
Abstract
Background
Microvesicles (MV) are extracellular vesicles known to be associated with cellular activation and inflammation. Hemofiltration is an effective blood purification technique for patients with renal failure and possibly also eliminates inflammatory mediators in the setting of sepsis. On the other hand, proinflammatory stimuli are induced by blood contacting the artificial membrane during extracorporeal blood purification. In chronic dialysis patients a systemic increase in MV has been described. The aim of the study was to investigate whether hemofilter passage of blood in continuous veno-venous hemofiltration (CVVH) alters MV composition and levels in critically ill patients with sepsis.
Methods
Pre- and postfilter bloods as well as ultrafiltrate samples from intensive care unit patients with severe sepsis were obtained during CVVH with regional citrate anticoagulation. MV subtypes in blood were analyzed by high-sensitivity flow cytometry. Additionally, tissue factor (TF) levels and MV-associated TF activities as well as MV activities were quantified. All parameters were corrected for hemoconcentration applied during CVVH.
Results
Twelve patients were analyzed. A significant increase in presumably mostly leukocyte-derived CD31+/CD41− MV (1.32 (1.09–1.93)-fold [median (25th–75th quartiles)], p = 0.021) was observed post- to prefilter, whereas platelet-derived MV as well as AnnexinV-binding MV were unaltered. Increments of AnnexinV+, CD42b+ and CD31+/CD41− MV post- to prefilter correlated with filtration fraction (FF) (all p < 0.05). Significant reductions in MV activity [0.72 (0.62–0.84)-fold, p = 0.002] and TF level [0.95 (0.87–0.99)-fold, p = 0.0093] were detected postfilter compared to prefilter. No MV activity was measurable in ultrafiltrate samples.
Conclusions
Despite clearing a fraction of small PS-exposing MV CVVH does not eliminate larger MV. Concurrently, CVVH induces the release of CD31+/CD4− MV that indicate leukocyte activation during hemofilter passage in septic patients. Increments of several MV subtypes within the hemofilter correlate with FF, which supports common recommendations to keep FF low. A fraction of TF is being cleared by CVVH via ultrafiltration.
Similar content being viewed by others

Background
Continuous veno-venous hemofiltration (CVVH) is a frequently used renal replacement therapy (RRT) modality in critically ill patients [1]. Although it is an effective and life-saving treatment in the presence of renal failure, its beneficial effect as sole supportive sepsis therapy is still controversial [2, 3]. It has been shown that hemofiltration is able to influence levels of circulating mediators such as inflammatory cytokines by adsorption or filtration [4]. However, these potentially favorable effects might be offset by proinflammatory stimuli that are induced by blood contacting an artificial membrane during extracorporeal blood purification itself [5–7]. This effect may be enhanced when heparin is used for anticoagulation and attenuated during regional citrate anticoagulation [8]. Microvesicles (MV) are extracellular vesicles sized between approximately 100 nm and 1 µm that are released by several cell types upon stimulation or apoptosis [9, 10]. They mediate pleiotropic inflammatory signals during sepsis [11] and are associated with the occurrence of disseminated intravascular coagulation [12]. In patients with chronic renal failure increased systemic levels of platelet-derived as well as procoagulatory AnnexinV-binding MV were found in blood following RRT [13]. However, it is not known whether this increase reflects secondary systemic effects of RRT or a MV release within the hemofilter.
The aim of this study was to examine the influence of blood membrane contact during a single hemofilter passage in CVVH with citrate anticoagulation on MV levels and composition in patients with severe sepsis.
Methods
Patients
Patients from a tertiary intensive care unit (ICU) that required RRT and fulfilled at least two systemic inflammatory response syndrome (SIRS) criteria [14] in the presence of a suspected or proven sepsis focus were prospectively enrolled in the study from January 2012 to March 2013. Patients were excluded if they were younger than 18 years, moribund, pregnant or breast-feeding. The study was approved by the Ethics Committee from the Medical University Innsbruck (protocol UN 2705a 244/4.20). Subjects provided written informed consent either prior to enrollment or post hoc. Vital parameters were obtained, routine laboratory values measured, as well as acute physiology and chronic health evaluation (APACHE II) score [15], simplified acute physiology score (SAPS II) [16] and sequential organ failure assessment score (SOFA) [17] computed.
