
Abstract
Prion diseases are progressive and fatal, neurodegenerative disorders described in humans and animals. According to the “protein-only” hypothesis, the normal host-encoded prion protein (PrPC) is converted into a pathological and infectious form (PrPSc) in these diseases. Transgenic knockout models have shown that PrPC is a prerequisite for the development of prion disease. In Norwegian dairy goats, a mutation (Ter) in the prion protein gene (PRNP) effectively blocks PrPC synthesis. We inoculated 12 goats (4 PRNP+/+, 4 PRNP+/Ter, and 4 PRNPTer/Ter) intracerebrally with goat scrapie prions. The mean incubation time until clinical signs of prion disease was 601 days post-inoculation (dpi) in PRNP+/+ goats and 773 dpi in PRNP+/Ter goats. PrPSc and vacuolation were similarly distributed in the central nervous system (CNS) of both groups and observed in all brain regions and segments of the spinal cord. Generally, accumulation of PrPSc was limited in peripheral organs, but all PRNP+/+ goats and 1 of 4 PRNP+/Ter goats were positive in head lymph nodes. The four PRNPTer/Ter goats remained healthy, without clinical signs of prion disease, and were euthanized 1260 dpi. As expected, no accumulation of PrPSc was observed in the CNS or peripheral tissues of this group, as assessed by immunohistochemistry, enzyme immunoassay, and real-time quaking-induced conversion. Our study shows for the first time that animals devoid of PrPC due to a natural mutation do not propagate prions and are resistant to scrapie. Clinical onset of disease is delayed in heterozygous goats expressing about 50% of PrPC levels.
Similar content being viewed by others

Introduction
Prion diseases are a group of fatal, neurodegenerative disorders that occur in humans and a range of animals. Among these are Creutzfeldt-Jakob disease in humans, bovine spongiform encephalopathy in cattle, chronic wasting disease in cervids, and scrapie in sheep and goats. According to the “protein-only” hypothesis, the host-encoded prion protein (PrPC) is converted into a pathological form (PrPSc), which accumulates in the central nervous system (CNS) and variably in the peripheral tissues in these diseases [1, 2]. Studies of transgenic mice with genetic ablation of the prion protein gene (Prnp) have confirmed that host expression of PrPC is obligatory for prion-disease development, infectivity, and neurodegeneration [3, 4]. Interestingly, the neurotoxicity of PrPSc also depends completely on host expression of PrPC [5,6,7].
Susceptibility of small ruminants to scrapie is influenced by allelic variants (polymorphisms) in the PRNP. In goats, polymorphisms including S127, M142, S146, H154, Q211, and K222 have been associated with a decrease in disease susceptibility during natural outbreaks of scrapie [8,9,10,11]. Of particular interest are the S146 and K222 polymorphisms that delay clinical disease beyond the productive lifetime of goats upon experimental inoculation with scrapie [12,13,14,15]. These polymorphisms are found in a variety of goat breeds although the allele frequencies are relatively low [16,17,18]. In 2009 and 2017, the European Food Safety Authority (EFSA) released scientific opinions on genetic resistance in goats as requested by the European Commission [19, 20]. EFSA recommended further studies of candidate PRNP genotypes and allele frequencies before starting any breeding program for genetic resistance towards scrapie in goats.
In 2012, Norwegian researchers discovered a nonsense mutation at codon 32 in the PRNP of Norwegian dairy goats that completely terminates PrPC synthesis [21]. An initial genetic survey of several unrelated Norwegian dairy-goat flocks (n = 192) revealed an allele frequency of the PRNPTer-mutation of about 11% [21]. In a later genotyping of Norwegian goat bucks (n = 1984), 216 (10.9%) animals carried the mutation, of which only 13 (0.7%) were homozygous (PRNPTer/Ter) [22]. Heterozygous goats (PRNP+/Ter) express approximately half the amount of PrPC on the surface of peripheral blood mononuclear cells (PBMCs) [23]. This indicates that no compensatory expression from the normal allele is present.
In this proof of principal study, we investigated the predicted resistance conveyed by the PRNPTer allele in goats, which are the first animals described with a natural lack of PrPC. Groups of PRNPTer/Ter, PRNP+/Ter, and PRNP+/+ goats were subjected to intracerebral inoculation with goat scrapie prions. Disease progression was studied by clinical and neurological examinations, as well as post-mortem histopathological, immunohistochemical, and gene expression analyses. Furthermore, a thorough longitudinal clinical evaluation provides information on the range of clinical signs in goat scrapie.
Materials and methods
Animals
Twelve female Norwegian dairy goats (4 PRNP+/+, 4 PRNP+/Ter, and 4 PRNPTer/Ter) were recruited from a research herd at the Norwegian University of Life Sciences. The average kinship coefficient of included animals was 10.8% (Additional file 1). The mean age was 3.3 (SD = 0.5) months and the mean body weight was 18.4 (SD = 2.8) kg. The animals were kept under a 16 h light/8 h dark cycle and housed in three groups of 4 goats according to genotype. Hay and water were provided ad libitum, and they were provided with a commercial goat pellet concentrate twice a day. The goats were acclimatized in the new facilities for 14 days, and clinical examinations and hematology were performed to ensure healthy animals before inoculation.
Inoculation procedure
The inoculum was derived from the brain of a 3.5-year-old goat with the wild-type PRNP genotype with natural clinical scrapie as described in [24]. The goats were inoculated intracerebrally with 400 µL of the goat scrapie brain inoculum (10% wt/vol). In brief, goats were anesthetized with Zoletil vet. (tiletamine and zolazepam, 5.5 mg/kg intramuscularly) and local anesthetics were administered subcutaneously. A midline incision was made in the skin at the junction of the parietal and frontal bones, and a 1 mm hole was drilled through the calvarium. The inoculum was injected into the midbrain via a 21G and 5 cm long needle while the needle was being withdrawn from the brain. The skin incision was closed with two sutures. The goats received a single dosage of flunixin meglumine (2.2 mg/kg) and antibiotics (procaine benzylpenicillin, 45 mg/kg) for 3 days.
