are known to modulate protein biosynthesis [21, 22]. Therefore, it is plausible that the growth retardation and other deformities observed after EDZ exposure might be due to perturbations in protein biosynthesis. However, further experiments are needed to support this notion.
In ZF, the heart is the first organ to develop and function; hence, it appears to be a primary target for developmental toxicity [23]. Etridiazole induced a series of cardiac-related deformities in ZF embryos (Table 1 and Fig. 3), suggesting that EDZ affects cardiac development and function. In agreement with these results, Long-Evans rats treated with TCAN showed cardiovascular deformities at a concentration as low as 1 mg/kg/day [6]. Similarly, a report showed that triazole-containing fungicides disrupt the development of the heart [24]. The occurrence of pericardial edema is an indicator of abnormal cardiac development [23]. The EDZ-treated groups displayed a high incidence of PE (Table 1). Similarly, ZF exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin [25] and endosulfan [26] displayed PE. Pericardial edema may also be an indicator of osmotic or metabolic dysfunction [25]. Some results suggest that YE can inhibit the supply of nutrients to the heart, resulting in pericardial edema [27]. Therefore, further experiments are required to determine the exact cause of EDZ-induced PE.
Heartbeat is considered a significant indicator of cardiac function [11]. In ZF, the regular heartbeat starts at an early point of 36 hpf [11]. Zebrafish embryos exposed to EDZ displayed reduced heartbeats (Fig. 3b). Earlier reports showed that organophosphate pesticides, which are acetylcholinesterase (AChE) inhibitors, reduce heartbeats in ZF [28]. Overstimulation of the mAChRs is also known to reduce heartbeats in ZF [29]. Interestingly, it was reported that 1,2,4-thiadiazoles derivatives modulate mAChRs [20]. Thus, it is possible that EDZ may reduce heartbeats by modulating the acetylcholine receptors. However, the exact cause of the reduction of the heartbeat in response to EDZ remains to be evaluated. Regardless, these results suggest that the progressive loss of heart function is one of the earliest responses to EDZ in the developing ZF.
Among the deformities, the skeletal malformations were significantly increased in ZF after exposure to EDZ (Table 1). Similar deformities have been reported in ZF embryos exposed to organophosphate insecticides [30]. Spine deformities might be the consequence of dysregulated Wnt signaling or decreased collagen in the spine column [31]. Spine deformities might also be linked to calcium and phosphorous ion depletion or deregulation, which are necessary for the normal development of ZF [32].
Swimming inability was another significant deformity observed in the EDZ-treated groups (Table 1). Similar results were observed in ZF after exposure to endosulfan [33], chlorpyrifos [34], and profenofos [35]. It has been well established that inhibition of AChE activity in the brain impairs swimming in ZF [32, 34]. Abnormal swim bladders (Additional file 1: Fig. S1) may also hamper swimming. The swim bladder is a gas-filled organ that is essential for the survival of most teleost species for the regulation of buoyancy. Swim bladder inflation and float up are visible shortly after hatching. In fact, the EDZ-treated fish that were unable to swim did not float up (not shown). The abnormal spine and tail shapes induced in response to EDZ (Table 1) might be the other causes for abnormal swimming [36].
In summary, these results demonstrate that EDZ strongly interferes with the normal development and function of the cardiovascular system of ZF. Teratogenic effects on the spine, tail, yolk sac, and heart were observed at concentrations lower than those causing lethality, indicating that EDZ is a teratogen. Our results primarily provide detailed documents of EDZ with regard to the developmental toxicity in ZF. This study highlights the importance of monitoring the EDZ content during the manufacturing process and the rational use of pesticides. This study also serves as a model to perform screening of thiadiazole-containing pesticides and drugs to a certain extent.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its additional file.
