
Abstract
Twelve solitary and platy, colonial coral taxa assigned to the families Microsolenidae, Misistellidae, Montlivaltiidae, Rayasmiliidae, and Thamnasteriidae are described and illustrated from the Callovian (Ifenthal Formation, Herznach Member) of the area of Andil near Liesberg, about 20 km WSW of the town of Basel, Switzerland. The platy growth forms and the presence of five species of the superfamily Cyclolitoidea suggests that these corals lived under low-level light conditions.
Similar content being viewed by others

Introduction
The Mesozoic rocks forming the Jura Mountains in NW Switzerland were deposited in an epicontinental setting. The Jurassic rocks accumulated under shallow-marine conditions and consist principally of limestone, marl, and mudrock. Due to differential subsidence and relative changes in sea level, the domain of the Swiss Jura Mountains became morphologically diversified and, accordingly, so did the depositional regime and related faunal associations. Partly, these deposits are very rich in fossils and their intensive investigation began in the nineteenth century through, for instance, Agassiz (1839, 1840) and Koby (1880–1889).
During the Middle and Late Jurassic, carbonate platforms developed in the domain of the Swiss Jura Mountains, housing local coral patches or even chains of patch reefs. For the Middle Jurassic (Bajocian), shallow and isolated coral meadows were reported, such as, for instance, from the Gisli-Fluh (Gonzalez, 1993; Gonzalez & Wetzel, 1996). Very prominent coral occurrences are described from the Late Jurassic (Oxfordian; Gygi, 2000; Gygi & Persoz, 1986). Starting with the coral meadows during the early Middle Oxfordian (Liesberg Member of the Bärschwil Fm.), locally, reefs developed further up during the Middle Oxfordian St. Ursanne Formation. These corals were taxonomically investigated, first by Thurmann and Etallon (1864), then by Koby (1880–1889), and later by Beauvais (1963), and Birenheide (1969). The material studied by these authors originated mainly from the Middle Oxfordian in the areas of Combe Chavatte, La Caquerelle, Saint-Ursanne, among others. In contrast, Middle Jurassic corals, late Bajocian to early Bathonian in age, from northern Switzerland, have only be reported (Gonzalez & Wetzel, 1996), but are not yet taxonomically dealt with in detail, except for the studies of Meyer (1888), Koechlin (1933, 1950), Beauvais (1966), and Kutz et al. (2020). The present study addresses a small coral fauna from the Callovian of Liesberg (Kanton Basellandschaft; Fig. 1) that has been reported for the first time by Gründel and Hostettler (2012).

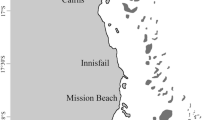
Location map, study area. Outcrop area marked by red asterisk
Geological setting
In NW Switzerland, the deposits of the Swiss Jura Mountains are built-up by Mesozoic rocks deposited in the southern part of the Mesozoic Central European Basin System, which was mainly covered by an intracontinental epeiric sea that became connected to the Tethys in the South during the Middle Jurassic (e.g., Maystrenko et al., 2008; Ziegler, 1990). During the Middle and Late Jurassic, the rocks forming the Swiss Jura Mountains record the changing morphology of the depositional realm (Allenbach & Wetzel, 2006; Burkhalter, 1996; Pieñkowski et al., 2008; Wetzel & Allia, 2000) that appears to have been considerably affected by pre-existing tectonic structures in the basement, which became temporarily re-activated (e.g., Wetzel et al., 2003).
The basement in central Europe was tectonically structured towards the end of the Variscan orogeny during the late Palaeozoic, when numerous basins, grabens, half-grabens, and fault systems formed (Arthaud & Matte, 1977; Boigk & Schöneich, 1974; Ménard & Molnar, 1988; von Raumer, 1998). In the study area and its surroundings, such tectonic structures in the basement were identified on seismic records (Madritsch et al., 2018; McCann et al., 2006; NAGRA, 2008). During the Triassic, peneplanation took place, and continental and restricted marine deposits accumulated, including up to 100-m-thick evaporites. At the beginning of the Jurassic, a major transgression occurred, and mainly fully marine rocks accumulated since then (Fig. 2).