Hemofiltration and sample collection
RRT was conducted in CVVH mode by using a ST150 (Gambro Hospital Austria GmbH, Wiener Neudorf, Austria) hemofilter in a Prismaflex® machine (Gambro Hospital Austria GmbH) with postdilution. Phoxilium® was used as substitution fluid. Anticoagulation was performed as regional citrate anticoagulation with Prismocitrate® 18/0, aiming at a postfilter iCa2+ level of 0.35–0.45 mmol/l. Prefilter blood was obtained from a port located after citrate and prefilter substitution fluid influx, and postfilter blood from a port located directly after the hemofilter. Blood was drawn in 3-ml S-Monovette® tubes (Sarstedt, AG & Co., Nümbrecht, Germany) with 3.2% citrate. Ultrafiltrate and blood samples were immediately centrifuged at 20 °C in a Hettich Rotanta 46 RC centrifuge at 1550 g for 15 min with acceleration and deceleration set at 8. The supernatant plasma from blood samples was then further centrifuged at 13000 g for 2 min in a Hettich Micro R22 centrifuge set at maximum acceleration and deceleration, and platelet-free plasma (PFP) obtained. Samples were flash-frozen in liquid nitrogen and stored at −80 °C. Upon analysis samples were thawed at 37 °C in a water bath for 2 min and then kept on wet ice.
Quantification of microvesicles by high-sensitivity flow cytometry
PFP was labeled for 30 min at room temperature with AnnexinV–FITC (1.5 µg/ml [final concentration]) to label phosphatidylserine (PS) on MV, CD31-PE (PECAM; platelet endothelial cell adhesion molecule; 0.39 µg/ml) which is present on MV from endothelial cells, platelets as well as leukocytes and CD42b-APC (GPIbα; glycoprotein Ib alpha; 0.78 µg/ml; all from BD Pharmingen, San Jose, CA) as well as CD41-PC7 (GPIIb; glycoprotein IIb; 0.78 µg/ml; Beckman-Coulter, Miami, FL) to label platelet-derived MV. AnnexinV and antibodies were filtered with a 0.1-µm filter (Millipore, Darmstadt, Germany), and PBS and AnnexinV-binding buffer with a 0.2-µm syringe filter (Sarstedt, Nümbrecht, Germany). Following the addition of CytoCount™ beads (DakoCytomation, Glostrup, Denmark) and 500 µl of AnnexinV-binding buffer (BD Pharmigen) specimens were measured with a Gallios™ flow cytometer with the threshold set at forward scatter (FS) by using Gallios™ Cytometry List Mode Data Acquisition and Analysis Software 1.2 (both Beckman Coulter, Bra, CA). Isotype controls prepared analogously were run in parallel to assess background signals of samples that showed potential artifacts in flow cytometry. These controls were labeled with AnnexinV–FITC and matched irrelevant antibodies of the same isotype and diluted in calcium-free phosphate-buffered saline (PBS from PAA Laboratories, Pasching, Austria) instead of AnnexinV-binding buffer. Artifact-containing parameters were excluded in subsequent analyses (Additional file 1: Fig. S1). Flow cytometric data were analyzed with Kaluza®, version 1.2 (Beckman Coulter). The gating strategy defined MV in a size range between 0.3 and 1.0 µm polystyrene bead-equivalents (LB3 and 89904 from Sigma-Aldrich, Saint Louis, MO) that showed a positivity concerning the aforementioned markers. The number of MV per µl PFP was determined by referring to CytoCount™ beads.
Microvesicle activity and tissue factor measurements
PFP and ultrafiltrate samples were analyzed with three solid-phase-capturing assays as recommended by the manufacturers. The quantity of PS, here referred to as MV activity, which is also one aspect of the procoagulatory potential of MV, was measured with the Zymuphen MP activity assay (Hyphen BioMed, Neuville sur Oise, France). The amount of tissue factor was assessed by using the Imubind® tissue factor ELISA (Sekisu Diagnostics, Stamford, CT). The procoagulatory ability of TF in the presence of PS on MV, here referred to as TF activity, was analyzed with the Zymuphen MP-TF assay (Hyphen BioMed).
