
Abstract
Schistosomiasis is a neglected tropical disease that affects humans and animals in tropical and subtropical regions worldwide. Schistosome eggs are responsible for the pathogenesis and transmission of schistosomiasis, thus reducing egg production is vital for prevention and control of schistosomiasis. However, the mechanisms underlying schistosome reproduction remain unclear. Annexin proteins (ANXs) are involved in the physiological and pathological functions of schistosomes, but the specific regulatory mechanisms and roles of ANX A13 in the development of Schistosoma japonicum and host–parasite interactions remain poorly understood. Therefore, in this study, the expression profiles of SjANX A13 at different life cycle stages of S. japonicum were assessed using quantitative PCR. In addition, the expression profiles of the homolog in S. mansoni were analyzed in reference to public datasets. The results of RNA interference showed that knockdown of SjANX A13 significantly affected the development and egg production of female worms in vivo. The results of an immune protection assay showed that recombinant SjANX A13 increased production of immunoglobulin G-specific antibodies. Finally, co-culture of S. japonicum exosomes with LX-2 cells using a transwell system demonstrated that SjANX A13 is involved in host–parasite interactions via exosomes. Collectively, these results will help to clarify the roles of SjANX A13 in the development of S. japonicum and host–parasite interactions as a potential vaccine candidate.
Similar content being viewed by others

Introduction
Schistosomiasis is a zoonotic parasitic disease that affects approximately 290 million people globally and is responsible for 1.4–3.3 million disability-adjusted life years annually [1, 2]. Among the three main pathogenic species of schistosomes, Schistosoma japonicum infects more than 40 kinds of animals, resulting in significant economic losses to the livestock industry [3]. Livestock is recognized as a key contributor to animal schistosomiasis in Asia [4]. Due to the large amount of feces produced by livestock and high degree of environmental overlap between humans and livestock during agricultural production, schistosome eggs released by infected animals threaten human health [5]. The increasing attention to animal schistosomiasis over the past few decades in China has facilitated significant improvements in agricultural practices [6]. National surveillance of schistosomiasis-related morbidity in China has detected no case of S. japonicum infection of livestock from 2017 to 2022 [7,8,9,10]. However, vast areas of China are suitable habitats for snails, which serve as intermediate hosts for schistosomes. Southeast Asian countries neighboring China, as well as some developing countries in Africa, are still suffering from severe schistosomiasis [11, 12]. In addition, the World Health Organization recommends praziquantel (PZQ) for treatment of schistosomiasis, which has led to the emergence of drug-resistant phenotypes, thereby further complicating prevention and control of schistosomiasis [13, 14]. Therefore, the development of vaccines or to explore novel drug targets against schistosomiasis is still necessary on a global basis.
Unlike most trematodes, which are hermaphroditic, schistosomes are dioecious, presenting as separate male and female sexes [15]. The sexual development and maintenance of the mature reproductive status of female worms require perpetual coupling with males [16]. Maturation and egg production lead to the pathogenesis and transmission of schistosomiasis. Hence, elucidation of the molecular mechanisms of the pathogenesis and transmission is vital for prevention and control of schistosomiasis. A proteomic study conducted by our group identified several highly expressed proteins involved in reproductive development of mated female (MF) worms and single-sex infected female (SF) worms at 18, 21, 23, and 25 days (four important time points from sexual maturation to egg production) and various recombinant proteins with immunoprotective effects [17, 18]. Subsequent analysis of S. japonicum identified several highly expressed proteins, including annexin A13 (SjANX A13), in MF worms at all four time points.
Multifunctional ANX proteins (ANXs) are expressed by a wide range of parasites [19,20,21]. Recent omics studies have identified several ANXs in the gut, vitelline gland, and tegument of S. mansoni [22,23,24], S. japonicum [25] and S. bovis [26]. Tissue-specific transcriptomic analyses of female S. japonicum found that SjANX B7 and B22 were expressed in the gut lining, while SjANX B5 was expressed in the vitelline gland, suggesting potential developmental roles in females [27]. Transcriptional profiling revealed that expression of SmANX B22 and B30 during schistosome development was correlated with life cycle stages characterized by the presence of the syncytial tegument layer [24, 28,29,30]. Besides, the native ANX protein of S. bovis (SbANX) was expressed on the tegument surface of the schistosomula and adult [26]. ANXs are frequently identified in the tegument and present important antigen sources for anti-schistosome vaccines and promising targets for drug development [31]. Although many ANXs have been identified in schistosomes, SjANX A13 is a promising vaccine candidate due to roles in the growth and development of S. japonicum.
In addition, schistosome ANXs are components of excretory/secretory products [32] and exosomes [33], which may mediate the secretion of anticoagulant ANXs and restrict host hemostatic responses to facilitate blood feeding. Thus, the identification and characterization of ANXs released from parasite excretory/secretory products or exosomes and crucial interactions with host cells could help to clarify host–parasite interactions.
Here, the molecular features of SjANX A13 and transcriptional levels at different maturation stages were assessed. RNA interference (RNAi) was conducted to explore the functional roles of SjANX A13 in growth, development, and fecundity of S. japonicum. In addition, the protective immunity of recombinant SjANX A13 (rSjANX A13) was assessed in a mouse model. Finally, the potential involvement of SjANX A13 in host–parasite interactions through exosomes was investigated.
Materials and methods
Parasites and animals
Specific-pathogen-free (SPF) male BALB/c mice (6–7 weeks old; body weight 18 ± 2 g) were purchased from Shanghai Jiesijie Laboratory Animal Co., Ltd. (Shanghai, China) and housed in SPF-grade animal rooms at the Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Shanghai, China).
The Chinese strain of S. japonicum cercariae used in this study was obtained from the National Reference Laboratory for Animal Schistosomiasis, Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Shanghai, China). BALB/c mice were percutaneously infected by applying cercariae to the shaved skin of the abdomen. Depending on different experiments, animals were euthanized and worms were collected at specific time points through hepatic-portal perfusion as previously described [34].
Sequence analysis of SjANX A13
Based on the amino acid sequence of SjANX A13 (GenBank no: CAX70812.1), the cDNA sequence of this protein was found in the S. japonicum genome sequence. Protein domains were analyzed by the Simple Modular Architecture Research Tool (SMART) [35]. The physicochemical properties (molecular weight, isoelectric point, transmembrane region, signal peptide) of SjANX A13 were analyzed using the Detaibio tool [36]. Alignments of multiple ANXs with the highest homology in other species included Schistosoma mansoni (XP_018652854.1), Schistosoma haematobium (XP_035587885.1), Homo sapiens (AAI25159.1) and Mus musculus (NP_081487.1) were performed using the ClustalX 1.83 program [37].