Schematic stratigraphic column of the rocks in northern Switzerland
When the Tethys Ocean to the south and the North Atlantic Ocean to the west opened during the Jurassic, the area in between was affected by extensional and transtensional stresses (e.g., Færseth, 1996; Philippe et al., 1996). Even short-term changes in the stress field might have occurred (Robin et al., 1998; Wetzel et al., 2003). Pre-existing basement structures became temporarily reactivated, as evidenced by vein mineralisation data, changes in lithofacies and rock thickness across and adjacent to basement faults, and in particular, thickness anomalies exceeding depositional water depth, which imply synsedimentary differential tectonic movements (e.g., Wetzel et al., 2003; Timar-Geng et al., 2006; Brockamp et al., 2011; Reisdorf & Wetzel, 2018). Vertical movements in the basement, however, were dissipated by Triassic salt and, thus, the deposits above deformed mainly flexurally, and the basin floor was morphologically differentiated into gentle swells and depressions (e.g., Wetzel & Meyer, 2006; Wetzel et al., 2003). Differential subsidence has been deciphered, especially for the Middle and Upper Jurassic, including the up to 100-m-thick oolites and shallow-water carbonates of the Hauptrogenstein Formation (Gonzalez & Wetzel, 1996), the Passwang Formation below (Burkhalter, 1996), and the Callovian Ifenthal Formation above (Bitterli-Dreher, 2012), as well as Oxfordian formations (e.g., Allenbach, 2001; Wetzel et al., 2003; Fig. 2). Shallow-water sedimentation continued until the Upper Jurassic–Lower Cretaceous, constituting a rock pile less than 1 km thick (NAGRA, 2002).
The investigated material was collected from Callovian deposits that are exposed in the now abandoned ‘Andil’ clay pit near Liesberg, which is about 20 km WSW of the town of Basel (Swiss coordinates 2′598′686/1′250′087, Fig. 1). The corals were found in the Herznach Member of the Ifenthal Formation (Figs. 2, 3). In northern Switzerland, the Callovian represented a period of slow, but nonetheless differential subsidence, structuring the seafloor into gentle depressions and swells, the latter being temporarily exposed to wave influence, resulting in rock reworking, omission, and/or condensation (e.g., Bitterli-Dreher, 2012). Hence, the approximately 1-m-thick Herznach Member encompasses iron-oolithic marly limestone and marls that accumulated during a relative sea-level rise. Stratigraphically, these deposits belong to the Calloviense to Lamberti Zones (Hostettler, 2014; Fig. 3). The stratigraphically lowest iron-oolithic bed was named ‘iron-oolithic lumachelle bank’ by Stäuble (1959), because of its richness in bivalve shells. It was included into the Herznach Member by Hostettler (2014). According to ammonite finds, it belongs to the Enodatum Subzone (Calloviense Zone), and it also contains the corals described here. In addition to the outcrop locality Andil, the Herznach Member is locally exposed from the Col de la Croix in the north to the Grenchenberg in the south, and from Liesberg village in the east to the marginal area of the Franches-Montagnes in the west (Fig. 1). The fossil-bearing Callovian deposits are only exposed in river beds, clay pits, or during temporal excavations.

Detailed section of the outcrop studied
The corals were collected from only one interval consisting of an iron-ooid-bearing, clayey limestone, which was deposited on top of a well-developed hardground in the lowermost part of the Herznach Member (see Fig. 3). The coral-bearing interval contains numerous shells of various bivalves; most common are specimens of the genera Plagiostoma, Ctenostreon, Chlamys, Trigonia, Orthotrigonia, Protocardia, Pseudotrapezium, and non-identified members of the family Arcidae. Fragments of tree-shaped bryozoan colonies and crinoids of the genus Millericrinus are also common. Ammonites are quite rare. The fauna is allochthonous and the corals are quite randomly dispersed in the rocks, and do not occur directly on the hardground, except for one juvenile single coral that was found attached to the hardground. Occasionally, the corals grew on thick-shelled bivalves. The coral colonies are generally flat, do not exceed ten centimetres in their largest lateral dimension, and are not higher than two centimetres. They are bored and, partly only preserved as fragments. The surrounding rock could not be completely removed from the corals and, therefore, the surface of the corals is not observable.
Methods
Coral specimens were collected from one stratigraphic horizon. After careful cleaning, they were cut and polished. Thin sections in both transversal and longitudinal orientations were prepared, whenever possible. Thin sections were scanned in plane light by using a flatbed scanner with an optical resolution of 6400 dpi. Scanned images were then transferred to greyscale bitmaps. Their quality was amended by histogram contrast manipulation (contrast stretching), whenever possible.
