
Abstract
In late 2014, a H5N8 highly pathogenic avian influenza (HPAI) virus, clade 2.3.4.4, spread by migratory waterfowl into North America reassorting with low pathogenicity AI viruses to produce a H5N2 HPAI virus. Since domestic waterfowl are common backyard poultry frequently in contact with wild waterfowl, the infectivity, transmissibility, and pathogenicity of the United States H5 HPAI index viruses (H5N8 and H5N2) was investigated in domestic ducks and geese. Ducks infected with the viruses had an increase in body temperature but no or mild clinical signs. Infected geese did not show increase in body temperature and most only had mild clinical signs; however, some geese presented severe neurological signs. Ducks became infected and transmitted the viruses to contacts when inoculated with high virus doses [(104 and 106 50% embryo infective dose (EID50)], but not with a lower dose (102 EID50). Geese inoculated with the H5N8 virus became infected regardless of the virus dose given, and transmitted the virus to direct contacts. Only geese inoculated with the higher doses of the H5N2 and their contacts became infected, indicating differences in infectivity between the two viruses and the two waterfowl species. Geese shed higher titers of virus and for a longer period of time than ducks. In conclusion, the H5 HPAI viruses can infect domestic waterfowl and easily transmit to contact birds, with geese being more susceptible to infection and disease than ducks. The disease is mostly asymptomatic, but infected birds shed virus for several days representing a risk to other poultry species.
Similar content being viewed by others


Introduction
The Asian-origin H5N1 A/goose/Guangdong/1/1996 (Gs/GD) lineage of highly pathogenic avian influenza (HPAI) viruses has spread across several continents affecting wild birds, poultry and humans. Despite great efforts to control H5N1 HPAI viruses, these viruses continue to circulate and evolve, which has led to the emergence of multiple genotypes or sublineages and the generation of reassortant H5 strains with novel gene constellations. Subclade 2.3.4.4 H5N1 viruses have mixed with several neuraminidase subtypes to generate widely circulating H5N2, H5N5, H5N6, and H5N8 subtypes of H5 HPAI viruses [1,2,3,4,5,6]. In early 2014, outbreaks of H5N8 HPAI virus were reported in South Korea and Japan in poultry and wild aquatic birds, with migratory aquatic birds strongly suspected in playing a key role in the spread of the virus [7, 8]. In late autumn 2014, H5N8 HPAI viruses were detected in Siberia, several countries in Europe, in South Korea, and in Japan [3, 4]. Concurrently, this virus was detected in the United States (U.S.) in captive falcons, wild birds, and backyard aquatic and gallinaceous poultry [5]. In addition, another novel reassortant H5 HPAI clade 2.3.4.4 virus (H5N2) was identified as the cause of an outbreak in poultry farms in British Columbia, Canada, during November 2014 [9], and was subsequently detected in the U.S. in wild waterfowl, raptors, and backyard poultry, including domestic ducks and geese [10]. From March to June 2015, this H5N2 virus predominated in the U.S., with extensive inter-farm transmission occurring in the Midwestern region. Over 7.5 million turkeys and 42.1 million chickens died or were depopulated in the USA during this outbreak which ended in June 2015 [11].
Wild and domestic waterfowl have played an important role in the maintenance and spread of Gs/GD lineage H5 HPAI viruses. Infected migratory waterfowl contributed to the spread of H5N1 and H5N8 HPAI viruses from Asia to other parts of the world [4, 8, 12, 13]. In 2016, H5N8 viruses of the same Gs/GD H5 lineage (HA clade 2.3.4.4) have been again detected in wild waterfowl in Russia, Europe, Middle East and Africa, these viruses causing outbreaks in poultry in many countries [14,15,16]. Domestic waterfowl have an important role in the maintenance and spread of H5N1 HPAI viruses, and have been shown to serve as intermediaries in the transmission of these viruses between wild waterfowl and other poultry species [17,18,19]. Wild waterfowl and domestic ducks are also important in the emergence and maintenance of H5N8 HPAI [3]. Hill et al. [20] found that wild waterfowl migration and domestic duck density were important factors in the epidemiology of H5N8 in the Republic of Korea. During the U.S. outbreak, H5N8 and H5N2 HPAI viruses were detected in backyard poultry including waterfowl [21], with possible contact with wild waterfowl or ponds reported in some cases [22]. H5N8 HPAI viruses have also affected commercial duck facilities in Europe and in the U.S. [16, 23, 24]. Recently, non-Gs/GD lineage H5 HPAI viruses (H5N1, H5N2, H5N9) have also caused outbreaks in domestic ducks in France in [25].
H5N1 Gs/GD lineage HPAI viruses are highly lethal to chickens; however, in domestic ducks these viruses can produce a range of clinical outcomes from asymptomatic infections to severe disease with mortality [26]. Both sick and asymptomatic infected ducks can shed high virus quantities into the environment favoring increased risk of transmission and potential outbreaks in commercial poultry. Naturally or experimentally, mortality in ducks caused by HPAI viruses had been infrequently reported before the Gs/GD H5N1 HPAI outbreaks in Asia [27, 28]. However, many Gs/GD lineage H5N1 HPAI viruses, and recently other Gs/GD-derived H5 viruses, have caused disease and death in domestic ducks (reviewed in [26]) [15, 16, 23, 24, 29]. Similarly, in domestic geese, outcome of infection with H5N1 HPAI viruses depends on the virus strain and the geese species. Domestic geese, naturally or experimentally infected with H5N1 viruses, showed from no clinical signs to neurological signs with or without mortality [30,31,32,33,34,35]. H5N8 and H5N2 HPAI viruses have also been detected in wild geese [16, 36]. In Taiwan, H5 clade 2.3.4.4 viruses (H5N2, H5N3 and H5N8) produced a severe epidemic in the domestic geese population in 2015, and more than 2.2 million geese died or were culled [37]. Recently, infections with H5N8 HPAI viruses have also been reported in domestic geese in Europe [16].
The recent emergence and recurrence of outbreaks of H5NX Gs/GD lineage HPAI in poultry underscore the need to better understand the pathobiology of these viruses in domestic waterfowl. In this study, in order to improve early detection of H5 HPAI viruses in domestic waterfowl, the infectivity, transmissibility and pathogenicity of the index H5N8 and H5N2 clade 2.3.4.4 HPAI viruses from the U.S. outbreak, was investigated in Pekin ducks and Chinese geese.
Materials and methods
Virus
The highly pathogenic avian influenza (HPAI) viruses A/gyrfalcon/Washington/40188-6/2014 H5N8 (GF/WA/14 H5N8) and A/Northern Pintail/Washington/40964/2014 H5N2 (NP/WA/14 H5N2) were used as challenge viruses. These were the first two Gs/GD H5 HPAI isolates, HA clade 2.3.4.4, from the U.S. outbreak and are considered representative of both the wholly Eurasian H5N8 lineage viruses and reassortant Eurasian/North American lineage H5N2 viruses, respectively [5]. The viruses were propagated and titrated in specific pathogen free (SPF) embryonating chicken eggs (ECE) using standard methods [38]. Stocks were diluted to the target dose with brain–heart infusion (BHI) broth (Becton, Dickinson and Company, Sparks, MD, USA). The experiments were performed in biosecurity level-3 enhanced (BSL-3E) facilities in accordance with procedures approved by the U.S. National Poultry Research Center (USNPRC) Institutional Biosecurity Committee.
