
Abstract
Background
Many species inhabiting Kihansi gorge ecosystem in Southern Udzungwa Mountains, Tanzania are threatened with extinction due to habitat loss caused by constructed hydropower plant that diverted over 90% the water off the Kihansi River. Understanding ecological correlates and structure of an animal community in perturbed ecosystems is an important way to enhancing species conservation particularly in this system, where several species are already threatened with extinction. This study assessed influences of local habitat characteristics on the butterfly abundance, species richness and diversity and examined how the butterfly community structures across three structurally heterogeneous habitats in the Kihansi gorge forest. Butterfly abundance data were collected using baited traps and measured environmental variables in the field, where trapping occurred. To understand the diversity and butterfly richness, these parameters were computed under Primer software. Non-metric multidimensional scaling and analysis of community similarity were used to analyse the butterfly community structure. Furthermore, the generalized linear models (GLM) were used to assess how the measured variables explained observed species diversity, abundance, and richness.
Results
In total, 72 species of butterfly in five families were recorded. Butterfly species diversity and richness did not vary across the habitat types or elevation gradient but showed strong correlation with some habitat characteristics. In addition, there was evidence of butterfly communities structuring into distinct groups that showed greater separation associated with the habitat characteristics they utilise. Furthermore, across the study area, butterfly species diversity increased significantly in high canopy and in trees with smaller DBH and at lower ground cover. Similarly, abundance significantly increased in the woodland and wooded grassland habitats, where there was high canopy and where tree density was high. Furthermore, butterfly species richness was strongly positively associated with high canopy cover and mostly increased in sites closer to water source.
Conclusion
First, this study shows strong positive correlation between the habitat characteristics and butterfly species and community structure, suggesting strongly that maintaining habitat quality should be a priority in conserving the butterfly communities in Kihansi. Second, conservation of environmental disturbance indicator butterfly taxa which are also endemic and rare is worthy of concern under the ongoing human-induced threats in the area. It is appropriate time that the Tanzania government should declare the Kihansi gorge forest among existing protected areas to secure effective conservation of biological diversity in this threatened ecosystem. Third, the ants–butterfly symbionts and their links to the potential lichen depletion are still unclear, and thus future research should explore the trophic cascades of depleting habitat quality on the endemic and rare butterfly species in the Kihansi gorge.
Similar content being viewed by others


Introduction
Human development process is one among factors that are widely known to affect biodiversity in many regions. Construction activities such as opening up of new roads, erection of power transmission lines, energy development and river excavation and impoundment conducted for development have been documented to affect land cover and several species (Gasparatos et al. 2017; Jones et al. 2015; Preston and Kim 2016). For example, the construction of new roads in central African rainforest increased access to remote forests by illegal hunters, increasing poaching and threatening forest elephants with extinction (Maisels et al. 2013). In addition, human population increase in newly established industrial sites such as hydropower plants (e.g., Bayo and Rija 2021) and wildlife parks (Pickering and Hill 2007) has been increasingly reported to affect local animal and plant biodiversity. Assessing how local biodiversity fairs under such development-related threats is important for detecting any future deleterious impacts on the species and to enable the designing of potential mitigative measures to improve biodiversity conservation.
Disturbance in natural areas resulting from construction activities directly removes important species requirements, such as food, cover and nesting sites. This can greatly reduce ability of the species to reproduce due to diminished suitable conditions for growth and reproduction (Decker 1994; Kocher and Williams 2000). The effect of such disturbances may be particularly severe for smaller organisms such as invertebrates due to their large surface area to body ratio which increases their vulnerability to even subtle changes in habitat conditions such as light penetration (Öckinger and Smith 2006). For butterfly, the loss of quality to their habitats such as removal of host larval plants, reduced nectars and nectar plants that provide source of energy to adult butterflies and vegetation structure may have significant severe impact on their populations (Pywell et al. 2004). Biodiversity assessment in perturbed systems conducted post-disturbance, thus can ensure habitat qualities through improved habitat management strategies informed by research.
Butterflies are considered among indicator species and have been used to track environmental changes and systems recovery following disturbance regimes (Thomas 2005). For example, butterflies have widely been used to detect climate change impacts through range shift polewards due to elevated global temperatures (Hickling et al. 2006; Mason et al. 2015). In Kihansi gorge forest in southern Udzungwa Mountains Tanzania, the habitats inhabited by butterfly and other taxa such as amphibian, birds and plant were greatly perturbed following a major hydropower plant constructed in the 1990s. The project caused significant changes in animal (Channing et al. 2006; Rija et al. 2010) and plant populations (Quinn et al. 2005; Rija et al. 2011) residing the gorge ecosystem. Assessment of arthropods in 1997 prior to the commencement of the hydropower production found high arthropod diversity in spray dominated forest and riverine habitats and low diversity in open grassland areas away from the riverine habitats (Zilihona et al. 1998). Subsequent studies following commencement of the power production revealed varied impacts on the biodiversity. For instance, there was remarkable decrease in the forest specialist birds such as greenbuls (Cordeiro et al. 2006) and decline in coleopteran diversity in sites closer to the river Kihansi (Zilihona et al. 2004). However, recent bird assessment reported no significant changes in bird community structures between pre- and post-hydropower construction periods (Werema and Msuya 2019), suggesting that impacts were probably only associated with some species. Furthermore, some amphibian species such as the critically endangered Kihansi spray toad was severed, leading to its declaration as extinct in the wild species by IUCN (Rija et al. 2010), although reintroduction of the species into the gorge is ongoing (Rija et al. 2014). In addition, some plant species, such as Coffea kihansiensis and Cupea jonii, remain critically endangered or threatened as their habitats have not fully recovered following the habitat disturbance (Vollesen 2000; Rija et al. 2011).