Phylogenetic analysis of SjANX A13
ANX A13 protein sequences of multiple species were acquired from the database of National Center for Biotechnology Information (NCBI). Amino acid sequences of ANX with the highest homology in other species including Schistosoma like Schistosoma bovis (CAH8605973.1), Schistosoma guineensis (CAH8598563.1), Schistosoma intercalatum (CAH8585262.1), Schistosoma mattheei (CAH8576064.1), Schistosoma margrebowiei (CAH8603292.1), Schistosoma rodhaini (CAH8645073.1) and Schistosoma spindale (CAI2731872.1); other trematodes (except Schistosoma) like Fasciola.hepatica (THD28190.1), Opisthorchis felineus (TGZ72241.1) and Clonorchis sinensis (GAA48684.1); cestoda like Echinococcus granulosus (KAH9278638.1) and Taenia solium (AAY17503.1); nematodes like Caenorhabditis elegans (CAE45742.1); mammals like Homo. sapiens (AAI25159.1) Mus. musculus (NP_081487.1) and R.norvegicus (NP_001128382.1) were used to construct the phylogenetic tree by MEGA-X software with the maximum likelihood method, then improved by the iTOL online tool [38].
Molecular modelling and evaluation of SjANX A13
The 3-dimensional (3D) structure for SjANX A13 was modelled by homology modelling method using the SWISS-MODEL [39]. The protein modelling was performed based on the reference sequence of crystal structure of calcium-bound annexin (Sm)1 as a template (Uniprot accession number: C4QH88) [40]. The validation of the 3D model was assessed using Ramachandran plots [41], QMEANDisCo global and GMQE estimate [42].
Bioinformatics analysis of SjANX A13
To get a general overview of the SjANX A13 expression in publicly available datasets, a volcano plot was presented to illustrate the SjANX A13 expression level in our previous proteomics data focused on 18, 21, 23 and 25-day SF and MF (dataset identifier PXD030834) [17]. In addition, the expression data of SmANX (GenBank no: XP_018652854.1, also referred as Smp_074140 obtained from WormBase ParaSite [43]) in a publicly RNAseq dataset [44] and a single-cell RNAseq (scRNAseq) dataset [45] of S. mansoni were taken as a reference for the expression of SjANX A13. A heatmap representing the Smp_074140 gene expression throughout life stage (egg, miracidium, sporocyst, cercaria, schistosomulum, 21-day juvenile, 28-day juvenile, 35-day adult, 38-day adult, adult older than 42-day) of mated male (MM), single-sex infected male (SM), MF and SF was constructed.
In addition, to explore the potential interacting molecules of SjANX A13, the online tool STRING v11.5 was used to construct a predicted interaction network [46]. Gene ontology (GO) [47] were performed to assess the molecular function, cellular component and biological process of the predicted interacting molecules, and Kyoto Encyclopedia of Genes (KEGG) [48] analysis were conducted to allocate proteins into metabolic and regulatory pathways.
Expression examination of SjANX A13 in life cycle by qPCR
To collect SF from different life stages, the sexes of S. japonicum cercariae were first identified by PCR based on previous publication [49]. After determining the sexes of cercariae, mice were infected with unisexual cercariae percutaneously to obtain SF through hepatic-portal perfusion at different life stages. To collect MM and MF, mice were infected with bisexual cercariae and worms were collected and separated manually at each time point, except for 7-day and 14-day worms.
All worms were washed three times in phosphate buffer solution (PBS, Corning, USA) at 4 °C to remove residual host debris and total RNA was then extracted by TRIzol reagent (Invitrogen, USA) according to the manufacturer’s instructions [50]. The reverse-transcription was performed using a Hifair® III 1st Strand cDNA Synthesis SuperMix for qPCR kit (Yeasen, China) and the resulting cDNA was used as template for qPCR with Hieff® qPCR SYBR Green Master Mix (Yeasen, China). The relative mRNA expression levels of genes were quantified with β-tubulin served as an endogenous control. The LightCycler 96 system (Roche, China) was used for qPCR analysis. The cycling conditions were as follows: preincubation, 95 °C for 60 s; 2 step amplification, 95 °C for 5 s, and 60 °C for 30 s, for 40 cycles; melting, 95 °C for 10 s, 65 °C for 60 s, 97 °C for 1 s. All samples were assessed in triplicate. Subsequently, the relative transcript levels of SjANX A13 were analyzed using the 2−ΔΔCt method [51]. The primers used in the qPCR analysis are listed in Table 1.
RNA interference (RNAi) assay in vivo
For biological characteristic analysis of SjANX A13, RNAi was performed using the method as described previously with modification [17, 52, 53]. Mice were exposed to approximately 200 cercariae and at 22 days post-infection (dpi), three specific siRNA (S1, S2 and S3) pairs and a non-specific (negative control, NC) siRNA pair, dissolved in 200 µL diethyl pyrocarbonate (DEPC)-treated water with the concentration of 100 nM, or 200 µL PBS were injected via the tail vein. After 48 h, worms were collected and the interference efficiency of these siRNA pairs was evaluated through qPCR.
After the interference efficiency was compared, SjANX A13 S1 was selected for long-term interference assay to assess the effect of SjANX A13 on the development and reproduction of S. japonicum. Mice were exposed to 40 cercariae, and SjANX A13 S1, NC siRNA (dissolved in 200 µL DEPC-treated water) or 200 µL PBS were given via the tail vein every 4 days since 4 dpi. Mice were sacrificed at 42 dpi, and worms were collected and counted. Part of worms were applied for interference efficiency assay with qPCR and the rests were prepared for morphological observation. In addition, livers were collected with the calculation of liver egg count and the liver egg-hatching rate according to previous studies [54, 55].
The siRNA pairs used in this study were synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China) and the detailed sequences of these siRNA pairs are shown in Table 2.
Morphological observation by scanning electron microscopy (SEM)
MM and MF worms from the long-term interference assay were washed with PBS for three times (15 min/time), and then fixed in 1% osmium tetroxide for 2 h, and then dehydrated in a graded ethanol series of 30%, 50%, 70%, 80%, 90%, 95%, and 100% for 20 min each. After desiccation in a CO2 critical point drying apparatus, the dehydrated worms were then coated with a gold film in a vacuum-coating device and then examined under a JSM-6380LV scanning electron microscope (JEOL, Ltd., Tokyo, Japan).
Expression and purification of recombinant SjANX A13
Primers of SjANX A13 were designed based on its nucleotide sequence (Genbank no: FN315080.1) using the corresponding restriction enzyme sites of BamHI and XhoI at the N-terminus and C-terminus, respectively. The verified SjANX A13 cDNA fragment was amplified and ligated into the expression vector pET-28a(+) (Novagen, Germany). Products were transformed into Escherichia coli BL21 (DE3) cells (Invitrogen, USA), and recombinant clones were obtained by antibiotic selection. The recombinant proteins were overexpressed in the presence of isopropyl-b-d-thiogalactopyranoside (IPTG). Transformed cells were grown in Luria broth (LB) with 1 mg/mL kanamycin at 37 °C until OD600nm = 0.7, and IPTG was added to the culture at a final concentration of 1 mM. After 8 h induction, cells were harvested and the expression of recombinant protein was analyzed by SDS-PAGE. The histidine-tagged fusion recombinant protein was then purified from E. coli lysates by metal affinity chromatography using Ni-NTA His·Bind Resin Chromatography according to the manufacturer’s instructions (Novagen, China). The purified recombinant SjANX A13 (rSjANX A13) protein was analyzed by SDS-PAGE and the protein concentration was measured by a bicinchoninic acid (BCA) assay kit (Yeasen, China). The primers used in this study are listed in Table 3.