To gain more insights into the intraspecific variation of these corals, and to obtain a better strategy for comparing species, the corallite dimensions, and number of septa or septal density were systematically measured using the scanned thin sections. To achieve statistical significance, the largest possible number of measurements was taken. This number was mainly determined by the size and quality of the thin sections, and the size of the single corallites in relation to the size of the thin sections.
For each type of measurement (such as the corallite diameter or number of septa), the following values were obtained:
n number of measurements.
min–max lowest and highest measured values.
µ arithmetic mean (average).
s standard deviation.
cv coefficient of variation according to K. Pearson.
µ ± s first interval.
Thin sections were measured and values were calculated using the Palaeontological Database System PaleoTax, module PaleoTax/Measure (https://www.paleotax.de/measure). Previous studies (for instance Löser, 2012, 2020) have shown that the corallite dimensions (septal counts, or septal density) have a certain range throughout the whole coral colony. The comparison of morphometric data may serve as a method to distinguish fossil coral species.
The morphometric data obtained from the corals of the study area were compared with the morphometric data of available specimens in worldwide collections of Jurassic and Cretaceous corals. The described and illustrated coral specimens are part of the Fondation paléontologique jurassienne (FPJ), Glovelier, Switzerland, and they are kept at the Natural History Museum Bern, Switzerland.
The distribution data (as reflected in the synonymy lists) are almost entirely based on examined material. Material that is only mentioned in the literature and material that is not available or is insufficiently described and illustrated in the literature were not taken into account. To obtain better insight into the distribution patterns of the present coral fauna, additional unpublished material has been included. This material is indicated by a collection acronym and sample number under ‘Other material’.
Systematic description
Order Scleractinia Bourne, 1900
The classification of the order Scleractinia follows the studies by Vaughan and Wells (1943), Wells (1956), and Alloiteau (1952, 1957) with the difference that, instead of suborders, superfamilies are applied. More explication and comparison of both classification systems are found in Löser (2016) and Löser et al. (2018).
The abbreviations used in the synonymy lists follow Matthews (1973): *, earliest valid publication of the species name; v, the specimen was observed by the first author. The following abbreviations are used to describe the dimensions of the corals: c, corallite diameter (outer diameter); ccd, distance between corallite centres; cdw, distance between corallite centres within series; cl, corallite diameter (lumen, calicular pit); cmax, large corallite diameter; cmin, small corallite diameter; crd, distance of corallite series; septa, number of septa in the adult corallite; sk, number of septa, which reach the columella. Collection abbreviations are as per the following: BSPG, Bayerische Staatssammlung für Paläontologie und Geologie (München, Germany); BUFGG, Bukarest University, Faculty of Geography and Geology (Bucuresti, Romania); CAMSM, The Sedgwick Museum of Earth Sciences (Cambridge, UK); CGS, Ceská geologická sluzba (Praha, Czech Republic); ERNO, Universidad Nacional Autónoma de México, Instituto de Geología, Estación Regional del Noroeste (Hermosillo, Mexico); FLH, Coll. Fritz Lang (Hirschaid, Germany); GZG, Geowissenschaftliches Zentrum Göttingen (Göttingen, Germany); MJSN, Musée jurassien des Sciences naturelles (Porrentruy, Switzerland); MNHN, Muséum National d'Histoire Naturelle (Paris, France); NMNH; National Museum of Natural History (Washington D.C., USA); RUB, Ruhr-Universität, Geologisches Institut (Bochum, Germany); TUM, The Tohoku University Museum (Sendai, Japan);. UP, Université de Provence (Marseille, France); UPV, Universidad del País Vasco (Bilbao, Spain); ZPAL, Polish Academy of Sciences, Institute of Paleobiology (Warszawa, Poland).
Superfamily Cyclolitoidea Milne Edwards & Haime, 1849
Family Microsolenidae Koby, 1889
Dimorpharaea Fromentel, 1861
Type species. Microsolena koechlini Milne Edwards, 1860
Remarks. The type material of the type species is not available. According to the author of the genus, Fromentel (1861), the genus has the same structure as Microsolena Lamouroux, 1821 but differs by the presence of a central corallite and subsequent corallites that are arranged in rows. Pandey et al. (1999) contribute to the knowledge of the genus.