Animals and housing
Pekin ducks (Anas platyrhynchos var. domestica) and White Chinese geese (Anser cygnoides) were obtained at 2 days of age from a commercial hatchery and reared in USNPRC facilities. At 2 weeks of age, birds were transferred to ABSL-3 enhanced facilities for virus challenge. Serum samples were collected from ten birds from each species prior to challenge to ensure that the birds were serologically negative for AI viruses by ELISA (FlockCheck Avian Influenza MultiS-Screen Antibody Test®, IDEXX Laboratories, Westbrook, ME, USA). Each experimental group was housed separately in self-contained isolation units ventilated under negative pressure with inlet and exhaust HEPA-filtered air within the animal BSL-3 enhanced facilities at Southeast Poultry Research Laboratory. This study and associated procedures were reviewed and approved by the USNPRC Institutional Animal Care and Use Committee (IACUC).
Experimental design and sampling
Similar experiments were conducted with each bird species to evaluate the mean bird infectious dose (BID50), transmissibility, and pathogenicity of the 2014 H5 HPAI index viruses. The 2-week-old ducks and geese were separated into virus-inoculated groups (4–5 birds for each species) as shown in Table 1. Non-inoculated control groups were included for each species. Groups containing 4–5 Pekin ducks were challenged with the appropriate dose per bird (102 50% egg infectious doses [EID50] per bird [low dose], 104 EID50 per bird [medium dose], or 106 EID50 per bird [high dose]), administered in 0.1 mL by the intrachoanal route. To examine pathogenesis, two additional birds were challenged with the high dose of the viruses. Sham-inoculated control ducks were inoculated with 0.1 mL of sterile allantoic fluid diluted 1:300 in BHI. To assess transmission by contact, three naïve ducks were introduced in the isolators with virus-inoculated ducks at one day post-inoculation (dpi). Groups containing four Chinese geese were challenged by intrachoanal inoculation with sham inoculum or with a dose of 102, 104, or 106 EID50/bird in 0.1 mL of either virus (Table 1). Two additional geese were challenged with the high dose of the viruses. Two naïve geese were introduced in the isolators with virus-inoculated geese at 1 dpi. The inoculum titers were subsequently verified by back titration in ECE as 1.7–1.9 (low dose), 3.5–3.9 (medium dose), and 5.7–6.1 (high dose) log10 EID50/0.1 mL.
Clinical signs were monitored daily for 10 days in the duck experiment and for 11 days in the geese experiment. Oropharyngeal (OP) and cloacal (CL) swabs were collected from all birds at days 2, 4, 7 and 10, 11 to determine virus shed. Body temperatures were taken at 2 and 4 dpi from birds inoculated with the high dose of the viruses and from the sham inoculated control birds. Significant difference for body temperatures between groups was analyzed using Kruskal–Wallis test (GraphPad Prism™ Version 5 software). A p value of < 0.05 was considered to be significant. For each species, the two additional birds challenged with the high dose of the viruses were euthanized and necropsied at 4 dpi to evaluate gross lesions. A full set of tissues were collected from each bird and fixed in 10% neutral buffered formalin solution, paraffin-embedded, sectioned, and stained with hematoxylin-and-eosin for histopathologic evaluation. Duplicate sections were stained by immunohistochemical (IHC) methods to determine influenza viral antigen distribution in individual tissues [39]. Portions of lung, heart, brain, muscle and spleen were also collected and stored at −80 °C for subsequent virus detection and quantification. Sera were collected from all surviving birds at the end of the experiments to evaluate infection status by antibody levels using hemagglutination inhibition (HI) assay. HI assays were performed using standard methods and homologous antigen [40]. The virus infectious dose was calculated by the Reed–Muench method [41], using the criteria that birds were considered infected if they shed detectable levels of virus at any time and/or were positive for antibody at the end of the study.
Viral RNA quantification in swabs and tissues
OP and CL swabs were collected in 1 mL of BHI broth with a final concentration of 10 μg/mL of gentamicin, 100 units/mL of penicillin G, and 56 μg/mL of amphotericin B, and kept frozen at −80 °C until processed. RNA was extracted using MagMAX™-96 AI/ND Viral RNA Isolation Kit® (Ambion, Inc.) following the manufacturer’s instructions. qRRT-PCR reactions targeting the influenza virus M gene [42] were conducted using AgPath-ID one-step RT-PCR Kit (Ambion, Austin, TX, USA) and the ABI 7500 Fast Real-Time PCR system (Applied Biosystem, Carlsbad, CA, USA). Viral RNA was extracted from tissues using Trizol LS reagent (Invitrogen, Carlsbad, CA, USA) and the Qiagen RNeasy Mini Kit (Qiagen Corp, Valencia, CA, USA). In tissue homogenates, and in order to standardize the amount of nonspecific RNA from the tissue, the resulting viral RNA extracts were quantified by NanoDrop™ 1000 Spectrophotometer (Thermo Fisher Scientific) following the manufacturer’s instructions and accordingly diluted with phosphate buffered saline to obtain 50 ng/µL. For virus quantification, a standard curve was established with RNA extracted from dilutions of the same titrated stock of the challenge virus. Results were reported as EID50/mL or EID50/g equivalents and the lower limit of detection was was 101.8 EID50/mL for both viruses. For statistical purposes, qRRT-PCR negative samples were given a numeric value of 1.7 log10 EID50/mL (1.7 log10 EID50/g).
Results
Infectivity, transmission and pathogenicity of the H5N8 and H5N2 HPAI viruses in domestic ducks
No ducks were infected in the groups inoculated with the lowest dose (102 EID50) of each virus (Table 1). Birds were considered infected if they had detectable virus or seroconverted. All ducks inoculated with the medium (104 EID50) or high (106 EID50) dose of the viruses were infected but no mortality occurred. The 50% bird infectious dose (BID50) for both viruses was 3 log10 EID50. One or 2 of 3 contact ducks in the groups inoculated with the medium dose for both viruses were infected, and 2 or 3 of 3 contact ducks in the high dose groups were infected. No mortality was observed in these ducks either.
A significant difference in body temperature was observed at 2 dpi, but not at 4 dpi, between sham-inoculated control ducks and H5N8-inoculated ducks (high dose group) (Figure 1). Four of six ducks inoculated with the H5N2 virus also showed high body temperatures. Conjunctivitis, tearing of the eyes, and diarrhea was observed intermittently in a low percentage of ducks in each group (1–2 ducks). One duck in the H5N2 high dose group presented mild ataxia starting at 2 dpi. At the end of the study (10 dpi) one duck from the H5N8 medium dose group and a second duck from the H5N2 high dose group also had mild ataxia. No other clinical signs were observed in the virus-inoculated and contact ducks. At 4 dpi, two ducks inoculated with the high dose of the viruses were euthanized for gross examination. One of the ducks inoculated with the H5N8 virus had airsacculitis and marbled spleen. No gross lesions were observed in the three other ducks.