Regular assessment of biodiversity in threatened ecosystems is important for several reasons. First, it can provide information on the population status and important species interactions; second, assessment can detect changes in the target populations or habitats towards a desired state especially in restored systems, such as the Kihansi gorge forests. Third, the assessment can provide information to improve future management actions, particularly protecting the forest from future anthropogenic threats, such as fire and logging (Mwasapi and Rija 2021). Although butterfly is an ideal taxon to monitor and track changes in habitat quality in threatened ecosystems, very scarce information exists on the butterfly communities of the Kihansi gorge forest after disturbance. Furthermore, with the exception of Kielland (1990) describing comprehensive butterfly collections from several parts of Tanzania, other existing studies so far have concentrated on specific localities and mostly biased in protected areas. For example, several studies have described the butterflies of Mount Kilimanjaro National Park in northern Tanzania (Liseki and Vane-Wright 2013, 2014, 2016, 2018), Katavi National Park in western Tanzania (Fitzherbert et al. 2006), Manyara National Park in northern Tanzania (Cordeiro 1990, 1995), Serengeti National Park in northern Tanzania (Sinclair et al. 2015), and Wildlife Management Areas in southern Tanzania (Nkwabi et al. 2017). Others have investigated into how butterflies respond to a gradient of utilization of the miombo forest products in western Tanzania (Jew et al. 2015). These studies are useful in improving our understanding of the butterfly communities and their responses to human utilization of biological resources thereby shedding light into how humans can better interact with the natural landscapes (Bossart et al. 2006; Jew et al. 2015). However, such studies can hardly be used to inform the conservation decisions for threatened and yet unprotected system, such as the Kihansi gorge forest of southern Tanzania.
To date, only one study has provided a checklist of butterflies of Kihansi based on field assessment conducted between 2005 and 2007 immediately after commencing the power production (Mtui et al. 2019). Although the study provides baseline, but detailed assessment of how important local habitat and environmental characteristics interact with species and how this influences the butterfly community structure in this perturbed ecosystem is still lacking. Such information could improve conservation management actions in the area. The current study was conducted to fill this gap and to provide information on how butterfly abundance and diversity is influenced by local habitat characteristics, habitat types and how the two vary spatially along an altitudinal gradient. The aims of this study were to:
-
i.
Assess butterfly diversity and species richness in three structurally heterogeneous habitats and elevation gradients in the Kihansi gorge forest.
-
ii.
Assess how the butterfly community structures in the Kihansi gorge forest.
-
iii.
Examine effects of local habitat characteristics on the butterfly diversity, abundance, and species richness as well as to describe their conservation status based on the IUCN red list.
It was hypothesised that butterfly diversity and richness would be different across various habitat types and elevation gradients due to potentially heterogeneous resources for the butterflies, such as food and cover in these patches. In addition, the characters of the assessed habitats would positively predict butterfly species diversity, abundance and richness and the effect of these would be variable within individual habitat types.
Materials and methods
Study site
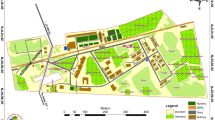
The study was conducted in 2014 in Kihansi gorge forests located at 8° 35′ S, 35° 51′ E along the eastern escarpment of the southern Udzungwa Mountains (Fig. 1). The area is distanced about 600 km East–south from Dar es Salaam city with an elevation of 340 m.a.s.l. to 1240 m.a.s.l. (this study). The annual rainfall ranges between 1000 and 3200 mm (LKEMP 2004) and the mean daily minimum and maximum temperatures is 13 °C and 25 °C, respectively (Cordeiro et al. 2006). The vegetation types comprise montane forest, miombo woodland and wetland spray meadow with various species composing each type (Lovett et al. 1997; Quinn et al. 2005).

Map of Tanzania (left) showing location of the Kihansi gorge forest study site (right) and where butterfly trapping (coloured bullets) was conducted
Sampling of butterfly community
Butterfly species were surveyed using cylindrical butterfly traps of 90 cm height and 30 cm diameter fitted with a round 30 cm diameter plastic bait plate placed at 3 cm below the net (Austin and Riley 1995). In total, 41 traps were set at 600 m interval along nine transects of various sizes (0.9–5 km) measuring a total of 25 km long. Due to limited accessibility to some parts of the Kihansi gorge (Poynton et al. 1998) available paths and trails were used to establish transects for this study. The survey transects intercepted three habitat types: woodland, montane forest and bushed grassland in the Kihansi gorge ecosystem. The habitat types were used as strata for later comparison of the butterfly distribution and abundance. Three transects (each of 3–5 km long) were laid along the woodland habitat, two transects (each of 3 km long) in the bushed grassland and four transects (0.9–3 km long) were used in the montane forest cutting through the gorge spray wetlands (Rija et al. 2011). Each trap was placed on a randomly selected tree c. 3–4 m (depending on the local terrain) above the ground (Bouyer et al. 2007) and on alternate sides of the transects at each trapping point. Depending on the length of transect, a total of 18, 13 and 10 traps were placed in the woodland, montane forest and the wooded grassland, respectively.
To attract fruit feeding butterflies, traps were baited with fermented banana prepared 48 h prior to the trapping (Alexander and DeVries 2012). This technique is very effective in surveying tropical butterflies and can greatly minimize errors of identification than when species are identified in flight (Sparrow et al. 1994; Walpole and Sheldon 1999). Traps were left for fourteen successive days, thereafter checked and emptied between 12:00 and 14:00 local time every day. Each time checking of traps started from the other end of the transect to avoid potential bias and to ensure independence of the data collected at individual trap level, allowing data pooling during analysis. The bait was inspected and replaced every day to prevent the bait from over-drying and replacing consumed or rain-washed bait (Houlihan et al. 2013). Trapped butterflies were removed, identified and marked with non-toxic silver markers, allowing identification of subsequent recaptures which were later excluded during analysis. After recording all captures released in the wild, where species were not identifiable in the field, voucher specimens were collected and later identified in the laboratory. Voucher specimens of all trapped species are stored in the Zoology laboratory at Sokoine University of Agriculture.
