
Abstract
Background
Duchenne muscular dystrophy (DMD) is a severe form of muscular dystrophy without an effective treatment, caused by mutations in the DMD gene, leading to the absence of dystrophin. DMD results in muscle weakness, loss of ambulation, and death at an early age. Metabolomics studies in mdx mice, the most used model for DMD, reveal changes in metabolites associated with muscle degeneration and aging. In DMD, the tongue muscles exhibit unique behavior, initially showing partial protection against inflammation but later experiencing fibrosis and loss of muscle fibers. Certain metabolites and proteins, like TNF-α and TGF-β, are potential biomarkers for dystrophic muscle characterization.
Methods
To investigate disease progression and aging, we utilized young (1 month old) and old (21–25 months old) mdx and wild-type tongue muscles. Metabolite changes were analyzed using 1H nuclear magnetic resonance, while TNF-α and TGF-β were assessed using Western blotting to examine inflammation and fibrosis. Morphometric analysis was conducted to assess the extent of myofiber damage between groups.
Results
The histological analysis of the mid-belly tongue showed no differences between groups. No differences were found between the concentrations of metabolites from wild-type or mdx whole tongues of the same age. The metabolites alanine, methionine, and 3-methylhistidine were higher, and taurine and glycerol were lower in young tongues in both wild type and mdx (p < 0.001). The metabolites glycine (p < 0.001) and glutamic acid (p = 0.0018) were different only in the mdx groups, being higher in young mdx mice. Acetic acid, phosphocreatine, isoleucine, succinic acid, creatine, and the proteins TNF-α and TGF-β had no difference in the analysis between groups (p > 0.05).
Conclusions
Surprisingly, histological, metabolite, and protein analysis reveal that the tongue of old mdx remains partially spared from the severe myonecrosis observed in other muscles. The metabolites alanine, methionine, 3-methylhistidine, taurine, and glycerol may be effective for specific assessments, although their use for disease progression monitoring should be cautious due to age-related changes in the tongue muscle. Acetic acid, phosphocreatine, isoleucine, succinate, creatine, TNF-α, and TGF-β do not vary with aging and remain constant in spared muscles, suggesting their potential as specific biomarkers for DMD progression independent of aging.
Similar content being viewed by others


Introduction
Duchenne muscular dystrophy (DMD) affects approximately 1 in 3600–6000 male live births [1] and is a severe form of muscular dystrophy without an effective treatment [2]. It is an X-linked recessive disease [2] caused by mutations in the DMD gene [3] which leads to the absence of dystrophin [4].
The lack of dystrophin causes continuous loss of muscle strength, myofiber damage, chronic inflammation, progressive fibrosis, and muscle stem cell dysfunction. This dystrophic scenario leads to a loss of ambulation in the early teens to 20s. Although patients’ life expectancy has improved with current standards of cardioprotective care and respiratory support, they often die around the third or fourth decade of life, mainly due to cardiac and respiratory complications [2, 5, 6].
The mdx mice is the most used animal model for research on DMD [7, 8]. Around 20 days old, these animals begin to show their first signs of muscle degeneration and regeneration [9]. The mdx mice have reduced life spans, by about 17 to 19% compared to wild type. At 26 months, average lifespans, the muscles present typical dystrophic characteristics: loss of muscle fibers with increased fibrosis, fat infiltration, necrotic fibers, and regenerated fibers [10].
Metabolomics are the quantitative analysis of metabolites produced by an organism under certain conditions. Metabolomics provide an integrated view of biochemical pathways in complex organisms, thus producing a more detailed and systematic overview of the cellular processes and its response to diseases. Therefore, this approach is essential for the definition of personalized medicine, through the establishment of metabolite profiles and biomarkers for certain pathological states [11]. Studies in mdx mice show changes in metabolites related to the progression of muscle degeneration and aging [12,13,14].
In DMD, the muscles of the oral cavity are also affected, causing dysphagia in late stages of the disease, which worsens with advancing age and disease progression [15, 16]. In mdx mice, the tongue muscles show an unusual behavior as the disease progresses compared to other muscles. In 3-month mdx mice, inflammatory cells were hardly found in the tongue, unlike other masticatory muscles. In addition, the expression of collagen did not change in the tongue of these mdx, while it was three times higher in masseter muscle, when compared to the control [17]. These results indicate partial protection of this muscle against myonecrosis and inflammation at 3 months of age. However, at 23 months of age, foci of inflammation and fibrosis were identified [18]. Additionally, Chamberlain et al. (2007) [10] described the tongue of the mdx as the second most affected muscle at 26 months of age, due to fibrosis and the loss of fibers in the central portion of the muscle, just after the diaphragm.
Some metabolites from dystrophic mice were analyzed by nuclear magnetic resonance (NMR) in different types of tissues and have been suggested to determine biomarkers for the state of muscle fibers [19]. Martins-Bach and colleagues [13] studied the diaphragm and quadriceps muscles, in 3-month and 6-month-old dystrophic and non-dystrophic mice, and found that the metabolites glutamate, glutamine, succinate, isoleucine, acetate, alanine, glycerol, carnosine, taurine, glycine, methionine, and creatine had alterations when compared to same age controls and also across time [13]. Griffin and colleagues [20] studied cardiac, cortex, cerebellum, diaphragm, and soleus tissue and stated that different ratios of the founding metabolites were responsible to distinguish the dystrophic and non-dystrophic tissue, including the following: phosphocholine/taurine, glutamate/creatine, and taurine/creatine [20]. Taurine itself has already been indicated as a biomarker for dystrophin deficiency in the quadriceps muscle, liver, and kidney [21].
Besides metabolites, there are some proteins that are well established in the current literature related to inflammation and fibrosis, like tumor necrosis factor alpha (TNF-α) and transforming growth factor beta (TGF-β), respectively. These proteins are very promising biomarkers for the dystrophic muscle characterization, for their relation to the myonecrosis as seen in the mdx animal model and human patients [22]. Given tongue’s unusual behavior within DMD progression, being spared in young mice and severely affected in old mice [10, 17], we evaluated metabolites and proteins in the tongue muscles, within young (30 days old) and old (21 to 25 months old) mdx and wild-type mice. Since the proteins and metabolites chosen were previously reported as possible biomarkers for the characterization of the dystrophy pathology and progression, in muscle and other tissues [13, 19,20,21,22,23,24,25,26,27], the present study aimed to validate these biomarkers in the tongue of mdx mice over time. Surprisingly, our histological results found that the tongue remains partially spared in older mdx mice (21 to 25 months). Our results allowed us to identify biomarkers that change with aging, regardless of the absence of dystrophin. Other biomarkers seem to be excellent candidates to indicate the progression of dystrophinopathy over time, as they do not change with aging and remain constant in the spared muscles. Furthermore, analyzing these possible biomarkers in the tongue at different ages can help to understand the protection mechanisms involved in the pathological process and support the development of future approaches for the diagnosis and monitoring progression of DMD.