Mean body temperatures of 2-week-old Pekin ducks and Chinese geese inoculated by the intrachoanal route with 10 6 EID 50 of H5N8 and H5N2 HPAI viruses, at 2 and 4 days post-inoculation. Bars represent the standard deviation of the mean. Significant difference in body temperature compared to controls (**p < 0.01).
Infectivity, transmission and pathogenicity of the H5N8 and H5N2 HPAI viruses in domestic geese
All geese inoculated with the H5N8 virus were infected regardless of the dose given (BID50 was less than 2 log10 EID50) (Table 1). No geese were infected in the group inoculated with the lowest dose of the H5N2 virus, but the virus did infect all geese at the medium and high doses (BID50 was 3 log10 EID50). No differences in body temperature were found among virus-inoculated geese (high dose groups) at 2 and 4 dpi when compared to sham-inoculated controls (Figure 1). In the groups of geese inoculated with the H5N8 virus, no or mild clinical signs (conjunctivitis) were observed during the study, but at 11 dpi one goose from the high dose group had mild tremors and incoordination, and one of the contacts from the same group was found dead. In the groups of geese inoculated with the H5N2 virus, one goose and a contact goose from the high and medium dose groups presented severe ataxia and torticollis, at 8 and 10 days respectively, and were euthanized and necropsied. At 11 dpi, one goose from each of the medium and high dose groups had mild ataxia. The rest of the geese inoculated with H5N2 showed no or mild clinical signs (conjunctivitis). No gross lesions were observed in the two geese that were euthanized and examined at 4 dpi from the H5N8 pathogenesis group. The two geese from the H5N2 pathogenesis group had multifocal areas of necrosis in the pancreas. The goose from the H5N2 high dose group euthanized at 8 dpi due to severe neurological signs had nasal discharge and empty intestines. The contact goose from the H5N2 medium dose group euthanized at 10 dpi had a pale spleen, enlarged heart, hemorrhagic thymus, malacic brain, and nasal discharge.
Microscopic lesions and viral antigen distribution
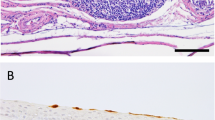
In order to evaluate microscopic lesions and sites of virus replication, tissues collected from ducks and geese necropsied at 4 dpi were examined, and immunohistochemical staining for AI virus nucleoprotein antigen was performed (Table 2; Figure 2). Tissues from the two geese that were euthanized at 8 and 10 dpi were also examined.

Histological lesions and immunohistochemical detection of viral antigen in 2-week old Pekin ducks and Chinese geese inoculated by the intrachoanal route with H5N8 and H5N2 HPAI viruses. 40×; viral antigen staining in red. A Heart. Goose inoculated with H5N2, 8 dpi; lymphoplasmacytic inflammation. B Skeletal muscle. Goose inoculated with H5N2, 10 dpi; lymphoplasmacytic inflammation. C Cerebrum. Goose inoculated with H5N2, 8 dpi; foci of malacia, and gliosis. D Cerebrum. Goose inoculated with H5N2, 10 dpi; viral antigen in neurons and glial cells. E Spleen. Goose inoculated with H5N2, 8 dpi; viral antigen in necrotic cells and mononuclear cells. F Airsac. Pekin duck inoculated with H5N8, 4 dpi; viral antigen present in epithelial cells. G Harderian gland. Pekin duck inoculated with H5N8, 4 dpi; viral antigen present in epithelial cells and infiltrating monocytes. H Lung. Pekin duck inoculated with H5N8, 4 dpi; viral antigen in epithelium of air capillaries and infiltrating monocytes.
In all ducks and geese examined at 4 dpi, mild catarrhal rhinitis and sinusitis was observed. The tracheas had mild degenerative changes of the overlying epithelium and mild lymphocytic infiltration in the submucosa. Also present: mild to moderate interstitial pneumonia, mild airsacculitis, focal necrosis of the epithelia of the Harderian glands, mild proliferation of gut-associated lymphoid tissues, and mild to moderate lymphoid depletion in cloacal bursa, thymus and spleen. Remaining organs lacked significant histopathologic changes. The two geese from the H5N2 pathogenesis group had, in addition, multifocal areas of necrosis in the pancreas, and the two geese examined at 8 and 10 dpi had individual cell necrosis of myofibers and focal mononuclear inflammation in the heart and thigh skeletal muscle (Figures 2A and B), and in the brain, randomly scattered foci of malacia, gliosis and perivascular cuffing (Figure 2C). Viral antigen staining was present in multiple tissues from ducks and geese infected with the H5 HPAI viruses indicating systemic infection (results in Table 2; Figures 2C–G). Viral antigen was present in epithelial cells and infiltrating phagocytes in the nasal turbinates, trachea, bronchus, lung air capillaries, air sac, Harderian glands, cloacal bursa, and in resident and infiltrating phagocytes of the spleen. Viral antigen staining was also present in pancreatic acinar epithelial cells, hepatocytes, neurons and glial cells of the brain, fragmented cardiac and skeletal myofibers of geese infected with the H5N2 virus.
Replication of H5N8 and H5N2 HPAI viruses in Pekin ducks and Chinese geese
Quantitation of viral shed was performed by qRRT-PCR using extrapolation of a standard curve generated with the challenge viruses via virus isolation and titration. OP and CL viral shed was examined at 2, 4, 7 and 10 dpi in ducks (Figures 3A, C, E and G) and at 2, 4, 7 and 11 dpi in geese (Figures 4A, C, E and G). To evaluate viral shed in contact birds, naïve birds were introduced in the isolator with inoculated birds at 1 dpi and sampled at 1, 3, 6 and 9 or 10 days post-exposure (dpe) (Figures 3 and 4B, D, F and H).

Mean oropharyngeal (OP) and cloacal (CL) viral shed from 2-week-old Pekin ducks directly inoculated (A, C, E, G) or contact-exposed (B, D, F, H) with H5N8 and H5N2 HPAI viruses. Ducks were inoculated with 102, 104 and 106 EID50 of either virus and titers determined by qRRT-PCR. Bars represent the standard deviation of the mean. Swabs from which virus was not detected were given a numeric value of 101.7 EID50/mL.

Mean oropharyngeal (OP) and cloacal (CL) viral shed from 2-week-old Chinese geese directly inoculated (A, C, E, G) or contact-exposed (B, D, F, H) with H5N8 and H5N2 HPAI viruses. Geese were inoculated with 102, 104 and 106 EID50 of either virus and titers determined by qRRT-PCR. Bars represent the standard deviation of the mean. Swabs from which virus was not detected were given a numeric value of 101.7 EID50/mL.
