
Abstract
Background
Watersheds dominated by agriculture often have significant, negative impacts on local stream habitats and fauna. Our study examined stream biota and habitats within an entire stream in a karst agricultural landscape to determine if several decades of watershed conservation and management had reversed the severe degradation caused by poor land use and catastrophic soil erosion during the 1920s and 1930s.
Results
Fish communities and stream habitats were degraded at over half of the 22 stream sites examined, and benthic macroinvertebrate communities were mostly poor throughout the entire stream. Poor fish communities were associated with missing or narrow riparian buffers, high percent fine sediments, high embeddedness of coarse substrates, and homogeneous run habitat. Invertebrate communities were dominated by filter-feeding taxa and those adapted to life on or within fine sediments. Stream sites with wide buffers and influenced by groundwater springs had higher fish community integrity and good to excellent instream habitat.
Conclusions
Our results suggest that wide riparian buffers and karstic springs may have allowed recovery of some stream reaches, but more extensive buffers and additional soil conservation practices in upland areas likely are necessary to protect most stream segments and their biota from on-going agricultural activities. New state stream buffer laws instituted after this study resulted in establishment of vegetated buffers at all study sites, which may lead to improved stream habitat and biotic communities in future years. Benthic invertebrate community improvement also may require translocation of sensitive taxa from nearby watersheds due to limited dispersal abilities of many species currently absent from the study stream. Increased frequency and intensity of storm events and flooding may hinder future recovery of stream habitats and biota.
Similar content being viewed by others


Introduction
Streams draining agricultural landscapes can be impacted by eroded soils (Armour et al. 1991; Waters 1995), excessive nutrients (Tesoriero et al. 2009; Dubrovsky and Hamilton 2010), altered hydrology (Freeman et al. 2007; Blann et al. 2009), and numerous herbicides and pesticides (Battaglin and Fairchild 2002; Schäfer et al. 2007). These factors in turn can lead to degraded stream habitats (Davis et al. 2003), changes in productivity (McTammany et al. 2007), reductions or loss of sensitive aquatic taxa (Lenat 1984), and modified food webs (Moss 2008). Impacts can range from local (Nerbonne and Vondracek 2001) to regional (Heitke et al. 2006), even leading to homogenization of habitats and communities across broad geographical areas (Li and Reynolds 1994).
The impacts of agriculture on streams can be influenced greatly by local soils and geology (Reynolds et al. 1986; Troelstrup and Perry 1989; Molenat et al. 2008). In regions with flat topography, precipitation falling on agricultural lands may be filtered and purified by soils or underlying bedrock prior to reaching either aquifers or local streams (Fragalà and Parkin 2010; Hénault-Ethier et al. 2017). In other areas, however, steep topography, thin hydrophobic soils, and/or karst geology may allow agricultural runoff to reach aquifers and streams quickly and unmodified (Troelstrup and Perry 1989; Boyer and Pasquarell 1996, 1999; Lindsey et al. 2010; Danz et al. 2013; Roussiez et al., 2013).
Streams may be insulated from land use effects if vegetated buffers are in place within the riparian zone (Talmage et al. 2002; Vondracek et al. 2005; Yates et al. 2007). Wide, intact forest and grass buffers can protect stream habitats from land use disturbances by intercepting eroded soils in runoff before they reach the stream, and by reducing nutrient concentrations in both surface runoff and subsurface waters (Osborne and Kovacic 1993). Such reductions in potential stream contaminants can allow for development of diverse and high-quality stream communities even in agriculture-dominated drainages (Wang et al. 1997; Fitzpatrick et al. 2001; King et al. 2005). Efforts are accelerating in many areas of the United States to expand the use of vegetated riparian buffers along waterways to better manage soil erosion and nutrient runoff from sensitive agricultural lands (United States Department of Agriculture 2016). For example, the State of Minnesota recently passed a law (Minnesota Statutes 2014, Sect. 103B.101, subdivision 12, as amended in 2016 and 2020; https://www.revisor.mn.gov/statutes/cite/103F.48) that mandates the presence of a continuous, vegetative buffer (perennially rooted vegetation, 15-m average width, 9-m minimum width) between all public waters and cultivated lands. Within vegetated buffers, haying and livestock grazing are permitted activities. To help meet the requirements of that law, the United States Department of Agriculture and the State of Minnesota agreed in 2017 to spend $500 million over 5 years to protect and improve waters in 54 counties primarily by installing > 12,000 hectares of vegetated buffer strips along streams, rivers, and lakes (Minnesota Board of Water and Soil Resources 2017).
The goal of our study was to examine the influence of instream and riparian habitat on aquatic community structure in a Minnesota, USA, stream flowing through an agricultural landscape underlain by karst geology. Specifically, we investigated whether watershed conservation and management practices, implemented beginning in 1941 after historic land misuse due to poor agricultural practices, have protected stream habitats from the impacts of row-crop production and intensive livestock grazing prior to establishment of riparian buffer laws and allowed for development of diverse aquatic communities after past degradation (Waters 1977; Thorn et al. 1997). We also were interested in determining the role of coldwater springs (resulting from the karst geology) in balancing the impacts of agriculture on stream habitats and communities.
Materials and methods
Study area
The Paleozoic Plateau (or Driftless Area) ecoregion of southeastern Minnesota, USA, has been an important agricultural area for > 160 years. Fertile, loess soils overlie ancient (> 100,000 ybp) glacial till (Williams and Vondracek 2010), but the region is most characterized by its rugged karst topography, created when meltwaters from the recent (15,000 ybp) Wisconsinan glaciation deeply dissected the plateau into ridges and steep valleys connecting to the Mississippi River (Fremling 2004). The area’s rich soils, within the transition zone between hardwood forests and tallgrass prairies, supported many decades of grain crops and livestock grazing during the late-1800s, elevating Minnesota as an agricultural producer of worldwide importance (Whitewater River Watershed Project 2015).
Poor land use (e.g., forest removal, plowing steep slopes, continuous cropping, burning and grazing hillsides) in southeastern Minnesota led to catastrophic soil erosion during the 1920s and 1930s (Whitewater River Watershed Project 2015). Impacts on area streams were severe, as soils washed from hillsides and repeatedly buried stream valleys under meters of sand and silt, extirpating native brook trout (Salvelinus fontinalis) from most streams (Thorn et al. 1997). As productivity plummeted, both ridgetop and valley farms were abandoned or sold to the government for a fraction of their former value.
The Whitewater River watershed was severely impacted by soil erosion and flooding, as mud and logging debris from the watershed clogged streams. This 830-km2 drainage suffered through many multi-flood years, the worst in 1938 with 28 flood events (Waters 1977). Trout were eliminated from 60% of the watershed’s 240 km of trout streams, and the remaining 40% were deteriorated to “poor condition” (Waters 1995). Once-prosperous communities of Beaver and Whitewater Falls were abandoned and > 15,000 hectares of land were purchased by the State of Minnesota (Whitewater River Watershed Project 2015). Federal workers were brought in during the mid-1940s to construct erosion control structures and perform forest management work. A complex dike system was constructed to retain runoff and create wildlife habitat while reducing erosion and improving soil health. These conservation measures and better land stewardship produced significant improvements within the drainage basin, but degraded water quality continues to plague the present-day Whitewater River system (Whitewater River Watershed Project 2015).