Materials and methods
Animals
Male and female, young and old mdx (C57BL/10-DMDmdx/PasUnib), and age-match wild-type mice (C57BL/10ScCr/PasUnib) were obtained and maintained by our institutional animal care facility of Institute of Bioscience (Botucatu) — UNESP. All mouse experimentation was approved by our institution committee and done in accordance with the guidelines of the Brazilian College for Animal Experimentation (protocol n° 1095-CEUA). The animals were divided into four groups: young mdx (1 month old), old mdx (21–25 months old), young wild type (1 month old), and old wild type (21–25 months old).
Tissue harvesting
The animals were euthanized with an overdose of intraperitoneal anesthesia of xylazine hydrochloride (30 mg/kg) and ketamine hydrochloride (300 mg/kg). The tongue was dissected and fixed for histological techniques or frozen in liquid nitrogen for Western blotting and metabolome assays.
Histology
The mid-belly tongue muscles of the young wild type (n = 5), young mdx (n = 5), old wild type (n = 4), and old mdx (n = 6) groups were sectioned and stained with Masson’s trichrome, to distinguish and quantify the areas of fibrosis (FIB) and areas of muscle fibers with peripheral nuclei (PN) and central nuclei (CN). The analyses were performed blindly, and the areas were expressed in relation to the area of total transverse fibers of the section.
Proteins analysis
Proteins related to the mechanisms of inflammation (TNF-α) and fibrosis (TGF-β) were quantified in the whole tongue with the Western blotting assay, as described previously [28]. The values were normalized with the glyceraldehyde 3-phosphate dehydrogenase protein (GAPDH), incubated on the same membrane after routine stripping methods.
Metabolomics by nuclear magnetic resonance (NMR) spectroscopy
In order to verify possible changes in the metabolic profile, metabolomics analysis was performed by NMR spectroscopy of the whole tongue muscles of young (n = 10) and old (n = 5) wild-type mice and young (n = 11) and old (n = 5) mdx mice.
Data acquisition was performed on a Varian INOVA spectrometer operating at a resonance frequency of 1 H of 600 MHz. The samples were homogenized in a methanol/chloroform solution (2:1). After 15 min, a solution of chloroform/Milli-Q water (1:1) was added to the pellet. Then, the samples were centrifuged at 4000 RPM for 20 min. The supernatant was collected and lyophilized. The obtained powder was resuspended in deuterated water (D2O) with trimethylsilyl tetradeuteropropionic acid (TSP, 10 mM). Subsequently, the samples were transferred to a standard NMR tube for spectral analysis. The D2O allowed the device to be monitored and blocked by the device’s resonant frequency. The TSP reference signals were used to assess the quality of the spectra as well as to quantify the identified substances. Proton spectra in one dimension, using pulse sequences, optimized for suppression of the water signal and were collected at 25 °C [29].
Spectrum treatment and identification and quantification of metabolites were performed using the Bruker Topspin 3.1 and Chenomx NMR Suite (Version 7.1; Chenomx Inc., Edmonton, Canada) application packages, in conjunction with the Human Metabolome Database [30] and literature already published. All the chemical shift intervals (ppm) are listed in Table 1. In parallel with the chemometric analysis using the separation of the spectrum in small intervals, the statistical analysis was performed directly with the concentrations of the identified metabolites, through the targeted profiling methodology, developed by the creators of the Chenomx NMR Suite application. In this methodology, the groups of peaks corresponding to each metabolite are identified and quantified using a database of pure substance spectra. We also used the spectral binning methodology, in which the spectra are divided into predefined frequency intervals, while the integrals of the signals within each interval are used in the statistical analysis [31].
Statistical analysis
Statistical analysis was performed through two-way analysis of variance (two-way ANOVA, p ≤ 0.05) with Bonferroni’s post hoc test for histological and Western blotting data.
For the metabolomics analysis, unsupervised interactive principal component analysis (iPCA) was used in order to visualize the complex sample space and multivariate after identification and quantification of metabolites. These discriminant analyses were performed through standard procedures implemented in Pirouette 4.0 (Infometrix, WA, USA) [32]. The input variables will consist of the integral of the area intervals (spectral binning) and/or the concentration values obtained with targeted profiling. Two-way analysis of variance (ANOVA, p ≤ 0.05) with Bonferroni’s post hoc test was performed directly with the concentrations of the identified metabolites, using the MetaboAnalyst platform.
Results
Mid-belly tongue muscles remain spared in old mdx
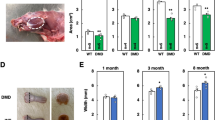
In the qualitative histopathological analysis of the sections, we observed different histological aspects in the muscle fibers. Fibers with peripheral nuclei (PN) were observed, indicating normal muscle tissue status; fibers with central nuclei (CN), indicating regenerated muscle fibers; and areas of fibrosis (FIB), shown in blue by Masson’s trichrome stain (Fig. 1). In the quantitative histological analysis of the tongue of the groups described (Table 2), there was no difference (p > 0.05) in the areas of PN, CN, or FIB, both between ages, young and old, and between lineages, wild type and mdx.

Cross sections of mid-belly tongue muscle. Young wild type (A), young mdx (B), old wild type (C), and old mdx (D) groups. In B, areas of fibrosis highlighted in blue by Masson’s trichrome (*). In C, muscle fibers with a central nucleus (arrow) and muscle fibers with a peripheral nucleus (arrowhead). Scale: 50 \(\mu\)m
Changes in proteins related to inflammation and fibrosis
The quantification of TNF-α and TGF-β in the whole tongue muscle was performed by Western blotting to verify the presence of inflammation and fibrosis, respectively. There was no significant difference in the concentrations of both proteins in the tongue between the groups (two-way ANOVA, TGF-β p = 9.968; TNF-α p = 7.558), as shown in Fig. 2.

A Quantification of TNF-⍺ and B TGF-β in the whole tongue muscles. The quantification of TNF-⍺ and TGF-β was performed by Western blotting analysis in crude extracts of tongue muscles from young wild type, young mdx, old wild type, and old mdx groups. Same blot reprobed for GAPDH as a loading control. Graphs represent the level of proteins normalized to GAPDH levels expressed in arbitrary units. Bars represent standard deviation. No significant difference was observed between the groups (two-way ANOVA with Bonferroni’s post hoc test, p > 0.05)
Changes in the metabolomic profile
The whole tongues of the mice are grouped according to their metabolic profile through the iPCA. It was possible to distinguish the tongues of the same lineage at different ages; however, it was not possible to distinguish mdx mice from wild-type mice, in both young or old for the tongue muscle (Fig 3).