Calculation of hemofiltration flow rates and statistical analyses
Absolute amounts of microvesicles entering and leaving the hemofilter were calculated by multiplying the measured MV counts (MV/µl) with the plasma flow or ultrafiltrate flow at the distinct sampling site per minute:
Cmeas—concentration of MV measured at specific port (MV/µl); Amountprefilter—amount of MV entering the hemofilter (MV/min); Amountpostfilter—amount of MV leaving the hemofilter via blood (MV/min); AmountUF—amount of MV leaving the hemofilter via ultrafiltrate (MV/min); BF—blood flow (ml/min); CF—flow of citrate-containing solution (ml/h); UF—ultrafiltrate flow (ml/h), as determined by the Prismaflex® machine; HCT—hematocrit (%); and FF—filtration fraction.
Statistical analyses were performed with GraphPad Prism® version 5 (GraphPad Software Inc., La Jolla, CA) and SPSS® version 23 (IBM, Armonk, NY). Data were tested for normality by using the Shapiro–Wilk test. For comparison of amounts before and after the hemofilter the Wilcoxon signed-rank test was used. Correlations were analyzed in SPSS by using the Spearman rho test. Quantitative data are presented as median and 25th–75th quartiles, if not indicated otherwise. P values below 0.05 were considered statistically significant.
Results
Patients
Twelve eligible patients from a tertiary ICU with severe sepsis or septic shock requiring RRT due to renal failure were included in the study. Blood culture was positive in six patients. Primary sepsis focus was the lung in six patients, urinary tract in two patients and soft tissue in one patient. In three patients no distinct focus could be determined in addition to a positive blood culture. Additional patient characteristics are presented in Table 1.
Table 2 provides an overview of the hemofiltration settings.
Microvesicles and tissue factor in pre- and postfilter blood samples
To examine whether hemofiltration alters counts of several MV subtypes in blood the absolute amounts of MV subtypes entering and leaving the hemofilter per time were compared. Ratios of the amounts of distinct parameters in blood that enter and leave the hemofilter are shown in Table 3.
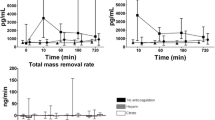
A significant increase in probably mostly leukocyte-derived CD31+/CD41− MV [8.45 × 106 (3.37 × 106 − 2.03 × 107) vs. 1.19 × 107 (5.49 × 106 − 2.53 × 107) MV/min; p = 0.021] (Fig. 1a) was detected after the hemofilter passage. However, no significant differences in counts of platelet-derived CD41+ and CD42b+ MV, AnnexinV+ MV or TF activity (all p > 0.05) were found (Fig. 1b–d, f). There were significant correlations between filtration fraction (FF) and the ratios of post- to prefilter MV (Fig. 2) of AnnexinV+, CD42b+ and CD31+/CD41− MV (Spearman’s correlation coefficient (rs) = 0.683, p = 0.042; rs = 0.664, p = 0.018; rs = 0.720, p = 0.008, respectively).

Postfilter-to-prefilter ratios of MV subsets, MV activity, TF activity and TF level. P values refer to the comparison of absolute amounts per minute pre- to postfilter

Correlations of MV subtypes (post- to prefilter Ratios) with filtration fraction
A measure of the total amount of PS-exposing MV, i.e., MV activity (Fig. 1e), revealed a statistically significant decrease during hemofilter [0.088 (0.027–0.243) vs. 0.067 (0.021–0.223) nmol/min; p = 0.002]. The comparison of TF levels (Fig. 1g) before and after the hemofilter showed a significant decrease after the hemofilter [27877 (22103–37793) vs. 25811 (18736–35164) pg/min, (p = 0.0376) pg/min; p = 0.0093)]. No significant correlation was detected between FF and alterations in MV activity, TF activity or TF level.
There were no correlations between platelet or leukocyte counts and the observed alterations of MV or TF during hemofilter passage. No associations between filter runtime and MV alterations were detected.
Three patients showing artifacts in AnnexinV+ MV counts and one patient with artifacts in measurements of MV activities were therefore excluded from analyses. However, this did not relevantly change statistics.