Ducks inoculated with the low dose of either the H5N8 or H5N2 virus did not shed virus and virus was not transmitted to contacts. Ducks inoculated or contact-exposed in the medium and high dose groups shed virus mostly through the OP route (Figures 3A, B, E and F); few ducks (including the contacts) shed through the CL route and, if so, at very low titers (Figures 3C, D, G and H). Ducks from the medium and high dose groups shed virus by the OP route up to 7 dpi (6 dpe for contacts). Ducks inoculated with the high doses of the viruses were able to transmit to 3–4 of the 6 contacts, but higher titers were shed by the H5N2 contacts (Figures 3F and H).
All geese inoculated with H5N8 virus shed virus through the OP and CL routes at high titers regardless of the inoculation dose (Figures 4A and C). H5N8 and H5N2 contact exposed geese became infected and showed a similar virus shedding pattern as the directly inoculated geese (Figures 4B and D). Only geese inoculated with the medium and high dose of the H5N2 virus shed virus (Figure 2E, G). Contact geese in these groups also shed virus similar to the inoculated birds (Figures 4F and H). H5N2 OP viral titers were similar to those observed with the H5N8 virus, but H5N2 virus was shed for longer by the CL route. Overall, the Pekin ducks shed lower virus titers and for a shorter period of time (7 days vs 11 days) than the Chinese geese. The peak of viral shedding for both viruses in both species was between 3 and 5 dpi.
To evaluate systemic replication of H5N8 and H5N2 HPAI viruses in ducks and geese, viral titers were determined in brain, heart, spleen, lung and muscle. Tissues were collected from 2 birds of each 106 EID50 inoculated groups at 4 dpi (Figure 5). Ducks and geese showed moderate to high H5N8 and H5N2 HPAI virus titers in all tissues examined, with the highest titers found in tissues of geese inoculated with the H5N2 virus. Tissues were also collected from the two geese exposed to H5N2 virus and showing neurological signs; both birds also presenting high virus titers in all tissues, especially in brain.

Virus titers in tissues from Pekin ducks and Chinese geese directly inoculated or contact-exposed with H5N8 and H5N2 HPAI viruses. Titers determined by qRRT-PCR. A Tissues from Pekin ducks, 4 dpi. B Tissues from geese, 4 dpi. C Tissues from geese showing neurological signs euthanized at 8 dpi and 10 dpi inoculated or exposed to H5N2 virus, respectively. Bars represent the standard error of the mean. Tissues from which virus was not detected were given a numeric value of 101.7 EID50/g.
Serology
Serum samples were examined for detectable titers of antibodies against the corresponding challenge virus at 10 or 11 dpi, for Pekin ducks or geese respectively. All Pekin ducks inoculated with the medium and high dose of either virus seroconverted (Table 1); 1 of 3 contacts placed in the medium dose groups and 2 or 3 of 3 in the highest dose groups seroconverted. Only one goose inoculated with the low dose, two geese inoculated with the high dose and one contact goose in the medium dose group seroconverted to the H5N8 virus. Two geese, one from the medium dose and one from the high dose group seroconverted to the H5N2 virus.
Discussion
In this study we describe the pathogenesis and transmission dynamics of the U.S. index H5N8 and H5N2 HPAI viruses (Gs/GD lineage, HA clade 2.3.4.4) in domestic ducks and geese with the objective of better understanding the infection process in order to improve strategies for early detection of HPAI viruses in domestic waterfowl. Based on viral shed and serology results, only ducks inoculated with the medium and high doses of the viruses (104 and 106 EID50, respectively) were infected and were able to efficiently transmit to contacts, resulting in a BID50 of 3 log10 EID50 for both viruses. Similarly, geese inoculated with the medium and high doses of the H5N2 HPAI virus and the contact birds in these groups were infected, resulting also in a BID50 of 3 log10 EID50. In contrast to ducks, all geese inoculated with the low, medium and high doses of the H5N8 HPAI virus were infected and transmitted the virus to contacts, with a resulting BID50 of < 2 log10 EID50. These findings support the conclusion that geese were more susceptible than ducks to infection by the H5N8 virus.
Although most ducks and geese in this study survived virus infection and showed only minimal clinical signs, occasionally neurological signs were present in both species but were more common and severe in geese. Virus was detected by qRRT-PCR and IHC in the brain of both ducks and geese, but virus titers were higher and presence of viral antigen was more common in the brain of infected geese, especially in geese infected with the H5N2 virus. In general, virus replication in tissues was higher in the geese than in the ducks. This higher virus replication and more severe clinical signs in geese compared to ducks has been also observed in other studies comparing H5N1 HPAI virus infections side by side using these species [31, 32]. In addition, geese shed both viruses for longer times and in higher titers, especially by the CL route, than ducks, indicating that the viruses tested in this study replicated better in this species.
The Gs/GD H5N1 HPAI viruses developed the unique capacity among HPAI viruses to infect and cause disease in domestic waterfowl and wild birds producing a range of syndromes from asymptomatic infections to systemic disease and death [31]. The pathogenicity of Gs/GD lineage H5N1 HPAI viruses in waterfowl is associated with the efficiency of virus replication [43]. Viral dissemination to the brain, leading to severe neurological dysfunction, is considered one of the causes of the high virulence of H5N1 viruses in ducks, but lesions to other important organs could lead to multi-organ failure and death [44,45,46,47]. In addition to the virus strain, the susceptibility of wild and domestic waterfowl to H5N1 HPAI virus infection and the presentation of disease vary depending on other factors, including the age and species of the birds and management practices [19, 48]. In wild geese, neurological disease along with systemic virus replication and mortality was observed following experimental inoculation with Gs/GD lineage H5N1 HPAI viruses [49, 50]. Canada geese (Branta canadiensis) experienced sporadic deaths in natural H5N1 HPAI virus outbreaks [51,52,53,54], and have shown high susceptibility to the virus as evidenced by systemic replication and high mortality rates after experimental infection [34, 50, 55], although age-related differences in susceptibility were described [56]. Similar findings have been reported for other species of wild geese, including bar-headed geese (Anser indicus) and cackling geese (B. utchinsii) [49, 57]. Domestic geese naturally infected with H5N1 HPAI virus showed severe clinical signs including neurological signs [33]. Domestic geese (Anser anser var. domestica) experimentally infected with H5N1 viruses presented neurological signs and 40–50% mortality [34]. Similarly, domestic White Chinese geese (Anser cygnoides) showed high mortality when experimentally infected [30]. However, Embden geese (Anser anser var. domestica) and Graylag geese (Anser anser) inoculated with different H5N1 HPAI viruses developed neurological signs, but lacked mortality [31, 32]. These observations indicate that both the virus strain and waterfowl species affect the outcome of infection with Gs/GD lineage H5 viruses, similar to what was observed in our study.