Interactive principal component analysis (iPCA) of whole tongue with the scores plot between groups. Samples spectrum plotted according to age (young, green; old, red) and genotype (mdx ◯; wild type ∆)
No differences were found in the comparison between the concentrations of metabolites from wild-type or mdx whole tongue muscles of the same age, whether young or old (Fig. 4), suggesting that the protection of the dystrophic tongue muscle is observed in both ages. Table 3 summarizes the metabolites responsible for the differentiation of the tongue muscle of same strain mice at different ages. The metabolites methionine, 3-methylhistidine, and alanine were higher in young animals in both wild type and mdx (p < 0.001, Fig. 4D–F), and the metabolites taurine and glycerol were lower in both the young wild-type and mdx groups (p < 0.001, Fig. 4C and G). The metabolites glycine (p < 0.001, Fig. 4A) and glutamic acid (p = 0.0018, Fig. 4B) were different only in the mdx groups, being more concentrated in young mdx mice. The metabolites acetic acid, creatine, isoleucine, succinic acid, and phosphocreatine had no difference in the analysis between groups (p > 0.05, Fig. 4H–L).

Differences in the concentrations of tongue metabolites between groups. C–G Difference between metabolite concentrations between young and old wild-type animals (*). A–G Difference between metabolite concentrations between young and old mdx animals (#), two-way ANOVA with Bonferroni’s post hoc test (p < 0.001). H–L The metabolites acetic acid, creatine, isoleucine, succinic acid, and phosphocreatine showed no differences between groups (p > 0.05)
Discussion
This work aimed to validate biomarkers for diagnosis and progression of DMD through the analysis of metabolites, protein, and histology of the tongue muscles of dystrophic mdx mice and non-dystrophic control mice, aged 30 days (young) and 21–25 months (old).
Dystropathology
Surprisingly, our histological results demonstrated that the mid-belly tongue of old mdx remains partially spared from fibrosis and myonecrosis. Previous studies from 23- and 26-month-old mdx mice described the tongue muscles as displaying significant histological abnormalities, including foci of inflammation and fibrosis, and localized loss of muscle fibers [10, 18]. However, in these studies, the analysis of the tongue muscle was approached qualitatively, yet without specific delineation of the particular region of the tongue analyzed. The region analyzed may interfere with the results, given the complex composition of the tongue muscle, the presence of intrinsic and extrinsic muscles by region, coupled with age-related shifts in MyHC isoform composition and differences in fiber size from the anterior, middle, and posterior regions of the tongue [33,34,35]. In contrast, here, the qualitative and quantitative analysis of mid-belly tongue did not show such severity. This protection was evidenced by metabolomics and Western blotting experiments performed with whole muscle of the tongue, analyzing the biomarker proteins for inflammation and fibrosis, and the concentrations of the metabolites 3-methylhistidine, acetic acid, glutamic acid, alanine, creatine, phosphocreatine, glycerol, glycine, isoleucine, methionine, succinic acid, and taurine already used for dystrophic muscle differentiation and that in this work presented similar results between wild type and mdx. This similarity between the chosen biomarkers was observed in young and old animals, corroborating the histological result.
Kunert-Keil and colleagues [36] report that dystrophic masseter and temporalis muscles are histologically similar to other skeletal muscles involved in the degeneration process, whereas the tongue remains with a milder phenotype. However, from the findings stated in the article, masseter and temporalis are more resistant to the calcium (Ca2+) overload when compared to tongue muscles of 100-day-old mdx. Hence, histological findings assert that inflammatory foci are hardly detectable, and dystrophic tongue contains only 11.2% of regenerated muscle, when compared to masseter, temporalis, and even soleus [36]. In DMD rats, the tongue showed hypertrophy of myofibers with less advanced dystrophic changes until 9 months old compared to masseter. This resistance against degeneration might be related to a higher transcription level of utrophin in tongue of wild-type and DMD rats compared to masseter of wild-type rats [37]. Increased expression of utrophin in the skeletal muscles of mdx mice results in a functional improvement in muscle function [38,39,40]. However, in mdx muscles spared from degeneration, as extraocular and intrinsic laryngeal muscles, the utrophin upregulation is unlikely to be a protective mechanism [41,42,43]. The extraocular muscles remain unaffected even in dystrophin-deficient utrophin double knockout mice [42]. In the intrinsic laryngeal muscles, the reduction of utrophin at 20 months in mdx mice did not instigate any degeneration in muscle fibers [43]. Further investigation of utrophin in partially spared tongue muscles throughout life span would be interesting, as utrophin and concomitant rescue of dystrophin-associated glycoproteins are irregularly correlated with other protected muscles.
The study of biomarkers in a muscle partially spared from the absence of dystrophin allows us to identify dystrophinopathy markers that change with age, regardless of muscle degeneration. Among the possible biomarkers analyzed, the metabolites alanine, methionine, 3-methylhistidine, taurine, and glycerol change with aging, but not between control and dystrophic whole tongue muscles. Thus, it is suggested that these biomarkers may be efficient for specific assessments, but care should be taken when using them to monitor the progression of the disease, since they change throughout life.
The analysis of the metabolic profile of tongue suggests that the muscle aging process has a high impact on its metabolism, regardless of the lineage, since it was possible to distinguish the different ages of animals of the same lineage (Fig. 3). High-resolution 1H NMR spectroscopy has been shown to differentiate skeletal muscle from adult and old mice. In addition, there is a general difference in composition between younger and older muscles in mice [44], and these results are confirmed in the scenario of dystrophinopathy.
It was not possible to distinguish the metabolic profile of the whole tongue between the wild-type and mdx strains. This result corroborates our histological analysis, which showed protection against fibrosis and myonecrosis in the mid-belly tongue of young and old dystrophic animals. The chosen metabolites (3-methylhistidine, acetate, alanine, creatine, glutamic acid, glycerol, glycine, isoleucine, methionine, phosphocreatine, succinic acid, and taurine) were altered between the wild-type and mdx animals in muscles that suffer degeneration, such as the diaphragm, quadriceps, and soleus [13, 20, 21]. Since these metabolites were not different between the strains in this study, it suggests that the protection previously observed in the tongue of 3-month-old adult animals [17] is also observed in young (1 month old) and old (21–25 months old) animals.
Metabolite and protein analysis
Taurine is considered a biomarker for the aging of skeletal muscle in mice [44], corroborating the data from this project, since its concentration was higher in old dystrophic and non-dystrophic mice. Its decrease in young mdx mice, compared to wild type, has already been seen in other muscles, indicating the possibility of a deficient taurine synthesis by the mdx muscles [21]. In the tongue, this decrease occurs both in the mdx and in the wild type compared to older age, corroborating the protection of the tongue muscle in dystrophic pathology. Taurine has an osmoregulatory function that helps to balance intracellular Ca2+ levels, helping with cell integrity and membrane stability [13]. Kunert-Keil et al. (2014) [34] studied the differential expressions of genes involved in Ca2+ homeostasis in dystrophic masticatory muscles and found uneven expressions in the studied muscles. Despite being a priori protected muscle, in its study, the tongue of 100-day-old mice presented levels of expression of Ca2+ regulatory proteins typical of dystrophic muscles. Analyzing Ca2+ regulatory proteins at the age of 21 to 25 months old would provide a better view on the relationship of increased taurine, found in this study, with Ca2+ homeostasis in old mice. In addition, in muscle regeneration, there is an increase in taurine, regardless of the type of muscle or genetic etiology of the damage to the fiber, suggesting that metabolic changes are significant indicators of muscle status [45]. The level of taurine diminishes with advancing age, and reinstating this decline via taurine supplementation protects against age-related muscle mass loss and decreased functionality and also improves both health and lifespan across various species [46,47,48]. Its greater concentration in the old group tongue, in this project, suggests the activation of fiber metabolism to prevent muscle degeneration.