Microvesicles and tissue factor in ultrafiltrate
To investigate whether MV or fragments of them are ultrafiltrated during hemofilter passage solid-phase-capturing assays were used to examine the ultrafiltrate, such as the MV activity assay to detect MV. This assay captures PS and is therefore able to detect also MV that are sized below the detection limit of high-sensitivity flow cytometry which is approximately 0.3 µm. However, no relevant MV activity could be detected in ultrafiltrate samples (i.e., absolute activities ≤0.05 nM). A remarkable fraction of TF (in average approximately 4.6% of the TF amount that enters the hemofilter) was found in the ultrafiltrate [1229 (408–2218) pg/min, corresponding to an average sieving coefficient of 0.12 (Fig. 1g). However, there was no relevant TF activity measurable in ultrafiltrate (i.e., absolute activities ≤0.05 pg/ml) (Fig. 1f).
Discussion
This is the first study analyzing MV levels as well as TF in blood before and after the hemofilter in CVVH. The major findings of this study are that (1) CVVH clears small PS-exposing MV but not larger MV, (2) subsets of MV are generated within the hemofilter indicating leukocyte activation during hemofilter passage, (3) the extent of MV generation correlates with FF, and (4) a fraction of TF is being cleared via ultrafiltration.
Effects of hemofiltration on microvesicle counts and composition
We could show that CVVH has differential effects on distinct MV subtypes. On the one hand, we observed a reduction in MV activity during hemofilter passage, but no change in AnnexinV+ MV. Although both parameters are measures of PS-exposing MV, they reflect different fractions of PS + MV: MV activity comprises the overall amount of PS + MV, whereas AnnexinV+ MV, measured with hs-FC, exclusively PS + MV that are larger than approximately 0.3 µm. Thus, our data indicate that CVVH clears a fraction small PS-exposing MV. Since no MV activity was measurable in the ultrafiltrate, we assume mechanisms other than filtration such as adsorption being responsible for the clearance of small MV. The findings from our clinical study are compatible with a study from Abdelhafeez et al. [18] who analyzed the effects of CVVH on MV counts in an ex vivo model of CVVH and reported a reduction in MV over a period of 70 min, as shown for endothelium-derived MV. In line with our findings, they propose that mechanisms other than filtration seem to be responsible for the reduction in MV [18]. In conclusion, our data suggest that CVVH clears a fraction of small PS-exposing MV from blood of septic patients.
On the other hand, we found a significantly higher amount of CD31+/CD41− MV after the hemofilter. According to their surface epitope pattern this MV subtype can either be derived from leukocytes or endothelial cells. However, CD31+/CD41− MV that are generated within the hemofilter are probably predominantly leukocyte derived. There are two aspects corroborating this assumption: Firstly, recent data suggest that the majority of MV exposing this not endothelium-specific CD31+/CD41− pattern are predominantly not derived from endothelial cells, but rather from leukocytes in patients with septic shock [19]. Secondly, it is evident that CD31+/CD41− MV that are released during a single hemofilter passage are originating from circulating cells. Despite low levels of circulating endothelial cells (CEC) present in patients with septic shock (approximately between 10 and 30 cells per ml) [20] it is likely that the observed significant increase in CD31+/CD41− MV is rather produced by leukocytes which are around half a million times more abundant than CEC. Our findings and conclusions are in accordance with a study from Kozek-Langenecker et al. [21] who directly assessed activation status of cells. Likewise, they report activation of leukocytes and also platelets within the hemofilter during CVVH with heparin anticoagulation [21]. In line with this, Faure et al. [13] found even higher systemic counts of leukocyte- and platelet-derived MV in patients with chronic renal failure that were dialyzed with heparin anticoagulation compared to patients without dialysis. Moreover, they observed a pronounced systemic increase in CD41+ platelet-derived MV and also in AnnexinV+ MV after the hemodialysis session [13]. We could not find increased levels of platelet-derived MV after hemofilter passage. This may be explained by the use of regional citrate anticoagulation in all patients, which has previously been demonstrated to result in significantly less platelet activation than heparin anticoagulation [6, 7].