Since 1987, the Whitewater River Watershed Project has worked with farmers to implement soil conservation practices (e.g., contour farming, strip cropping, conservation tillage, grass waterways, grade stabilization structures and ponds) in upland areas to reduce erosion, increase infiltration, slow runoff, and improve soil quality (Whitewater River Watershed Project 2015). Self-sustaining coldwater fisheries for brown (Salmo trutta) and brook trout within the watershed, benefitting from increased coldwater spring flows from limestone karst, now attract both regional and international anglers (Thorn et al. 1997). Recreational angling is very important to the economy of southeastern Minnesota, contributing at least US$48 million to the regional economy each year (Gartner et al. 2002). However, streams within the watershed remain impacted by agricultural activities; most are impaired for aquatic recreation (due to high fecal coliform bacteria) and aquatic life (due to turbidity; U.S. Environmental Protection Agency Clean Water Act, Section 303(d) listing of impaired waters), with high (> 10 mg/l) nitrate–nitrogen concentrations common throughout the system (Minnesota Pollution Control Agency 2013). Fish and macroinvertebrate communities in this system may be limited further by combinations of warm water temperatures, low dissolved oxygen, and degraded stream habitats (Schauls and Carter 2015). In addition, a major fish kill (nearly 10,000 fish) of undetermined cause(s) occurred during July 2015 on the South Fork Whitewater River (SFWR) shortly after a 5-cm rainfall event (Hunt 2015).
The SFWR drains the southern and eastern portions (241 km2, 29%) of the Whitewater River basin in Olmsted and Winona counties in southeastern Minnesota, USA (Fig. 1), an area dominated by agriculture (49% cropland, 28% livestock pastureland; Minnesota Pollution Control Agency 2013). From its headwaters (387 m above sea level) in the highlands of the Paleozoic Plateau, the SFWR flows 56.5 km to the east and north, descending 168 m through a valley carved by glacial meltwaters to its confluence with the mainstem Whitewater River. In addition to cropland and pastureland, the SFWR watershed includes 110 livestock feedlots that generate 635 metric tons of manure/day, which are applied on watershed croplands at a rate 15 metric tons/hectare/year (Minnesota Pollution Control Agency 2013). The U.S. Interstate 90 highway corridor lies within the southern portion of the watershed.

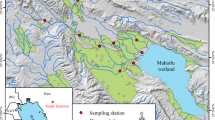
Map of the Whitewater River watershed in southeastern Minnesota, USA. The Mississippi River is located to the upper right. Sampling sites along the South Fork Whitewater River are numbered from upstream to downstream and are indicated by + symbols. The ellipse encloses sites located near natural springs
The entire SFWR is listed as impaired for aquatic recreation and aquatic life (Minnesota Pollution Control Agency 2013). However, 13 coldwater springs enter the SFWR along the lower 21 km (average spring elevation = 277 m above sea level, with a 25% slope; Williams and Vondracek 2010), transitioning the system from coolwater/warmwater to coldwater and creating a popular, self-sustaining fishery for brown trout. The Minnesota Department of Natural Resources maintains Crystal Springs Hatchery at the largest spring (7500 L/min at 9 °C), raising 20,000 kg/year of brook, rainbow (Oncorhynchus mykiss), and lake (Salvelinus namaycush) trout for stocking within the state.
We selected 22 stream sites along the SFWR for habitat and biotic assessments (Fig. 1). We attempted to space sites evenly along the stream, but inaccessibility in some locations (due to topography and private property access restrictions) created some gaps in coverage. Site locations included public and private property, croplands, livestock pastures, and forests. Ultimately, the downstream-most 10 sites were located within the spring-fed section of the SFWR, whereas the 12 upstream sites were located within the river section fed largely by wetland seepage and surface runoff (Fig. 1).
Field work
During the period June to August 2005 (prior to state buffer laws), stream sites were sampled for fish, aquatic macroinvertebrates, and stream habitat. Fish at each site were sampled with one or more backpack electrofishing units (Smith-Root LR-24, 12-B POW, and Type VII) as needed, based on stream size. All fish captured within a 150-m stream reach (downstream to upstream, single pass) were identified, counted, and released. Specimens of uncertain identity were retained for later identification. This sampling procedure was the standard protocol recommended when using a coldwater Index of Biotic Integrity (IBI; Mundahl and Simon 1998) to assess fish communities.
Benthic macroinvertebrates were collected in D-frame net (500 µm mesh) samples at each stream site. Each sample was the combination of organisms collected by disturbing the substrate upstream from the net (0.1-m2 area) for 30 s in each of two sections (fast and slow) of a single riffle. Three such samples were collected at each site, each from a separate riffle. If riffles were absent, collections were made from available coarse (gravel, cobble, boulder) substrates. Invertebrate samples were preserved in ethanol and later sorted, identified (mostly to genus level except for a few dipteran [Chironomidae, Empididae] and non-insect [Hirudinea, Sphaeriidae, Oligochaeta] taxa), and counted in the laboratory. This sampling protocol followed the procedure recommended when using a regional Benthic Index of Biotic Integrity (B-IBI; Wittman and Mundahl 2003; Magner et al. 2008) to assess benthic aquatic invertebrate communities.
Physical habitat at each stream site was examined in two ways. First, a simple fish habitat rating index designed for assessing regional streams (Simonson et al. 1993) was used. Seven instream and riparian features (bank land use, bank stability, pool area, width:depth ratio, riffle-to-riffle distance:stream width ratio, silt substrate abundance, fish cover abundance) were measured, scored, and used to produce a fish habitat rating score (scale = 0 to 100; > 80 = excellent, 60–80 = good, 30–60 = fair, < 30 = poor) for each site. Additional, more detailed habitat assessments also were conducted at each of the sites. These were based on protocols developed or adapted for use regionally (Platts et al. 1983; Simonson et al. 1993; Minnesota DNR 2007). Sampling at each site was conducted using a modification of the transect method (Simonson et al. 1993). Habitats at each site were sampled across each of 12 transects spaced approximately two to three mean stream widths apart.
Along each transect, instream habitat features were assessed, and riparian conditions were recorded. Instream measures were taken at four, evenly spaced points along each transect. At each point, depth was measured using a wading rod, and velocity measured at 0.6-depth (Marsh-McBirney model 2000 flow meter). Dominant substrate composition was estimated visually according to a modification of the Wentworth Scale (clay, silt, sand, gravel, cobble, boulder; Minnesota DNR 2007). Embeddedness, the percent of large substrates such as cobbles covered by fine materials, was visually estimated on a five-category scale: < 5%, 5–25%, 26–50%, 51–75%, and > 75% (Platts et al. 1983).
Other instream measures were estimated visually within a reach one mean stream width in length, centered on the transect (Simonson et al. 1993). The percentage of the stream shaded by the canopy at noon, the percentage of riffle, pool, and run, and the percent of the reach providing cover for fish 200 mm or larger, were estimated to the nearest 5%. Cover for fish included overhanging bank vegetation, woody debris, instream vegetation, boulders, and water > 60 cm deep.
Riparian measures were made on only one stream bank per transect, alternating the side measured with each transect. Width of the riparian buffer was measured to the nearest meter. Average length of vegetation overhanging the stream was measured to the nearest 0.1 m. The percentages of bank vegetation as grass, forb, tree, and shrub, and the percentage of bank as bare soil or rock, were estimated visually to the nearest 5% for each category. Data collected from all transects were averaged to determine overall site values.
Data analyses
Lacking historical data on stream biota prior to stream degradation over a century ago, we assessed fish and aquatic macroinvertebrates in SFWR by calculating community integrity indices for each group and comparing them to regional standards. Fish data were used to calculate a coldwater IBI score and rating for each site (Mundahl and Simon 1998), based on the presence of brown trout throughout most of the SFWR. Twelve fish community metrics (five species richness metrics: total, coldwater specialists, minnows, benthic, tolerant; five percentage metrics: salmonids as brook trout, intolerant individuals, coldwater individuals, white suckers, top carnivores; number of coldwater individuals, number of warmwater individuals) were quantified, scored, and summed to produce a coldwater IBI score for each site (scale = 0 to 120; 105–120 = excellent, 70–100 = good, 35–65 = fair, 10–30 = poor, 0–5 = very poor). These ratings allowed us to compare the fish communities observed at SFWR sites to regional benchmarks (Mundahl and Simon 1998). Using this coldwater IBI, ratings of poor and very poor indicate high to severe impairment, with degraded conditions and trout being very rare or absent. In contrast, fair to excellent ratings indicate reduced impairment and trout abundance ranging from common to abundant.