Sampling
Clinical and neurological examinations were performed approximately once a month from day 432 post-inoculation (dpi). The examination included assessment of behavior, cranial nerve function, proprioception, signs of pruritus (alopecia and scratch test), body condition score, and evaluation of locomotion in a corridor. The full neurological protocol was modified from [25] and can be found in Additional file 2. The groups of normal goats (PRNP+/+) and heterozygous goats (PRNP+/Ter) were euthanized when at least one animal in the group had advanced clinical signs of prion disease. The PRNPTer/Ter goats were euthanized approximately 1 year after the heterozygous group. Euthanasia was performed by an overdose of pentobarbital, and a full necropsy was performed. The left half of the brain, the whole spinal cord and cauda equina with corresponding dorsal root ganglia, and a range of peripheral organs were sampled. Tissue samples for histology and immunohistochemistry (IHC) were immersion-fixed in 4% formaldehyde for approximately 1 week, dehydrated in graded ethanol, and paraffin embedded. Tissues for enzyme immunoassay (EIA) and real-time quaking-induced conversion (RT-QuIC) were frozen at −70 °C until further processing. Samples for RNA extraction were collected from hippocampus within 20 min after euthanasia, and immediately immersed in RNA-later (Invitrogen, Oslo, Norway) and stored at −70 °C.
Histopathology
The brain was sectioned at six levels: (1) the olfactory lobe; (2) section through the frontal cortex at the level of ansate sulcus; (3) transverse section through the piriform lobe and thalamus; (4) transverse section through the rostral aspect of the superior colliculus; (5) transverse section through the brainstem and cerebellum at the level of the caudal cerebellar peduncles; and (6) transverse section through the obex. In addition, one section of cervical, thoracic, and lumbar spinal cord and cauda equina with corresponding dorsal root ganglia were selected for analysis. Peripheral tissues included in this study were the trigeminal ganglion, parotideal-, medial retropharyngeal- and superficial cervical lymph nodes, spleen, adrenal gland, and mucosa-associated lymphoid tissue at the recto-anal border. From each of these tissues and CNS areas, 4 µm sections were stained with hematoxylin and eosin and evaluated. Serial sections for IHC were mounted on Superfrost® Plus slides (Menzel-Gläser, Thermo Scientific, Oslo, Norway).
To generate a brain-lesion profile, vacuolation was graded from 0 to 5 as follows: 0, no vacuoles; 1, few vacuoles, unevenly distributed; 2, few vacuoles, evenly distributed; 3, moderate numbers of vacuoles, 10–20 per 40x-field; 4, many vacuoles, with tendency of coalescence; and 5, dense vacuolation with coalescence. The brain areas and scores were slightly modified from Fraser and Dickinson [26], since the original lesion profile is described in mice.
Immunohistochemistry
Sections were dried overnight at 58 °C. All tissues listed above were PrP-immunolabelled with the monoclonal antibody F89 (Abcam, Cambridge, UK), dilution 1:2000. Glial fibrillary acidic protein (GFAP) labeling, dilution 1:1500 (Dako, Glostrup, Denmark, Z0334), was performed on sections including thalamus and hippocampus. The sections were deparaffinized in xylene and rehydrated through decreasing concentrations of graded ethanol. The protocol for PrP-immunolabeling included demasking steps in 98% formic acid for 5 min, followed by hydrated autoclaving in citrate buffer (pH 6.0) at 121 °C for 15 min. Endogenous peroxidase activity was blocked by incubation in 3% H2O2 in methanol for 20 min at room temperature. Sections were then blocked in normal goat serum (1:50) diluted in phosphate-buffered saline (PBS) for 20 min and incubated with the primary antibody for 1 h at room temperature. Further steps were performed with the EnVison+ System-HRP AEC (Dako, K4005, mouse or K4009, rabbit). Sections were counterstained in hematoxylin and mounted using Faramount medium (Agilent, Oslo, Norway). Washing between steps was performed with Tris-buffered saline (TBS). All runs included a negative control section where the primary antibody was replaced with 1% BSA, and a brain or lymph node section from a known scrapie-negative animal.
Sections were examined by light microscopy, and both PrPSc and GFAP signals were semi-quantitatively scored as follows: 0 = negative; 1 = sparse; 2 = moderate; 3 = marked, including half-step grading. Astrocyte GFAP evaluation also included scoring of the number and localization of cells, and the appearance of primary and secondary processes.
IDEXX EIA test
Tissue homogenates (hippocampus, superficial cervical lymph node, and spleen) were prepared from 200 mg tissue and 1000 µL dH2O and analyzed with the HerdChek Scrapie/BSE Antigen EIA test (IDEXX, Hoofddorp, Netherlands) according to the manufacturer’s instructions. In brief, 120 µL brain homogenate (20% wt/vol) was mixed with 30 µL of the working plate diluent. For lymph node and spleen tissue, 100 µL homogenate (20% wt/vol) was mixed with 50 µL of the working plate diluent. One hundred microliter of the diluted sample was transferred onto the assay plate, incubated for 45 min with low shaking and washed six times. A conditioning buffer (100 µL) was added to each well and the plate was incubated for 10 min and then washed three times. The plate was incubated with a HRPO-conjugated anti-PrP antibody for 45 min, washed five times, before adding 100 µL tetramethylbenzidine substrate per well. After 15 min incubation, the reaction was stopped by adding 100 µL hydrochloric acid (1 M) and the absorbance was read at 450 nm and 620 nm. The cut-off value for a scrapie-positive result was calculated based on the following formula: the negative control mean (NCx̄) + 0.180.
RT-QuIC analysis
Truncated Syrian hamster (90–231) recombinant PrP (SHrPrP) solution (purchased from Colorado State University, USA) was used as substrate for all samples. The substrate was thawed at room temperature and filtered through a 100 kDa Nanosep centrifugal device (Pall Corporation, New York, USA). The RT-QuIC reaction was performed by adding 2 µL of 10 −2/10 −3 diluted seeding samples (hippocampus, 20% wt/vol) to 98 µL reaction mix composed of 20 mM NaH2PO4 (pH 7.4), 320 mM NaCl, 1.0 mM EDTA, 10 µM Thioflavin T (ThT), 0.001% sodium dodecyl sulfate, 0.1% N2, and 0.04 mg/mL SHrPrP substrate in wells of a black 96-well plate with clear bottom. The plate was sealed with plate-sealer film (Nalgene Nunc International, Roskilde, Denmark) and incubated at 42 °C in a BMG FLUOstar Omega plate reader with cycles of 60 s shaking (700 rpm, double orbital) followed by 60 s of rest. In one sample with low prion levels (#536), longer shaking intervals (90 s shaking and 30 s rest) were used to promote faster RT-QuIC kinetics [27]. ThT measurements (450 ± 10 nm excitation and 480 ± 10 nm emission; bottom read; gain 1.700) in each well were read every 15 min, for a total of at least 60 h. Fluorescence values were plotted as the average of triplicate reactions versus incubation time (GraphPad software Inc, San Diego, USA). RT-QuIC reaction was classified as positive if the fluorescence of at least two out of three replicates exceeded a threshold determined to be 5 SD above the average baseline fluorescence.