Most Pekin ducks infected with either of the H5 viruses examined in this study had high body temperatures at 2 dpi when compared to controls, but this difference was only significant in the H5N8 group. An increase in body temperature after infection with the same H5N8 virus was seen in our previous studies with mallards [58, 59], and it has also been reported with H5N1 virus infections in several breeds of Anas platyrrhynchs var. domestica ducks (Pekin, Mallards, Black Runners, Rouen, Khaki Campbell), but not in Muscovy ducks (Cairina moschata) [60, 61]. Similar to Muscovy ducks, no increase in body temperature was observed in the geese, indicating differences in the innate immune response between Pekin ducks and geese, which could also explain the differences in disease severity. Most surviving geese examined at 11 dpi did not have antibodies against the challenge virus, contrary to most ducks, also indicating differences in humoral immune responses between these two species. This low seroconversion rate in geese after AI virus infection was also reported in our previous study examining the pathogenesis of a H7N9 virus in different avian species [62]. The immunological differences observed between waterfowl species could affect virus infection, replication, tissue tropism, and virus clearing, consequently affecting the severity of the pathogenic outcome observed.
When comparing the present study with our previous studies examining the pathobiology of the same H5N8 and H5N2 HPAI viruses in mallards, we observe that both viruses are more infectious for mallards (BID50 of < 2 log10 EID50) than for Pekin ducks (3 log10 EID50) [58, 59]. Similar to Pekin ducks, infected mallards showed minimal clinical signs and transmitted the viruses to all contacts; however, mallards shed virus for longer than Pekin ducks. Interestingly, when examining two H5N2 HPAI viruses isolated later in the U.S. outbreak from commercial poultry, both had a similar high infectivity as the index H5N2 virus in mallards, but one of the viruses showed lower replication and one caused some mortality when given at high doses [58]. These results show that individual H5N2 Gs/GD clade 2.3.4.4 viruses have different pathobiology in infected ducks, and that the duck type can also affect the pathogenic outcome of infection.
Likewise, a range of pathobiological outcomes, from no clinical signs to severe disease including neurological signs, were observed in wild and domestic ducks experimentally inoculated with H5N8 viruses of the same Gs/GD lineage (clade 2.3.4.4), with mortality rates varying from 0 to 20% [1, 29, 58, 59, 63,64,65,66,67,68], and the viruses transmitting efficiently to naïve contacts [63,64,65, 67, 68]. A study describing the pathobiologic characteristics of a H5N1 virus isolated in Canada also belonging to HA clade 2.3.4.4, found that the virus was highly pathogenic to juvenile Muscovy ducks and adult Chinese geese causing systemic infections with neurological signs and mortality (36 and 22% respectively) in both species [69]. The virus was also efficiently transmitted and caused mortality (40 and 80% respectively) in naïve contact ducks and geese of the same species.
Natural infections of domestic ducks with H5N8 HPAI viruses have been associated with disease and mortality, but such mortality is typically low. Hungary reported a H5N8 HPAI outbreak during late winter of 2015 at a Pekin duck fattening facility [70]. In addition to increased mortality in the flock and respiratory symptoms, the affected birds showed lethargy and neurological signs. The H5N8 HPAI outbreak in Korea in 2014 mostly resulted in drops in egg production [63], and the outbreak in the United Kingdom in 2014 showed a gradual reduction in egg production and mild increased mortality over a 7-day period [24]. Similarly, an outbreak with H5N8 HPAI virus in commercial Pekin ducks in California in early 2015 resulted in decreased feed consumption, moderate increase in mortality and severe neurological signs in 2% of the ducks [23]. In these natural infections other factors could have increased the severity of disease presentation. In line with this, fungal and bacterial lesions probably exacerbated the mortality and clinical presentation in the outbreak in the UK, motivating disease investigation [24]. If concurrent infections with other pathogens or added stress under intensive conditions do not prompt an avian notifiable disease investigation, the virus can continue to circulate with mild clinical signs or asymptomatically in domestic waterfowl and potentially spread to other holdings, resulting in a more extensive outbreak of HPAI.
As the Gs/GD H5 HPAI viruses continue to evolve and reassort, antigenic and genetic divergent strains have emerged, many expressing distinct pathobiological features and increased virulence for waterfowl. New H5N8 reassortants recently detected in Europe have caused disease and death in both domestic and wild ducks [15, 16], these viruses appearing to be more pathogenic for waterfowl and other wild bird species than the 2014–2015 H5N8 HPAI viruses.
In conclusion, the results of the present study indicate that infection of naïve domestic Pekin ducks and Chinese geese with clade 2.3.4.4 H5 HPAI viruses resulted in efficient virus replication and transmission to contacts. Mortality was low and clinical signs were uncommon, consisting mostly of mild neurological signs. In the field, clinical signs could be exacerbated by other infectious and non-infectious factors. Our findings emphasize the need to implement and improve active surveillance in domestic waterfowl (backyard and commercial), and increase biosecurity compliance to reduce direct and indirect contact between poultry and wild waterfowl in order to detect, prevent and control AI in poultry.