Vaccination, immune response assays and evaluation of immune protection
To explore whether SjANX A13 provides immune-protective effect, an immune protection trial was conducted. Mice were randomly distributed into three groups (ten mice each). Mice in the vaccination group were injected subcutaneously with rSjANX A13 emulsified with ISA206 adjuvant (Seppic, France) (50 µg/100 µL/mouse). Following this, the immunity of the mice was enhanced twice at intervals of 2 weeks with 25 µg rSjANX A13/mouse. Each mouse in the adjuvant and control groups was injected with 100 µL ISA206 adjuvant in PBS or 100 µL PBS only, respectively. Fourteen days after the last vaccination, all mice were challenged percutaneously with 40 S. japonicum cercariae and mice were sacrifice 42 days after challenge. Worms were collected and counted and blood samples were collected from mice in each group by retro-orbital bleeding [56]. Specific IgG antibodies against SjANX A13 were detected by ELISA in the serum, as described in a previous study [17].
Cell line and cell culture
Human hepatic stellate cell (HSC) line LX-2 was obtained from Boster company (Wuhan, China) and cultured in the Dulbecco’s modified Eagle’s medium (DMEM, Corning, USA), supplemented with 10% heat-inactivated fetal bovine serum (FBS, Gibco, USA), 1% penicillin-streptomycin (Thermo Fisher Scientific, USA) in a humidified incubator at 37 °C with 5% CO2.
Construction of transwell model
Mice were percutaneously infected with approximately 100 cercariae and worms were collected from the infected mice at 28 dpi. Worms were thoroughly and gently washed three times with 30 mL PBS and then maintained in preheated DMEM. In each transwell system of 12-well plate (PET membrane, pore size 0.4 μm) (Corning, USA), LX-2 cells (5 × 105 cells/well) were placed in the lower well with 1.5 mL DMEM containing 10% FBS and 1% penicillin-streptomycin. Five pair adult worms were transferred from the previous DMEM culture and placed in the upper well with 900 µL fresh DMEM containing 10% FBS and 1% penicillin–streptomycin. A transwell insert with only an unused schistosomal medium was utilized as a control. Each transwell system had at least three technical replicates and were cultured in a humidified incubator at 37 °C with 5% CO2. Cells were harvested for qPCR analysis 48 h after co-incubation was established. The relative mRNA expression of SjANX A13 were quantified with GAPDH served as an endogenous control.
Isolation and characterization of S. japonicum exosomes
The isolation of S. japonicum exosomes was performed as previous described with modification [33, 57]. Briefly, New Zealand rabbits were percutaneously infected with approximately 3000 cercariae and worms were collected through hepatic-portal perfusion at 28 dpi. Worms were gently washed 3–5 times with 50 mL PBS and the dead or fragmentary ones were discarded. The remaining worms were then maintained in preheated RPMI-1640 culture medium (Corning, USA) containing 1% penicillin-streptomycin at 37 °C under 5% CO2 at a density of ~15 worm pairs/mL for 2 h. After 2 h incubation, the supernatant was collected and fresh culture medium was added for the next collection (the whole collecting procedure could last for 3–4 days until the worms were less active). The pellets in the collected supernatant were discarded by centrifugation at 2000 × g and 14 000 × g for 30 min each at 4 °C, respectively. Then, the supernatant was collected and dialyzed in PBS for 24 h at 4 °C followed by centrifugal ultrafiltration through a 3K Omega membrane (Pall, USA). The supernatant was then filtered using a 0.22 μm syringe filter (Pall, USA) and a total exosome isolation kit (Thermo Fisher Scientific, USA) was used according to the manufacturer’s instructions. The exosome pellet was resuspended in 200 µL of PBS and then stored at −80 ℃ until further analysis. Each time before proceeding with downstream experiments, S. japonicum exosomes were subjected to transmission electron microscopy (TEM) analysis to confirm their morphology, and the concentration of S. japonicum exosomes was measured using a BCA assay kit according to the manufacturer’s instructions.
Exosomes uptake experiment
The isolated S. japonicum exosomes were labeled with PKH67 using a Green Fluorescent Labeling Kit (Sigma Aldrich, USA), and the procedures were performed according to the manufacturer’s protocol. Briefly, 10 µg S. japonicum exosomes were stained with PKH67 dye in 500 µL of Diluent C fluid for 5 min at room temperature. Next, 1 mL 1% bull serum albumin (BSA, Yeasen, China) was added to stop the labeling process. Then, the labeled exosomes were re-purified via ultracentrifugation at 100 000 × g with PBS rinsing for 90 min. As a control for non-specific labeling of cells, PBS was PKH67-stained, washed, and added to the cells as a parallel experiment. The whole procedure was conducted at 4 °C. Then, the PKH67-labeled exosomes were co-incubated with LX-2 for 2 h in a humidified incubator at 37 °C with 5% CO2. Afterward, the culture medium was discarded, and the cells were washed in PBS three times, fixed with 4% formaldehyde solution (Servicebio, China) for 15 min and washed twice more with PBS, nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI, Sigma Aldrich, USA) for 3 min. After washed in PBS three times to remove the remaining DAPI, the cells were observed using a fluorescence microscopy (Olympus, Japan). In addition, to detect the expression of SjANX A13 in LX-2 cells after co-incubated with S. japonicum exosomes, cells were harvested for qPCR analysis 48 h after co-incubation. The relative mRNA expression of SjANX A13 gene were quantified with GAPDH served as an endogenous control.
Statistical analysis
Data were analyzed with SPSS 25.0 software (SPSS Inc., USA) and expressed as mean ± standard deviation (SD) of three independent biological replicates. Data were statistically analyzed with Student’s t-tests. A P-value of < 0.05 was considered statistically significant in statistical analysis.
Results
In silico characterization of SjANX A13
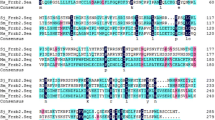
SjANX A13 (molecular weight, 39.62 kDa; isoelectric point, 5.11) consisted of 1253 bp and encoded 354 amino acid residues. Analysis with the SMART, which is used for the identification and annotation of genetically mobile domains and analysis of domain architectures, showed that SjANX A13 has four conserved repeat domains, each consisting of approximately 60 amino acids containing a calcium (Ca+)-binding motif (amino acids 43–95, 115–175, 221–274, and 296–348) (Figure 1A). In addition, sequence analysis revealed that SjANX A13 had no signal peptide structure and the whole protein was expressed extracellularly, therefore lacking a transmembrane helix, as confirmed by a model to predict membrane protein topology based on a hidden Markov model (Additional file 1). Sequence alignment showed that SjANX A13 shared 74.01%, 77.05%, 43.96%, and 44.27% identity with orthologs in S. mansoni, S. haematobium, H. sapiens, and M. musculus, respectively (Figure 1B).

Domain and multiple sequences alignment of SjANX A13. A Protein domains of SjANX A13. B Comparison of amino acid sequences of SjANX A13 with associated sequences from other species, including Schistosoma mansoni (XP_018652854.1), Schistosoma haematobium (XP_035587885.1), Homo sapiens (AAI25159.1) and Mus musculus (NP_081487.1).