To understand species distribution across the habitats and altitudes, GPS coordinates of each trapping point were recorded using hand-held GPS, Garmin eTrex 20. Two butterfly trapping events were conducted during late dry season (September–October 2014) and during mid-rain season (April 2015) and measured habitat cover at trap location to assess the influence of these generic variables on the butterfly richness and diversity. Vegetation cover (%) at ground, low (2 m above ground), and understorey and canopy cover were estimated and recorded. Other variables measured included tree diameter at breast height (DBH), plot tree density, distance to nearby water source (measured using GPS, Garmin eTrex 10), trap elevation above sea level and habitat type, where a trap was set. The habitat characteristics were used to find out their influence on the abundance, species richness and diversity of butterfly in the study area.
Data analyses
Prior to analysis, butterfly abundance data were pooled from individually sampled days for each trap and habitat characteristics recorded for each sampling station. First, to understand how species-rich and diverse the study area was, butterfly richness and diversity were analysed using Chao1 estimator and Shannon diversity index, respectively, under program Estimate (Colwell 2013). To evaluate species richness and diversity indices, the numbers of runs were set to 100 to get smoothed curves at all levels of species accumulation (Gotelli and Colwell 2001). At this stage, randomization with sample replacement was checked, ensuring a target diversity index is selected to obtain richness and diversity estimates. Species richness and diversity data were further compared between habitat types and altitudinal gradients using Kruskal-Wallis tests after confirming non-normal distribution (Shapiro test, p > 0.05).
Second, to understand how butterfly populations organize across the surveyed habitat types, a cluster analysis was performed on a Bray–Curtis similarity matrix of grouped butterfly samples under program Primer ver.6 (Gorley and Clarke 2006). To do this, butterfly abundance data were first square root transformed for normality, reducing the effect of high abundance species (Clarke et al. 2014). Furthermore, non-metric multidimensional scaling (NMDS) and cluster plots were used to understand similarity in butterfly communities between habitat types. When clustering was evident a similarity profile (SIMPROF) test was used to examine if there was statistically significant clustering in the habitat sites (Gorley and Clarke 2006). To find out significant differences in butterfly communities collected from the surveyed habitat types, analysis of similarity (ANOSIM) test (i.e., ANOSIM is similar to classical ANOVA test) was used between pairs of the butterfly communities (Clarke et al. 2014). This test produces a global R-statistic which ranges from 0 to 1, indicating relative significant difference between the butterfly community groups being compared (Clarke 1993).
Third, to investigate how local habitat and environmental characteristics (henceforth termed as variables) affect butterfly diversity, abundance and species richness in the Kihansi gorge forest, the data were analysed using generalised linear models (GLM) under R-software ver. 4.0.5 (R Core Team 2021). Prior to model fitting, correlation analysis was conducted to check potential collinearity between independent variables. For the highly correlated variables (r > 0.5), only one was included in the model due to redundancy. Three models, one for each dependent variable (i.e., species diversity, abundance and species richness) were built incorporating eight explanatory variables: habitat type, tree density, tree DBH, distance to water point, ground cover, and herbaceous cover at 2 m height, canopy cover and elevation. The GLM model with Poisson family and log link function was used to fit the species diversity data, while abundance and species richness data were fitted with quasipoisson family and log link functions after confirming model overdispersion (theta ≥ 1.4). Model validation was conducted through examining deviance residuals distribution which looked fine. To evaluate the relative effect of each variable on the dependent variable, a backwards stepwise single deletion of non-significant terms was performed. For each deletion, model significance was tested using F test for the model assessing diversity and Chi-test for models on abundance and richness (Kamil 2018). For any two competing models observed during the stepwise deletion of non-significant terms, analysis of variance (ANOVA) test was used to select the most parsimonious model based on the model showing the smallest residual deviance. Furthermore, prediction models were built for the final significant model terms using the “predict function” in the package “ggplot2” to understand how well the independent variables predicted the dependent variables. Explained deviance for each model was calculated as pseudo R2. Finally, to understand the conservation status, each taxon was crosschecked against the current IUCN red list (IUCN 2021) and available literature in Africa and globally (New 1993; Edge and Mecenero 2015) to discuss their threat level based on observed abundance data. Endemism of some taxa were obtained from available literature (Congdon and Bampton 2001; Collins and Larsen 2008).
Results
Description of butterfly biodiversity
In total, 1271 individuals representing 72 species of the order Lepidoptera, belonging to 5 families were recorded in the study area (Additional file 1: Table S1). Of the species, 23 were common, eight uncommon and forty-one rare species (Additional file 1: Fig. S1). Butterfly abundance was on average relatively higher in montane forest than other habitat types (Additional file 1: Fig. S2) but showed no significant difference (χ2 = 0.858, df = 2, p = 0.651). Similarly, there was no significant variation in the median diversity across the elevation (χ2 = 3.437, df = 2, p = 0.179 and habitat types (χ2 = 2.115, df = 2, p = 0.347) nor richness at elevation gradient (χ2 = 4.541, df = 2, p = 0.103) or habitat types (χ2 = 0.858, df = 2, p = 0.651; Fig. 2).