Glycerol is a component of triglycerides (fats and oils) and phospholipids [30]. In muscles affected by DMD, the glycerol concentration was increased in relation to the wild type, at 6 months of age, being one of the metabolites responsible for the differentiation between the dystrophic and non-dystrophic muscles [13]. The results of this work showed that glycerol is more concentrated in the tongue of old animals, both in wild-type and mdx mice, with no difference between strains, suggesting its relationship with the aging process in the tongue mice muscle.
Alanine is a nonessential amino acid resulting from the conversion of pyruvate or the breakdown of DNA and carnosine and anserine dipeptides [30]. It can be used as a fuel for gluconeogenesis directly from muscle tissue and therefore plays an important role in glucose homeostasis [49, 50]. With the progression of dystrophinopathy in affected muscles, there is an increase in energy expenditure [10, 51], for the incorporation of amino acids into new proteins [52]. Therefore, the lower concentration of alanine in the tongue muscle of old animals may indicate incorporation of alanine into proteins for processes of muscle maintenance and regeneration related to age, since the decrease is seen in old dystrophic and non-dystrophic mice.
Methionine is an essential amino acid, substrate for protein synthesis, necessary for normal mammalian growth and development [30]. Martins-Bach et al. (2012) [13] showed that methionine increased with aging in quadriceps muscle samples from control mice from 3 months to 6 months of age. Besides, it showed that methionine concentrations did not change for mdx samples. However, in the whole tongue muscles, with a greater interval between the ages studied, methionine changed for both strains, being more concentrated in younger animals. The growth of the animals can elucidate this difference between the concentrations of methionine throughout the aging of the mice. From the 26th day of life until the 150th–200th, approximately 7 months of age, the mice’s growth is observed, then remaining on a plateau until the end of life [53]. The high concentration of methionine at a young age in this study suggests growth in young animals. In older animals, which are not in the growth phase, the metabolite is decreased. Furthermore, this similarity of methionine concentrations between strains suggests that the tongue muscle in mdx is metabolically closer to wild type, since this metabolite participates in the regulation of the immune system, lipid metabolism, oxidative stress, and other metabolic regulation processes (for review, see [54]). New studies should be carried out to characterize the involvement of this metabolite in the metabolism of DMD.
3-Methylhistidine (3-MeH) is an amino acid present in actin and myosin. It has been determined that more than 90% of body 3-MeH is located in the skeletal muscle. When skeletal muscle is degraded, 3-MeH is released but is not reused for protein synthesis [55]; therefore, the urinary excretion of 3-MeH can be used to indicate muscle protein degradation [56]. Furthermore, there is a progressive loss of skeletal muscle mass and muscle strength with aging [57, 58]. The higher concentration of 3-MeH observed in the tongue of the young groups may reflect the decrease in muscle mass and the consequent reduction in 3-MeH concentrations seen in older muscles.
Muscle degeneration biomarkers, which in other studies proved to be efficient (acetic acid, phosphocreatine, isoleucine, succinic acid, creatine, TNF-α, and TGF-β protein) [13, 19, 23], did not show any difference with aging and in the analysis between strains. The TGF-β protein is actively involved in the proliferation of fibrous connective tissue in the skeletal muscles of patients and mdx mice [26, 27]. In previous studies, the levels of TGF-β have already been related to the presence of fibrous tissue in mdx’s respiratory muscles [23]. Spassov et al. [17] studied changes in the expression of collagen in the masticatory muscles in 100-day-old animals and concluded that there was no difference between the tongue of mdx and control groups. Our results corroborate theirs, due to the equality in the quantification of TGF-β in young animals indicating protection against tissue fibrosis of this muscle in the initial phase of the disease. In old animals, the levels of TGF-β were also equal between strains and suggest that this muscle protection against fibrosis continues until advanced stages of dystrophy in the mice, corroborating the histological results for fibrosis found in our study, for both ages.
TNF-α is a protein mediator of inflammatory response produced by activated macrophages, mast cells, endothelial cells, fibroblasts, myoblasts and myotubes, and injured myofibers [59, 60]. It stimulates the expression of adhesion molecules in endothelial cells, increasing leukocyte recruitment and adhesion. TNF-α is secreted at the site of inflammation and can enter the bloodstream [59]. The TNF-α is related to the progression of DMD, and its absence is related to muscle protection for the disease. Maranhão et al. [23] evidenced the progressive increase in the concentration of TNF-α in mdx diaphragm muscles, in comparison with the control and over age, with 1, 4, and 9 months. The intrinsic muscles of the larynx were identified as muscles spared from dystrophinopathy in the mdx mice, as they did not show signs of muscle damage, degeneration, or regeneration, during the course of the disease [24]. The concentrations of TNF-α in intrinsic muscles of the larynx were comparable to those of the control animal, during the progression of the disease, even in later stages, at 20 months of age [23]. These results corroborate with this work, since the concentrations of TNF-α in the tongue of the mdx animals did not differ from the wild-type animals, in young and old. Messina et al. (2011) [25] analyzed vastus lateralis muscle samples from DMD patients and showed an increase in the concentration of TNF-α in relation to the control group, besides pointing out the increase in its expression with age and disease progression, from 2 to 9 years of age. In this work, the concentration of TNF-α did not increase with aging in tongue in both strains, corroborating with the protection previously mentioned, including with disease progression and aging.
The metabolites glycine and glutamic acid were different only in dystrophic animals, being more concentrated in young mdx mice. Glycine is a nonessential amino acid involved in the production of DNA, phospholipids, and collagen, in addition to being involved in the release of energy [30]. Ham, Gardner, Kennedy, Trieu, Naim, Chee, Alves, Caldow, Lynch, and Koopman [61] demonstrated that glycine supplementation in mdx mice and dystrophin/utrophin double knockout mice can attenuate the progression of dystrophic pathology, as well as improve the effectiveness of prednisolone, the current gold standard treatment for DMD. Anderson and Skrabek [62] studied the heart of mdx mice treated with deflazacort, from a metabolomic perspective. They showed that the concentration of glycine in the mdx heart decreased with disease progression, but with a 2-week high-dose treatment with deflazacort, glycine levels increased above the levels of the mdx placebo group. Glycine supplementation has already been shown to protect against loss of myotubes in nutrient restricted/growth factor restriction models with C2C12 muscle cells in vitro. This in vitro protection has been shown to be dependent on mammalian target of rapamycin complex 1 (mTORC1) signaling [63] and also a specific glycine activation of mTORC1 involved in muscle regeneration in dystrophic mice [48, 64]. Thus, the protection observed in the 1-month-old tongue of the mdx muscle, close to the 20th day degeneration peak, may be related to the high levels of glycine presented in our study. In mdx and control mice, a gradual decline in muscle mass becomes apparent around 18 months, marking the onset of sarcopenia [65]. Mouisel and colleagues (2010) [66] have reported decreased levels of recovery of maximum force after myotoxic injury and of phosphorylated Akt and S6 (i.e., participants in the mTOR signaling pathway) in the tibialis anterior of 18–24-month-old mdx mice in comparison to 5-month-old mdx mice [64]. The decrease in regeneration of skeletal muscles from 18 months of age would explain the lower glycine levels in the tongue of old mdx found in this study, since a specific glycine activation of mTORC1 pathway is linked to muscle regeneration in dystrophic mice.