Benthic invertebrate data were used to calculate regional B-IBI scores and ratings for each stream site (Wittman and Mundahl 2003; Magner et al. 2008). The index was developed based on mostly genus- and family-level identifications (Wittman and Mundahl 2003). Ten invertebrate metrics (seven taxa richness metrics: total, Plecoptera, Trichoptera, Diptera, long-lived, intolerant, filterer; two percentage metrics: predators, long-lived individuals) were quantified, scored, and summed to produce a B-IBI score (scale = 0–100; 65–100 = excellent, 50–60 = good, 30–45 = fair, 10–25 = poor, 0–5 = very poor) for each individual sample. Scores for the triplicate invertebrate samples for each site were averaged to produce a single value and rating for that site. These ratings allowed us to compare the benthic invertebrate communities observed at SFWR sites to regional standards (Wittman and Mundahl 2003; Magner et al. 2008). With this B-IBI, ratings of fair to very poor indicate moderate to severe impairment, with degraded conditions producing reduced taxa richness (especially Ephemeroptera, Plecoptera, and Trichoptera) and increasing dominance by tolerant taxa. In contrast, good to excellent ratings indicate slight or no impairment and high taxa richness (including intolerant or sensitive taxa).
We used non-metric multidimensional scaling (NMDS; JMP Pro 12 software, SAS Institute Inc. 2015; open-source software package R with additional packages [dyplyr, readxl, vegan, ggplot2], R Core Team 2020) on the fish and benthic invertebrate count data (individuals per taxon for all taxa observed for all sites) and the stream habitat variables to create ordination plots visualizing the level of community similarity and potential site clustering among the 22 SFWR stream sites. Separate fish-habitat and benthic invertebrate-habitat NMDS models were constructed to determine if the different taxa types produced similar site clustering. We used linear fit (r2) to compare each of the 18 physical habitat variables and all of the fish and benthic invertebrate taxa individually to each NMDS model as a whole, to assess which variables were most influential in structuring potential site clusters. PERMANOVA (vegan package in R; P values based on 999 permutations; Oksanen et al. 2020) was used to compare assemblages between stream site clusters.
In a different approach, we used a model selection procedure to evaluate the influence of stream and riparian habitat on biotic community integrity in the SFWR. Specifically, we explored a series of simple models separately comparing fish IBI and macroinvertebrate B-IBI scores to a suite of stream habitat variables (9 in-stream variables, 9 riparian variables). For simplicity, we focused only on those potential models that included five or fewer habitat variables. The explanatory power of these possible competing models was compared using Akaike’s Information Criterion for small sample sizes (AICc; Burnham and Anderson 2002; Brown and Guy 2007). We used JMP Pro 12 (SAS Institute Inc. 2015) to examine and evaluate these models. Only those explanatory models with Akaike weights (wi) ≥ 10% of the wi of the best model were interpreted (Burnham and Anderson 2002). Explanatory powers of competing models were calculated by dividing the wi value of the best model by that of the competing model (Burnham and Anderson 2002).
Finally, we used a variety of simple statistical tests to address additional study questions. Simple linear regression was used to examine potential cause–effect relationships between various fixed and response variables. These included the potential effects of buffer width on instream habitat variables typically indicative of habitat quality (fish habitat rating, % fines, embeddedness, % run habitat), and also the potential effects of these habitat variables on typical measures of stream biotic integrity (coldwater fish IBI, benthic IBI, % Ephemeroptera–Plecoptera–Trichoptera [EPT]). We also used Student’s t tests to compare habitat variables (fish habitat rating index, buffer width, % fines, substrate embeddedness, % run habitat) and biotic metrics (coldwater fish IBI score, fish taxa richness, benthic IBI score, % EPT) between groups of sites in stream reaches with and without groundwater springs.
Results
Habitats
Both instream and riparian habitats varied among the SFWR sites examined (Additional file 1: Tables S1 and S2). Upstream sites flowed through cattle pastures and along crop fields, with fine bottom sediments, homogeneous run habitats, and grassy banks often lacking buffers. Downstream sites flowed through forests and pastures, had greater widths, wider buffers, improved habitat diversity (more pools and riffles), more fish cover, faster current velocities, and coarser bottom substrates. Fish habitat ratings at upstream sites 1 through 12 (non-spring sites) ranged from poor to good, whereas at downstream sites 13 through 22 (spring-fed sites) ratings ranged from fair to excellent (Additional file 1: Table S3).
Buffer width was statistically (α = 0.05) correlated with important instream habitat variables that measure habitat quality. Specifically, there was a significant, positive relationship between buffer width and the fish habitat rating index (simple linear regression: fish habitat rating index = 0.17 * buffer width + 40.7, F1,20 = 16.3, P < 0.001, r2 = 0.45). In addition, we observed significant, negative relationships between buffer width and % fines (simple linear regression: % fines = − 0.18 * buffer width + 60.9, F1,20 = 7.81, P = 0.01, r2 = 0.28), and between buffer width and embeddedness (simple linear regression: embeddedness = − 0.006 * buffer width + 4.82, F1,20 = 9.09, P = 0.006, r2 = 0.31). However, there was no significant relationship between buffer width and % run habitat (simple linear regression: F1,20 = 1.00, P = 0.330, r2 = 0.05).
Fish communities
Eighteen species of fishes were represented among the 9167 individuals collected from the SFWR (Table 1). White sucker and longnose dace were most frequently encountered, collected at 21 and 19 sites, respectively. Most species (10) were found at half or fewer of the stream sites. Slimy sculpin was the most abundant fish, representing > 31% of all fish collected, but it was found at fewer than one-third of the sites (downstream sites 14 through 19). Only six of the 18 species were represented by < 100 individuals. Species richness at the spring-fed downstream sites (mean ± SD; 5.0 ± 1.7 species/site) was half that observed at non-spring upstream sites (10.4 ± 2.5 species/site).
Fish IBI ratings generally were very poor at upstream sites, becoming fair to good at downstream sites (Additional file 1: Table S3). Model selection produced three simple, competing models (with Akaike weights [wi] ≥ 10% of the wi of the best model) that best explained the relationship between instream and riparian variables and fish IBI scores (Table 2). The best model contained four instream and riparian variables (% fish cover, buffer width, % bank forbs, % bank rocks) that together explained > 87% of the variation in fish IBI scores. Individually, fish IBI scores were statistically (α = 0.05) correlated to both buffer width (F1,20 = 11.39, P = 0.003, r2 = 0.33) and fish habitat rating score (F1,20 = 19.66, P < 0.001, r2 = 0.50).
Aquatic macroinvertebrates
We collected > 13,500 aquatic macroinvertebrates representing 39 taxa from SFWR sites (Table 3). Midges were found at every site but represented < 7% of all individuals collected. Caddisflies (Trichoptera), mayflies (Ephemeroptera), beetles (Coleoptera), and amphipods (Gammarus) were the other common taxa encountered at many sites in large numbers. Most common taxa were either filter-feeders or those adapted to living on or within fine sediments. Densities (1027 ± 753 organisms/m2, grand mean ± SD) were highly variable among sites but increased statistically (α = 0.05) (simple linear regression: F1,20 = 12.9, P = 0.002) from upstream to downstream (river km as X variable, density as Y variable). The percentage of individuals that were EPT taxa also increased statistically (α = 0.05) (simple linear regression: F1,20 = 18.8, P < 0.001) from upstream to downstream sites. In contrast, total invertebrate taxa richness was low (grand mean = 14 ± 3 taxa/site) and did not change statistically (α = 0.05) throughout the river system (simple linear regression: F1,20 = 0.003, P = 0.96).