RNA extraction and cDNA synthesis
RNA was extracted from approximately 30 mg of tissue using the RNeasy Lipid Tissue Mini Kit (Qiagen, Hilden, Germany, 74804) according to the manufacturer’s instruction. The isolated RNA was quantified at optical density (OD)260 and purity was assessed by OD260/280 and OD260/230 absorbance readings with a DeNovix DS-11 spectrophotometer. RNA integrity (RIN) was assessed using the 2100 BioAnalyzer with RNA 6000 Nano kits (Agilent). Mean RIN value of included samples ± SD was 7.2 ± 0.30. Complementary DNA synthesis was performed using the QuantiTect Reverse Transcription Kit (Qiagen) according to the manufacturer’s instructions, with 600 ng RNA from each sample as templates. For each primer assay, a non-reverse transcriptase control was included to evaluate potential genomic DNA background signals, and a no template control was used to assess non-specific amplification or sample contamination.
Quantitative PCR
The expression of seven target genes (PRNP, GFAP, SAA3, CXCL10, CD14, S100A9, and IL1B) was investigated by the LightCycler 480 quantitative PCR (qPCR) system. Primers were designed to span exon/exon boundaries and to cover known splice variants by the Primer3 software [28]. ACTB and SAA3 sequences were adapted from [29, 30]. Primer sequences are given in Additional file 3. Quantitative PCR reactions were performed using SYBR Green PCR Master Mix, including 10 µL cDNA (1:10) in each reaction and standard cycling conditions: initial denaturation for 5 min at 95 °C, followed by 42 amplification cycles (10 s at 95 °C, 10 s at 60 °C and 15 s at 72 °C) and final construction of melting curves. A standard curve was generated for each target gene to obtain primer amplification efficiencies, correlations, and dynamic range. Normalization was performed against the ACTB reference gene, and relative expression was calculated using the 2 −ΔΔCq method, as described in [31].
Descriptive and statistical analysis
Data are presented as mean ± standard error of the mean (SEM). Graphical and statistical analyses were performed in GraphPad Prism 6 (GraphPad software Inc.). Associations between clinical signs and scrapie were analyzed by Barnard’s test for contingency tables. The Mann–Whitney U test was used to compare differences in ordinal data including the brain lesion profile and immunohistochemical scoring. Differences in gene expression were analyzed by one-way ANOVA with Tukey post hoc test for multiple comparisons. A p value < 0.05 was considered statistically significant.
Results
Clinical signs of prion disease are delayed in PRNP +/Ter goats
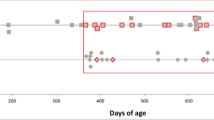
Figure 1 summarizes the clinical progression of nine selected clinical signs in the 12 goats (4 PRNP+/+, 4 PRNP+/Ter and 4 PRNPTer/Ter) included in the study. The mean incubation period until clinical signs of prion disease was 601 dpi (SD = 14) in the PRNP+/+ goats. This group was euthanized 615 dpi, when goat #417 had developed advanced clinical signs dominated by aggressive behavior over a short period of time. The mean incubation period in the heterozygous group was 773 dpi (SD = 46) and the goats were euthanized 909 dpi. Goat #527 gradually developed depression, social withdrawal, alopecia, and pruritus before euthanasia. One heterozygous animal (#469) had early signs of alopecia from day 432 dpi and intermittent periods of pruritus, not associated with scrapie. Histopathological evaluation of skin biopsies indicated an allergic dermatitis, but the etiology was not identified. The progression of clinical signs was much slower in in PRNP+/Ter goats than in PRNP+/+ goats (136 vs 14 days). The four PRNPTer/Ter goats remained healthy without any clinical signs of prion disease until they were euthanized 1260 dpi. However, one goat (#476) had mild signs of alopecia, and a dull and flaking coat throughout the last year. The complete open reading frame of PRNP was sequenced in all 12 goats to make sure that already-known polymorphisms associated with scrapie susceptibility would not bias the results. In addition to the stop-mutation in codon 32 of PRNPTer/Ter and PRNP+/Ter goats, polymorphisms were observed at codon 220 and 240 (Additional file 4), but none of these have been reported to affect incubation times. Moreover, the high average kinship of the goats (Additional file 1) indicates that the PRNP genotype is the main factor causing the observed differences between the groups.

Timeline of clinical and neurological signs. Goats of three different genotypes (4 PRNP+/+, 4 PRNP+/Ter and 4 PRNPTer/Ter) were intracerebrally inoculated with goat scrapie prions at day 0. The figure illustrates the progression of nine selected clinical signs at the indicated days after inoculation (dpi). Neurological exams were performed in all animals at each time point until euthanasia. *Goat 469 had early signs of alopecia associated with an allergic dermatitis. Goat 476 had mild signs of alopecia, and generally a dull and flaking coat not related to scrapie.
Clinical signs are linked to scrapie
Table 1 shows the distribution of clinical signs in goats that were confirmed scrapie-positive (8 goats) or scrapie-negative (4 goats). At the time of euthanasia, all scrapie-positive goats displayed mild to moderate signs of alopecia. Various areas were affected, but most frequently were the poll (7 of 8), pelvic region (7 of 8), and neck (5 of 8). A positive scratch test, teeth grinding, and overreactivity of head tests were also significantly overrepresented in scrapie-positive goats. Aggressive behavior, such as biting towards the tail and perineal area and horning, was observed in 4 of 8 goats, whereas depression was observed in 2 of 8 goats.
Histopathological changes
Vacuolation was scored in nine defined brain regions to generate a lesion profile (Figure 2A). Although the mean vacuolation score was higher in PRNP+/Ter goats than in PRNP+/+ goats in 8 of 9 brain regions, this difference was not statistically significant. As expected, the most severe changes were seen in the midbrain and thalamus (Figure 2C), near the inoculation site. The vacuoles were primarily associated with neuropil of grey matter and only a limited number of vacuoles were intraneuronal. Neuropil vacuolation was found throughout the spinal cord of both PRNP+/Ter and PRNP+/+ goats, and the dorsal horn was more affected in both groups. Scar tissue associated with the inoculation, also macroscopically visible, was found in two PRNP+/+ animals and one PRNPTer/Ter animal.