Pattern of butterfly diversity (lower) and species richness (upper) at elevation gradient and between habitat types in the Kihansi gorge forest. No significant difference was found in these butterfly parameters across the elevation and habitat types (see text for statistic)
Among the butterfly families, Nymphalidae showed highest contribution (65.28%, n = 47) to the total observed species, while Hesperiidae was the least dominant. On another hand, family Nymphalidae ranked first in terms of species richness (Woodland, 26 species; Montane forest, 28 species; Wooded grassland, 26 species) when study sites were considered independently (Additional file 1: Fig. S3). Considering the number of individuals across the entire study area, family Nymphalidae scored the highest with 72.46% (n = 921), followed by Pieridae 13.14% (n = 167), Papilionidae (10.62%, n = 135), Lycaenidae (2.28%, n = 29) and last Hesperiidae (1.49%, n = 19).
Influence of habitat characteristics on abundance, species richness and diversity
Butterfly species diversity, abundance and species richness were significantly influenced by the measured local habitat variables (Table 1). Butterfly species diversity showed contrasting response to canopy cover, tree DBH and ground cover, where the diversity increased significantly at high canopy cover and decreased mostly in sites with high ground cover and trees with larger DBH (Fig. 3). Furthermore, the abundance of butterfly was significantly positively associated with increasing canopy cover and tree density and in the woodland and wooded grassland vegetation types but was lower in the moist forest habitat (Fig. 4). Species richness was significantly positively associated with the canopy cover and tended to increase in sites closer to water than further away (Fig. 5).

Effect of canopy cover, tree DBH and ground cover on the diversity of butterfly in the Kihansi gorge forest. Butterfly diversity increased with increasing canopy cover and in trees with lower DBH and at lower ground cover. The shaded part indicates 95% confidence interval of the estimated effect size. Grey dots are raw data points

Effect of habitat type, canopy cover and tree density on the abundance of butterfly in Kihansi gorge forest indicating butterfly abundance increasing with increasing canopy cover, tree density and at woodland and wooded grassland habitats. The habitat type plot (right) shows median estimates with standard errors, while the shaded part (left) indicates 95% confidence interval of the estimated effect size. Grey dots are raw data points

Effect of canopy cover and distance to water points on the species richness of butterfly in the Kihansi gorge forest. Canopy cover and distance to water were positive and negative predictors of species richness, respectively. The shaded part indicates 95% confidence interval of the estimated effect size. Grey dots are raw data points
Butterfly community structure
The sampled butterflies were grouped into three distinct butterfly communities that showed significant structuring within (Pi = 3.367, p = 0.2%, Fig. 6) and greater separation across the communities (Global R = 0.102, p = 1.1%). Furthermore, examining the pattern of group separation showed that only the butterfly samples collected from the woodland and montane forest habitats contributed significantly to the observed group separation (ANOSIM test, Global R = 0.156, p = 0.5%). The samples from the woodland and wooded grassland or from montane forest and wooded grassland were closely similar.

Dendrogram of butterfly samples collected in three habitats Woodland (W), Montane forest (MF) and Wooded grassland (WG) within the Kihansi gorge forest ecosystem showing three broad butterfly communities A–C. There was some evidence of internal structuring within individual community groups but also greater separation between the distinct butterfly communities (see text for more statistics)
Butterfly taxa of conservation concerns in Kihansi
Of the species collected, 20.8% (n = 15) are listed as Least Concern, while the rest have not been assessed, thus unavailable in the IUCN red list. Three species from the Family Hesperiidae (one species) and Family Nymphalidae (two species) in the genera Celaenorrhinus and Charaxes, respectively, are endemic to the study area (Table 2). Furthermore, based on the conservation assessment of butterfly elsewhere is Southern Africa, the genus Athene contains species that are either critically endangered or vulnerable, while the genera Papilio, Colotis and Charaxes have species considered to be extremely rare.
Discussion
This study aimed to understand how butterfly biodiversity varies across a heterogeneous habitat, elevation gradient and the effect of local habitat characteristics on the butterfly species diversity, abundance and richness in the Kihansi gorge forest. The butterfly diversity and richness were not significantly different across the habitat types and elevation gradient. There was evidence of butterfly community groups showing greater separation associated with the habitats they utilise. Furthermore, species diversity increased in areas with high canopy cover, in trees with smaller DBH and at low ground cover. The butterfly abundance, on another hand, increased in sites with high tree density and high canopy cover within the woodland and wooded grassland habitats. Furthermore, species richness tended to increase at high canopy cover and in sites closer to the water source.
The butterfly records observed in the current study are lower than previously published for this area. In their study, Mtui et al. (2019) used Malaise traps to collect butterflies over a 2-year period between 2005 and 2007, just 5 years after commencing power production in the Kihansi gorge forest. They recorded > 200 butterfly species distributed in six families. These species are almost three times more than recorded in the present study, 14 years after hydropower production. The difference in the species records between these studies could partly be attributed to the sampling methods used and the time spent collecting data but also more so on the possible changes in the conditions that favour survival and reproduction of the butterflies in this ecosystem. For instance, several studies on various plant and animal taxa conducted within 5 years of commencing power production in the gorge reported significant changes in biological species (e.g., specialist birds and Coleopterans) associating the changes to the reduced river flow particularly in areas closer to the river waterfall (Zilihona et al. 2004; Cordeiro et al. 2006). In addition, increased weed encroachment in the Kihansi gorge wetlands (Quinn et al. 2005), increased exposure to new disease infestation in the critically endangered Coffea kihansiensis (Rija et al. 2011) and extinction in the wild of the critically endangered amphibian Nectophrynoides asperginis after 7 years of commencing power production (Rija et al. 2010) are some impacts of the environmental change previously recorded in the gorge forest ecosystem. Based on the impacts on these species, it is fair to suggest that the difference in the butterfly records is due to the disturbances and temporal changes of conditions in their habitats which may have influenced decline in their abundance and species richness. Although there is no study done to date specifically to quantify the impact of the reduced river flow on the abundance of food resources (e.g., nectars) and availability of suitable breeding cover (e.g., presence of larval host plants) for the butterflies in the Kihansi gorge forest ecosystem, such impacts have been observed in other ecosystems elsewhere (Kocher and Williams 2000; Pywell et al. 2004), highlighting the importance of carrying out biodiversity monitoring especially in perturbed systems, such as the Kihansi gorge forest.