Benthic IBI ratings generally were poor or fair throughout the entire stream system, with no obvious pattern of change throughout the river system (Additional file 1: Table S3). Model selection produced five simple, competing models (with Akaike weights [wi] ≥ 10% of the wi of the best model) that best explained the relationship between instream and riparian variables and benthic IBI scores (Table 2). The best model contained four instream and riparian variables (% bank grass, % fish cover, % pool, % run) that together explained 60% of the variation in benthic IBI scores in the SFWR (Table 2). Neither benthic IBI scores nor EPT relative abundance (%) were statistically (α = 0.05) correlated (simple linear regression) with any of the expected measures of stream habitat quality (buffer width, fish habitat rating score, % fines, embeddedness, or % run habitat). Fish IBI scores were not correlated with either benthic IBI scores (r2 = 0.010) or EPT relative abundance (r2 = 0.035).
NMDS ordination
NMDS ordination of stream sites based on abundances of fish and invertebrate taxa and stream habitat variables produced two models that adequately (i.e., stress < 0.20) positioned stream sites in two-dimensional space (Figs. 2 and 3). Abundances of 10 species of fish and seven stream habitat variables were correlated statistically (α = 0.05) with the overall fish-habitat NMDS model (Fig. 2, Table 4). Similarly, eight taxa of benthic macroinvertebrates and four stream habitat variables were correlated statistically (α = 0.05) with the invertebrate-habitat NMDS model (Fig. 3, Table 4). Both fish-habitat and invertebrate-habitat NMDS models separated stream sites into two clusters: upstream sites (sites 1 through 12) and downstream sites (sites 13 through 22; Figs. 2 and 3). Although clusters of upstream and downstream sites exhibited greater visual overlap in the invertebrate-habitat model than in the fish-habitat model, PERMANOVA indicated that cluster centroids and dispersion of the clusters differed statistically (α = 0.05) in both models (Table 5).

Non-metric multidimensional scaling ordination (NMDS) biplots of 22 stream sites on the South Fork Whitewater River, Minnesota, USA, based on abundances of fish and stream habitat variables at each site. Numbers represent sites (see Fig. 1), separate ellipses enclose the 12 upstream sites and the 10 downstream sites, and arrows represent locations of greatest abundance for fish (A) and strongest influence of stream habitat variables (B) whose values were statistically (α = 0.05) correlated with the complete NMDS model (see Table 4)

Non-metric multidimensional scaling ordination (NMDS) biplots of 22 stream sites on the South Fork Whitewater River, Minnesota, USA, based on abundances of benthic invertebrates and stream habitat variables at each site. Numbers represent sites (see Fig. 1), separate ellipses enclose the 12 upstream sites and the 10 downstream sites, and arrows represent locations of greatest abundance for invertebrates (A) and strongest influence of stream habitat variables (B) whose values were statistically (α = 0.05) correlated with the complete NMDS model (see Table 4)
By referencing both NMDS models, upstream sites were characterized by high abundances of five species of cyprinids, johnny darter, and white sucker and low abundances of trout, sculpin, Baetis mayflies, Hydropsyche caddisflies, and several dipteran taxa (Figs. 2 and 3). In contrast, downstream sites were characterized by high abundances of trout, sculpin, Baetis mayflies, Hydropsyche caddisflies, and several dipteran taxa and low abundances of cyprinids, suckers, and darters. Both models indicated that downstream sites had greater widths, greater % riffle habitat, and wider buffers compared to upstream sites. In general, more stream habitat variables exhibited stronger correlation with the fish-habitat NMDS model than with the invertebrate-habitat NMDS model (Table 4).
Proximity to springs
Spring-fed stream sites (sites 13 through 22) differed statistically (α = 0.05) from sites without springs (sites 1 through 12) in several ways. Fish IBI scores were higher at spring-fed sites than at non-spring locations; benthic IBI scores did not differ among sites based on spring proximity, but EPT relative abundance was statistically (α = 0.05) higher at spring-fed sites (Table 6). Fish habitat rating scores and buffer widths were greater at spring-fed sites, whereas fine substrate relative abundance, substrate embeddedness, and relative abundance of run habitats all were greater at non-spring sites (Table 6).
Discussion
Our study of the habitats and biota of the SFWR, draining a largely agricultural basin, produced four important findings. First, large sections of the stream were impaired by sedimentation, with riparian buffer width directly related to instream habitat quality. Second, better fish community structure was associated with wider buffers and improved instream habitats. Third, benthic community structure generally was fair or poor throughout the stream, and not influenced significantly by either riparian or instream habitats. Finally, stream habitats and fish communities were better at stream sites influenced by springs, although these sites also benefitted from wider buffers than non-spring sites.
Sedimentation and degraded water quality were evident in the SFWR during 2005, especially for sites surrounded by intensive agricultural activities. Only 12 of the 22 sites examined had riparian buffers that would meet current (2021) state standards for public waters (i.e., perennially rooted vegetation, 15-m average width, 9-m minimum width). This study was conducted prior to establishment of stream buffer laws in Minnesota, but an examination of study sites in 2019 indicated that all 22 sites are now in compliance with state buffer laws (average buffer width ≥ 15 m). In previous studies, inadequate buffers have allowed eroded soils and agricultural chemicals to enter stream systems, smothering coarse substrates and filling pools to create homogeneous run habitats (Armour et al. 1991; Osborne and Kovacic 1993; Nerbonne and Vondracek 2001) and altering stream productivity (McTammany et al. 2007; Piscart et al. 2009). These conditions have been reported in streams draining agricultural lands elsewhere in Minnesota (Troelstrup and Perry 1989; Nerbonne and Vondracek 2001; ZumBerge et al. 2003), the United States (Walser and Bart 1999; Agouridis et al. 2005), and other countries (Soulsby et al. 2001; Matthaei et al. 2010; Burdon et al. 2013; Teufl et al. 2013). When vegetated buffers are inadequate or lacking, there usually are strong relationships between the proportion of agricultural lands within a drainage and instream physical and chemical variables (Lammert and Allan 1999; Walser and Bart 1999; Fitzpatrick et al. 2001; Davis et al. 2003; King et al. 2005).
Both the type of buffer present (e.g., grass versus trees) and the width of the buffer are important in protecting streams from adjacent cultivated and grazed lands. Grassy riparian areas can be more effective than wooded or brushy areas at filtering eroded soils and nutrients from overland flows, and at preventing bank erosion (Barfield et al. 1998; Lyons et al. 2000; Vigiak et al. 2007). Grassy buffers at SFWR sites usually were associated with well-vegetated banks with little bare soil, whereas wooded buffers often had high relative abundance (> 30%) of streambank bare soil, similar to levels found in areas lacking buffers. A previous study in this same watershed also reported considerable bare soil within wooded buffers, especially beneath riparian stands of box elder (Acer negundo; Nerbonne and Vondracek 2001). Although less effective as filters, forested buffers can contribute branches or fallen trees that modify channel morphology and improve habitat complexity, often keeping coarse substrates from becoming embedded by flushing away fine sediments (Angermeier and Karr 1984; Talmage et al. 2002).
Buffer width can be critical in protecting streams from agricultural lands (Barfield et al. 1998; Nerbonne and Vondracek 2001; Vondracek et al. 2005), and buffer width was strongly correlated with better fish habitat ratings and higher fish IBI scores in the SFWR. Current Minnesota state law mandates an average buffer width of 15 m on all public waterways (excluding ditches). Although buffers < 15 m wide can be capable of filtering > 90% of sediments and chemicals from surface runoff (Barfield et al. 1998), much wider buffers may be necessary in many areas (Davis et al. 2003; Snyder et al. 2003; Vondracek et al. 2005). Buffer effectiveness frequently is determined by many site variables in addition to width (Johnson and Buffler 2008). For example, greater buffer slope, high degree of soil saturation, reduced surface roughness, poor soil permeability, and other variables can limit the ability of buffers to retain both sediments and nutrients (Barfield et al. 1998; Graff et al. 2005; Mander and Hayakawa 2005; Johnson and Buffler 2008). Consequently, a “one-size-fits-all” buffer width in Minnesota may not provide adequate protection for streams in many of the state’s agricultural areas (Johnson and Buffler 2008).