Phylogenetic analysis of SjANX A13
ANX proteins of other species acquired from NCBI database were used to construct a phylogenetic tree, which showed that the Schistosoma genus has one clade and SjANX A13 is closely related to orthologs of all schistosome species (Figure 2).

Phylogenetic analysis of SjANX A13 and its homologues. The sequence ID consists of the Genbank no.+ species name. Protein sequences were divided into five groups that are color-coded (see list to the top left of figure): Schistosoma like Schistosoma bovis (CAH8605973.1), Schistosoma guineensis (CAH8598563.1), Schistosoma intercalatum (CAH8585262.1), Schistosoma mattheei (CAH8576064.1), Schistosoma margrebowiei (CAH8603292.1), Schistosoma rodhaini (CAH8645073.1) and Schistosoma spindale (CAI2731872.1); other trematodes (except Schistosoma) like Fasciola hepatica (THD28190.1), Opisthorchis felineus (TGZ72241.1) and Clonorchis sinensis (GAA48684.1); cestoda like Echinococcus granulosus (KAH9278638.1) and Taenia solium (AAY17503.1); nematodes like Caenorhabditis elegans (CAE45742.1); mammals like Homo. sapiens (AAI25159.1) Mus. musculus (NP_081487.1) and R.norvegicus (NP_001128382.1).
Molecular modelling and evaluation
The 3D structure of SjANX A13 was obtained based on the crystal structure of calcium-bound annexin (Sm)1 as a template (UniProt accession number: C4QH88). There was no significant difference in the conformation of the two molecules of the dimer (Figure 3A). The predicted model obtained with the SWISS-MODEL homology modelling pipeline had a Qualitative Model Energy Analysis-Distance Constraint (QMEANDisCo) global score of 0.75 ± 0.05 and Global Model Quality Estimation (GMQE) score of 0.78, where higher QMEANDisCo global and GMQE scores (range 0–1) indicate higher expected quality (Figure 3B). Ramachandran plot analysis revealed that the protein model had 96.85% of residues in most favored regions, 0.29% in allowed regions, and only 0.49% in disallowed regions (Figure 3C). Collectively, these findings indicate that the SjANX A13 model had correct topology with high expected quality.

Validation results of the refined 3D structure of SjANX A13. A A 3D structure predicted for SjANX A13 (the structure was presented by “confidence” color, blue: confident, yellow: low, orange: very low) B The average per-residue QMEANDisCo score. C Ramachandran plot of the model showed that 96.85%, 0.29% and 0.49% of residues were located in the favored, allowed and disallowed regions, respectively.
Bioinformatics analysis of SjANX A13
To obtain a comprehensive profile of SjANX A13 expression in the publicly available dataset, the proteomics data included in a previous report by our group was analyzed by mainly focusing on SF and MF worms at 18, 21, 23, and 25 days [17]. In this dataset, SjANX A13 expression was relatively higher in MF worms at all four time points, suggesting a potential role in female development (Figure 4A). In addition, the expression profiles of the S. mansoni homolog (Smp_074140) of SjANX A13 were analyzed in references to more well-established RNAseq [44] and scRNAseq [45] datasets. In terms of the life history of S. mansoni, SmANX (Smp_074140) was highly expressed in single-sex cercariae and mated male (MM) worms at 35 and > 42 days post-infection (dpi). In addition, Smp_074140 expression was generally higher in males than females at the same developmental time point, both in unisexual and bisexual infections (Figure 4B). Further comparison of unisexual- and bisexual-infected worms found that Smp_074140 expression was higher in SM than MM worms, with the exception of 42 dpi at the late maturation stage, while expression was generally higher in MF than SF worms, with the exception of 35 and 42 dpi, which was similar to the proteomic data of S. japonicum females mentioned above, as expression at 18–25 dpi in S. japonicum was almost identical to that at 28–35 dpi in S. mansoni [58] (Figure 4B).

Bioinformatics analysis of SjANX A13. A Volcano plots showing the SjANX A13 protein expression in 18, 21, 23 and 25-day MF verses SF proteome. B A heatmap showing the SmANX (Smp_074140) gene expression during the schistosome life cycle. C ScRNAseq projections depicting the expression profiles of SmANX (Smp_074140) in different cell clusters in all cells, male, female and immature female worms of S. mansoni worms. D A network of SjANX A13 and its potential interacting proteins, and the line thickness indicates the strength of data support.
Analysis of the scRNAseq data allowed assessment of the expression profiles of Smp_074140 in different cell types of S. mansoni worms. As shown in Figure 4C, Smp_074140 was enriched in almost all cell types. Smp_074140 was mainly enriched in the tegument lineage, neurons, and neoblasts cell clusters of MM worms, the neurons, muscle, and parenchyma cell clusters of MF worms, and relatively limited in the neurons and parenchyma of SF worms (Figure 4C).
A protein–protein interaction network that was constructed based on the STRING database (Figure 4D) identified 10 proteins that potentially interact with SjANX A13, which included beta-galactosidase-1-like protein, dysferlin, IST1, nicotinate phosphoribosyltransferase, otoferlin, T-complex protein 1, UTP-glucose-1-phosphate uridylyltransferase, T-complex protein 1 subunit beta, Nucleolysin TIA-1, and UDP-glucose 4-epimerase. SjANX A13 and predicted interacting proteins were enriched in the GO terms “chaperonin-containing T-complex” and “cytoplasm”, in addition to the KEGG pathway “galactose metabolism”, suggesting potential roles in cellular component and physiological processes (Additional file 2).
Expression patterns of SjANX A13 at different time points after infection
To further clarify the mRNA expression profiles of SjANX A13, schistosomes were recovered from infected mice at different time points after infection. The qPCR results revealed that the mRNA levels of SjANX A13 tended to increase at 7–42 dpi (Figure 5A). After mating (around 15 dpi for S. japonicum), SjANX A13 expression gradually increased in MM worms and surpassed that of females at 25 dpi, whereas SjANX A13 expression slightly fluctuated in MF worms at 18–42 dpi (Figure 5B). Such differences in expression pattern may indicate that SjANX A13 plays different roles in males and females during reproductive development. Further comparisons of SF and MF worms showed that SjANX A13 was highly expressed in SF worms before eggshell formation (around 21 dpi for S. japonicum), whereas after eggshell formation, SjANX A13 expression was significantly higher in MF than SF worms (Figure 5C).

Transcriptional profile of SjANX A13. A Transcript levels of SjANX A13 in 7, 14, 18, 21, 25, 28, 35 and 42-day worms come from mixed cercariae-infected mice. B Transcript levels of SjANX A13 in 18, 21, 25, 28, 35 and 42-day mated male and female worms. C Transcript levels of SjANX A13 in 18, 21, 23 and 25-day SF and MF. All experiments were performed in triplicate and are expressed as the mean ± SD. Significant differences are indicated (*P < 0.05, **P < 0.01)
SjANX A13-knockdown affects the development and reproduction of S. japonicum
RNAi was performed to determine the function of SjANX A13 in vivo. Three specific SjANX A13 siRNA pairs were first tested in vivo to identify the most efficient. At 22 dpi, siRNAs were injected via the tail vein and the parasites were harvested 48 h later (Figure 6A). The effects of gene silencing were determined by qPCR and S1 siRNA was chosen for further experiments (Figure 6B).