Glutamic acid, also known as glutamate (the anion), is a nonessential amino acid, one of the 20 proteinogenic amino acids and the most abundant rapid excitatory neurotransmitter in the nervous system [30]. Glutathione is the main cellular antioxidant and regulates free radical homeostasis. Glutamate plays an essential regulatory role in the synthesis of glutathione and is also related to its functions (for review, see [67, 68]). It is proven the relationship between decreased intracellular glutathione and stress states, such as chronic diseases [68], besides demonstrating benefits in the use of different antioxidant drugs for preclinical studies in mdx mice, such as improvement in dystrophinopathy with decreased necrosis [69]. In a metabolomic study of the biceps femoris muscle of the canine model for DMD (golden retriever muscular dystrophy), there was an increase in the concentration of glutamic acid when compared to the control [70]. Laferte, Rosenkrantz, and Berlinguet (1963) [71] observed an increase in weight loss and acceleration of the beginning of the terminal phase of the disease in mice that received exogenous glutamic acid. The increased glutamate concentration in the dystrophic muscles of the quadriceps and diaphragm suggests its connection with muscle regeneration and its requirement of protein replacement [13] and its relationship with the pathology of DMD. In this study, the amounts of glutamic acid in the tongue mdx muscles were equivalent to those of control muscles of the same age, indicating protection against myonecrosis on the tongue. As previously mentioned, skeletal muscle of older mdx mice (18-24-month-old) present sarcopenia with reduced regeneration, including reduced mTORC1 activation for muscle protein synthesis [66]. Thus, lower glutamate levels in old versus young mdx mice are expected. The observed decrease of glycine and glutamate levels suggests that mTOR-related pathways may not contribute to the protective mechanisms against the dystropathology seen in the old mdx tongue.
These results corroborate the protection observed in histology and the use of these as muscle biomarkers of degeneration and regeneration. Other studies on affected muscles can elucidate their specificities as biomarkers for the progression of DMD, without the interference of aging.
Conclusions
The tongue of young (1 month) and old (21 to 25 months) mdx animals remains spared from the intense and progressive myonecrosis described in other muscles, as evidenced by mid-belly histological analysis. This protection was verified by the biomarker proteins for inflammation and fibrosis and by the concentrations of the metabolites 3-methylhistidine, acetic acid, glutamic acid, alanine, creatine, phosphocreatine, glycerol, glycine, isoleucine, methionine, succinic acid, and taurine already used for dystrophic muscle differentiation and in this work presented similar results between wild-type and mdx whole tongue muscles.
Among the possible biomarkers analyzed, the metabolites alanine, methionine, 3-methylhistidine, taurine, and glycerol may be efficient for specific assessments, but care should be taken when using them to monitor the progression of the disease, since they change throughout life.
Muscle degeneration biomarkers, which in other studies proved to be efficient (acetic acid, phosphocreatine, isoleucine, succinate, creatine, TNF-α, and TGF-β), did not show any difference with aging and in the spared muscles. Other studies on affected muscles can elucidate their specificities as biomarkers for the progression of DMD, without the interference of aging.
Availability of data and materials
Not applicable.
Abbreviations
- 3-MeH:
-
3-Methylhistidine
- ANOVA:
-
Analysis of variance
- CEUA:
-
Brazilian College for Animal Experimentation
- CN:
-
Central nuclei
- D2O:
-
Deuterated water
- DMD:
-
Duchenne muscular dystrophy
- FIB:
-
Areas of fibrosis
- GAPDH:
-
Glyceraldehyde 3-phosphate dehydrogenase protein
- mdx :
-
X chromosome-linked muscular dystrophy
- mTORC1:
-
Mammalian target of rapamycin complex 1
- NMR:
-
Nuclear magnetic resonance
- PN:
-
Areas of muscle fibers with peripheral nuclei
- iPCA:
-
Interactive principal component analysis
- TNF-α:
-
Tumor necrosis factor alpha
- TGF-β:
-
Transforming growth factor beta
- TSP:
-
Trimethylsilyl tetradeuteropropionic acid
- UNESP:
-
São Paulo State University
References
Mendell JR, Shilling C, Leslie ND, Flanigan KM, al-Dahhak R, Gastier-Foster J, et al. Evidence-based path to newborn screening for Duchenne muscular dystrophy. Ann Neurol 2012;71(3):304–13. https://doi.org/10.1002/ana.23528
Hoffman EP, Brown RH, Kunkel LM. Dystrophin: the protein product of the duchenne muscular dystrophy locus. Cell. 1987;51(6):919–28. https://doi.org/10.1016/0092-8674(87)90579-4.
Tuffery-Giraud S, Béroud C, Leturcq F, Yaou RB, Hamroun D, Michel-Calemard L, et al. Genotype-phenotype analysis in 2,405 patients with a dystrophinopathy using the UMD-DMD database: a model of nationwide knowledgebase. Hum Mutat. 2009;30(6):934–45. https://doi.org/10.1002/humu.20976.
Tinsley JM, Blake DJ, Zuellig RA, Davies KE. Increasing complexity of the dystrophin-associated protein complex. Proc Natl Acad Sci. 1994;91(18):8307–13. https://doi.org/10.1073/pnas.91.18.8307.
Bogdanovich S, Perkins KJ, Krag TOB, Khurana TS. Therapeutics for Duchenne muscular dystrophy: current approaches and future directions. J Mol Med. 2004;82(2):102–15. https://doi.org/10.1007/s00109-003-0484-1.
Landfeldt E, Thompson R, Sejersen T, McMillan HJ, Kirschner J, Lochmüller H. Life expectancy at birth in Duchenne muscular dystrophy: a systematic review and meta-analysis. Eur J Epidemiol. 2020;35(7):643–53. https://doi.org/10.1007/s00109-003-0484-1.
Bulfield G, Siller WG, Wight PA, Moore KJ. X chromosome-linked muscular dystrophy (mdx) in the mouse. Proc Natl Acad Sci. 1984;81(4):1189–92. https://doi.org/10.1073/pnas.81.4.1189.