Fish communities in agricultural streams depend on intact riparian buffers that protect instream habitats from degradation (Talmage et al. 2002; Vondracek et al. 2005). In systems lacking adequate buffers, fish communities suffer due to reduced water quality, habitat siltation, reduced cover, and less stable hydrology and water temperatures (Fitzpatrick et al. 2001; Nerbonne and Vondracek 2001; Blann et al. 2002; Snyder et al. 2003). As in our study in the SFWR, fish IBI scores usually are correlated positively with buffer width and/or negatively with instream habitat variables (fine sediments, embeddedness, run habitats) that are indicative of poor buffers (Walser and Bart 1999; Fitzpatrick et al. 2001; Nerbonne and Vondracek 2001; Heitke et al. 2006). In the SFWR, the association of good fish communities with wide, intact riparian buffers emphasizes what buffers can accomplish even in watersheds dominated by agricultural activities.
Benthic macroinvertebrate communities in the SFWR were consistently poor throughout the system, and not correlated with riparian, instream, or fish community variables. Most invertebrate taxa were filter-feeders and or adapted to life on or within fine sediments (see feeding strategies and habitats described throughout Merritt and Cummins 1996). It is widely recognized that agriculture can affect stream macroinvertebrates directly via toxic runoff of various pesticides, herbicides, and fungicides (Lenat 1984; Troelstrup and Perry 1989; Helfrich et al. 2009) and indirectly via enhanced algal (food) production from nutrients (Matthaei et al. 2010) and habitat degradation from sedimentation (Waters 1995; Burdon et al. 2013). Several studies have reported significant relationships between various macroinvertebrate indexes (e.g., benthic IBI) or metrics (e.g., EPT measures) and agricultural land use (Troelstrup and Perry 1989; Davis et al. 2003; ZumBerge et al. 2003; King et al. 2005; Burdon et al. 2013), but others have reported few or inconsistent relationships (Sovell et al. 2000; Fitzpatrick et al. 2001; Nerbonne and Vondracek 2001; this study). Similarly, fish indexes can be good or poor at detecting the effects of agriculture on stream ecosystems (e.g., Lammert and Allan 1999; Sovell et al. 2000; Fitzpatrick et al. 2001), possibly the result of differing responses among species to land use versus instream (watershed scale versus local scale) habitat measures (Plafkin et al. 1989; Lammert and Allan 1999).
Macroinvertebrate taxa richness generally was low (average = 14 taxa/site, 39 total taxa) in the SFWR, with EPT representing 30% of taxa and 60% of total individuals. Even in downstream areas with improved habitat, invertebrate taxa richness and community structure remained poor, although EPT relative abundance averaged 50% higher at downstream sites than at upstream sites. During the historical degradation of the SFWR in the early 1900s (Waters 1995; Thorn et al. 1997), many macroinvertebrate taxa likely were eliminated along with intolerant fish species. When stream habitats later improved, fish community recovery benefited from early salmonid stocking, subsequent recolonization of stream reaches, and natural reproduction (Thorn et al. 1997), and colder water temperatures (caused by improved spring discharges) became less supportive of warmwater- and coolwater-adapted species (high-quality coldwater sites have fewer fish species than high-quality warmwater sites; Mundahl and Simon 1998). However, macroinvertebrate community recovery was left to natural mechanisms of downstream drift and adult flight dispersal (Waters 2000; Trekels et al., 2011; Petersen et al. 2004; Mundahl and Hunt 2011). Lacking any upstream or interstitial refugia that escaped degradation (Williams and Hynes 1976; Angradi 1997), and isolated from other streams (Collier et al. 2002) by tall, wooded bluffs, recovery of macroinvertebrate communities in the lower SFWR has been hindered, despite improved instream habitats. Streams in watersheds immediately adjacent (Zumbro River to the northwest, Root River to the south) to the Whitewater River have more diverse benthic invertebrate communities with greater EPT taxa richness compared to those in the SFWR and the other two forks (Middle and North) of the Whitewater River (N. Mundahl, unpublished data), but they have been unable to colonize the SFWR. A similar lack of dispersal by benthic macroinvertebrate taxa also has limited the success of stream restoration efforts in Finland (Louhi et al. 2011). Short of reintroducing additional invertebrate taxa by translocation from other systems (Grégoire Taillefer and Wheeler 2013) into the lower reaches of the SFWR, where stream habitats are better than in upstream reaches, macroinvertebrate communities will remain unrepresentative of habitat quality within this downstream reach. Only the greater densities of benthic invertebrates observed at downstream sites in the SFWR are indicative of improved habitat conditions (Dolph et al. 2015).
The question remains whether state-mandated, vegetated buffers along all stream reaches, coupled with existing soil conservation practices within the agricultural watershed, will lead to improved instream habitats and better fish and benthic invertebrates throughout the entire SFWR. Both upland land use and localized habitat conditions affect stream fish and benthic invertebrate community structure and abundance (Lammert and Allan 1999; Heitke et al. 2006; Dolph et al. 2015). However, numerous studies indicate that small-scale restoration projects or localized instream and or riparian habitat restorations often fail to successfully improve biotic communities or stream habitat to desirable levels (e.g., Moerke et al. 2004; Alexander and Allan 2007; Louhi et al. 2011; Ernst et al. 2012; Kroll et al. 2019), as the area(s) of restored habitat may simply be too small compared to the size of nearby, degraded habitats (Palmer 2009). Conservation practices applied over greater spatial areas (i.e., watershed-wide), which collectively comprise significant proportions of total watershed area, often are necessary to produce observable improvements in both stream habitats and biotic communities (Wang et al. 2002; Marshall et al. 2008; Palmer 2009; Richardson et al. 2011). Consequently, complete recovery of aquatic habitats and communities in the SFWR likely will require both upland and riparian restoration efforts. Buffer law compliance within the watershed presently exceeds 99%, but upland soil conservation practices are far less commonly implemented (23% of agricultural lands within the watershed utilize soil conservation practices in 2021) within the SFWR drainage despite many years of encouragement and governmental monetary assistance (Daryl Buck, district manager, Winona County Soil and Water Conservation District, personal communication). More widespread implementation of conservation tillage and other soil conservation practices may be needed, especially in the upper portions of the SFWR watershed, to facilitate improvement in stream habitat and fish and benthic invertebrate communities.
The frequency and intensity of storm events within the SFWR watershed, and the southeastern Minnesota region in general, have increased in recent decades (Minnesota Interagency Climate Adaptation Team 2017), and flash floods associated with these storm events can significantly alter stream habitats and impact fish and benthic invertebrate communities for months or years at a time (Mundahl and Hunt 2011; Mundahl 2017). Such continuing disturbance, especially one that differs from the historic disturbance regime within which stream communities evolved and adapted, can disrupt metacommunity dynamics (e.g., dispersal) within stream networks (Holyoak et al. 2020). Temporal variability in dispersal within metacommunities could lead to frequent shifts in both community structure and function (Datry et al. 2016), further compromising development of a self-sustaining stream community and offsetting any mitigation efforts being undertaken to improve stream habitats and communities (Holyoak et al. 2020).