The butterfly communities were broadly grouped into three distinct groups but no single habitat type alone appears to better explain these broad structures as most butterfly populations from various habitat types seem to group together. This may suggest the characters within these habitats are more important in supporting the utilization of a particular habitat by the butterfly species than the habitat types themselves. This supports results from the GLM that identified important local habitat characters on the butterfly assemblage in a particular habitat. The implications of these results on broader conservation of the butterfly communities and perhaps other threatened species in the Kihansi ecosystem is that these habitats require greater conservation attention in preventing more anthropogenic threats that will lead to erosion of the habitat qualities that have shown to influence butterfly populations in the area. Although the main conservation threat to the various taxa inhabiting the Kihansi gorge forest ecosystem has been the reduced river flow, several other threats, such as sporadic logging, tree pole cutting and agricultural encroachment have been reported in Kihansi (Rija et al. 2010) and nearby forests (Mwasapi and Rija 2021). Such threats have severe potential effects on the butterfly communities as they directly influence the quality of the habitats through reducing suitable larval host plants and reducing shade that influences butterfly breeding. These threats if not checked, will compound the situation of this fragile ecosystem thereby sever the biological diversity in the area.
Local habitat characteristics variously influenced species diversity, abundance and species richness in the study area. The high butterfly diversity deduced in sites with high canopy cover and in tree vegetation with smaller DBH could be due to the presence of more shade that is associated with high canopy that predicts high species diversity. Shade availability is known to favour egg ovipositioning and larval development during butterfly breeding (Warren 1985; Grundel et al. 1998). Although shade was not directly measured in the current study, it is not clear yet what proportion of the shade is most suitable for attaining maximum species diversity in Kihansi. In a study conducted elsewhere, for example, slight than dense shade was reported to be more important for some butterfly species (Greatorex-Davies et al. 1993). In addition, it appears that various species have differing requirements for survival and breeding due to possessing various traits, e.g., feeding niche, dispersal propensity, egg number, number of generations per year, shade requirement etc. (Börschig et al. 2013). Linking these traits to explain the population and community structure of the Kihansi butterflies would be prudent as future research gaps in this threatened ecosystem.
The strong positive association between high canopy cover and butterfly abundance and species richness particularly in the woodland and wooded grassland suggests that these habitats possess unique resources on which the butterfly species depend. High canopy is associated with more light penetration—an important resource that favours foraging especially for forest canopy foraging species (Hill et al. 2001; Houlihan et al. 2013). This result suggests also that a large proportion of the butterfly records observed in this study was canopy specialists. This may further suggest that the woodlands and wooded grassland habitat qualities were probably not affected by the environmental change caused by the reduction of the river flow or that suitable conditions may have recovered in these habitats 14 years after disturbance. While these speculations may be valid, there is no existing ecological study to date upon which to benchmark these observations. Elsewhere in the rainforest of Manu Biosphere Reserve in Peruvian Amazon, Whitworth et al. (2016) reported fewer canopy forest than the terrestrial species suggesting that the forest disturbance had more impact on the canopy species or that they recovered relatively slower than their terrestrial counterparts. Despite this, our system has had no history of intensive logging as that in Peru, thus, the development-induced disturbance could still hold water. Furthermore, the importance of forest canopy in the Kihansi gorge forest is further supported by the observed butterfly community structuring with high abundant butterflies in the two habitat types. The distinct separation among the community groups inhabiting the woodland and wooded grasslands suggests presence of important subtle conditions that are necessary for the butterfly survival and reproduction, such as availability of larval host plants and food resources in such habitats (Hellmann 2002; Curtis et al. 2015). Furthermore, an alternative explanation to the low abundance and species richness recorded in the montane forest could be related to the habitat disturbance caused by illegal logging and tree cutting for poles happening in the montane forest patch (Rija et al. 2010). Such habitat disturbances directly remove habitat conditions required for butterfly breeding thereby affecting their overall abundance and richness (Hellmann 2002).
The recorded butterfly taxa have great conservation significance in the Kihansi gorge because of being endemic and potentially rare in this area. In addition, several genera including Eurema (from Subfamily Coliadinae), Belenois (Subfamily Pierinae), Bycyclus (Subfamily Satyrinae), Charaxes (Subfamily Charaxini) and Neptis (Subfamily Limenitidinae) are known to be good indicators of environmental disturbances associated with the human impacts in the miombo woodlands in western Tanzania (Jew et al. 2015). The finding of these genera in the current study suggests that the Kihansi gorge is facing habitat disturbances ranging from low to high levels. The impact of these disturbances is far reaching, both on the endemic taxa and other species whose conservation statuses are yet to be established. Furthermore, these disturbances, coupled with the declining microclimatic conditions (i.e., reduced relative humidity) may deplete favourable conditions for the survival of ants which are known to be host for many other insects including butterflies (Edge et al. 2008). Although there has been no study conducted on the ant–butterfly symbiotic relations in the Kihansi gorge, if such symbionts do exist and as many as 84% of butterflies elsewhere are myrmecophilous (Edge and Macenero 2015), then the Kihansi butterflies will be no exception. The decline of microclimates in the Kihansi gorge (Rija et al. 2011; Quinn et al. 2005) may directly reduce lichen, thereby further threatens the butterfly communities that are dependent on them for food. The existing disturbances and their cascades are likely to sever the endemic and those seemingly rare taxa. These results call for immediate conservation measures to effectively protect the whole Kihansi ecosystem.