Histopathological changes and GFAP immunohistochemistry. A Vacuolation was graded 0–5 in nine defined brain areas (modified from Fraser and Dickinson 1968): 1, rostral part of medulla oblongata at the level of the obex; 2, transverse section of cerebellar cortex; 3, dorsal grey substance of the superior colliculus; 4, hypothalamus; 5, thalamus: massa intermedia and central nuclei; 6, hippocampus; 7, septal nuclei; 8, cerebral cortex dorsal to corpus callosum; and 9, cerebral cortex dorsal to septal nuclei. B GFAP signals were scored from 0 to 3 in thalamus and in three layers of the hippocampus (ML, molecular layer; SGZ, sub-granular zone; and hilus). C Vacuoles were primarily located in the neuropil of grey matter. Pictures show a representative vacuolation score 4 (PRNP+/+) and score 5 (PRNP+/Ter) in the thalamus. D Astrocyte GFAP labeling displayed increased length and thickening of primary and secondary processes in the thalamus of some scrapie-positive animals (goat #417 and #527). Values are mean ± SEM. n = 12 (4 PRNP+/+, 4 PRNP+/Ter, and 4 PRNPTer/Ter). Magnification: C, D ×200.
No significant differences in the number of GFAP-positive astrocytes, or in the length and thickness of primary and secondary processes, were observed between genotypes (Figure 2B). Nevertheless, the two animals with the most severe clinical signs and vacuolation (#417 and #527) had longer and thicker astrocytic primary and secondary processes in the thalamus when compared with PRNPTer/Ter goats (Figure 2D and Additional file 5).
Distribution of PrPSc in the CNS and peripheral tissues
The distribution of PrPSc in the CNS (Figure 3) correlated with vacuolation and was similar in the PRNP+/+ and PRNP+/Ter goat groups, reaching the olfactory lobe (Figure 4A) cranially and cauda equina caudally. The animals with the mildest clinical signs, such as goat #416, also had less severe pathology and accumulation of PrPSc (Additional file 6).

Schematic overview of PrPSc deposition in the CNS and ganglia. PrPSc was observed throughout the CNS and in dorsal root ganglia and trigeminal ganglia of scrapie-positive animals. Generally, the PRNP+/Ter animals had slightly higher levels of PrPSc than PRNP+/+ animals in the cranial part of the brain and in the spinal cord. The PRNP+/+ goats had more PrPSc accumulation in the ganglia than the heterozygous goats. No PrPSc was observed in PRNPTer/Ter goats. Abbreviations: C/P, caudate nucleus and putamen; T, thalamus; H, hippocampus; TG, trigeminal ganglion; DH, dorsal horn; VH, ventral horn; WM, white matter; DRG, dorsal root ganglion. Graded red color indicates the magnitude of PrPSc accumulation, whereas grey color are brain regions that were not examined. This scheme summarizes the data described in Additional file 4.

PrPSc immunohistochemistry of the CNS and peripheral tissues. A In the olfactory lobe, distinct granular PrPSc labeling of mitral cells was observed, but also neuropil and glial labeling involving the other layers was found. B In thalamus, there is moderate to marked PrPSc labeling that is mainly found in the neuropil. C There is marked PrPSc accumulation in the neuropil of the dorsal horn of both PRNP+/+ and PRNP+/Ter goats. D There is sparse to moderate labelling of both neurons and surrounding satellite cells in the trigeminal ganglion. E A follicle in the retropharyngeal lymph node of a PRNP+/+ goat is PrPSc positive, while this lymph node was negative in the heterozygous group. A–E There was no accumulation of PrPSc in the CNS or peripheral tissues in any of the PRNPTer/Ter goats. Magnification: A ×400; B, D, E ×200; C ×100.
Generally, neuropil deposits of PrPSc dominated, with less intraneuronal and intraglial labeling. The highest levels of PrPSc deposition were observed in brain regions near the area of inoculation (Figure 4B). In three of the heterozygous animals, fibrillar PrPSc-positive plaques were observed in several brain areas. PrPSc accumulation was observed throughout the spinal cord, more pronounced in the dorsal than in the ventral horns (Figure 4C). No PrPSc labeling of white matter occurred in the spinal cord. In the dorsal root ganglia and trigeminal ganglia there was sparse to moderate labeling of both neurons and satellite cells (Figure 4D). Nerve fibers in these ganglia were not labeled.
All four PRNP+/+ goats were positive in the retropharyngeal lymph node, but only in a few or moderate number of follicles (Figure 4E). There were also a few positive cells in sinuses associated with the positive follicles. In contrast, none of the PRNP+/Ter animals were positive in the retropharyngeal lymph node, but one heterozygous goat was positive in the parotideal lymph node (Additional file 6). In this animal, positive cells, most likely macrophages, were found in the capsular sinus, and associated with positive follicles. Unfortunately, this lymph node was not sampled in the PRNP+/+ goats. All animals were negative in the superficial cervical lymph nodes, rectoanal mucosa-associated lymphoid tissue, spleen, and adrenal glands.
There was no accumulation of PrPSc in the CNS or peripheral tissues in any of the PRNPTer/Ter goats (Figures 3 and 4A–E). Individual scoring-data of PrPSc in the CNS and peripheral tissues can be found in Additional file 6.
Validation of PrPSc-immunohistochemistry by EIA and RT-QuIC
There was a high similarity between the results of the three methods used for detection of prions: IHC, EIA and RT-QuIC (Table 2). Sections from hippocampus (#416 and #536) with a low IHC score (0.5), also had lower EIA absorbance values than sections with a higher IHC score (> 1.5). Peripheral tissues, including the spleen and superficial cervical lymph node, were confirmed negative by EIA in all animals. The RT-QuIC method was optimized by running samples (brain and lymphatic tissues) from sheep inoculated with classical scrapie. Homogenate from a known scrapie-negative sheep was included as negative control (Figure 5A). Different dilutions (10 −2 to 10 −5) of brain homogenate were analyzed, of which the 10 −2/10 −3 dilutions were considered optimal for detecting an increase in ThT fluorescence. Hippocampal prion seeding activity was detected in at least 2 of 3 replicates from all scrapie-positive goats (Figure 5B). All samples from PRNPTer/Ter goats were confirmed scrapie-negative by EIA and RT-QuIC.