Stream habitat and fish communities were better in the lower portion of the SFWR, where wider riparian buffers and groundwater springs were present. Buffers and springs both likely influenced habitat and fish communities within this reach via reduced sedimentation, increased discharges (> 6X greater than at upstream sites) that helped to clean coarse substrates, and colder summer water temperatures (shading by riparian trees, groundwater discharges at 9 °C). Karst geology throughout the region contains many aquifers that connect with the land surface in a mix of conduit and diffuse springs (Troelstrup and Perry 1989). Most springs are associated with the St. Lawrence–Franconia–Ironton–Galesville aquifer group, which becomes exposed at elevations of 230 to 290 m above sea level within the lower stream valleys (Williams and Vondracek 2010). These aquifers, which receive recharge waters at a high rate (15–25 cm/year; Delin and Falteisek 2007) from upland agricultural lands, are the likely source of nitrates that impair lower sections of the SFWR (Dubrovsky and Hamilton 2010; Minnesota Pollution Control Agency 2013). These nutrient-rich spring waters support extensive macrophyte beds in non-shaded stream sections (N. Mundahl, personal observation), and spring discharge volumes have increased substantially since the 1930s due to increased precipitation and better infiltration resulting from improved conservation practices on agricultural lands (Thorn et al. 1997). Although karst geology in agricultural regions can impair streams by bypassing natural filtration systems for nutrients, bacteria, and other chemicals (Pasquarell and Boyer 1995; Boyer and Pasquarell 1996, 1999; Lindsey et al. 2010), high spring discharges and intact riparian buffers in the lower SFWR appear to have counteracted many of the negative effects of agriculture observed in the upper reaches and may have allowed for recovery of both stream habitat and fish communities.
Conclusions
Stream habitats and biota in many portions of the SFWR are impacted by on-going agricultural activities, even though watershed conservation and management activities have been conducted within the drainage basin since the 1940s. With 77% of the watershed dedicated to crop and livestock production, upland soil conservation practices and intact riparian buffers are needed along the SFWR to reduce sedimentation and absorb nutrients and agricultural chemicals. Inadequate buffers were correlated with poor instream habitats (high embeddedness and % fines), low habitat diversity (high % run habitat), and poor fish and macroinvertebrate communities. In areas, where wide buffers were present and groundwater springs discharged large volumes of cold water, good stream habitats supported good, coldwater fish communities, although macroinvertebrate communities remained poor. Additional and more widespread implementation of soil conservation practices throughout the watershed may be necessary to reduce or eliminate the impact of on-going agricultural activities on the stream environment. Buffers and springs may have allowed downstream reaches and fish communities to recover from past degradation, but it may require macroinvertebrate reintroductions or translocations to restore benthic communities due to lack of dispersal from other potential source populations and communities. In the future, stream sites in the upper reaches of the SFWR should be reassessed to determine if the presence of buffers put in place in response to state law, plus the implementation of additional upland soil conservation practices, have reduced the impacts of agricultural activities on stream habitats and biota.
Availability of data and materials
The data sets used during the current study are available from the corresponding author on reasonable request.
Abbreviations
- B-IBI:
-
Benthic Index of Biotic Integrity
- DNR:
-
Department of Natural Resources
- EPT:
-
Ephemeroptera–Plecoptera–Trichoptera
- IBI:
-
Index of Biotic Integrity
- NMDS:
-
Non-metric Multidimensional Scaling
- PERMANOVA:
-
Permutational Multivariate Analysis of Variance
- SD:
-
Standard deviation
- SFWR:
-
South Fork Whitewater River
- USA:
-
United States of America
References
Agouridis CT, Workman SR, Warner RC, Jennings GD (2005) Livestock grazing management impacts on stream water quality: a review. J Am Water Resour Assoc 41(3):591–606
Alexander GG, Allan JD (2007) Ecological success in stream restoration: case studies from the midwestern United States. Environ Manag 40(2):245–255
Angermeier PL, Karr JR (1984) Relationship between woody debris and fish habitat in a small warmwater stream. Trans Am Fish Soc 113(6):716–726
Angradi TR (1997) Hydrologic context and macroinvertebrate community response to floods in an Appalachian headwater stream. Am Midl Nat 138(2):371–386
Armour CL, Duff DA, Elmore W (1991) The effects of livestock grazing on riparian and stream ecosystems. Fisheries 16(1):7–11
Barfield BJ, Blevins RL, Fogle AW, Madison CE, Inamdar S, Carey DI, Evangelou VP (1998) Water quality impacts of natural filter strips in karst areas. Trans Am Soc Agric Eng 41:371–381
Battaglin W, Fairchild J (2002) Potential toxicity of pesticides measured in Midwestern streams to aquatic organisms. Water Sci Technol 45(9):95–103
Blann K, Nerbonne JF, Vondracek B (2002) Relationship of riparian buffer type to water temperature in the Driftless Area Ecoregion of Minnesota. N Am J Fish Manag 22(2):441–451
Blann KL, Anderson JL, Sands GR, Vondracek B (2009) Effects of agricultural drainage on aquatic ecosystems: a review. Crit Rev Environ Sci Technol 39(11):909–1001
Boyer DG, Pasquarell GC (1996) Agricultural land use effects on nitrate concentrations in a mature karst aquifer. J Am Water Resour Assoc 32(3):565–573
Boyer DG, Pasquarell GC (1999) Agricultural land use impacts on bacterial water quality in a karst groundwater aquifer. J Am Water Resour Assoc 35(2):291–300
Brown ML, Guy CS (2007) Science and statistics in fisheries research. In: Guy CS, Brown ML (eds) Analysis and interpretation of freshwater fisheries data. American Fisheries Society, Bethesda, p 1
Burdon FJ, McIntosh AR, Harding JS (2013) Habitat loss drives threshold response of benthic invertebrate communities to deposited sediment in agricultural streams. Ecol Appl 23(5):1036–1047
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretical approach. Springer-Verlag, New York
Collier K, Parkyn S, Quinn J, Scarsbrook M (2002) Bouncing back: how fast can stream invertebrates recolonize? Water Atmosphere 10(2):9–11
Danz ME, Corsi SR, Brooks WR, Bannerman RT (2013) Characterizing response of total suspended solids and total phosphorus loading to weather and watershed characteristics for rainfall and snowfall events in agricultural watersheds. J Hydrol 507:249–261
Datry T, Bonada N, Heino J (2016) Towards understanding the organisation of metacommunities in highly dynamic ecological systems. Oikos 125(2):149–159
Davis S, Golladay SW, Vellidis G, Pringle CM (2003) Macroinvertebrate biomonitoring in intermittent coastal plain streams impacted by animal agriculture. J Environ Qual 32(3):1036–1043
Delin GN, Falteisek JD (2007) Groundwater recharge in Minnesota. U.S. Geological Survey, Minnesota Water Science Center Fact Sheet 2007–3002. Mounds View, Minnesota
Dolph CL, Eggert SL, Magner J, Ferrington LC Jr, Vondracek B (2015) Reach-scale stream restoration in agricultural streams of southern Minnesota alters structural and functional responses of macroinvertebrates. Freshw Sci 34(2):535–546
Dubrovsky NM, Hamilton PA (2010) Nutrients in the nation’s streams and groundwater: national findings and implications. U.S. Geological Survey Fact Sheet 2010–3078
Ernst AG, Warren DR, Baldigo BP (2012) Natural-channel-design restorations that changed geomorphology have little effect on macroinvertebrate communities in headwater streams. Restor Ecol 20(4):532–540
Fitzpatrick FA, Scudder BC, Lenz BB, Sullivan DJ (2001) Effects of multi-scale environmental characteristics on agricultural stream biota in eastern Wisconsin. J Am Water Resour Assoc 37(6):1489–1507