Abbreviations
- AI:
-
avian influenza
- Gs/GD:
-
A/goose/Guangdong/1/1996
- dpi:
-
day post-inoculation
- dpe:
-
days post-exposure
- ECE:
-
embryonating chicken eggs
- HA:
-
hemagglutinin
- HI:
-
hemagglutination inhibition
- HPAI:
-
highly pathogenic avian influenza
- LPAI:
-
low pathogenic avian influenza
- BID50 :
-
mean bird infectious dose
- MDT:
-
mean death time
- EID50 :
-
mean egg infectious dose
- na:
-
not applicable
- qRRT-PCR:
-
quantitative real-time RT-PCR
- SPF:
-
specific pathogen free
References
Zhao K, Gu M, Zhong L, Duan Z, Zhang Y, Zhu Y, Zhao G, Zhao M, Chen Z, Hu S, Liu W, Liu X, Peng D, Liu X (2013) Characterization of three H5N5 and one H5N8 highly pathogenic avian influenza viruses in China. Vet Microbiol 163:351–357
Wong FY, Phommachanh P, Kalpravidh W, Chanthavisouk C, Gilbert J, Bingham J, Davies KR, Cooke J, Eagles D, Phiphakhavong S, Shan S, Stevens V, Williams DT, Bounma P, Khambounheuang B, Morrissy C, Douangngeun B, Morzaria S (2015) Reassortant highly pathogenic influenza A(H5N6) virus in Laos. Emerg Infect Dis 21:511–516
Verhagen JH, Herfst S, Fouchier RA (2015) Infectious disease. How a virus travels the world. Science 347:616–617
Lee DH, Torchetti MK, Winker K, Ip HS, Song CS, Swayne DE (2015) Intercontinental spread of Asian-origin H5N8 to North America through Beringia by migratory birds. J Virol 89:6521–6524
Ip HS, Torchetti MK, Crespo R, Kohrs P, DeBruyn P, Mansfield KG, Baszler T, Badcoe L, Bodenstein B, Shearn-Bochsler V, Killian ML, Pedersen JC, Hines N, Gidlewski T, DeLiberto T, Sleeman JM (2015) Novel Eurasian highly pathogenic avian influenza A H5 viruses in wild birds, Washington, USA, 2014. Emerg Infect Dis 21:886–890
Wu H, Peng X, Xu L, Jin C, Cheng L, Lu X, Xie T, Yao H, Wu N (2014) Novel reassortant influenza A(H5N8) viruses in domestic ducks, eastern China. Emerg Infect Dis 20:1315–1318
Lee YJ, Kang HM, Lee EK, Song BM, Jeong J, Kwon YK, Kim HR, Lee KJ, Hong MS, Jang I, Choi KS, Kim JY, Lee HJ, Kang MS, Jeong OM, Baek JH, Joo YS, Park YH, Lee HS (2014) Novel reassortant influenza A(H5N8) viruses, South Korea, 2014. Emerg Infect Dis 20:1087–1089
Jeong J, Kang HM, Lee EK, Song BM, Kwon YK, Kim HR, Choi KS, Kim JY, Lee HJ, Moon OK, Jeong W, Choi J, Baek JH, Joo YS, Park YH, Lee HS, Lee YJ (2014) Highly pathogenic avian influenza virus (H5N8) in domestic poultry and its relationship with migratory birds in South Korea during 2014. Vet Microbiol 173:249–257
Pasick J, Berhane Y, Joseph T, Bowes V, Hisanaga T, Handel K, Alexandersen S (2015) Reassortant highly pathogenic influenza A H5N2 virus containing gene segments related to Eurasian H5N8 in British Columbia, Canada, 2014. Sci Rep 5:9484
United States Department of Agriculture, Animal and Plant Health Inspection Service. Highly pathogenic H5 avian influenza confirmed in wild birds in Washington State H5N2 found in Northern pintail ducks and H5N8 found in captive Gyrfalcons. https://www.usda.gov/wps/portal/usda/usdahome?contentid=2014/12/0273.xml&contentidonly=true. Accessed 15 Jan 2017
United States Department of Agriculture, animal and plant health inspection service. Animal disease information, avian influenza, HPAI 2014/2015 confirmed detections. https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/animal-disease-information/avian-influenza-disease/sa_detections_by_states/hpai-2014-2015-confirmed-detections. Accessed 15 Jan 2017
Cattoli G, Monne I, Fusaro A, Joannis TM, Lombin LH, Aly MM, Arafa AS, Sturm-Ramirez KM, Couacy-Hymann E, Awuni JA, Batawui KB, Awoume KA, Aplogan GL, Sow A, Ngangnou AC, El Nasri Hamza IM, Gamatié D, Dauphin G, Domenech JM, Capua I (2009) Highly pathogenic avian influenza virus subtype H5N1 in Africa: a comprehensive phylogenetic analysis and molecular characterization of isolates. PLoS One 4:e4842
Keawcharoen J, van Riel D, van Amerongen G, Bestebroer T, Beyer WE, van Lavieren R, Osterhaus AD, Fouchier RA, Kuiken T (2008) Wild ducks as long-distance vectors of highly pathogenic avian influenza virus (H5N1). Emerg Infect Dis 14:600–607
Lee DH, Sharshov K, Swayne DE, Kurskaya O, Sobolev I, Kabilov M, Alekseev A, Irza V, Shestopalov A (2017) Novel reassortant clade 2.3.4.4 avian influenza A(H5N8) virus in wild aquatic birds, Russia, 2016. Emerg Infect Dis 23:359–360
Pohlmann A, Starick E, Harder T, Höper D, Globig A, Staubach C, Dietze K, Grund C, Strebelow G, Ulrich RG, Schinköthe J, Teifke JP, Conraths FJ, Mettenleiter TC, Beer M (2017) Outbreaks among wild birds and domestic poultry caused by reassorted influenza A(H5N8) clade 2.3.4.4 viruses, Germany, 2016. Emerg Infect Dis 23:633–636
Food and Agricultural Organization of the United Nations. Animal Production and Health. H5N8 HPAI Global situation update. http://www.fao.org/ag/againfo/programmes/en/empres/H5N8/situation_update.html. Accessed 15 Jan 2017
Gilbert M, Chaitaweesub P, Parakamawongsa T, Premashthira S, Tiensin T, Kalpravidh W, Wagner H, Slingenbergh J (2006) Free-grazing ducks and highly pathogenic avian influenza, Thailand. Emerg Infect Dis 12:227–234
Henning J, Wibawa H, Morton J, Usman TB, Junaidi A, Meers J (2010) Scavenging ducks and transmission of highly pathogenic avian influenza, Java, Indonesia. Emerg Infect Dis 16:1244–1250
Songserm T, Jam-on R, Sae-Heng N, Meemak N, Hulse-Post DJ, Sturm-Ramirez K, Webster RG (2006) Domestic ducks and H5N1 influenza epidemic, Thailand. Emerg Infect Dis 12:575–581
Hill SC, Lee YJ, Song BM, Kang HM, Lee EK, Hanna A, Gilbert M, Brown IH, Pybus OG (2015) Wild waterfowl migration and domestic duck density shape the epidemiology of highly pathogenic H5N8 influenza in the Republic of Korea. Infect Genet Evol 34:267–277
United States Department of Agriculture. HPAI 2014-2015 Infected premises. https://www.aphis.usda.gov/animal_health/animal_dis_spec/poultry/downloads/hpai-positive-premises-2014-2015.pdf. Accessed 15 Jan 2017
United States Department of Agriculture, animal and plant health inspection service. Stakeholder announcement. H5N8 found in backyard poultry in Oregon https://www.aphis.usda.gov/publications/animal_health/2014/SA_H5N8_Oregon.pdf. Accessed 15 Jan 2017