Detection of prion seeding activity by RT-QuIC. RT-QuIC was performed using SHrPrP (90–231) as substrate. A Brain homogenate from two sheep inoculated with classical scrapie (positive control 1 and 2) and homogenate from a known scrapie-negative sheep (negative control) were used to assess the overall performance of the RT-QuIC. B Hippocampal homogenates diluted at 10 −2 or 10 −3 were analyzed as described in materials and methods. Inserted graph shows amplification of sample #536 at a slightly modified experimental protocol. Samples were analyzed in triplicates and the curves show the average fluorescence intensity at each time point.
Gene expression analysis
The expression of PRNP and six selected genes (GFAP, SAA3, CXCL10, CD14, S100A9, and IL1B) associated with inflammation and stress response was investigated in hippocampal tissue (Figure 6). The genes were selected based on differential expression in different scrapie models [32,33,34,35]. The levels of PRNP correlated with genotype and was 54% in PRNP+/Ter goats and about 5% in PRNPTer/Ter goats compared with PRNP+/+ goats. None of the other six target genes were differentially expressed between genotypes. However, goat #527, which was most clinically affected, had increased expression levels of GFAP and S100A9.

Relative gene expression in hippocampus at the time of euthanasia. Gene expression of seven target genes was measured by qPCR. For each gene, the expression levels are normalized relative to the PRNP+/+ group (= 1). Bars display mean expression (± SEM) with asterisks representing significant differences (p < 0.001) assessed by one-way ANOVA with Tukey post hoc test. n = 12 (4 PRNP+/+, 4 PRNP+/Ter, and 4 PRNPTer/Ter).
Discussion
In this study we show that goats naturally devoid of PrPC do not replicate prions and therefore are resistant to prion diseases. The PRNP+/+ goats were euthanized at 615 dpi, when clinical signs of scrapie were observed in all four animals in the group. Histopathological examination showed vacuolation and accumulation of PrPSc in the CNS of all four goats, which confirms that the goat scrapie inoculum was consistently infectious. In contrast to the PRNP+/+ goats, none of the PRNPTer/Ter goats had clinical signs of scrapie or evidence of PrPSc accumulation at 1260 dpi, which confirms that the PRNPTer-mutation confer resistance against scrapie in animals without PrPC. These results are in accordance with PrPSc inoculation of genetically modified Prnp-knock out animals, which are unable to replicate prions and do not develop prion disease [3, 4].
Heterozygous goats had, on average, 29% longer incubation time (773 versus 601 days) and a much slower progression in the development of clinical signs than PRNP+/+ goats. Interestingly, inoculation of heterozygous (Prnp0/+) mice with RML mouse-adapted scrapie resulted in a 70 to 120% increase in incubation time when compared with wild type mice [36,37,38]. This suggests that the protective effect of PRNPTer heterozygosity in goats, which is a natural host for scrapie, seems lower than expected based on the results from laboratory rodents. Studies of transgenic mice carrying different copy numbers of Prnp have shown that time to onset of clinical disease is inversely correlated with PrPC expression [38, 39]. This is probably because neurotoxic PrP-isoforms accumulate in a process that is rate limited by, and directly proportional to, PrPC concentrations [39, 40]. In addition, it has been demonstrated that neuronal expression of PrPC is essential to mediate neurotoxic effects of PrPSc oligomers [5,6,7]. In the present study, hippocampal PRNP levels in heterozygous goats were 54% of those of PRNP+/+ goats, which is in line with studies of PrPC levels in heterozygous animals [23, 39]. Thus, the delayed onset and slower progression of clinical disease in these animals probably reflect a combination of reduced availability of PrPC as a substrate for prion replication, in addition to less PrPC at the cell surface to mediate the toxic effects.
Although intracerebral inoculation does not mimic the natural oral route of prion infection, the current study provides insights into the spread of prions within the CNS and the centrifugal spread to peripheral tissues. Not surprisingly, the severity of histopathological changes and PrPSc accumulation was centered on the site of inoculation and spread cranially and caudally throughout the CNS. The distribution in the brain was similar to previous descriptions in goats and sheep upon intracerebral inoculation with scrapie [13, 41]. Interestingly, the severity of histopathological changes and PrPSc accumulation did not differ between the cervical region of the spinal cord and caudal segments, including cauda equina. As the inoculation site was near the ventricular system, one possible explanation could be that infectious prions were spread in the cerebrospinal fluid (CSF) during and after inoculation. Notably, high levels of prions have been observed in the CSF already at day 1 after intracerebral inoculation of hamsters [42]. In addition, neuronal spread likely contributed to the distribution of PrPSc within the CNS. We found only a limited centrifugal spread of prions to peripheral tissues, and positive tissues were either closely associated with the CNS, such as the dorsal root and trigeminal ganglia, or were head lymph nodes. If prions enter the CSF, it is likely that the meningeal lymphatic system [43, 44] could participate in transmission of PrPSc to head lymph nodes. The limited spread of PrPSc to peripheral organs, contrasts to some extent similar studies [13, 24]. The discrepancies could be due to differences in prion strains used for inoculation or associated with host factors, such as goat breed and genetic background. Whereas accumulation of PrPSc in the CNS did not differ between the genotypes, PrPSc was detected in head lymph nodes in all PRNP+/+ goats but only in 1 of 4 PRNP+/Ter animals. Since levels of PrPC is much higher in the CNS than in the periphery [45], it is possible that the amount of PrPC is a more critical factor for prion replication in peripheral tissues than in the CNS. Thus, it would be of great interest to investigate scrapie-resistance in PRNP+/Ter animals upon oral inoculation, since PRNP-expression in gut-associated lymphatic tissue is obligate for transmission of scrapie under natural conditions [46].