Fragalà FA, Parkin G (2010) Local recharge processes in glacial and alluvial deposits of a temperate catchment. J Hydrol 389(1–2):90–100
Freeman MC, Pringle CM, Jackson CR (2007) Hydrologic connectivity and the contribution of stream headwaters to ecological integrity at regional scales. J Am Water Resour Assoc 43(1):5–14
Fremling CR (2004) Immortal river: the Upper Mississippi River in ancient and modern times. University of Wisconsin Press, Madison
Gartner W, Love L, Erkkila D, Fulton D (2002) Economic impact and social benefits study of coldwater angling in Minnesota. University of Minnesota Extension Service, St. Paul
Graff CD, Sadeghi AM, Lowrance RR, Williams RG (2005) Quantifying the sensitivity of the riparian ecosystem management model (REMM) to changes in climate and buffer characteristics common to conservation practices. Trans Am Soc Agric Eng 48(4):1377–1387
Grégoire Taillefer A, Wheeler TA (2013) Animal colonization of restored peatlands: inoculation of plant material as a source of insects. Restor Ecol 21(1):140–144
Heitke JE, Pierce CL, Gelwicks GT, Simmons GA (2006) Habitat, land use, and fish assemblage relationships in Iowa streams: preliminary assessment in an agricultural landscape. Am Fish Soc Symp 48:287–303
Helfrich LA, Weigmann DL, Hipkins P, Stinson ER (2009) Pesticides and aquatic animals: a guide to reducing impacts on aquatic systems. Virginia Polytechnic Polytechnic and State University, College of Agriculture and Life Sciences Publication 420-013, Blacksburg
Hénault-Ethier L, Larocque M, Perron R, Wiseman N, Labreque M (2017) Hydrological heterogeneity in agricultural riparian buffer strips. J Hydrol 546:276–288
Holyoak M, Caspi T, Redosh LW (2020) Integrating disturbance, seasonality, multi-year temporal dynamics, and dormancy into the dynamics and conservation of metacommunities. Front Ecol Evol 8:571130. https://doi.org/10.3389/fevo.2020.571130
Hunt L (2015) South Branch Whitewater River unified fish kill response 2015. Minnesota Department of Agriculture, St Paul
Johnson CW, Buffler S (2008) Riparian buffer design guidelines for water quality and wildlife habitat functions on agricultural landscapes in the Intermountain West. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station General Technical Report RMRS-GTR-203
King RS, Baker ME, Whigham DF, Weller DE, Jordan TE, Kazyak PF, Hurd MK (2005) Spatial considerations for linking watershed land cover to ecological indicators in streams. Ecol Appl 15(1):137–153
Kroll SA, Horwitz RJ, Keller DH, Sweeney BW, Jackson JK, Perez LB (2019) Large-scale protection and restoration programs aimed at protecting stream ecosystem integrity: the role of science-based goal-setting, monitoring, and data management. Freshw Sci 38(1):23–39
Lammert M, Allan JD (1999) Assessing biotic integrity of streams: effects of scale in measuring the influence of land use/cover and habitat structure on fish and macroinvertebrates. Environ Manag 23(5):257–270
Lenat DR (1984) Agriculture and stream water quality: a biological evaluation of erosion control practices. Environ Manag 8(4):333–343
Li H, Reynolds JF (1994) A simulation experiment to quantify spatial heterogeneity in categorical maps. Ecology 75(8):2446–2455
Lindsey BD, Katz BG, Berndt MP, Ardis AF, Skach KA (2010) Relations between sinkhole density and anthropogenic contaminants in selected carbonate aquifers in the eastern United States. Environ Earth Sci 60(5):1073–1090
Louhi P, Mykrä H, Paavola R, Huusko A, Vehanen T, Mäki-Petäys A, Muotka T (2011) Twenty years of stream restoration in Finland: little response by benthic macroinvertebrate communities. Ecol Appl 21(6):1950–1961
Lyons J, Trimble SW, Paine LK (2000) Grass versus trees: managing riparian areas to benefit streams of central North America. J Am Water Resour Assoc 36(4):919–930
Magner JA, Vondracek B, Brooks KN (2008) Grazed riparian management and stream channel response in southeastern Minnesota (USA) streams. Environ Manag 42(3):377–390
Mander U, Hayakawa Y (2005) Purification processes, ecological functions, planning and design of buffer zones in agricultural watersheds. Ecol Eng 24(5):421–432
Marshall DW, Fayram AH, Panuska JC, Baumann J, Hennessy J (2008) Positive effects of agricultural land use changes on coldwater fish communities in southwest Wisconsin streams. N Am J Fish Manag 28(3):944–953
Matthaei CD, Piggott JJ, Townsend CR (2010) Multiple stressors in agricultural streams: interactions among sediment addition, nutrient enrichment and water abstraction. J Appl Ecol 47(3):639–649
McTammany ME, Benfield EF, Webster JR (2007) Recovery of stream ecosystem metabolism from historical agriculture. J North Am Benthological Soc 26(3):532–545
Merritt RW, Cummins KW (1996) An introduction to the aquatic insects of North America, 3rd edn. Kendall/Hunt, Dubuque
Minnesota Board of Water and Soil Resources (2017) The Minnesota Conservation Reserve Enhancement Program (MN CREP). Minnesota Board of Water and Soil Resources, St Paul
Minnesota Department of Natural Resources (2007) Fisheries stream survey manual: stream survey methods. Minnesota Department of Natural Resources, Division of Fish and Wildlife, Section of Fisheries Special Publication No. 165, St. Paul
Minnesota Interagency Climate Adaptation Team (2017) Adapting to climate change in Minnesota. Minnesota Pollution Control Agency, St. Paul. https://www.pca.state.mn.us/sites/default/files/p-gen4-07c.pdf
Minnesota Pollution Control Agency (2013) Mississippi River (Winona) watershed monitoring and assessment report. Minnesota Pollution Control Agency, St. Paul
Moerke AH, Gerard KJ, Latimore JA, Hellenthal RA, Lamberti GA (2004) Restoration of an Indiana, USA, stream: bridging the gap between basic and applied lotic ecology. Freshw Sci 23(3):647–660
Molenat J, Gascuel-Odoux C, Ruiz L, Gruau G (2008) Role of water table dynamics on stream nitrate export and concentration in agricultural headwater catchment (France). J Hydrol 348(3–4):363–378
Moss B (2008) Water pollution by agriculture. Philos Trans R Soc b: Biol Sci 363(1491):659–666
Mundahl ND (2017) Population dynamics of brown trout in a Minnesota (USA) stream: a 25-year study. River Res Appl 133(8):1235–1245
Mundahl ND, Hunt AM (2011) Recovery of stream invertebrates after catastrophic flooding in southeastern Minnesota, USA. J Freshw Ecol 26(4):445–457
Mundahl ND, Simon TP (1998) Development and application of an index of biotic integrity for coldwater streams of the upper midwestern United States. In: Simon TP (ed) Assessing the sustainability and biological integrity of water resource quality using fish assemblages. Lewis, Boca Raton, p 383
Nerbonne BA, Vondracek B (2001) Effects of land use on benthic macroinvertebrates and fish in the Whitewater River, Minnesota. Environ Manag 28(1):87–99
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) vegan: Community ecology package. R package version 2.5-7
Osborne LL, Kovacic DA (1993) Riparian vegetated buffer strips in water-quality restoration and stream management. Freshw Biol 29:243–258
Palmer MA (2009) Reforming watershed restoration: science in need of application and applications in need of science. Estuar Coast 32(1):1–17
Pasquarell GC, Boyer DG (1995) Agricultural impacts on bacterial water quality in karst groundwater. J Environ Qual 24(5):959–969
Petersen I, Masters Z, Hildrew AG, Ormerod SJ (2004) Dispersal of adult aquatic insects in catchments of differing land use. J Appl Ecol 41(5):934–950
Piscart C, Genoel R, Doledec S, Chauvet E, Marmonier P (2009) Effects of intense agricultural practices on heterotrophic processes in streams. Environ Pollut 157(3):1011–1018
Plafkin JL, Barbour MT, Porter KD, Gross SK, Hughes RM (1989) Rapid bioassessment protocols for use in streams and rivers: benthic macroinvertebrates and fish. EPA/444/4-89-001. U.S. Environmental Protection Agency, Washington, DC