SjANX A13 knockdown affects the vitality and reproduction of S. japonicum. A Time schedule for siRNA efficiency assay and long-term siRNA experiment. B Comparison of the efficacies of the negative control and three SjANX A13-specific siRNA pairs. C Effects of SjANX A13 S1 siRNA on SjANX A13 gene expression after 10 injections between 4 and 42 dpi. D Effect of long-term siRNA treatment on the worm burden. E Effect of long-term siRNA treatment on reproductive capacity of female worms. F Effect of long-term siRNA treatment on the liver egg burden. G Effect of long-term siRNA treatment on egg hatching rate. All experiments were performed in triplicate and are expressed as the mean ± SD. Significant differences are indicated (*P < 0.05, **P < 0.01). The reduction rates are presented in the panels. A was created with Biorender.com.
A long-term RNAi study revealed that SjANX A13 mRNA expression levels had significantly decreased after 10 injections with S1 siRNA, as detected by qPCR (Figures 6A and C). Morphological observations by SEM demonstrated that the MM and MF worms in the NC group had a continuous and well-organized network structure with clearly recognizable flower-like papillae. As compared to the NC group, most of the tegument of worms in the SjANX A13 S1 group showed no significant morphological changes, while the tegument surface of some areas appeared smoother (Additional file 3).
To further characterize the role of SjANX A13 in reproduction, the worm and liver egg burdens in mice were quantified. As compared to the PBS and NC groups, the number of worms and liver egg burden were reduced in the SjANX A13 S1 group by 14.5% and 29.5% (Figure 6D), while egg counts were decreased by 77.1% and 82.8%, respectively (Figure 6E). The loss of worms and reproductive capacity reduced the egg burden in the liver. As compared to the PBS and NC groups, the liver egg burden was decreased in the SjANX A13-knockdown group by 61.5% and 71.5%, respectively, while 44.2% and 24.2% of eggs harvested from the liver failed to hatch into miracidia (Figures 6F and G).
Vaccine efficacy of rSjANX A13 against S. japonicum
To determine whether rSjANX A13 exhibits immunoprotective effects, the SjANX A13 gene was cloned into the plasmid pET28a(+) and successfully expressed in E. coli BL21 (DE3) cells. Following initiation of gene transcription with IPTG, the bacterial culture was briefly sonicated and the lysate was separated into soluble and insoluble fractions. Most of the recombinant protein was contained in the supernatant and then purified by affinity chromatography using a His-tag purification column. The purified rSjANX A13 protein was verified by SDS–PAGE and stained with Coomassie brilliant blue, which showed a single band of the expected size (Additional file 4). Then, the immunoprotective effects of rSjANX A13 were assessed with an immune protection assay (Figure 7A). Serum levels of IgG-specific antibodies against rSjANX A13 of immunized and control mice were measured with an ELISA. The results showed that rSjANX A13 induced increased production of IgG-specific antibodies, with extremely elevated levels after the first immunization (Figure 7B). During the second and third subsequent immunizations, antibody levels were increased as compared to the first immunization and remained at high levels for 2 weeks after immunization (Figure 7B). Unfortunately, however, perhaps due to the imbalanced sex ratio of cercariae, which resulted in the collection of fewer female worms and complicated calculation of the liver egg burden, only the rate of worm reduction could be calculated, and the results showed that the worm burden was reduced (Figure 7C). However, this assay should be repeated because the infection process may have been flawed.

Vaccine efficacy of rSjANX A13 against S. japonicum. A Time schedule for immune protection assays. B Changes in IgG antibody levels. C Effect of rSjANX A13 immunization on the worm burden. All experiments were performed in triplicate and are expressed as the mean ± SD. Significant differences are indicated (ns > 0.05, **P < 0.01). A was created with Biorender.com.
SjANX A13 participates in host–parasite interactions via exosomes
Previous studies have shown that SjANX A13 is an important component of the excretory/secretory cargo of the exosomes of S. japonicum [32, 33]. To determine whether SjANX A13 is involved in host–parasite interactions through exosomes, a transwell model was constructed to simulate the activities of S. japonicum in host cells (Figure 8A). Following co-culture for 48 h, the presence of the SjANX A13 gene in LX-2 cells was verified by qPCR (Figure 8B), suggesting that S. japonicum worms can be delivered into host cells via exosomes. Thus, S. japonicum-derived exosomes were isolated from host cells as described in previous studies [33, 57, 59]. As shown in Figure 8C, TEM analysis confirmed enrichment of exosomes with the typical cup-like morphology. Next, the ability of worm-derived exosomes to deliver A13 directly into host cells was investigated. The worm-derived exosomes were purified and labeled with a green fluorescent dye (PKH67). Uptake of the PKH67-labeled exosomes by LX-2 cells was verified by fluorescence microscopy. The green fluorescence of worm-derived exosomes labeled with PKH67 was observed in LX-2 cells, demonstrating uptake of worm-derived exosomes by LX-2 cells, while LX-2 cells co-cultured with PKH67-stained PBS showed no green fluorescence (Figure 8D). Subsequently, exosome proteins at 10 µg/mL were selected for subsequent in vitro studies (Figure 8E). The results of qPCR analysis verified the presence of SjANX A13 in LX-2 cells (Figure 8F). Collectively, these results suggest that SjANX A13 might be involved in host–parasite interactions via exosomes.

SjANX A13 is involved in the host–parasite interaction via exosomes. A Schema of the transwell system. B Transcript levels of SjANX A13 in LX-2 cells of the transwell system. C Observation of S. japonicum worm-derived exosomes through TEM. The white arrows highlight typical structure of exosomes. Scale bar = 200 nm. D Representative images of the uptake of PKH67-labeled S. japonicum worm-derived exosomes (green) by LX-2 cells (DAPI blue). The black arrows highlight representative internalization. Scale bar = 50 μm. E Schema of exosomes and LX-2 cells co-incubation. F Transcript levels of SjANX A13 in LX-2 cells after coculture with S. japonicum worm-derived exosomes after 48 h. A and E were created with Biorender.com.
Discussion
Despite the similarities in amino acid sequences among vertebrates, invertebrates, fungi, plants, and protists, ANXs of eukaryotes have unique structural features and biological functions. The current nomenclature classifies ANXs as “A” for vertebrates, “B” for invertebrates (including schistosomes), “C” for fungi, “D” for plants, and “E” for protists [60]. With the recent and rapid updates of high-throughput databases for genomic and transcriptomics studies of parasitic helminths, there have been some inconsistencies in the nomenclature of ANXs in group B [61]. For instance, SjANX A13 is currently classified as group A (vertebrates) rather than group B. Therefore, the current nomenclature of SjANX A13 was maintained in the present study. However, a rational and consistent nomenclature is required to reflect the biological roles of ANXs.