Goats without PrPC could be useful as breeding goats to avoid scrapie in endemic regions and might also be valuable for production of “prion-free” bio-products, such as vaccines and antibodies. Such use would require animals with good general health and production parameters, and adverse effects due to lack of PrPC should be ensured. During the last decades, substantial work has been directed towards understanding the normal function of PrPC. For instance, PrPC is abundantly expressed in the male reproductive system [47, 48], and loss of PrPC resulted in murine spermatozoa being more susceptible to Cu2+-induced stress [49]. To date, PRNPTer/Ter goats appear healthy under normal herd conditions [21] and detailed analyses of spermatozoa, both at rest and under acute stress, have not detected any abnormalities [50]. This suggests that PRNPTer/Ter goat bucks have normal sperm quality and fertility. In a study of acute systemic inflammation, however, goats without PrPC displayed a prolonged sickness behavior [51] and increased activation of genes encoding for pro-inflammatory cytokines in the lungs [52]. This indicates that PrPC could have a protective role against inflammatory stress [Reviewed in 22], and that PRNPTer/Ter goats might be more susceptible to infections. Larger epidemiological studies would be necessary to investigate the potential loss-of function phenotypes in PRNPTer/Ter goats in conjunction with important production-related diseases, such as mastitis and pneumonia. A possible drawback with breeding for the PRNPTer-mutation is the modest protection against scrapie observed in heterozygous animals, at least upon intracerebral inoculation. In addition, the mutation has only been observed in Norwegian dairy goats, which limits the availability of breeding animals. Nevertheless, two other PRNP polymorphisms, S146 and K222, have been identified to confer strong resistance towards scrapie in goats, even in heterozygous animals [13,14,15, 53,54,55]. Although the allelic variants S146 and K222 are commonly found in many goat breeds, there is a wide variation in allele frequency across countries and regions [16,17,18]. Recently, a breeding model for scrapie resistance was carried out in two European dairy breeds reported to have low K222 allele frequencies [56]. The authors concluded that breeding for scrapie resistance can be implemented in goats, even though the rate at which resistant animals increased was slow.
Although prion infection does not induce a proper immune response, cells of the innate immune system have proven to be critical players in the initial pathogenesis of prion disease [57]. Moreover, activated glial cells release pro-inflammatory cytokines and chemokines that probably contribute to the disease development [58]. In the current study, none of the target genes (GFAP, SAA3, CXCL10, CD14, S100A9, and IL1B) associated with the acute phase response and inflammation were differentially expressed in the hippocampus of animals with and without scrapie. In contrast, previous inoculation studies in sheep and rodents have reported several differentially expressed genes in the brain [32,33,34,35]. The discrepancies could reflect differences in timing of euthanasia and thus the magnitude of neuropathology and inflammation in the brain. For example, gliosis and increased GFAP were observed in the terminal stage of a mouse scrapie model, but not in the earlier stages [33]. In the present study, increased GFAP expression was only observed in goat #527, which also exhibited profound clinical signs. Taken together, these findings underline the difficulty of identifying expression-based biomarkers of early prion disease in small ruminants.
In conclusion, our findings confirm that goats naturally devoid of PrPC do not replicate prions and are therefore resistant to prion disease. The onset of disease and the progression of clinical signs is delayed in heterozygous animals. Pathological changes and the distribution of PrPSc in the CNS were similar in PRNP+/+ and PRNP+/Ter goats, and there was only a limited peripheral spread of PrPSc to head lymph nodes and nervous tissue in close proximity to the CNS.
References
Prusiner SB (1982) Novel proteinaceous infectious particles cause scrapie. Science 216:136–144
Kovacs GG, Budka H (2008) Prion diseases: from protein to cell pathology. Am J Pathol 172:555–565
Büeler H, Aguzzi A, Sailer A, Greiner RA, Autenried P, Aguet M, Weissmann C (1993) Mice devoid of PrP are resistant to scrapie. Cell 73:1339–1347
Prusiner SB, Groth D, Serban A, Koehler R, Foster D, Torchia M, Burton D, Yang SL, DeArmond SJ (1993) Ablation of the prion protein (PrP) gene in mice prevents scrapie and facilitates production of anti-PrP antibodies. Proc Natl Acad Sci U S A 90:10608–10612
Brandner S, Isenmann S, Raeber A, Fischer M, Sailer A, Kobayashi Y, Marino S, Weissmann C, Aguzzi A (1996) Normal host prion protein necessary for scrapie-induced neurotoxicity. Nature 379:339–343
Mallucci G, Dickinson A, Linehan J, Klöhn PC, Brandner S, Collinge J (2003) Depleting neuronal PrP in prion infection prevents disease and reverses spongiosis. Science 302:871–874
Resenberger UK, Harmeier A, Woerner AC, Goodman JL, Muller V, Krishnan R, Vabulas RM, Kretzschmar HA, Lindquist S, Hartl FU, Multhaup G, Winklhofer KF, Tatzelt J (2011) The cellular prion protein mediates neurotoxic signalling of beta-sheet-rich conformers independent of prion replication. EMBO J 30:2057–2070
Bouzalas IG, Dovas CI, Banos G, Papanastasopoulou M, Kritas S, Oevermann A, Papakostaki D, Evangelia C, Papadopoulos O, Seuberlich T, Koptopoulos G (2010) Caprine PRNP polymorphisms at codons 171, 211, 222 and 240 in a Greek herd and their association with classical scrapie. J Gen Virol 91:1629–1634
Billinis C, Panagiotidis CH, Psychas V, Argyroudis S, Nicolaou A, Leontides S, Papadopoulos O, Sklaviadis T (2002) Prion protein gene polymorphisms in natural goat scrapie. J Gen Virol 83:713–721