Shivaprasad HSS, Carnaccini S, Crossley B, Senties-Cue G, Chin R (2016) An overview of outbreaks of LPAI and HPAI H5N8 in commercial poultry in California. In: Sixty-fifth Western poultry disease conference, Vancouver, Canada April 24–27:261
Nuñez A, Brookes SM, Reid SM, Garcia-Rueda C, Hicks DJ, Seekings JM, Spencer YI, Brown IH (2016) Highly pathogenic avian influenza H5N8 clade 2.3.4.4 virus: equivocal pathogenicity and implications for surveillance following natural infection in Breeder ducks in the United Kingdom. Transbound Emerg Dis 63:5–9
HPAI outbreaks continue in France (2016) Vet Rec 179:158
Pantin-Jackwood MJ, Swayne DE (2009) Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev Sci Tech 28:113–136
Alexander DJ, Allan WH, Parsons DG, Parsons G (1978) The pathogenicity of four avian influenza viruses for fowls, turkeys and ducks. Res Vet Sci 24:242–247
Capua I, Mutinelli F (2001) Mortality in Muscovy ducks (Cairina moschata) and domestic geese (Anser anser var. domestica) associated with natural infection with a highly pathogenic avian influenza virus of H7N1 subtype. Avian Pathol 30:179–183
Lee DH, Kwon JH, Noh JY, Park JK, Yuk SS, Erdene-Ochir TO, Lee JB, Park SY, Choi IS, Lee SW, Song CS (2016) Pathogenicity of the Korean H5N8 highly pathogenic avian influenza virus in commercial domestic poultry species. Avian Pathol 45:208–211
Webster RG, Guan Y, Peiris M, Walker D, Krauss S, Zhou NN, Govorkova EA, Ellis TM, Dyrting KC, Sit T, Perez DR, Shortridge KF (2002) Characterization of H5N1 influenza viruses that continue to circulate in geese in Southeastern China. J Virol 76:118–126
Kwon YK, Thomas C, Swayne DE (2010) Variability in pathobiology of South Korean H5N1 high-pathogenicity avian influenza virus infection for 5 species of migratory waterfowl. Vet Pathol 47:495–506
Perkins LE, Swayne DE (2002) Pathogenicity of a Hong Kong-origin H5N1 highly pathogenic avian influenza virus for emus, geese, ducks, and pigeons. Avian Dis 46:53–63
Szeredi L, Dán A, Pálmai N, Ursu K, Bálint A, Szeleczky Z, Ivanics E, Erdélyi K, Rigó D, Tekes L, Glavits R (2010) Tissue tropism of highly pathogenic avian influenza virus subtype H5N1 in naturally infected mute swans (Cygnus Olor), domestic geese (Aser Anser var. domestica), pekin ducks (Anas platyrhynchos) and mulard ducks (Cairina moschata x anas platyrhynchos). Acta Vet Hung 58:133–145
Smietanka K, Minta Z, Reichert M, Olszewska M, Wyrostek K, Jóźwiak M, van den Berg T (2013) Experimental infection of juvenile domestic and Canada geese with two different clades of H5N1 high pathogenicity avian influenza virus. Vet Microbiol 163:235–241
Yuan R, Cui J, Zhang S, Cao L, Liu X, Kang Y, Song Y, Gong L, Jiao P, Liao M (2014) Pathogenicity and transmission of H5N1 avian influenza viruses in different birds. Vet Microbiol 168:50–59
United States Department of Agriculture. Wild bird highly pathogenic avian influenza cases in the United States. https://www.aphis.usda.gov/wildlife_damage/downloads/DEC%202014%20-%20JUNE%202015%20WILD%20BIRD%20POSITIVE%20HIGHLY%20PATHOGENIC%20AVIAN%20INFLUENZA%20CASES%20IN%20THE%20UNITED%20STATES.pdf. Accessed 15 Jan 2017
Lee MS, Chen LH, Chen YP, Liu YP, Li WC, Lin YL, Lee F (2016) Highly pathogenic avian influenza viruses H5N2, H5N3, and H5N8 in Taiwan in 2015. Vet Microbiol 187:50–57
Spackman E, Killian ML (2014) Avian influenza virus isolation, propagation, and titration in embryonated chicken eggs. Methods Mol Biol 1161:125–140
Pantin-Jackwood MJ (2014) Immunohistochemical staining of influenza virus in tissues. Methods Mol Biol 1161:51–58
Pedersen JC (2014) Hemagglutination-inhibition assay for influenza virus subtype identification and the detection and quantitation of serum antibodies to influenza virus. Methods Mol Biol 1161:11–25
Reed LJ, Muench H (1938) A simple method for estimating fifty percent endpoints. Am J Epidemiol 27:493–497
Spackman E, Senne DA, Bulaga LL, Myers TJ, Perdue ML, Garber LP, Lohman K, Daum LT, Suarez DL (2003) Development of real-time RT-PCR for the detection of avian influenza virus. Avian Dis 47:1079–1082
Wasilenko JL, Arafa AM, Selim AA, Hassan MK, Aly MM, Ali A, Nassif S, Elebiary E, Balish A, Klimov A, Suarez DL, Swayne DE, Pantin-Jackwood MJ (2012) Pathogenicity of two Egyptian H5N1 highly pathogenic avian influenza viruses in domestic ducks. Arch Virol 156:37–51
Kajihara M, Sakoda Y, Soda K, Minari K, Okamatsu M, Takada A, Kida H (2013) The PB2, PA, HA, NP, and NS genes of a highly pathogenic avian influenza virus A/whooper swan/Mongolia/3/2005 (H5N1) are responsible for pathogenicity in ducks. Virol J 10:45
Bingham J, Green DJ, Lowther S, Klippel J, Burggraaf S, Anderson DE, Wibawa H, Hoa DM, Long NT, Vu PP, Middleton DJ, Daniels PW (2009) Infection studies with two highly pathogenic avian influenza strains (Vietnamese and Indonesian) in Pekin ducks (Anas platyrhynchos), with particular reference to clinical disease, tissue tropism and viral shedding. Avian Pathol 38:267–278
Löndt BZ, Nunez A, Banks J, Nili H, Johnson LK, Alexander DJ (2008) Pathogenesis of highly pathogenic avian influenza A/turkey/Turkey/1/2005 H5N1 in Pekin ducks (Anas platyrhynchos) infected experimentally. Avian Pathol 37:619–627
Hulse-Post DJ, Sturm-Ramirez KM, Humberd J, Seiler P, Govorkova EA, Krauss S, Scholtissek C, Puthavathana P, Buranathai C, Nguyen TD, Long HT, Naipospos TS, Chen H, Ellis TM, Guan Y, Peiris JS, Webster RG (2005) Role of domestic ducks in the propagation and biological evolution of highly pathogenic H5N1 influenza viruses in Asia. Proc Natl Acad Sci USA 102:10682–10687
Pantin-Jackwood MJ, Smith DM, Wasilenko JL, Cagle C, Shepherd E, Sarmento L, Kapczynski DR, Afonso CL (2012) Effect of age on the pathogenesis and innate immune responses in Pekin ducks infected with different H5N1 highly pathogenic avian influenza viruses. Virus Res 167:196–206
Brown JD, Stallknecht DE, Swayne DE (2008) Experimental infection of swans and geese with highly pathogenic avian influenza virus (H5N1) of Asian lineage. Emerg Infect Dis 14:136–142
Pasick J, Berhane Y, Embury-Hyatt C, Copps J, Kehler H, Handel K, Babiuk S, Hooper-McGrevy K, Li Y, Le Mai Q, Lien Phuong S (2007) Susceptibility of Canada geese (Branta canadensis) to highly pathogenic avian influenza virus (H5N1). Emerg Infect Dis 13:1821–1827