Correlation of filtration fraction with MV generation
Interestingly, we found correlations between FF and increments of platelet- and presumably leukocyte-derived as well as procoagulant AnnexinV+ MV. The usually recommended FF is below 25% in postdilution mode [22], since higher FF frequently leads to hemofilter clotting [5]. As shown in Fig. 2, the induction of MV generation within the hemofilter seems to be more pronounced when FF is above 30%, probably due to increased shear stress at higher FF. This prominent effect at higher FF seems even to apply to platelet-derived and AnnexinV+ MV which did not significantly change in septic patients overall. We speculate that the generation of MV within the hemofilter might contribute to hemofilter clotting occurring when high FF is applied. However, this remains to be investigated in a larger study.
Mechanisms contributing to the presumed cellular activation and the subsequent MV generation within the hemofilter might include a high shear stress or contact with the hemofilter membrane [5–7, 23–25]. Such effects of CVVH might even be more pronounced in a septic state where cells are already preactivated or highly susceptible to activation. Accordingly, septic shock patients exhibit already threefold higher levels of presumably mostly leukocyte-derived CD31+/CD41− MV compared to healthy subjects, as recently reported [19]. As calcium is a key mediator for neutrophil degranulation that is induced by dialysis membranes [23] and as influx of extracellular calcium is required for vesiculation of platelets induced by shear stress [24] one might speculate that MV release might even be more pronounced in CVVH without citrate anticoagulation [6], but this needs to be tested.
Clearance of tissue factor by hemofiltration
Subsets of leukocytes, such as monocytes, are able to release MV and also associated TF upon activation [26, 27]. Based on the assumption that the above-mentioned increase in CD31+/CD41− predominantly reflects leukocyte activation we wanted to investigate whether we can find evidence for an accompanying TF generation during CVVH. Former studies reporting TF levels during hemofiltration analyzed exclusively systemic TF levels [28, 29]. Although Cardigan et al. [28] observed an increase in 50% of patients while on CVVH, no significant systemic increase could be detected in the most recent study from Bouman et al. [29]. So far, it was unclear whether potential alterations in systemic TF levels reflect secondary systemic inflammatory effects that are triggered by hemofiltration or if these alterations are due to TF generation or clearance within the hemofilter. Our study adds two new key answers to this matter: First, we provide evidence that hemofilter passage leads to a reduction in TF levels. Additionally, we demonstrate that a fraction of TF is being ultrafiltrated.
Strengths and limitations
A major strength of our study is that we assessed alterations in MV levels occurring directly and exclusively within the hemofilter. Limitations of the study are the relatively small number of patients as well as the lack of information concerning kinetics in the extracorporeal system or the systemic circulation because we did not take serial measurements due to the complexity of analysis. Since no conclusion can be drawn regarding whether the generation of CD31+/CD41− MV within the hemofilter may impact systemic levels or patient outcome, the clinical implications of MV generation within the hemofilter remain to be further elucidated.
Conclusions
In conclusion, CVVH clears a fraction of small PS-exposing MV by mechanisms other than filtration, but does not eliminate larger MV. CVVH induces the release of CD31+/CD41− MV that indicate leukocyte activation during hemofilter passage in septic patients. The generation of MV within the hemofilter seems to be particularly pronounced when FF is above 30%, which supports the clinical relevance of the usual recommendation to keep FF below 25%. A fraction of TF is being cleared by CVVH via ultrafiltration. Our findings show that MV analysis can be used to assess the effects caused by contact of blood with the hemofilter. Evaluation of MV patterns might be a promising instrument helping to refine extracorporeal therapeutic systems and biocompatibility of hemofilter membranes in future.
Abbreviations
- CVVH:
-
continuous veno-venous hemofiltration (CVVH)
- RRT:
-
renal replacement therapy
- MV:
-
microvesicles
- ICU:
-
intensive care unit
- SIRS:
-
systemic inflammatory response syndrome
- APCAHE II:
-
acute physiology and chronic health evaluation score
- SAPS II:
-
simplified acute physiology score
- SOFA:
-
sequential organ failure assessment score
- PFP:
-
platelet-free plasma
- PECAM:
-
platelet endothelial cell adhesion molecule
- GPIbα:
-
glycoprotein Ib alpha
- GPIIb:
-
glycoprotein IIb
- PBS:
-
phosphate-buffered saline
- CRP:
-
C-reactive protein
- PCT:
-
procalcitonin
- CEC:
-
circulating endothelial cells
- TAT:
-
thrombin–antithrombin
- FVII:
-
factor VII
- C meas :
-
concentration of MV measured at specific port (MV/µl)
- Amountprefilter :
-
amount of MV entering the hemofilter (MV/min)
- Amountpostfilter :
-
amount of MV leaving the hemofilter via blood (MV/min)
- AmountUF :
-
amount of MV leaving the hemofilter via ultrafiltrate (MV/min)
- BF:
-
blood flow (ml/min)
- CF:
-
flow of citrate-containing solution (ml/h)
- UF:
-
ultrafiltrate flow (ml/h)
- FF:
-
filtration fraction
References
Jörres, A. Acute kidney injury: choice of the initial modality for renal replacement therapy. Med Klin Intensivmed Notfmed. 2015;110(4):251–5.