Barillet F, Mariat D, Amigues Y, Faugeras R, Caillat H, Moazami-Goudarzi K, Rupp R, Babilliot JM, Lacroux C, Lugan S, Schelcher F, Chartier C, Corbiére F, Andréoletti O, Perrin-Chauvineau C (2009) Identification of seven haplotypes of the caprine PrP gene at codons 127, 142, 154, 211, 222 and 240 in French Alpine and Saanen breeds and their association with classical scrapie. J Gen Virol 90:769–776
Goldmann W, Ryan K, Stewart P, Parnham D, Xicohtencatl R, Fernandez N, Saunders G, Windl O, González L, Bossers A, Foster J (2011) Caprine prion gene polymorphisms are associated with decreased incidence of classical scrapie in goat herds in the United Kingdom. Vet Res 42:110
Aguilar-Calvo P, Espinosa JC, Pintado B, Gutiérrez-Adan A, Alamillo E, Miranda A, Prieto I, Bossers A, Andréoletti O, Torres JM (2014) Role of the goat K222-PrPC-polymorphic variant in prion infection resistance. J Virol 88:2670–2676
Acutis PL, Martucci F, D’Angelo A, Peletto S, Colussi S, Maurella C, Porcario C, Iulini B, Mazza M, Dell’atti L, Zuccon F, Corona C, Martinelli N, Casalone C, Caramelli M, Lombardi G (2012) Resistance to classical scrapie in experimentally challenged goats carrying mutation K222 of the prion protein gene. Vet Res 43:8
Cinar MU, Schneider DA, Waldron DF, O’Rourke KI, White SN (2018) Goats singly heterozygous for PRNP S146 or K222 orally inoculated with classical scrapie at birth show no disease at ages well beyond 6 years. Vet J 233:19–24
Vaccari G, Di Bari MA, Morelli L, Nonno R, Chiappini B, Antonucci G, Marcon S, Esposito E, Fazzi P, Palazzini N, Troiano P, Petrella A, Di Guardo G, Agrimi U (2006) Identification of an allelic variant of the goat PrP gene associated with resistance to scrapie. J Gen Virol 87:1395–1402
Goldmann W, Marier E, Stewart P, Konold T, Street S, Langeveld J, Windl O, Ortiz-Pelaez A (2016) Prion protein genotype survey confirms low frequency of scrapie-resistant K222 allele in British goat herds. Vet Rec 178:168
White S, Herrmann-Hoesing L, O’Rourke K, Waldron D, Rowe J, Alverson J (2008) Prion gene (PRNP) haplotype variation in United States goat breeds. Genet Sel Evol 40:553–561
Kanata E, Humphreys-Panagiotidis C, Giadinis ND, Papaioannou N, Arsenakis M, Sklaviadis T (2014) Perspectives of a scrapie resistance breeding scheme targeting Q211, S146 and K222 caprine PRNP alleles in Greek goats. Vet Res 45:43
Ricci A, Allende A, Bolton D, Chemaly M, Davies R, Escamez PSF, Girones R, Herman L, Koutsoumanis K, Lindqvist R, Norrung B, Robertson L, Ru G, Sanaa M, Skandamis P, Speybroeck N, Simmons M, Ter Kuile B, Threlfall J, Wahlstrom H, Acutis PL, Andreoletti O, Goldmann W, Langeveld J, Windig JJ, Pelaez AO, Snary E, Hazards EPB (2017) Genetic resistance to transmissible spongiform encephalopathies (TSE) in goats. EFSA J 15:4962
Andreoletti O, Budka H, Buncic S, Colin P, Collins JD, De Koeijer A, Griffin J, Havelaar A, Hope J, Klein G, Kruse H, Magnino S, Martinez L, McLauchlin J, Nguyen-Thé C, Nöckler K, Nørrung B, Maradona MP, Roberts T, Vågsholm I, Vanopdenbosh E (2009) Genetic TSE resistance in goats. EFSA J 7:1371
Benestad SL, Austbø L, Tranulis MA, Espenes A, Olsaker I (2012) Healthy goats naturally devoid of prion protein. Vet Res 43:87
Salvesen Ø, Tatzelt J, Tranulis MA (2019) The prion protein in neuroimmune crosstalk. Neurochem Int 130:104335
Reiten MR, Bakkebø MK, Brun-Hansen H, Lewandowska-Sabat AM, Olsaker I, Tranulis MA, Espenes A, Boysen P (2015) Hematological shift in goat kids naturally devoid of prion protein. Front Cell Dev Biol 3:44
Lacroux C, Perrin-Chauvineau C, Corbière F, Aron N, Aguilar-Calvo P, Torres JM, Costes P, Brémaud I, Lugan S, Schelcher F, Barillet F, Andréoletti O (2014) Genetic resistance to scrapie infection in experimentally challenged goats. J Virol 88:2406–2413
Konold T, Bone GE, Phelan LJ, Simmons MM, González L, Sisó S, Goldmann W, Cawthraw S, Hawkins SA (2010) Monitoring of clinical signs in goats with transmissible spongiform encephalopathies. BMC Vet Res 6:13
Fraser H, Dickinson AG (1968) The sequential development of the brain lesion of scrapie in three strains of mice. J Comp Pathol 78:301–311
Orru CD, Hughson AG, Groveman BR, Campbell KJ, Anson KJ, Manca M, Kraus A, Caughey B (2016) Factors that improve RT-QuIC detection of prion seeding activity. Viruses 8:E40
Primer3 software. http://bioinfo.ut.ee/primer3-0.4.0/primer3/. Accessed 28 Feb 2018
Zhang Y, Zhang XD, Liu X, Li YS, Ding JP, Zhang XR, Zhang YH (2013) Reference gene screening for analyzing gene expression across goat tissue. Asian-Australas J Anim Sci 26:1665–1671
Brenaut P, Lefevre L, Rau A, Laloe D, Pisoni G, Moroni P, Bevilacqua C, Martin P (2014) Contribution of mammary epithelial cells to the immune response during early stages of a bacterial infection to Staphylococcus aureus. Vet Res 45:16
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25:402–408
Xiang W, Windl O, Wunsch G, Dugas M, Kohlmann A, Dierkes N, Westner IM, Kretzschmar HA (2004) Identification of differentially expressed genes in scrapie-infected mouse brains by using global gene expression technology. J Virol 78:11051–11060
Brown AR, Rebus S, McKimmie CS, Robertson K, Williams A, Fazakerley JK (2005) Gene expression profiling of the preclinical scrapie-infected hippocampus. Biochem Biophys Res Commun 334:86–95
Riemer C, Queck I, Simon D, Kurth R, Baier M (2000) Identification of upregulated genes in scrapie-infected brain tissue. J Virol 74:10245–10248
Meling S, Skovgaard K, Bardsen K, Helweg Heegaard PM, Ulvund MJ (2018) Expression of selected genes isolated from whole blood, liver and obex in lambs with experimental classical scrapie and healthy controls, showing a systemic innate immune response at the clinical end-stage. BMC Vet Res 14:281
Büeler H, Raeber A, Sailer A, Fischer M, Aguzzi A, Weissmann C (1994) High prion and PrPSc levels but delayed onset of disease in scrapie-inoculated mice heterozygous for a disrupted PrP gene. Mol Med 1:19–30
Mays CE, van der Merwe J, Kim C, Haldiman T, McKenzie D, Safar JG, Westaway D (2015) Prion infectivity plateaus and conversion to symptomatic disease originate from falling precursor levels and increased levels of oligomeric PrPSc species. J Virol 89:12418–12426