Ellis TM, Bousfield RB, Bissett LA, Dyrting KC, Luk GS, Tsim ST, Sturm-Ramirez K, Webster RG, Guan Y, Malik Peiris JS (2004) Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2002. Avian Pathol 33:492–505
Teifke JP, Klopfleisch R, Globig A, Starick E, Hoffmann B, Wolf PU, Beer M, Mettenleiter TC, Harder TC (2007) Pathology of natural infections by H5N1 highly pathogenic avian influenza virus in mute (Cygnus olor) and whooper (Cygnus cygnus) swans. Vet Pathol 44:137–143
Liu J, Xiao H, Lei F, Zhu Q, Qin K, Zhang XW, Zhang XL, Zhao D, Wang G, Feng Y, Ma J, Liu W, Wang J, Gao GF (2005) Highly pathogenic H5N1 influenza virus infection in migratory birds. Science 309:1206
Chen H, Smith GDJ, Zhang SY, Qin K, Wang J, Li KS, Webster RG, Peiris JSM, Guan Y (2005) Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature 436:191–192
Berhane Y, Embury-Hyatt C, Leith M, Kehler H, Suderman M, Pasick J (2014) Pre-exposing Canada Geese (Branta canadensis) to a low-pathogenic H1N1 avian influenza virus protects them against H5N1 HPAI virus challenge. J Wildl Dis 50:84–97
Neufeld JL, Embury-Hyatt C, Berhane Y, Manning L, Ganske S, Pasick J (2009) Pathology of highly pathogenic avian influenza virus (H5N1) infection in Canada geese (Branta canadensis): preliminary studies. Vet Pathol 46:966–970
Nemeth NM, Brown JD, Stallknecht DE, Howerth EW, Newman SH, Swayne DE (2013) Experimental infection of bar-headed geese (Anser indicus) and ruddy shelducks (Tadorna ferruginea) with a clade 2.3.2 H5N1 highly pathogenic avian influenza virus. Vet Pathol 50:961–970
DeJesus E, Costa-Hurtado M, Smith D, Lee DH, Spackman E, Kapczynski DR, Torchetti MK, Killian ML, Suarez DL, Swayne DE, Pantin-Jackwood MJ (2016) Changes in adaptation of H5N2 highly pathogenic avian influenza H5 clade 2.3.4.4 viruses in chickens and mallards. Virology 499:52–64
Pantin-Jackwood MJ, Costa-Hurtado M, Shepherd E, DeJesus E, Smith D, Spackman E, Kapczynski DR, Suarez DL, Stallknecht D, Swayne DE (2016) Pathogenicity and transmission of H5 and H7 highly pathogenic avian influenza viruses in mallards. J Virol 90:9967–9982
Pantin-Jackwood M, Swayne DE, Smith D, Shepherd E (2013) Effect of species, breed and route of virus inoculation on the pathogenicity of H5N1 highly pathogenic influenza (HPAI) viruses in domestic ducks. Vet Res 44:62
Cagle C, To TL, Nguyen T, Wasilenko J, Adams SC, Cardona CJ, Spackman E, Suarez DL, Pantin-Jackwood MJ (2011) Pekin and Muscovy ducks respond differently to vaccination with a H5N1 highly pathogenic avian influenza (HPAI) commercial inactivated vaccine. Vaccine 29:6549–6557
Pantin-Jackwood MJ, Miller PJ, Spackman E, Swayne DE, Susta L, Costa-Hurtado M, Suarez DL (2014) Role of poultry in the spread of novel H7N9 influenza virus in China. J Virol 88:5381–5390
Kang HM, Lee EK, Song BM, Jeong J, Choi JG, Jeong J, Moon OK, Yoon H, Cho Y, Kang YM, Lee HS, Lee YJ (2015) Novel reassortant influenza A(H5N8) viruses among inoculated domestic and wild ducks, South Korea, 2014. Emerg Infect Dis 21:298–304
Kim YI, Pascua PNQ, Kwon HI, Lim GJ, Kim EH, Yoon SW, Park SJ, Kim SM, Choi EJ, Si YJ, Lee OJ, Shim WS, Kim SW, Mo IP, Bae Y, Lim YT, Sung MH, Kim CJ, Webby RJ, Webster RG, Choi YK (2014) Pathobiological features of a novel, highly pathogenic avian influenza A(H5N8) virus. Emerg Microbes Infect 3:e75
Sun H, Pu J, Hu J, Liu L, Xu G, Gao GF, Liu X, Liu J (2016) Characterization of clade 2.3.4.4 highly pathogenic H5 avian influenza viruses in ducks and chickens. Vet Microbiol 182:116–122
Kanehira K, Uchida Y, Takemae N, Hikono H, Tsunekuni R, Saito T (2015) Characterization of an H5N8 influenza A virus isolated from chickens during an outbreak of severe avian influenza in Japan in April 2014. Arch Virol 160:1629–1643
Kang HM, Lee EK, Song BM, Heo GB, Jung J, Jang I, Bae YC, Jung SC, Lee YJ (2017) Experimental infection of mandarin duck with highly pathogenic avian influenza A (H5N8 and H5N1) viruses. Vet Microbiol 198:59–63
Li J, Gu M, Liu D, Liu B, Jiang K, Zhong L, Liu K, Sun W, Hu J, Wang X, Hu S, Liu X, Liu X (2016) Phylogenetic and biological characterization of three K1203 (H5N8) like avian influenza A virus reassortants in China in 2014. Arch Virol 161:289–302
Berhane Y, Kobasa D, Embury-Hyatt C, Pickering B, Babiuk S, Joseph T, Bowes V, Suderman M, Leung A, Cottam-Birt C, Hisanaga T, Pasick J (2016) Pathobiological haracterization of a novel reassortant highly pathogenic H5N1 virus isolated in British Columbia, Canada, 2015. Sci Rep 6:23380
Banyái K, Bistyák AT, Thuma A, Gyuris E, Ursu K, Marton S, Farkas SL, Hortobágyi E, Bacsadi Á, Dán Á (2016) Neuroinvasive influenza virus A(H5N8) in fattening ducks, Hungary, 2015. Infect Genet Evol 43:418–423
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MPJ conceived this project. MPJ, MCH, EDJ, and DS conducted the animal experiments and sample processing. MPJ, MCH, DS, KB analyzed the data. MPJ, MCH, EDJ, DS, KB, and DES help write and reviewed the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors appreciate the technical assistance provided by Scott Lee and Nikolai Lee, and the animal care provided by Roger Brock, Keith Crawford, and Gerald Damron in conducting these studies.
Ethics approval
All procedures were performed according to the requirements of protocols approved by the Institutional Animal Care and Use Committee and the Institutional Biosecurity Committee.
Funding
This research was supported by the USDA/ARS CRIS Project 6612-32000-063-00D and by Center of Research in Influenza Pathogenesis (CRIP) an NIAID funded Center of Excellence in Influenza Research and Surveillance (CEIRS, Contract HHSN272201400008C). Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the USDA or NIH. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA. USDA is an equal opportunity provider and employer.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Pantin-Jackwood, M.J., Costa-Hurtado, M., Bertran, K. et al. Infectivity, transmission and pathogenicity of H5 highly pathogenic avian influenza clade 2.3.4.4 (H5N8 and H5N2) United States index viruses in Pekin ducks and Chinese geese. Vet Res 48, 33 (2017). https://doi.org/10.1186/s13567-017-0435-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-017-0435-4