Platts WS, Megahan WF, Minshall GW (1983) Methods for evaluating stream, riparian, and biotic conditions. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experimental Station Technical Report INT-138
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org
Reynolds B, Neal C, Hornung M, Stevens PA (1986) Baseflow buffering of streamwater acidity in five mid-Wales catchments. J Hydrol 87(1–2):167–185
Richardson CJ, Flanagan NE, Ho M, Pahl JW (2011) Integrated stream and wetland restoration: a watershed approach to improved water quality on the landscape. Ecol Eng 37(1):25–39
Roussiez V, Probst A, Probst J-L (2013) Significance of floods in metal dynamics and export in a small agricultural catchment. J Hydrol 499:71–81
SAS Institute Inc (2015) JMP Pro Version 12.2.0. Cary, North Carolina
Schäfer RB, Caquet T, Siimes K, Mueller R, Lagadic L, Liess M (2007) Effects of pesticides on community structure and ecosystem functions in agricultural streams of three biogeographical regions in Europe. Sci Total Environ 382(2–3):272–285
Schauls T, Carter C (2015) Mississippi River-Winona Watershed biotic stressor identification report. Minnesota Pollution Control Agency, St. Paul
Simonson TD, Lyons J, Kanehl PD (1993) Guidelines for evaluating fish habitat in Wisconsin streams. U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station General Technical Report NC-164
Snyder CD, Young JA, Villella R, Lemarié DP (2003) Influences of upland and riparian land use patterns on stream biotic integrity. Landsc Ecol 18(7):647–664
Soulsby C, Youngson AF, Moir HJ, Malcolm IA (2001) Fine sediment influence on salmonid spawning habitat in a lowland agricultural stream: a preliminary assessment. Sci Total Environ 265(1–3):295–307
Sovell LA, Vondracek B, Frost JA, Mumford KG (2000) Impacts of rotational grazing and riparian buffers on physicochemical and biological characteristics of southeastern Minnesota, USA, streams. Environ Manag 26(6):629–641
Talmage PJ, Perry JA, Goldstein RM (2002) Relation of instream habitat and physical conditions to fish communities of agricultural streams in the northern Midwest. N Am J Fish Manag 22(3):825–833
Tesoriero AJ, Duff JH, Wolock DM, Spahr NE (2009) Identifying pathways and processes affecting nitrate and orthophosphate inputs to streams in agricultural watersheds. J Environ Qual 38(5):1892–1900
Teufl B, Weigelhofer G, Fuchsberger J, Hein T (2013) Effects of hydromorphology and riparian vegetation on the sediment quality of agricultural low-order streams: consequences for stream restoration. Environ Sci Pollut Res 20(3):1781–1793
Thorn W, Anderson C, Lorenzen W, Hendrickson D, Wagner J (1997) A review of trout management in southeast Minnesota streams. N Am J Fish Manag 17(4):860–872
Trekels H, van de Meutter F, Stoks R (2011) Habitat isolation shapes the recovery of aquatic insect communities from a pesticide pulse. J Appl Ecol 48(6):1480–1489
Troelstrup NH Jr, Perry JA (1989) Water quality in southeastern Minnesota streams: observations along a gradient of land use and geology. J Minn Acad Sci 55(1):6–13
United States Department of Agriculture (2016) USDA building blocks for climate smart agriculture and forestry. United States Department of Agriculture, Washington, D.C.
Vigiak O, Ribolzi O, Pierret A, Valentin C, Sengtaheuanghoung O, Noble A (2007) Filtering of water pollutants by riparian vegetation: bamboo versus native grasses and rice in a Lao catchment. Unasylva 58(4):11–16
Vondracek B, Blann KL, Cox CB, Nerbonne JF, Mumford KG, Nerbonne BA, Sovell LA, Zimmerman JKH (2005) Land use, spatial scale, and stream systems: lessons from an agricultural region. Environ Manag 36(6):775–791
Walser CA, Bart HL Jr (1999) Influence of agriculture on in-stream habitat and fish community structure in Piedmont watersheds of the Chattahoochee River system. J Freshw Fish 8(4):237–246
Wang L, Lyons J, Kanehl P, Gatti R (1997) Influences of watershed land use on habitat quality and biotic integrity in Wisconsin streams. Fisheries 22(6):6–12
Wang L, Lyons J, Kanehl P (2002) Effects of watershed best management practices on habitat and fish in Wisconsin streams. J Am Water Res Assoc 38(3):663–680
Waters TF (1977) The streams and rivers of Minnesota. University of Minnesota Press, Minneapolis
Waters TF (1995) Sediment in streams: sources, biological effects, and control. American Fisheries Society, Bethesda
Waters TF (2000) Wildstream: a natural history of the free-flowing river. Riparian Press, St. Paul
Whitewater River Watershed Project (2015) A history of the Whitewater Watershed in Minnesota. Whitewater River Watershed Project, Lewiston, Minnesota. http://www.whitewaterwatershed.org/wp-content/uploads/2017/01/Whitewater-Watershed-Conservation-History_Minnesota.pdf
Williams DD, Hynes HBN (1976) The recolonization mechanisms of stream benthos. Oikos 27(2):265–272
Williams MA, Vondracek B (2010) Spring distributions and relationships with land cover and hydrogeologic strata in a karst landscape in Winona County, Minnesota, USA. Carbonates Evaporites 25(4):333–347
Wittman E, Mundahl ND (2003) Development and validation of a benthic index of biotic integrity (B-IBI) for streams in southeastern Minnesota. Winona State University Floruit (Student and Faculty Research from the College of Science and Engineering) 1(1):41–48
Yates AG, Bailey RC, Schwindt JA (2007) Effectiveness of best management practices in improving stream ecosystem quality. Hydrobiologia 583(1):331–344
ZumBerge JR, Perry JA, Lee KE (2003) Influence of Local Riparian Cover and Watershed Runoff Potential on Invertebrate Communities in Agricultural Streams in the Minnesota River Basin. U.S. Geological Survey, Water-Resources Investigations Report 03-4068. Mounds View, Minnesota.
Acknowledgements
We are grateful to the private property owners who provided access to many of the stream sites. Special thanks go to Kristen Nelson-Thomas and Nate Zwonitzer for assisting with field collections and macroinvertebrate sorting. Silas Bergen assisted with statistical analyses. Comments by the Associate Editor and two anonymous reviewers greatly improved the manuscript.
Funding
Funding for this project was provided by the Minnesota Pollution Control Agency. The Agency contributed to general project design, but played no role in collection, analysis, or interpretation of data or in writing of the manuscript.
Author information
Authors and Affiliations
Contributions
NM contributed to project design, conducted field collections, analyzed and interpreted the data, and wrote the manuscript. EM conducted field collections, processed and identified macroinvertebrates, and assisted with data analysis and interpretation. Both authors read and approved the final manuscript.
Author information
Neal D. Mundahl is a Certified Fisheries Professional as accredited by the American Fisheries Society. Erik D. Mundahl is a Professional Environmental Engineer registered in the State of Alaska.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Fish surveys were made under special permit (No. 12769) from the Minnesota Department of Natural Resources, Division of Fish and Wildlife, Section of Fisheries, and were conducted with the approval of the Winona State University Institutional Animal Care and Use Committee.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Mean (1 standard deviation in parentheses) stream habitat variables at 22 sites on the South Fork Whitewater River, Minnesota, USA. CV = current velocity, and Embed = embeddedness. Table S2. Mean (1 standard deviation in parentheses) riparian habitat variables at 22 sites on the South Fork Whitewater River, Minnesota, USA. Table S3. Scores and ratings for fish IBI, macroinvertebrate B-IBI, and fish habitat at 22 sites on the South Fork Whitewater River, Minnesota, USA.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mundahl, N.D., Mundahl, E.D. Aquatic community structure and stream habitat in a karst agricultural landscape. Ecol Process 11, 18 (2022). https://doi.org/10.1186/s13717-022-00365-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-022-00365-1