Manson JC, Clarke AR, McBride PA, McConnell I, Hope J (1994) PrP gene dosage determines the timing but not the final intensity or distribution of lesions in scrapie pathology. Neurodegeneration 3:331–340
Sandberg MK, Al-Doujaily H, Sharps B, Clarke AR, Collinge J (2011) Prion propagation and toxicity in vivo occur in two distinct mechanistic phases. Nature 470:540–542
Sandberg MK, Al-Doujaily H, Sharps B, De Oliveira MW, Schmidt C, Richard-Londt A, Lyall S, Linehan JM, Brandner S, Wadsworth JD, Clarke AR, Collinge J (2014) Prion neuropathology follows the accumulation of alternate prion protein isoforms after infective titre has peaked. Nat Commun 5:4347
Hamir AN, Kunkle RA, Bulgin MS, Rohwer RG, Gregori L, Richt JA (2008) Experimental transmission of scrapie agent to susceptible sheep by intralingual or intracerebral inoculation. Can J Vet Res 72:63–67
Orrù CD, Hughson AG, Race B, Raymond GJ, Caughey B (2012) Time course of prion seeding activity in cerebrospinal fluid of scrapie-infected hamsters after intratongue and intracerebral inoculations. J Clin Microbiol 50:1464–1466
Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS, Harris TH, Kipnis J (2015) Structural and functional features of central nervous system lymphatic vessels. Nature 523:337–341
Aspelund A, Antila S, Proulx ST, Karlsen TV, Karaman S, Detmar M, Wiig H, Alitalo K (2015) A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J Exp Med 212:991–999
Kobayashi S, Ano Y, Sakudo A, Yukawa M, Sugiura K, Manabe N, Nakayama H, Onodera T (2009) Quantification of PrPC in bovine peripheral tissues: analysis in wild-type and PrPC-deficient cattle. Mol Med Report 2:561–566
Donaldson DS, Kobayashi A, Ohno H, Yagita H, Williams IR, Mabbott NA (2012) M cell-depletion blocks oral prion disease pathogenesis. Mucosal Immunol 5:216–225
Fujisawa M, Kanai Y, Nam SY, Maeda S, Nakamuta N, Kano K, Kurohmaru M, Hayashi Y (2004) Expression of Prnp mRNA (prion protein gene) in mouse spermatogenic cells. J Reprod Dev 50:565–570
Ford MJ, Burton LJ, Morris RJ, Hall SM (2002) Selective expression of prion protein in peripheral tissues of the adult mouse. Neuroscience 113:177–192
Shaked Y, Rosenmann H, Talmor G, Gabizon R (1999) A C-terminal-truncated PrP isoform is present in mature sperm. J Biol Chem 274:32153–32158
Reiten MR, Malachin G, Kommisrud E, Østby GC, Waterhouse KE, Krogenæs AK, Kusnierczyk A, Bjørås M, Jalland CMO, Nekså LH, Røed SS, Stenseth EB, Myromslien FD, Zeremichael TT, Bakkebø MK, Espenes A, Tranulis MA (2018) Stress resilience of spermatozoa and blood mononuclear cells without prion protein. Front Mol Biosci 5:1
Salvesen Ø, Reiten MR, Espenes A, Bakkebø MK, Tranulis MA, Ersdal C (2017) LPS-induced systemic inflammation reveals an immunomodulatory role for the prion protein at the blood-brain interface. J Neuroinflammation 14:106
Salvesen Ø, Reiten MR, Kamstra JH, Bakkebø MK, Espenes A, Tranulis MA, Ersdal C (2017) Goats without prion protein display enhanced proinflammatory pulmonary signaling and extracellular matrix remodeling upon systemic lipopolysaccharide challenge. Front Immunol 8:1722
Ortiz-Pelaez A, Georgiadou S, Simmons MM, Windl O, Dawson M, Arnold ME, Neocleous P, Papasavva-Stylianou P (2015) Allelic variants at codon 146 in the PRNP gene show significant differences in the risk for natural scrapie in Cypriot goats. Epidemiol Infect 143:1304–1310
Georgiadou S, Ortiz-Pelaez A, Simmons MM, Windl O, Dawson M, Neocleous P, Papasavva-Stylianou P (2017) Goats with aspartic acid or serine at codon 146 of the PRNP gene remain scrapie-negative after lifetime exposure in affected herds in Cyprus. Epidemiol Infect 145:326–328
Papasavva-Stylianou P, Simmons MM, Ortiz-Pelaez A, Windl O, Spiropoulos J, Georgiadou S (2017) Effect of polymorphisms at codon 146 of the goat PRNP gene on susceptibility to challenge with classical scrapie by different routes. J Virol 91:01142–01217
Sacchi P, Rasero R, Ru G, Aiassa E, Colussi S, Ingravalle F, Peletto S, Perrotta MG, Sartore S, Soglia D, Acutis P (2018) Predicting the impact of selection for scrapie resistance on PRNP genotype frequencies in goats. Vet Res 49:26
Bradford BM, Mabbott NA (2012) Prion disease and the innate immune system. Viruses 4:3389–3419
Riemer C, Gültner S, Heise I, Holtkamp N, Baier M (2009) Neuroinflammation in prion diseases: concepts and targets for therapeutic intervention. CNS Neurol Disord Drug Targets 8:329–341
Acknowledgements
The authors acknowledge Lucy Robertson for proofreading the manuscript. We also thank Solfrid Nevland, Tom Honnemyr, Asbjørn Haga, Liv Helland, Garmann Auklend, Wenche Okstad, Siri B. Hamre, and Susan Skogtvedt Røed for their technical assistance. We thank Jette Jakobsen in The Norwegian Sheep and Goat Association for calculating the kinship coefficients of the goats.
Funding
The research was supported by The Research Council of Norway (grant ID 227386/E40).
Author information
Authors and Affiliations
Contributions
CE, MAT and AE designed the experiment. CE, ØS, MAT, AE, MRR, and GM performed the experiment. ØS carried out laboratory procedures, performed the statistical analysis, and drafted the manuscript. OA and SB provided and classified the inoculation material. CE did the pathology and PrPSc-immunohistochemical evaluation. TTV performed the RT-QuIC-analysis and LT performed the EIA. IO planned the matings to obtain animals with appropriate genotypes for the experiment and provided information on the goats. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The animal experiment was performed in compliance with ethical guidelines and approved by the Norwegian Animal Research Authority (ID 5826) with reference to the Norwegian regulation on animal experimentation (FOR-2015-06-18-761).
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Salvesen, Ø., Espenes, A., Reiten, M.R. et al. Goats naturally devoid of PrPC are resistant to scrapie. Vet Res 51, 1 (2020). https://doi.org/10.1186/s13567-019-0731-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-019-0731-2