McGreevy JW, Hakim CH, McIntosh MA, Duan D. Animal models of Duchenne muscular dystrophy: from basic mechanisms to gene therapy. Dis Model Mech. 2015;8(3):195–213. https://doi.org/10.1242/dmm.018424.
Coulton GR, Morgan JE, Partridge TA, Sloper JC. The mdx mouse skeletal muscle myopathy: I. A histological, morphometric and biochemical investigation. Neuropathol Appl Neurobiol. 1988;14(1):53–70. https://doi.org/10.1111/j.1365-2990.1988.tb00866.x
Chamberlain JS, Metzger J, Reyes M, Townsend D, Faulkner JA. Dystrophin-deficient mdx mice display a reduced life span and are susceptible to spontaneous rhabdomyosarcoma. FASEB J. 2007;21(9):2195–204. https://doi.org/10.1096/fj.06-7353com.
Donald S, Chen R, Snyder M. Systems biology approaches to disease marker discovery. Disease markers. 2013;28(4). https://doi.org/10.3233/DMA-2010–0707
Gulston MK, Rubtsov DV, Atherton HJ, Clarke K, Davies KE, Lilley KS, et al. A combined metabolomic and proteomic investigation of the effects of a failure to express dystrophin in the mouse heart. J Proteome Res. 2008;7(5):2069–77. https://doi.org/10.1021/pr800070p.
Martins-Bach AB, Bloise AC, Vainzof M, Rahnamaye RS. Metabolic profile of dystrophic mdx mouse muscles analyzed with in vitro magnetic resonance spectroscopy (MRS). Magn Reson Imaging. 2012;30(8):1167–76. https://doi.org/10.1016/j.mri.2012.04.003.
Maurício AF, de Carvalho SC, Santo Neto H, Marques MJ. Effects of dietary omega-3 on dystrophic cardiac and diaphragm muscles as evaluated by 1H magnetic resonance spectroscopy: metabolic profile and calcium-related proteins. Clinical Nutrition ESPEN. 2017;20:60–7. https://doi.org/10.1016/j.clnesp.2017.03.005.
Archer SK, Garrod R, Hart N, Miller S. Dysphagia in Duchenne muscular dystrophy assessed by validated questionnaire. Int J Lang Commun Disord. 2013;48(2):240–6. https://doi.org/10.1111/j.1460-6984.2012.00197.x.
van den Engel-Hoek L, de Groot IJM, Sie LT, van Bruggen HW, de Groot SAF, Erasmus CE, et al. Dystrophic changes in masticatory muscles related chewing problems and malocclusions in Duchenne muscular dystrophy. Neuromuscul Disord. 2016;26(6):354–60. https://doi.org/10.1016/j.nmd.2016.03.008.
Spassov A, Gredes T, Gedrange T, Lucke S, Pavlovic D, Kunert-Keil C. Histological changes in masticatory muscles of mdx mice. Arch Oral Biol. 2010;55(4):318–24. https://doi.org/10.1016/j.archoralbio.2010.02.005.
Lefaucheur JP, Pastoret C, Sebille A. Phenotype of dystrophinopathy in old mdx mice. Anat Rec. 1995;242(1):70–6.
Al-KhaliliSzigyarto C, Spitali P. Biomarkers of Duchenne muscular dystrophy: current findings. Degener Neurol Neuromuscul Dis. 2018;8:1–13. https://doi.org/10.2147/DNND.S121099.
Griffin JL, Williams HJ, Sang E, Clarke K, Rae C, Nicholson JK. Metabolic profiling of genetic disorders: a multitissue 1H nuclear magnetic resonance spectroscopic and pattern recognition study into dystrophic tissue. Anal Biochem. 2001;293(1):16–21. https://doi.org/10.1006/abio.2001.5096.
Terrill JR, Grounds MD, Arthur PG. Taurine deficiency, synthesis and transport in the mdx mouse model for Duchenne muscular dystrophy. Int J Biochem Cell Biol. 2015;66:141–8. https://doi.org/10.1016/j.biocel.2015.07.016.
Grounds MD, Terrill JR, Al-Mshhdani BA, Duong MN, Radley-Crabb HG, Arthur PG. Biomarkers for Duchenne muscular dystrophy: myonecrosis, inflammation and oxidative stress. Dis Models Mechan. 2020;13(2). https://doi.org/10.1242/dmm.043638
Maranhão J, de Oliveira MD, Maurício AF, de Carvalho SC, Ferretti R, Pereira JA, et al. Changes in calsequestrin, TNF-α, TGF-β and MyoD levels during the progression of skeletal muscle dystrophy in mdx mice: a comparative analysis of the quadriceps, diaphragm and intrinsic laryngeal muscles. Int J Exp Pathol. 2015;96(5):285–93. https://doi.org/10.1111/iep.12142.
Marques MJ, Ferretti R, Vomero VU, Minatel E, Neto HS. Intrinsic laryngeal muscles are spared from myonecrosis in the mdx mouse model of Duchenne muscular dystrophy. Muscle Nerve. 2007;35(3):349–53. https://doi.org/10.1002/mus.20697.
Messina S, Vita GL, Aguennouz M, Sframeli M, Romeo S, Rodolico C, et al. Activation of NF-kappaB pathway in Duchenne muscular dystrophy: relation to age. Acta Myologica. 2011;30(1):16–23.
Bernasconi P, Torchiana E, Confalonieri P, Brugnoni R, Barresi R, Mora M, et al. Expression of transforming growth factor-beta 1 in dystrophic patient muscles correlates with fibrosis. Pathogenetic role of a fibrogenic cytokine. J Clin Invest. 1995;96(2):1137–44. https://doi.org/10.1172/JCI118101
Carvalho SC, Matsumura CY, Santo Neto H, Marques MJ. Identification of plasma interleukins as biomarkers for deflazacort and omega-3 based Duchenne muscular dystrophy therapy. Cytokine. 2018;102:55–61. https://doi.org/10.1016/j.cyto.2017.12.006.
Ferretti R., Marques M. J., Pertille A., & Santo Neto H. (2009). Sarcoplasmic-endoplasmic-reticulum Ca2+-ATPase and calsequestrin are overexpressed in spared intrinsic laryngeal muscles of dystrophin-deficient mdx mice. Muscle Nerve. 2019;39(5):609–615. https://doi.org/10.1002/mus.21154
Sukumaran DK, Garcia E, Hua J, Tabaczynski W, Odunsi K, Andrews C, et al. Standard operating procedure for metabonomics studies of blood serum and plasma samples using a 1H-NMR micro-flow probe. Magn Reson Chem. 2009;47(S1):S81–5. https://doi.org/10.1002/mrc.2469.