Finally, this study has painted the first ecological picture on the butterfly communities in relation to the habitat characteristics in the threatened Kihansi gorge forest—an area that has already recorded extinction of one amphibian species in the recent decade due to the human development activities. To date, it is still unknown how the existing threats in this ecosystem have affected the butterfly taxa that were originally there as there is no effective population monitoring system available to track changes in the butterfly communities inhabiting this ecosystem. As is shown in the present study and from others elsewhere that subtle qualities of the habitat are necessary for the butterfly population, but the influences of these on the individual butterfly species in the study area have not yet been quantified. Future studies looking into how various individual butterfly species are influenced by the available qualities of the habitats will be necessary in generating information that will be useful in identifying species-specific needs for improving conservation of the butterfly community in the Kihansi gorge ecosystem.
Availability of data and materials
Data is available and will be shared to individuals when requested from the author.
Abbreviations
- DBH:
-
Diameter at breast height
- NMDS:
-
Non-metric multidimensional scaling
- SIMPROF:
-
Similarity profile
- ANOSIM:
-
Analysis of similarity
- ANOVA:
-
Analysis of variance
- GLM:
-
Generalized linear modelling
- WWCT:
-
The Whitley Wildlife Conservation Trust
- TBA:
-
Tropical Biology Association
References
Alexander LG, DeVries PJ (2012) Variation in capture height and trap persistence among three Costa Rican understorey butterfly species. J Trop Ecol 28:585–589
Austin GT, Riley TJ (1995) Portable bait traps for the study of butterflies. Trop Lepid 6:5–9
Bayo MJ, Rija AA (2021) Distribution and human-otter interaction in a man-made Mtera Dam: how fishers perceive their Neighbour. IUCN Otter Spec Group Bull 38:95–105
Börschig C, Klein AM, von Wehrden H, Krauss J (2013) Traits of butterfly communities change from specialist to generalist characteristics with increasing land-use intensity. Basic Appl Ecol 14:547–554. https://doi.org/10.1016/j.baae.2013.09.002
Bossart JL, Opuni-Frimpong E, Kuudaar S, Nkrumah E (2006) Richness, abundance, and complementarity of fruit-feeding butterfly species in relict sacred forests and forest reserves of Ghana. In: Hawksworth DL, Bull AT (eds) Arthropod diversity and conservation. Springer, Netherlands, Dordrecht, pp 319–345
Bouyer J, Sana Y, Samandoulgou Y, Cesar J, Guerrini L, Kabore-Zoungrana C, Dulieu D (2007) Identification of ecological indicators for monitoring ecosystem health in the trans-boundary W Regional park: a pilot study. Biol Conserv 138:73–88. https://doi.org/10.1016/j.biocon.2007.04.001
Channing A, Finlow-Bates KS, Haarklau SE, Hawkes PG (2006) The biology and recent history of the critically endangered Kihansi spray toad Nectophrynoides asperginis in Tanzania. J East Afri Nat Hist 95:117–138
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aus J Ecol 18:117–143. https://doi.org/10.1111/j.1442-9993.1993.tb00438.x
Clarke KR, Gorley R, Somerfield PJ, Warwick R (2014) Change in marine communities: an approach to statistical analysis and interpretation: Primer-E Ltd
Collins SC, Larsen TB (2008) Eighteen new species, five new subspecies, and interesting data on other African butterflies - Fourth ABRI research paper. Metamorphosis 19:42–113
Colwell RK (2013) EstimateS: Statistical estimation of species richness and shared species from samples. Version 9 and earlier. User’s Guide and application. http://purl.oclc.org/estimates
Congdon TCE, Bampton I (2001) Some endemic butterflies of Eastern Africa and Malawi. Available at https://www.coastalforests.tfcg.org/pubs/Eastern%20Africa%20Butterfly%20Endemics%20DRAFT.pdf
Cordeiro NJ (1990) A provisional, annotated checklist of the butterflies in Lake Manyara National Park, Arusha Region, Tanzania. J East Afri Nat Hist Soc Nat Mus 80:25–41
Cordeiro NJ (1995) Interesting distribution records of butterflies from northern Tanzania. Metamorphosis 6:194–198
Cordeiro NJ, Lovett JC, Mulungu E, Maina GG, Gerstle JH (2006) Initial trends of bird assemblages before and after river diversion in an endemic-rich African forest. Biodivers Conserv 15:971–983. https://doi.org/10.1007/s10531-004-3104-7
Curtis RJ, Brereton TM, Dennis RLH, Carbone C, Isaac NJB (2015) Butterfly abundance is determined by food availability and is mediated by species traits. J App Ecol 52:1676–1684. https://doi.org/10.1111/1365-2664.12523
Decker BS (1994) Effects of habitat disturbance on the behavioral ecology and demographics of the Tana River red colobus (Colobus badius rufomitratus). Int J Primatol 15:703–737
Edge DA, Mecenero S (2015) Butterfly conservation in southern Africa. J Insect Conserv 19:325–339
Edge DA, Robertson HG, Van Hamburg H (2008) Ant assemblages at three potential breeding sites for the Brenton blue butterfly, Orachrysops niobe (Trimen). Afr Entomol 16:253–262
Fitzherbert E, Gardner T, Davenport TRB, Caro T (2006) Butterfly species richness and abundance in the Katavi ecosystem of western Tanzania. Afr J Ecol 44:353–362. https://doi.org/10.1111/j.1365-2028.2006.00655.x
Gasparatos A, Doll CN, Esteban M, Ahmed A, Olang TA (2017) Renewable energy and biodiversity: implications for transitioning to a Green Economy. Renew Sustain Energy Rev 70:161–184
Gorley CK, Clarke K (2006) PRIMER v6: user manual/tutorial. PRIMER-E, Plymouth, UK.