Payen D, Mateo J, Cavaillon JM, Fraisse F, Floriot C, Vicaut E, et al. Impact of continuous venovenous hemofiltration on organ failure during the early phase of severe sepsis: a randomized controlled trial. Crit Care Med. 2009;37(3):803–10.
Honore PM, Jamez J, Wauthier M, Lee PA, Dugernier T, Pirenne B, et al. Prospective evaluation of short-term, high-volume isovolemic hemofiltration on the hemodynamic course and outcome in patients with intractable circulatory failure resulting from septic shock. Crit Care Med. 2000;28(11):3581–7.
De Vriese AS, Colardyn FA, Philippe JJ, Vanholder RC, De Sutter JH, Lameire NH. Cytokine removal during continuous hemofiltration in septic patients. J Am Soc Nephrol. 1999;10(4):846–53.
Joannidis M, Oudemans-van Straaten HM. Clinical review: patency of the circuit in continuous renal replacement therapy. Crit Care. 2007;11(4):218.
Gritters M, Grooteman MP, Schoorl M, Schoorl M, Bartels PC, Scheffer PG, et al. Citrate anticoagulation abolishes degranulation of polymorphonuclear cells and platelets and reduces oxidative stress during haemodialysis. Nephrol Dial Transplant. 2006;21(1):153–9.
Oudemans-van Straaten HM, Kellum JA, Bellomo R. Clinical review: anticoagulation for continuous renal replacement therapy–heparin or citrate? Crit Care. 2011;15(1):202.
Oudemans-van Straaten HM. Hemostasis and thrombosis in continuous renal replacement treatment. Semin Thromb Hemost. 2015;41(1):91–8.
Thery C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–93.
Gyorgy B, Szabo TG, Pasztoi M, Pal Z, Misjak P, Aradi B, et al. Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell Mol Life Sci. 2011;68(16):2667–88.
Mastronardi ML, Mostefai HA, Meziani F, Martinez MC, Asfar P, Andriantsitohaina R. Circulating microparticles from septic shock patients exert differential tissue expression of enzymes related to inflammation and oxidative stress. Crit Care Med. 2011;39(7):1739–48.
Delabranche X, Boisrame-Helms J, Asfar P, Berger A, Mootien Y, Lavigne T, et al. Microparticles are new biomarkers of septic shock-induced disseminated intravascular coagulopathy. Intensive Care Med. 2013;39(10):1695–703.
Faure V, Dou L, Sabatier F, Cerini C, Sampol J, Berland Y, et al. Elevation of circulating endothelial microparticles in patients with chronic renal failure. J Thromb Haemost. 2006;4(3):566–73.
Bone RC, Balk RA, Cerra FB, Dellinger RP, Fein AM, Knaus WA, et al. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest. 1992;101(6):1644–55.
Knaus WA, Draper EA, Wagner DP, Zimmerman JE. APACHE II: a severity of disease classification system. Crit Care Med. 1985;13(10):818–29.
Le Gall JR, Lemeshow S, Saulnier F. A new simplified acute physiology score (SAPS II) based on a European/North American multicenter study. JAMA. 1993;270(24):2957–63.
Vincent JL, Moreno R, Takala J, Willatts S, De Mendonca A, Bruining H, et al. The SOFA (Sepsis-related Organ Failure Assessment) score to describe organ dysfunction/failure. On behalf of the Working Group on sepsis-related problems of the European Society of Intensive Care Medicine. Intensive Care Med. 1996;22(7):707–10.