Abbreviations
- EC50 :
-
Median effective concentration
- EDZ:
-
Etridiazole
- hpf:
-
Hours post fertilization
- LC50 :
-
Median lethal concentration
- mAChRs:
-
Muscarinic acetylcholine receptors
- OECD:
-
Organization for economic co-operation and development
- PE:
-
Pericardial edema
- TCAN:
-
Trichloroacetonitrile
- YE:
-
Yolk sac edema
- ZF:
-
Zebrafish
References
Reregistration eligibility decision (RED) Etridiazole (Terrazole®) (2000) US EPA 738-R-00-019: 1-124
Agrochemical yearbook (2018) Korean crop protection association, pp396
National Center for Biotechnology Information. PubChem compound summary for CID 17432, Etridiazole. https://pubchem.ncbi.nlm.nih.gov/compound/Etridiazole. Accessed 10 Oct 2020
Dalvi R, Howell C (1977) Toxic effects of a fungicide, 5-ethoxy-3-(trichloromethyl)1, 2, 4thiadiazole (terrazole), on the hepatic drug metabolizing enzyme system in mice. Bull Environ Contam Toxicol 17:225–232
Borzelleca JF, Egle JL, Hennigar GR, Klein HH, Kuchar EJ, Lane RW, Larson PS (1980) A toxicologic evaluation of 5-ethoxy-3-trichloromethyl-1, 2, 4-thiadiazole (ETMT). Toxicol Appl Pharmacol 56:164–170
Smith M, Randall J, Tocco D, York R, Stober J, Read E (1988) Teratogenic effects of trichloroacetonitrile in the Long-Evans rat. Teratology 38:113–120
Nagel R (2002) DarT: the embryo test with the zebrafish Danio rerio a general model in ecotoxicology and toxicology. Altex 19:38–48
Lammer E, Carr G, Wendler K, Rawlings J, Belanger S, Braunbeck T (2009) Is the fish embryo toxicity test (FET) with the zebrafish (Danio rerio) a potential alternative for the fish acute toxicity test? Comp Biochem Physiol C: Toxicol Pharmacol 149:196–209
Busquet F, Strecker R, Rawlings JM, Belanger SE, Braunbeck T, Carr GJ, Cenijn P, Fochtman P, Gourmelon A, Hübler N (2014) OECD validation study to assess intra-and inter-laboratory reproducibility of the zebrafish embryo toxicity test for acute aquatic toxicity testing. Regul Toxicol Pharmacol 69:496–511
OECD guidelines test number 236 (2013) Fish embryo acute toxicity (FET) test. Guidelines for the Testing of Chemicals
Yalcin HC, Amindari A, Butcher JT, Althani A, Yacoub M (2017) Heart function and hemodynamic analysis for zebrafish embryos. Dev Dyn 246:868–880
Pesticide ecotoxicity database (2017) United States environmental protection agency. https://ecotox.ipmcenters.org/details.cfm?recordID=36559. Accessed 03 Mar 2020
Pesticide properties database (PPDB) (2009) University of Hertfordshire. https://sitem.herts.ac.uk/aeru/ppdb/en/Reports/285.htm. Accessed 17 June 2020
Glaberman S, Padilla S, Barron MG (2016) Evaluating the zebrafish embryo toxicity test for pesticide hazard screening. Environ Toxicol Chem 36:1221–1226
Van Leeuwen CJ, Griffioen PS, Maas-Diepeveen Vergouw WHA (1985) Differences in susceptibility of early life stages of rainbow trout (Salmo gairdneri) to environmental pollutants. Aquat 7:59–78
Kais B, Schneider KE, Keiter S, Henn K, Ackermann C (2013) DMSO modifies the permeability of the zebrafish (Danio rerio) chorion-implications for the fish embro test (FET). Aquat Toxicol 140:229–238
Uddin MH, Alim MS, Islam SMM, Rashid H, Shahajahan M (2018) Temperature changes alter the acute toxicity responses of cypermethrin in zebrafish. Progress Agric 29:64–70
Uechi T, Nakajima Y, Nakao A, Torihara H, Chakraborty A, Inoue K, Kenmochi N (2006) Ribosomal protein gene knockdown causes developmental defects in zebrafish. PLoS ONE. https://doi.org/10.1371/journal.pone.0000037
Palasin K, Uechi T, Yoshihama M, Srisowanna N, Choijookhuu N, Hishikawa Y, Kenmochi N, Chotigeat W (2019) Abnormal development of zebrafish after knockout and knockdown of ribosomal protein L10a. Sci Rep 9:1–11
Suzuki J, Okamura D, Gushikawa T, Hirai K, Ando T (2011) Synthesis and insecticidal activity of 1, 2, 4-oxadiazole and 1, 2, 4-thiadiazole derivatives. J Pestic Sc 36:392–401
Park YS, Liu Z, Vasamsetti BMK, Cho NJ (2016) The ERK1/2 and mTORC1 sgnaling pathways are involved in the muscarinic acetylcholine receptor-mediated proliferation of SNU-407 Colon Cancer Cells. J Cell Biochem 117:2854–2863
Vasamsetti BMK, Liu Z, Park Y-S, Cho NJ (2019) Muscarinic acetylcholine receptors regulate the dephosphorylation of eukaryotic translation elongation factor 2 in SNU-407 colon cancer cells. Biochem Biophys Res Commun 516:424–429