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Let 4:379–391. https://doi.org/10.1046/j.1461-0248.2001.00230.x
Greatorex-Davies JN, Sparks TH, Hall ML, Marss RH (1993) The influence of shade on butterflies in rides of coniferised lowland woods in Southern England and implications for conservation management. Biol Conserv 63:31–41. https://doi.org/10.1016/0006-3207(93)90070-H
Grundel R, Pavlovic NB, Sulzman CL (1998) Habitat use by the endangered Karner blue butterfly in oak woodlands: the influence of canopy cover. Biol Conserv 85:47–53. https://doi.org/10.1016/S0006-3207(97)00165-1
Hellmann JJ (2002) The effect of an environmental change on mobile butterfly larvae and the nutritional quality of their hosts. J Anim Ecol 71:925–936. https://doi.org/10.1046/j.1365-2656.2002.00658.x
Hickling R, Roy DB, Hill JK, Fox R, Thomas CD (2006) The distributions of a wide range of taxonomic groups are expanding polewards. Glob Change Biol 12:450–455. https://doi.org/10.1111/j.1365-2486.2006.01116.x
Hill J, Hamer K, Tangah J, Dawood M (2001) Ecology of tropical butterflies in rainforest gaps. Oecologia 128:294–302
Houlihan PR, Harrison ME, Cheyne SM (2013) Impacts of forest gaps on butterfly diversity in a Bornean peat-swamp forest. J Asia-Pacific Entomol 16:67–73. https://doi.org/10.1016/j.aspen.2012.10.003
IUCN (2021) The IUCN Red List of Threatened Species. https://www.iucnredlist.org/ (Accessed on Sept 15, 2021)
Jew EKK, Loos J, Dougill AJ, Sallu SM, Benton TG (2015) Butterfly communities in miombo woodland: biodiversity declines with increasing woodland utilisation. Biol Conserv 192:436–444. https://doi.org/10.1016/j.biocon.2015.10.022
Jones NF, Pejchar L, Kiesecker JM (2015) The energy footprint: how oil, natural gas, and wind energy affect land for biodiversity and the flow of ecosystem services. Bioscience 65:290–301
Kamil B (2018) MuMIn: multi-model inference. R package version 1.42.1
Kielland J (1990) Butterflies of Tanzania: Hill House
Kocher SD, Williams EH (2000) The diversity and abundance of North American butterflies vary with habitat disturbance and geography. J Biogeogr 27:785–794
Liseki SD, Vane-Wright RI (2013) Butterflies (Lepidoptera: Papilionoidea) of Mount Kilimanjaro: family Pieridae, subfamily Coliadinae. J Nat Hist 47:1309–1323
Liseki SD, Vane-Wright RI (2014) Butterflies (Lepidoptera: Papilionoidea) of Mount Kilimanjaro: family Pieridae, subfamily Pierinae. J Nat Hist 48:1543–1583
Liseki SD, Vane-Wright RI (2016) Butterflies (Lepidoptera: Papilionoidea) of Mount Kilimanjaro: Nymphalidae subfamilies Libytheinae, Danainae, Satyrinae and Charaxinae. J Nat Hist 50:865–904
Liseki SD, Vane-Wright RI (2018) Butterflies (Lepidoptera: Papilionoidea) of Mount Kilimanjaro: Nymphalidae subfamily Helconiinae. J Nat Hist 52:2511–2552
LKEMP (2004) Lower Kihansi Hydropower project: immediate rescue and emergency measures. Final Specialist report: amphibian studies: report produced for Tanzania Electric Supply Company Ltd (TANESCO), Dar es Salaam, Tanzania
Lovett JC, Hatton J, Mwasumbi LB, Gerstle JH (1997) Assessment of the impact of the Lower Kihansi Hydropower Project on the forests of Kihansi Gorge, Tanzania. Biodivers Conserv 6:915–934. https://doi.org/10.1023/A:1018307412267
Maisels F, Strindberg S, Blake S, Wittemyer G, Hart J, Williamson EA et al (2013) Devastating decline of forest elephants in Central Africa. PLoS ONE 8:e59469. https://doi.org/10.1371/journal.pone.0059469
Mason SC, Palmer G, Fox R, Gillings S, Hill JK, Thomas CD, Oliver TH (2015) Geographical range margins of many taxonomic groups continue to shift polewards. Biol J Linnean Soc 115:586–597. https://doi.org/10.1111/bij.12574
Mtui D, Congdon C, Bampton I, Kalenga P, Haule L (2019) Altitudinal distribution and monthly occurrence of butterflies in the Kihansi Gorge Forest, Tanzania, with a checklist of species. Tanzania J Sci 45:543–558
Mwasapi BS, Rija AA (2021) Local habitat characteristics influence abundance and community structure of rodents in a regenerating Lulanda reserved forest, southern Tanzania. Restor Ecol e13501
New TR (1993) Conservation biology of Lycaenidae (butterflies): occasional paper of the IUCN species survival commission. IUCN, Gland, pp 1–21
Nkwabi AK, Liseki S, Bukombe JK, Kija H, Mmassy E, Otsyina RM et al (2017) Species richness and composition of butterfly with reference to anthropogenic activities in the wildlife management areas, Southern Tanzania. Int J Fauna Biol Stud 4:34–40
Öckinger E, Smith HG (2006) Landscape composition and habitat area affects butterfly species richness in semi-natural grasslands. Oecologia 149:526–534
Pickering CM, Hill W (2007) Impacts of recreation and tourism on plant biodiversity and vegetation in protected areas in Australia. J Environ Manage 85:791–800