Abdelhafeez AH, Jeziorczak PM, Schaid TR, Hoefs SL, Kaul S, Nanchal R, et al. Clinical CVVH model removes endothelium-derived microparticles from circulation. J Extracell Vesicles. 2014;3(1):23498. doi:10.3402/jev.v3.23498.
Lehner GF, Harler U, Haller VM, Feistritzer C, Hasslacher J, Dunzendorfer S, et al. Characterization of microvesicles in septic shock using high-sensitivity flow cytometry. Shock. 2016;46(4):373–81.
Mutunga M, Fulton B, Bullock R, Batchelor A, Gascoigne A, Gillespie JI, et al. Circulating endothelial cells in patients with septic shock. Am J Respir Crit Care Med. 2001;163(1):195–200.
Kozek-Langenecker SA, Spiss CK, Michalek-Sauberer A, Felfernig M, Zimpfer M. Effect of prostacyclin on platelets, polymorphonuclear cells, and heterotypic cell aggregation during hemofiltration. Crit Care Med. 2003;31(3):864–8.
Klein SJ, Joannidis M. Renal replacement therapy in acute kidney injury. Med Klin Intensivmed Notfmed. 2017;112(5):437–443.
Bohler J, Donauer J, Birmelin M, Schollmeyer PJ, Horl WH. Mediators of complement-independent granulocyte activation during haemodialysis: role of calcium, prostaglandins and leukotrienes. Nephrol Dial Transplant. 1993;8(12):1359–65.
Miyazaki Y, Nomura S, Miyake T, Kagawa H, Kitada C, Taniguchi H, et al. High shear stress can initiate both platelet aggregation and shedding of procoagulant containing microparticles. Blood. 1996;88(9):3456–64.
Lehner GF, Schopf M, Harler U, Pechlaner C, Joannidis M. Repeated premature hemofilter clotting during regional citrate anticoagulation as indicator of heparin induced thrombocytopenia. Blood Purif. 2014;38(2):127–30.
Egorina EM, Sovershaev MA, Olsen JO, Osterud B. Granulocytes do not express but acquire monocyte-derived tissue factor in whole blood: evidence for a direct transfer. Blood. 2008;111(3):1208–16.
Key NS, Mackman N. Tissue factor and its measurement in whole blood, plasma, and microparticles. Semin Thromb Hemost. 2010;36(8):865–75.
Cardigan RA, McGloin H, Mackie IJ, Machin SJ, Singer M. Activation of the tissue factor pathway occurs during continuous venovenous hemofiltration. Kidney Int. 1999;55(4):1568–74.
Bouman CS, de Pont AC, Meijers JC, Bakhtiari K, Roem D, Zeerleder S, et al. The effects of continuous venovenous hemofiltration on coagulation activation. Crit Care. 2006;10(5):R150.
Authors’ contributions
GFL designed and coordinated the study, enrolled patients, carried out the analyses and wrote the manuscript. UH enrolled patients, participated in the coordination of the study and refinement of the manuscript. CF participated in the design of the study, the analysis and refinement of the manuscript. VMH took measurement of microvesicles and participated in the refinement of the manuscript. JH participated in the design of the study. RB participated in the enrollment of patients, calculation of hemofilter flow rates and refinement of the manuscript. MJ designed and coordinated the study, enrolled patients and refined the manuscript.
Acknowledgements
None.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Consent for publication
Not applicable.
Ethics approval and consent to participate
The study was approved by the Ethics Committee from the Medical University Innsbruck (protocol UN 2705a 244/4.20). Subjects provided written informed consent either prior to enrollment or post hoc.
Funding
Supported by funds of the Oesterreichische Nationalbank (Oesterreichische Nationalbank, Anniversary Fund, project numbers 13861 and 15708).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional file
13613_2017_312_MOESM1_ESM.pdf
Additional file 1: Fig. S1. Workflow showing excluded parameters (filter survival and experimental data) relevant to final analyses. CVVH = Continuous veno-venous hemofiltration; MV activity = Microvesicle activity.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Lehner, G.F., Harler, U., Feistritzer, C. et al. Hemofiltration induces generation of leukocyte-derived CD31+/CD41− microvesicles in sepsis. Ann. Intensive Care 7, 89 (2017). https://doi.org/10.1186/s13613-017-0312-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13613-017-0312-3