Structurally, ANXs are characterized by similar C-terminal structural domains that consist of four homologous repeats of approximately 70 amino acids each, which have similar functions [61]. Recent studies have revealed that several ANXs, such as SmANX B22 of S. mansoni [29] and SbANX of S. bovis [26], are tegument components. In the present study, the structural domain composition of SjANX A13 was analyzed in reference to the SMART website, which showed that each homologous repeat was composed of approximately 60 amino acids. Meanwhile, the potential expression patterns of SjANX A13 were explored in reference to SmANX (Smp_074140), which has the highest homology (74.01%) with SjANX A13, and the scRNAseq database. According to the S. mansoni scRNAseq profile, Smp_074140 was only slightly expressed in the tegument cells of MM worms, which were not the predominant cell type. Although only the S. mansoni database was used as a reference, in conjunction with the results of the SjANX A13 RNAi assay in the present study, the lack of significant changes to the tegument after SjANX A13-knockdown in MM worms also suggests that SjANX A13 is not mainly expressed in tegument cells. In fact, mouse ANX A13 is an intestine-specific protein that is localized at the tips of the intestinal microvilli [62].
To further explore the function of SjANX A13, it was determined that the highest expression of Smp_074140 occurs in neurons, which are essential to the reproductive development of schistosomes [63]. In addition, the high expression of SjANX A13 in MF worms at 18, 21, 23, and 25 days also suggests potential roles in the growth and development of S. japonicum [17]. In the present study, long-term RNAi of SjANX A13 significantly reduced the fecundity of female worms and hatch capacity of eggs, indicating the involvement of SjANX A13 in the development of S. japonicum. Moreover, a protein-protein interaction network was constructed to identify proteins that potentially interact with SjANX A13. Among the identified proteins, expression of UDP-glucose 4-epimerase [64] and dysferlin [65] was mainly limited to the tegument cells of S. japonicum and a recombinant protein of UDP-glucose 4-epimerase and dysferlin showed potential as a vaccine target. Besides, T-complex protein 1 is involved cytoskeletal proteostasis [66] and nucleolysin TIA-1 is critical to the stress response and neurodevelopment in mammals [67]. Nonetheless, further studies are needed to determine whether SjANX A13 is related to the functions of these proteins.
Several ANXs located at the schistosome tegument interface with immunogenic properties have been proposed as potential vaccine candidates [24, 68]. In the present study, the immunoprotective effects of rSjANX A13 were evaluated. Due to the possible imbalance between male and female cercariae used in the assay, it was not possible to determine the extent of the liver egg burden, even though the worm burden was reduced. Therefore, the experiments must be repeated to further verify the results.
In addition to the potential biological activities against schistosomes, the effects of SjANX A13 as an excretory/secretory protein on the host cell were explored [32]. Notably, SbANX of S. bovis exhibits fibrinolytic and anticoagulant activities [26]. Recently, SjANX A13 was also identified in S. japonicum exosomes, which are considered important vectors for the regulation of host–parasite interactions [33, 69]. In the present study, through the construction of a transwell system and co-culture S. japonicum exosomes with LX-2, SjANX A13 was detected in host cells, suggesting participation in host–parasite interactions through exosomes.
In conclusion, based on the results of a previous proteomic study conducted by our group, bioinformatics, RNAi, and qPCR analyses were used to assess the potential involvement of SjANX A13 in the reproductive development of S. japonicum. The results confirmed the involvement of SjANX A13 in the host–parasite interactions via exosomes. However, further investigations are needed to assess the suitability of SjANX A13 as a vaccine candidate.
Availability of data and materials
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
References
GBD 2017 Diet Collaborators (2019) Health effects of dietary risks in 195 countries, 1990–2017: a systematic analysis for the global burden of disease study 2017. Lancet 393:1958–1972
Lo NC, Bezerra FSM, Colley DG, Fleming FM, Homeida M, Kabatereine N, Kabole FM, King CH, Mafe MA, Midzi N, Mutapi F, Mwanga JR, Ramzy RMR, Satrija F, Stothard JR, Traoré MS, Webster JP, Utzinger J, Zhou XN, Danso-Appiah A, Eusebi P, Loker ES, Obonyo CO, Quansah R, Liang S, Vaillant M, Murad MH, Hagan P, Garba A (2022) Review of 2022 WHO guidelines on the control and elimination of schistosomiasis. Lancet Infect Dis 22:e327–e335
Charlier J, van der Voort M, Kenyon F, Skuce P, Vercruysse J (2014) Chasing helminths and their economic impact on farmed ruminants. Trends Parasitol 30:361–367
Guo J, Li Y, Gray D, Ning A, Hu G, Chen H, Davis GM, Sleigh AC, Feng Z, McManus DP, Williams GM (2006) A drug-based intervention study on the importance of buffaloes for human Schistosoma japonicum infection around Poyang Lake, People’s Republic of China. Am J Trop Med Hyg 74:335–341
Zhou YB, Liang S, Jiang QW (2012) Factors impacting on progress towards elimination of transmission of Schistosomiasis japonica in China. Parasit Vectors 5:275
Grover E, Paull S, Kechris K, Buchwald A, James K, Liu Y, Carlton EJ (2022) Predictors of bovine Schistosoma japonicum infection in rural Sichuan, China. Int J Parasitol 52:485–496
Dang H, Li YL, Guo JY, Xu J, Li SZ, Lu S (2021) National surveillance of schistosomiasis morbidity in China, 2015–2019. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 33:120–126 (In Chinese)
Zhang LJ, Xu ZM, Yang F, Dang H, Li YL, Lü S, Cao CL, Xu J, Li SZ, Zhou XN (2021) Endemic status of schistosomiasis in People’s Republic of China in 2020. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 33:225–233 (In Chinese)
Zhang LJ, Xu ZM, Yang F, He JY, Dang H, Li YL, Cao CL, Xu J, Li SZ, Zhou XN (2022) Progress of schistosomiasis control in People’s Republic of China in 2021. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 34:329–336 (In Chinese)
Zhang L, He J, Yang F, Dang H, Li Y, Guo S, Li S, Cao C, Xu J, Li S, Zhou X (2023) Progress of schistosomiasis control in People’s Republic of China in 2022. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 35:217–224 (In Chinese)
Luo C, Wang Y, Su Q, Zhu J, Tang S, Bergquist R, Zhang Z, Hu Y (2023) Mapping schistosomiasis risk in Southeast Asia: a systematic review and geospatial analysis. Int J Epidemiol 52:1137–1149