Wishart D. S., Guo A., Oler E., Wang F., Anjum A., Peters H., Dizon R., Sayeeda Z., Tian S., Lee B. L., Berjanskii M., Mah R., Yamamoto M., Jovel J., Torres-Calzada C., Hiebert-Giesbrecht M., Lui V. W., Varshavi D., Varshavi D., Allen D., … Gautam V. (2022). HMDB 5.0: the Human Metabolome Database for 2022. Nucl Acids Res. 50(D1):D622–D631. https://doi.org/10.1093/nar/gkab1062
Weljie AM, Newton J, Mercier P, Carlson E, Slupsky CM. Targeted profiling: quantitative analysis of 1H NMR metabolomics data. Anal Chem. 2006;78(13):4430–42. https://doi.org/10.1021/ac060209g.
Beckonert O, Keun HC, Ebbels TMD, Bundy J, Holmes E, Lindon JC, et al. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nature Protocol. 2007;2(11):2692–703. https://doi.org/10.1038/nprot.2007.376.
McClung JR, Goldberg SJ. Functional anatomy of the hypoglossal innervated muscles of the rat tongue: a model for elongation and protrusion of the mammalian tongue. Anat Rec. 2000;260:378–86.
Cullins MJ, Connor NP. Alterations of intrinsic tongue muscle properties with aging. Muscle Nerve. 2017;56:E119–25. https://doi.org/10.1002/mus.25605.
Doyle ME, Premathilake HU, Yao Q, Mazucanti CH, Egan JM. Physiology of the tongue with emphasis on taste transduction. Physiol Rev. 2023;103(2):1193–246.
Kunert-Keil CH, Gredes T, Lucke S, Botzenhart U, Dominiak M Gedrange T. Differential expression of genes involved in the calcium homeostasis in masticatory muscles of MDX mice. Journal of physiology and pharmacology : an official journal of the Polish Physiological Society. 2014;65(2).
Yamanouchi K, Tanaka Y, Ikeda M, Kato S, Okino R, Nishi H, Hakuno F, Takahashi SI, Chambers J, Matsuwaki T, Uchida K. Macroglossia and less advanced dystrophic change in the tongue muscle of the Duchenne muscular dystrophy rat. Skelet Muscle. 2022;12(1):24. https://doi.org/10.1186/s13395-022-00307-7.
Tinsley JM, Potter AC, Phelps SR, Fisher R, Trickett JI, Davies KE. Amelioration of the dystrophic phenotype of mdx mice using a truncated utrophin transgene. Nature. 1996;384:349–53.
Deconinck N, Tinsley J, De Backer F, Fisher R, Kahn D, Phelps S, Davies K, Gillis JM. Expression of truncated utrophin leads to major functional improvements in dystrophin-deficient muscles of mice. Nat Med. 1997;3:16–21.
Tinsley JM, Deconinck N, Fisher R, Kahn D, Phelps S, Gillis JM, Davies K. Expression of full-length utrophin prevents muscular dystrophy in mdx mice. Nat Med. 1998;4:1441–4.
Porter JD, Merriam AP, Khanna S, Andrade FH, Richmonds CR, Leahy P, Cheng G, Karathanasis P, Zhou X, Kusner LL, Adams ME, Willem M, Mayer U, Kaminski HJ. Constitutive properties, not molecular adaptations, mediate extraocular muscle sparing in dystrophic mdx mice. FASEB J. 2003;17(8):893–5. https://doi.org/10.1096/fj.02-0810fje. (Epub 2003 Mar 28).
McDonald AA, Hebert SL, Kunz MD, Ralles SJ, McLoon LK. Disease course in mdx:utrophin+/- mice: comparison of three mouse models of Duchenne muscular dystrophy. Physiol Rep. 2015;3(4):e12391. https://doi.org/10.14814/phy2.12391.
Ferretti R, Pertille A, Neto HS, Marques MJ. Age-related changes in dystrophin–glycoprotein complex and in utrophin are not correlated with intrinsic laryngeal muscles protection in mdx mice. Muscle Nerve. 2011;44:978–80. https://doi.org/10.1002/mus.22235.
Sobolev AP, Mannina L, Costanzo M, Cisterna B, Malatesta M, Zancanaro C. Age-related changes in skeletal muscle composition: a pilot nuclear magnetic resonance spectroscopy study in mice. Exp Gerontol. 2017;92:23–7. https://doi.org/10.1016/j.exger.2017.03.005.
McIntosh LM, Garrett KL, Megeney L, Rudnicki MA, Anderson JE. Regeneration and myogenic cell proliferation correlate with taurine levels in dystrophin- and MyoD-deficient muscles. Anat Rec. 1998;252(2):311–24.
Barbiera A, Sorrentino S, Lepore E, Carfì A, Sica G, Dobrowolny G, Scicchitano BM. Taurine attenuates catabolic processesrelated to the onset of sarcopenia. Int J Mol Sci. 2020;21:8865.
Singh P, Gollapalli K, Mangiola S, Schranner D, Yusuf MA, Chamoli M, Shi SL, Lopes Bastos B, Nair T, Riermeier A, Vayndorf EM, Wu JZ, Nilakhe A, Nguyen CQ, Muir M, Kiflezghi MG, Foulger A, Junker A, Devine J, Sharan K, Chinta SJ, Rajput S, Rane A, Baumert P, Schönfelder M, Iavarone F, di Lorenzo G, Kumari S, Gupta A, Sarkar R, Khyriem C, Chawla AS, Sharma A, Sarper N, Chattopadhyay N, Biswal BK, Settembre C, Nagarajan P, Targoff KL, Picard M, Gupta S, Velagapudi V, Papenfuss AT, Kaya A, Ferreira MG, Kennedy BK, Andersen JK, Lithgow GJ, Ali AM, Mukhopadhyay A, Palotie A, Kastenmüller G, Kaeberlein M, Wackerhage H, Pal B, Yadav VK. Taurine deficiency as a driver of aging. Science. 2023;380(6649):abn9257. https://doi.org/10.1126/science.abn9257.
De Paepe B. What nutraceuticals can do for Duchenne muscular dystrophy: lessons learned from amino acid supplementation in mouse models. Biomedicines. 2023;11(7):2033. https://doi.org/10.3390/biomedicines11072033.
Nishio H, Wada H, Matsuo T, Horikawa H, Takahashi K, Nakajima T, Matsuo M, Nakamura H. Glucose, free fatty acid and ketone body metabolism in Duchenne muscular dystrophy. Brain Develop. 1990;12(4):390–402. https://doi.org/10.1016/s0387-7604(12)80071-4.
Srivastava NK, Yadav R, Mukherjee S, Sinha N. Perturbation of muscle metabolism in patients with muscular dystrophy in early or acute phase of disease: in vitro, high resolution NMR spectroscopy based analysis. Clin Chim Acta. 2018;478:171–81. https://doi.org/10.1016/j.cca.2017.12.036.
Radley-Crabb HG, Marini JC, Sosa HA, Castillo LI, Grounds MD, Fiorotto ML. Dystropathology increases energy expenditure and protein turnover in the Mdx mouse model of Duchenne muscular dystrophy. Fraidenraich D, editor. PLoS ONE. 2014;9(2):e89277. https://doi.org/10.1371/journal.pone.0089277 .