Poynton JC, Howell KM, Clarke BT, Lovett JC (1998) A critically endangered new species of Nectophrynoides (Anura: Bufonidae) from the Kihansi Gorge, Udzungwa Mountains, Tanzania. Afr J Herpetol 47:59–67. https://doi.org/10.1080/21564574.1998.9650003
Preston TM, Kim K (2016) Land cover changes associated with recent energy development in the Williston Basin; Northern Great Plains, USA. Sci Total Environ 566:1511–1518
Pywell R, Warman E, Sparks T, Greatorex-Davies J, Walker K, Meek W et al (2004) Assessing habitat quality for butterflies on intensively managed arable farmland. Biol Conserv 118:313–325
Quinn CH, Ndangalasi HJ, Gerstle J, Lovett JC (2005) Effect of the lower Kihansi hydropower project and post-project mitigation measures on wetland vegetation in Kihansi Gorge, Tanzania. Biodivers Conserv 14:297–308. https://doi.org/10.1007/s10531-004-5048-3
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
Rija AA, Khatibu FH, Mwamende KA (2010) Reclaiming the lost world: Kihansi spray toad re-introduction in Tanzania. In: Soore PS (ed) Global re-introduction perspectives: additional case-studies from around the globe. IUCN/SSC Re-introduction Specialist Group, Abu Dhabi, UAE
Rija AA, Mwamende KA, Hassan SN (2011) The aftermath of environmental disturbance on the critically endangered Coffea kihansiensis in the Southern Udzungwa Mountains, Tanzania. Trop Conserv Sci 4:359–372
Rija AA, Goboro EM, Mwamende KA, Said A, Kohi EM, Hassan SN (2014) Activity patterns and fine-scale resource partitioning in the gregarious Kihansi spray toad Nectophrynoides asperginis in captivity. Zoo Biol 33:411–418
Sinclair AR, Nkwabi AK, Metzger KL (2015) The butterflies of serengeti: impact of environmental disturbance on biodiversity. Serengeti IV: sustaining biodiversity in a coupled human-natural system. University of Chicago Press, pp 301–322
Sparrow HR, Sisk TD, Ehrlich PR, Murphy DD (1994) Techniques and guidelines for monitoring neotropical butterflies. Conserv Biol 8:800–809. https://doi.org/10.1046/j.1523-1739.1994.08030800.x
Thomas JA (2005) Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Phil Trans Royal Soc B: Biol Sci 360:339–357. https://doi.org/10.1098/rstb.2004.1585
Vollesen K (2000) Two new Tanzanian Acanthaceae. Kew Bull 55:965–969. https://doi.org/10.2307/4113643
Walpole MJ, Sheldon IR (1999) Sampling butterflies in tropical rainforest: an evaluation of a transect walk method. Biol Conserv 87:85–91. https://doi.org/10.1016/S0006-3207(98)00037-8
Warren MS (1985) The influence of shade on butterfly numbers in woodland rides, with special reference to the wood white Leptidea sinapis. Biol Conserv 33:147–164. https://doi.org/10.1016/0006-3207(85)90101-6
Werema C, Msuya C (2019) Understorey bird communities 8 and 18 years after river diversion in Kihansi Gorge, Udzungwa Mountains in the Eastern Arc Mountains, Tanzania. Tanzania J Sci 45:661–671
Whitworth A, Villacampa J, Brown A, Huarcaya RP, Downie R, MacLeod R (2016) Past human disturbance effects upon biodiversity are greatest in the canopy; a case study on rainforest butterflies. PLoS ONE 11:e0150520. https://doi.org/10.1371/journal.pone.0150520
Zilihona IJE, Heinonen J, Nummelin M (1998) Arthropod diversity and abundance along the Kihansi Gorge (Kihansi River) in the Southern Udzungwa Mountains, Tanzania. J East Afri Nat Hist 87(1):233–240
Zilihona IJE, Niemelä J, Nummelin M (2004) Effects of a hydropower plant on Coleopteran diversity and abundance in the Udzungwa Mountains, Tanzania. Biodivers Conserv 13:1453–1464. https://doi.org/10.1023/B:BIOC.0000021325.90554.0b
Acknowledgements
This study was funded through the support of two small grants from The Whitley Wildlife Conservation Trust (WWCT) in UK and Tropical Biology Association (TBA). I thank Abubakari S. Mgelwa, Munga J, Aloyce Mtenga and Person Kalenga for their assistance during data collection in the field. Anonymous reviewers provided comments on the manuscript.
Funding
Field surveys were supported to WWCT and TBA acknowledged fully in the acknowledgement section.
Author information
Authors and Affiliations
Contributions
The author designed, carried out field surveys, data analysis and writing of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Research permit was granted by relevant authority and approved by Sokoine University of Agriculture.
Consent for publication
Not applicable.
Competing interests
There is no competing interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Additional results showing records (based on presence/absence) of butterfly species, rarity and number of butterfly individuals recorded across the three sampled habitat types.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rija, A.A. Local habitat characteristics determine butterfly diversity and community structure in a threatened Kihansi gorge forest, Southern Udzungwa Mountains, Tanzania. Ecol Process 11, 13 (2022). https://doi.org/10.1186/s13717-022-00359-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-022-00359-z