Díaz AV, Walker M, Webster JP (2023) Reaching the World Health Organization elimination targets for schistosomiasis: the importance of a one health perspective. Philos Trans R Soc Lond Ser B Biol Sci 378:20220274
Vale N, Gouveia MJ, Rinaldi G, Brindley PJ, Gärtner F, Correia da Costa JM (2017) Praziquantel for schistosomiasis: single-drug metabolism revisited, mode of action, and resistance. Antimicrob Agents Chemother 61:e02582-16
Yu QF, Zhang JY, Sun MT, Gu MM, Zou HY, Webster JP, Lu DB (2021) In vivo praziquantel efficacy of Schistosoma japonicum over time: a systematic review and meta-analysis. Acta Trop 222:106048
Cort WW (1921) Sex in the trematode family Schistosomidae. Science 53:226–228
Popiel I, Cioli D, Erasmus DA (1984) The morphology and reproductive status of female Schistosoma mansoni following separation from male worms. Int J Parasitol 14:183–190
Zhong H, Ren Y, Qin F, Li X, Hou L, Gu S, Jin Y (2022) Schistosoma japonicum translationally controlled tumour protein, which is associated with the development of female worms, as a target for control of schistosomiasis. Int J Parasitol 52:569–579
Zhong H, Qin F, Ren Y, Li X, Hou L, Gu S, Jin Y (2023) Functional characterization of differentially expressed proteins coming from unisexual and bisexual infected Schistosoma japonicum female worms. Exp Parasitol 248:108504
Peattie DA, Alonso RA, Hein A, Caulfield JP (1989) Ultrastructural localization of giardins to the edges of disk microribbons of Giarida lamblia and the nucleotide and deduced protein sequence of alpha giardin. J Cell Biol 109:2323–2335
Hongli Y, Shuhan S, Ruiwen C, Yingjun G (2002) Cloning and functional identification of a novel annexin subfamily in Cysticercus cellulosae. Mol Biochem Parasitol 119:1–5
Weiland ME, McArthur AG, Morrison HG, Sogin ML, Svärd SG (2005) Annexin-like alpha giardins: a new cytoskeletal gene family in Giardia lamblia. Int J Parasitol 35:617–626
Castro-Borges W, Dowle A, Curwen RS, Thomas-Oates J, Wilson RA (2011) Enzymatic shaving of the tegument surface of live schistosomes for proteomic analysis: a rational approach to select vaccine candidates. PLoS Negl Trop Dis 5:e993
Braschi S, Borges WC, Wilson RA (2006) Proteomic analysis of the schistosome tegument and its surface membranes. Mem Inst Oswaldo Cruz 101:205–212
Leow CY, Willis C, Leow CH, Hofmann A, Jones M (2019) Molecular characterization of Schistosoma mansoni tegument annexins and comparative analysis of antibody responses following parasite infection. Mol Biochem Parasitol 234:111231
Mulvenna J, Moertel L, Jones MK, Nawaratna S, Lovas EM, Gobert GN, Colgrave M, Jones A, Loukas A, McManus DP (2010) Exposed proteins of the Schistosoma japonicum tegument. Int J Parasitol 40:543–554
de la Torre-Escudero E, Manzano-Román R, Siles-Lucas M, Pérez-Sánchez R, Moyano JC, Barrera I, Oleaga A (2012) Molecular and functional characterization of a Schistosoma bovis annexin: fibrinolytic and anticoagulant activity. Vet Parasitol 28:25–36
Gobert GN, McManus DP, Nawaratna S, Moertel L, Mulvenna J, Jones MK (2009) Tissue specific profiling of females of Schistosoma japonicum by integrated laser microdissection microscopy and microarray analysis. PLoS Negl Trop Dis 3:e469
Braschi S, Wilson RA (2006) Proteins exposed at the adult schistosome surface revealed by biotinylation. Mol Cell Proteom 5:347–356
Leow CY, Willis C, Osman A, Mason L, Simon A, Smith BJ, Gasser RB, Jones MK, Hofmann A (2014) Crystal structure and immunological properties of the first annexin from Schistosoma mansoni: insights into the structural integrity of the schistosomal tegument. FEBS J 281:1209–1225
Leow CY, Willis C, Hofmann A, Jones MK (2015) Structure-function analysis of apical membrane-associated molecules of the tegument of schistosome parasites of humans: prospects for identification of novel targets for parasite control. Br J Pharmacol 7:1653–1663
Fonseca CT, Braz Figueiredo Carvalho G, Carvalho Alves C, de Melo TT (2012) Schistosoma tegument proteins in vaccine and diagnosis development: an update. J Parasitol Res 2012:541268
Cao X, Fu Z, Zhang M, Han Y, Han Q, Lu K, Li H, Zhu C, Hong Y, Lin J (2016) Excretory/secretory proteome of 14-day schistosomula, Schistosoma japonicum. J Proteom 1:221–230
Zhu L, Liu J, Dao J, Lu K, Li H, Gu H, Liu J, Feng X, Cheng G (2016) Molecular characterization of S. japonicum exosome-like vesicles reveals their regulatory roles in parasite-host interactions. Sci Rep 13:25885
Smithers SR, Terry RJ (1965) The infection of laboratory hosts with cercariae of Schistosoma mansoni and the recovery of the adult worms. Parasitology 55:695–700
Smart Tool (2023) http://smart.embl-heidelberg.de/. Accessed 20 May 2023
Detaibio Tool (2023) http://www.detaibio.com/tools/. Accessed 20 May 2023
Clustal Omega Tool. http://www.clustal.org/. Accessed 20 May 2023
iTOL Tool. https://itol.embl.de/index.shtml. Accessed 20 May 2023
Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, Heer FT, de Beer TAP, Rempfer C, Bordoli L, Lepore R, Schwede T (2018) SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res 46:W296–W303
Wang Y, Wang Q, Huang H, Huang W, Chen Y, McGarvey PB, Wu CH, Arighi CN, UniProt Consortium (2021) A crowdsourcing open platform for literature curation in UniProt. PLoS Biol 19:e3001464
Chen VB, Arendall WB 3rd, Headd JJ, Keedy DA, Immormino RM, Kapral GJ, Murray LW, Richardson JS, Richardson DC (2010) MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr D Biol Crystallogr 66:12–21
Studer G, Tauriello G, Bienert S, Biasini M, Johner N, Schwede T (2021) ProMod3-A versatile homology modelling toolbox. PLoS Comput Biol 17:e1008667
Howe KL, Bolt BJ, Shafie M, Kersey P, Berriman M (2017) WormBase ParaSite—a comprehensive resource for helminth genomics. Mol Biochem Parasitol 215:2–10
LuZ Zhang Y, Berriman M (2018) A web portal for gene expression across all lifestages of Schistosoma mansoni. BioRxiv. https://doi.org/10.1101/308213
Wendt G, Zhao L, Chen R, Liu C, O’Donoghue AJ, Caffrey CR, Reese ML, Collins JJ 3rd (2020) A single-cell RNA-seq atlas of Schistosoma mansoni identifies a key regulator of blood feeding. Science 369:1644–1649
STRING Tool (2023) https://cn.string-db.org. Accessed 20 May 2023
Conesa A, Götz S, García-Gómez JM, Terol J, Talón M, Robles M (2005) Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21:3674–3676
Kanehisa M, Goto S (2000) KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res 28:27–30
Liu S, Piao X, Hou N, Cai P, Ma Y, Chen Q (2020) Duplex real-time PCR for sexing Schistosoma japonicum cercariae based on w chromosome-specific genes and its applications. PLoS Negl Trop Dis 14:e0008609