Strakova J, Kamdar F, Kulhanek D, Razzoli M, Garry DJ, Ervasti JM, Bartolomucci A, Townsend D. Integrative effects of dystrophin loss on metabolic function of the mdx mouse. Sci Rep. 2018;8(1). https://doi.org/10.1038/s41598-018-31753-3.
Schäfer R, Zweyer M, Knauf U, Mundegar RR, Wernig A. The ontogeny of soleus muscles in mdx and wild type mice. Neuromuscul Disord. 2005;15(1):57–64. https://doi.org/10.1016/j.nmd.2004.09.011.
Martínez Y, Li X, Liu G, Bin P, Yan W, Más D, Valdivié M, Hu CA, Ren W, Yin Y. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids. 2017;49:2091–8. https://doi.org/10.1007/s00726-017-2494-2.
Williams, AP. Amino acids | Determination. Encyclopedia Of Food Sciences And Nutrition , [s.l.], p.192–197, 2003. Elsevier.
Keller U. Nutritional laboratory markers in malnutrition. J Clin Med. 2019;8(6):775. https://doi.org/10.3390/jcm8060775.
Janssen I, Heymsfield SB, Wang Z, Ross R. Skeletal muscle mass and distribution in 468 men and women aged 18–88 yr. J Appl Physiol. 2000;89(1):81–8. https://doi.org/10.1152/jappl.2000.89.1.81.
Nilwik R, Snijders T, Leenders M, Groen BB, van Kranenburg J, Verdijk LB, van Loon LJ. The decline in skeletal muscle mass with aging is mainly attributed to a reduction in type II muscle fiber size. Exp Gerontol. 2013;48(5):492–8. https://doi.org/10.1016/j.exger.2013.02.012.
Kumar V, Abbas A, Fausto N. Robbins e Cotran Pathologic basis of disease. 8th ed. Rio de Janeiro: Elsevier; 2010.
Collins RA, Grounds MD. The role of tumor necrosis factor-alpha (TNF-alpha) in skeletal muscle regeneration. Studies in TNF-alpha(-/-) and TNF-alpha(-/-)/LT-alpha(/) mice. J Histochem Cytochem. 2001;49(8):989–1001. https://doi.org/10.1177/002215540104900807.
Ham DJ, Gardner A, Kennedy TL, Trieu J, Naim T, Chee A, Alves FM, Caldow MK, Lynch GS, Koopman R. Glycine administration attenuates progression of dystrophic pathology in prednisolone-treated dystrophin/utrophin null mice. Sci Rep. 2019;9(1):12982. https://doi.org/10.1038/s41598-019-49140-x.
Skrabek RQ, Anderson JE. Metabolic shifts and myocyte hypertrophy in deflazacort treatment of mdx mouse cardiomyopathy. Muscle Nerve. 2016;24(2).
Caldow MK, Ham DJ, Trieu J, Chung JD, Lynch GS, Koopman R. Glycine protects muscle cells from wasting in vitro via mTORC1 signaling. Front Nutr. 2019. https://doi.org/10.3389/fnut.2019.00172.
Lin C, Han G, Ning H, Song J, Ran N, Yi X, Seow Y, Yin H. Glycine enhances satellite cell proliferation, cell transplantation, and oligonucleotide efficacy in dystrophic muscle. Mol Ther. 2020;28(5):1339–58. https://doi.org/10.1016/j.ymthe.2020.03.003.
Grounds MD, Radley HG, Lynch GS, Nagaraju K, De Luca A. Towards developing standard operating procedures for pre-clinical testing in the mdx mouse model of Duchenne muscular dystrophy. Neurobiol Dis. 2008;31(1):1–19. https://doi.org/10.1016/j.nbd.2008.03.008.
Mouisel E, Vignaud A, Hourdé C, Butler-Browne G, Ferry A. Muscle weakness and atrophy are associated with decreased regenerative capacity and changes in mTOR signaling in skeletal muscles of venerable (18–24-month-old) dystrophic mdx mice. Muscle Nerve. 2010;41(6):809–18. https://doi.org/10.1002/mus.21624.
Wu G, Fang YZ, Yang S, Lupton JR, Turner ND. Glutathione metabolism and its implications for health. J Nutr. 2004;34(3):489–92. https://doi.org/10.1093/jn/134.3.489
Rutten EP, Engelen MP, Schols AM, Deutz NE. Skeletal muscle glutamate metabolism in health and disease: state of the art. Curr Opin Clin Nutr Metab Care. 2005;8(1):41–51. https://doi.org/10.1097/00075197-200501000-00007.
Petrillo S, Pelosi L, Piemonte F, Travaglini L, Forcina L, Catteruccia M, et al. Oxidative stress in Duchenne muscular dystrophy: focus on the NRF2 redox pathway. Hum Mol Genet. 2017;26(14):2781–90. https://doi.org/10.1093/hmg/ddx173.
Abdullah M, Kornegay JN, Honcoop A, Parry TL, Balog-Alvarez CJ, O’Neal SK, Bain JR, Muehlbauer MJ, Newgard CB, Patterson C, et al. Non-targeted metabolomics analysis of golden retriever muscular dystrophy-affected muscles reveals alterations in arginine and proline metabolism, and elevations in glutamic and oleic acid in vivo. Metabolites. 2017;7(3):38. https://doi.org/10.3390/metabo7030038.
Laferté RO, Rosenkrantz H, Berlinguet L. Transamination in muscular dystrophy and the effect of exogenous glutamate: a study on vitamin E deficient rabbits, and mice with hereditary dystrophy. Can J Biochem Physiol. 1963;41(1):1423–32. https://doi.org/10.1139/y63-162.
Acknowledgements
Not applicable.
Funding
This work was supported by São Paulo Research Foundation (FAPESP) grant no. 2018/25598–0, grant no. 2019/20162–1, and grant no. 2013/00312–2 (to MSVL and CYM). CYM was supported by CNPq grant no. 456357/2014–9. JSC is supported by NIH grant no. R01AR040864. GLO is supported by DOD grant award no. W81XWH-18–1-0624.
Author information
Authors and Affiliations
Contributions
MSVL, JSC and CYM conceived and designed experiments; MSVL, EKS, NGGA and CYM performed experiments; MSVL, RF, NGGA, GLO, JSC and CYM analyzed the data; MSVL, EKS, GLO, JSC and CYM interpreted the results of the experiments and wrote the manuscript with editing from MSVL. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All experiments were carried out in accordance with the guidelines for animal experimentation of the Ethics Committee on Animal Use — CEUA-IBB/UNESP, protocol no. 1095.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lorena, M., Santos, E.K., Ferretti, R. et al. Biomarkers for Duchenne muscular dystrophy progression: impact of age in the mdx tongue spared muscle. Skeletal Muscle 13, 16 (2023). https://doi.org/10.1186/s13395-023-00325-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13395-023-00325-z