Hill AJ, Teraoka H, Heideman W, Peterson RE (2005) Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol Sci 86:6–19
Yekti APA, Hsu HJ, Wang WD (2014) The effect of paclobutrazol on the development of zebrafish (Danio rerio) embryos. Zebrafish 11:1–9
Hill AJ, Bello SM, Prasch AL, Peterson RE, Heideman W (2004) Water permeability and TCDD-induced edema in zebrafish early-life stages. Toxicol Sci 78:78–87
Stanley KA, Curtis LR, Simonich SLM, Tanguay RL (2009) Endosulfan I and endosulfan sulfate disrupts zebrafish embryonic development. Aquat Toxicol 95:355–361
Mu X, Pang S, Sun X, Gao J, Chen J, Chen X, Li X, Wang C (2013) Evaluation of acute and developmental effects of difenoconazole via multiple stage zebrafish assays. Environ Pollut 175:147–157
Watson FL, Schmidt H, Turman ZK, Hole N, Garcia H, Gregg J, Tilghman J, Fradinger EA (2014) Organophosphate pesticides induce morphological abnormalities and decrease locomotor activity and heart rate in Danio rerio and Xenopus laevis. Environ Toxicol Chem 33:1337–1345
Hsieh DJY, Liao CF (2002) Zebrafish M2 muscarinic acetylcholine receptor: cloning, pharmacological characterization, expression patterns and roles in embryonic bradycardia. Br J Pharmacol 137:782–792
Rahman MS, Islam SM, Haque A, Shahjahan M (2020) Toxicity of the organophosphate insecticide sumithion to embryo and larvae of zebrafish. Toxicol Rep 7:317–323
Çelik ES, Kaya H, Yılmaz S (2012) Effects of phosalone on mineral contents and spinal deformities in common carp (Cyprinus carpio, L. 1758). Turk J Fish Aquat SC 12:259–264
Muramoto S (1981) Vertebral column damage and decrease of calcium concentration in fish exposed experimentally to cadmium. Environ Pollut Ecol Biol 24:125–133
Pereira VM, Bortolotto JW, Kist LW, de Azevedo MB, Fritsch RS, da Luz Oliveira R, Pereira TCB, Bonan CD, Vianna MR, Bogo MR (2012) Endosulfan exposure inhibits brain AChE activity and impairs swimming performance in adult zebrafish (Danio rerio). Neurotoxicology 33:469–475
Tilton FA, Bammler TK, Gallagher EP (2011) Swimming impairment and acetylcholinesterase inhibition in zebrafish exposed to copper or chlorpyrifos separately, or as mixtures. Comp Biochem Physiol C 153:9–16
Pamanji R, Yashwanth B, Bethu M, Leelavathi S, Ravinder K, Rao JV (2015) Toxicity effects of profenofos on embryonic and larval development of zebrafish (Danio rerio). Environ Toxicol Pharmacol 39:887–897
Müller UK, van Leeuwen JL (2004) Swimming of larval zebrafish: ontogeny of body waves and implications for locomotory development. J Exp Biol 207:853–868
Acknowledgements
This research was supported by a grant from the Research Program for Agricultural Science and Technology Development (Project No. PJ01423702), National Institute of Agricultural Sciences, Rural Development Administration, Republic of Korea.
Funding
This research was supported by a grant from the Research Program for Agricultural Science and Technology Development (Project No. PJ01423702), National Institute of Agricultural Sciences, Rural Development Administration, Republic of Korea.
Author information
Authors and Affiliations
Contributions
BMKV, HHP and KC designed the experiments. BMKV and N-SK performed experiments. BMKV and KC analyzed the data. BMKV wrote the manuscript. KC revised the manuscript. All authors read and approved the final manuscript
Corresponding author
Ethics declarations
Competing interests
The authors declare that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1. Fig. S1.
Representative images (96 hpf) showing the effect of EDZ on swim bladder development (red dotted). EDZ inhibited the development of swim bladders in ZF. The ZF were treated with indicated doses of EDZ and the images were captured at 96 hpf. The dotted red line indicates the morphology of the swim bladder.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vasamsetti, B.M.K., Kim, NS., Chon, K. et al. Teratogenic and developmental toxic effects of etridiazole on zebrafish (Danio rerio) embryos. Appl Biol Chem 63, 80 (2020). https://doi.org/10.1186/s13765-020-00566-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13765-020-00566-2