Rio DC, Ares M Jr, Hannon GJ, Nilsen TW (2010) Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb Protoc 6:pdb .prot5439
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25:402–408
Zhong H, Wu L, Ren Y, Qin F, Jin Y (2022) Comparative proteomic profiles of Schistosoma japonicum male worms derived from single-sex and bisexual infections. Int J Parasitol 52:815–828
Cheng G, Li X, Qin F, Xu R, Zhang Y, Liu J, Gu S, Jin Y (2019) Functional analysis of the Frzb2 gene in Schistosoma japonicum. Vet Res 50:108
Li X, Cheng G, Qin F, Liu J, Li H, Jin Y (2019) Function of the lesswright (lwr) gene in the growth, development, and reproduction of Schistosoma japonicum. Vet Parasitol 272:31–39
Yang Y, Jin Y, Liu P, Shi Y, Cao Y, Liu J, Shi Y, Li H, Lin J (2012) RNAi silencing of type V collagen in Schistosoma japonicum affects parasite morphology, spawning, and hatching. Parasitol Res 111:1251–1257
Shang Z, Guo Q, Zhou X, Yue Y, Zhou K, Tang L, Zhang Z, Fu Z, Liu J, Lin J, Xu B, Zhang M, Hong Y (2022) Characterization of aspartyl aminopeptidase from Schistosoma japonicum. Acta Trop 232:106519
Liu J, Zhu L, Wang L, Chen Y, Giri BR, Li J, Cheng G (2018) Isolation and characterization of extracellular vesicles from adult Schistosoma japonicum. J Vis Exp 22:57514
Armstrong JC (1965) Mating behavior and development of schistosomes in the mouse. J Parasitol 51:605–616
Liu J, Zhu L, Wang J, Qiu L, Chen Y, Davis RE, Cheng G (2019) Schistosoma japonicum extracellular vesicle miRNA cargo regulates host macrophage functions facilitating parasitism. PLoS Pathog 15:e1007817
Rios-Valencia DG, Mompala-García Y, Marquez-Navarro A, Tirado-Mendoza R, Ambrosio J (2022) Annexin in Taenia crassiceps ORF strain is localized in the osmoregulatory system. Acta Parasitol 67:827–834
Cantacessi C, Seddon JM, Miller TL, Leow CY, Thomas L, Mason L, Willis C, Walker G, Loukas A, Gasser RB, Jones MK, Hofmann A (2013) A genome-wide analysis of annexins from parasitic organisms and their vectors. Sci Rep 11:2893
McCulloch KM, Yamakawa I, Shifrin DA Jr, McConnell RE, Foegeding NJ, Singh PK, Mao S, Tyska MJ, Iverson TM (2019) An alternative N-terminal fold of the intestine-specific annexin A13a induces dimerization and regulates membrane-binding. J Biol Chem 294:3454–3463
Chen R, Wang J, Gradinaru I, Vu HS, Geboers S, Naidoo J, Ready JM, Williams NS, DeBerardinis RJ, Ross EM, Collins JJ 3rd (2022) A male-derived nonribosomal peptide pheromone controls female schistosome development. Cell 185:1506–1520.e17
Liu P, Shi Y, Yang Y, Cao Y, Shi Y, Li H, Liu J, Lin J, Jin Y (2012) Schistosoma japonicum UDP-glucose 4-epimerase protein is located on the tegument and induces moderate protection against challenge infection. PLoS One 7:e42050
Xiong Y, Ai D, Meng P, Wei M, Hong Y, Zhang M, Huang L, Fu Z, Shi Y, Lin J (2013) Cloning, expression, and preliminary characterization of the dysferlin tegument protein in Schistosoma japonicum. Parasitol Int 62:522–529
Han W, Jin M, Liu C, Zhao Q, Wang S, Wang Y, Yin Y, Peng C, Wang Y, Cong Y (2023) Structural basis of plp2-mediated cytoskeletal protein folding by TRiC/CCT. Sci Adv 9:eade1207
Byres LP, Mufteev M, Yuki KE, Wei W, Piekna A, Wilson MD, Rodrigues DC, Ellis J (2021) Identification of TIA1 mRNA targets during human neuronal development. Mol Biol Rep 48:6349–6361
Leow CY, Willis C, Chuah C, Leow CH, Jones M (2020) Immunogenicity, antibody responses and vaccine efficacy of recombinant annexin B30 against Schistosoma mansoni. Parasite Immunol 42:e12693
Du P, Giri BR, Liu J, Xia T, Grevelding CG, Cheng G (2020) Proteomic and deep sequencing analysis of extracellular vesicles isolated from adult male and female Schistosoma japonicum. PLoS Negl Trop Dis 14:e0008618
Acknowledgements
The authors thank International Science Editing for editing this manuscript and their valuable comments.
Funding
This work was supported by grants from the Natural Science Foundation of Shanghai, China (no. 20ZR1469300) and the Chinese National Natural Science Foundation (No. 31672245).
Author information
Authors and Affiliations
Contributions
Conceptualization: YJ. Data curation: HZ, KL, HL, RY. Formal analysis: ZF, BD, DZ, HL. Investigation: HZ, HL, ZF, JL, GS. Funding acquisition: YJ. Writing—original draft: HZ, LH. Writing—review and editing: YJ. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All animal experiments were performed in accordance with the guidelines of the Committee for the Care and Use of Laboratory Animals of the Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Shanghai, China, permit no. SYXK-20160010). The study protocol was approved by the Ethics and Animal Welfare Committee of the Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Shanghai, China, experiment no. SHVRImo-0124).
Competing interests
The authors declare that they have no competing interests.
Additional information
Handling editor: Frank Katzer.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Signal peptide and transmembrane region analysis of SjANX A13. A Analysis of signal peptide structure of SjANX A13. B Transmembrane region analysis of SjANX A13.
Additional file 2.
Functional analysis of SjANX A13 and its potential interacting molecules.
Additional file 3.
Morphological observation through SEM after long-term SjANX A13 knockdown. MM and MF worms from NC group had a continuous and well-organized network structure with clearly recognizable flower-like papillae, compared with the NC group, most of the tegument from worms of SjANX A13 S1 group showed no significant morphological changes, while the tegument in some areas (head part of male and middle part of female) appeared a smoother surface. ns: network structure, flp: fower-like papillae, b: bubbles, p: protrusion, pmp: pedicle mastoid process. Scale bar = 10 μm.
Additional file 4.
Amplification of SjANX A13 gene digested with vector pET-28a(+) and SDS-PAGE analysis of the expression of recombinant SjANX A13 protein. A Amplification of SjANX A13. Lane M: DL 2000 DNA marker; Lanes 1 and 2, PCR product of target gene SjANX A13. B Lane M: protein marker; Lanes 1, 2, 3, 4 and 5 are 0 h, 2 h, 4 h, 6 h, 8 h of rSjANX A13 induction, respectively. C SDS-PAGE detection of recombinant SjANX A13 protein (Coomassie). Lane M: protein marker; Lane 1: r SjANX A13 induced supernatant; Lane 2: r SjANX A13 induced precipitation; Lane 3: r SjANX A13 recombinant protein purified product.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhong, H., Hou, L., Qin, F. et al. Molecular and functional characterization of Schistosoma japonicum annexin A13. Vet Res 54, 116 (2023). https://doi.org/10.1186/s13567-023-01244-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-023-01244-z