Dimorpharaea fungiformis (Milne Edwards & Haime, 1851)
Fig. 4 1–3

1–3 Dimorpharaea fungiformis (Milne Edwards & Haime, 1851), FPJ 452. 1 transversal thin section. 2 transversal thin section, detail. 3 longitudinal thin section. 4–6 Dimorpharaea ornata (Koby, 1887), FPJ 375. 4 transversal thin section. 5 transversal thin section, detail. 6 longitudinal thin section. 7–9 Dimorpharaea cf. tosaensis Eguchi, 1951, FPJ 377. 7 transversal thin section 8 transversal thin section, detail. 9 longitudinal thin section. Scale bar 1 mm
*v 1851Thamnastrea fungiformis: Milne Edwards & Haime, p. 141, Pl. 30, Fig. 4
v 1970Dimorpharaea fungiformis (ME & H): Beauvais, p. 68, Pl. A, Fig. 1, Pl. E, Fig. 5
v 2003Dimorpharaea koechlini (Milne Edwards & Haime, 1860): Baron-Szabo & González León, p. 217, Fig. 9G
v 2003Synastrea bellula d'Orbigny, 1850: Baron-Szabo & González León, p. 220, Fig. 9I
v 2013Dimorpharaea manchacanensis Wells, 1933: Löser, p. 28, Figs. 10a–c
Material FPJ 452; 2 thin sections.
Dimensions (FPJ 452).
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 5 | 4.56–6.53 | 5.64 | 0.82 | 14.6 | 4.82–6.47 |
cdw | 12 | 2.78–3.61 | 3.26 | 0.27 | 8.3 | 2.99–3.54 |
septa | 15 | 23–36 | 28.9 | 3.86 | 13.3 | 25–33 |
Description Thamnasterioid colony with corallites arranged in rows. The rows are irregular. Septa regularly perforated. Microstructure of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. No septal symmetry and no septal generations. Septa not connected to each other. Septal lateral face with pennulae. Pali absent. Costae present, confluent. Synapticulae fairly common, mainly in the space between the corallites. Columella substyliform and clearly recognisable. Endotheca absent. Budding intracalicinal, polystomodeal and complete.
Remarks In its corallite dimensions (distance between corallite rows) and septal counts the present specimen compares well to the (only illustrated) syntype of Thamnastrea fungiformis Milne Edwards & Haime, 1851 (CAMSM J6115).
Other material BSPG 2003 XX 5492 (lower Aptian of Greece, Viotía, Arachova), BSPG 2003 XX 6343 (lower upper Albian of Spain, Cataluña, Montech de Rubies), RUB P17 (Lower Kimmerdigian of Spain, Aragón, Pozuel del Campo), UP 5784 (lowermost Hauterivian of France, Yonne, Gy-l’Evêque),
Other occurrences Aalenian to Callovian of the European Boreal (UK), Lower Kimmeridgian of the Western Tethys (Spain), Hauterivian of the European Boreal (France), Upper Barremian to Lower Aptian of the Western Atlantic (Mexico), Aptian to Middle Albian of the Central Tethys (Greece), Upper Aptian to Lower Upper Aptian of the Western Tethys (Spain), Lower Albian of the Western Atlantic (Mexico) and the Western Tethys (France).
Dimorpharaea ornata (Koby, 1887)
Fig. 4 4–6
*v 1887 Microsolena ornata: Koby, p. 399, Pl. 107, Figs. 1–2
v 1997Eocomoseris raueni Löser, 1993: Baron-Szabo, p. 84, Pl. 14, Fig. 5, Pl. 15, Figs. 1, 2, 4
v 2013Dimorpharaea sp. 1: Löser, p. 28, Figs. 8kl
Material FPJ 375; 2 thin sections.
Dimensions (FPJ 375).
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 12 | 3.69–5.71 | 4.55 | 0.67 | 14.7 | 3.88–5.22 |
cdw | 12 | 1.74–3.05 | 2.23 | 0.39 | 17.6 | 1.83–2.62 |
septa | 10 | 18–23 | 20.1 | 1.91 | 9.5 | 18–22 |
Description Thamnasterioid colony with corallites arranged in irregular rows. Septa regularly perforated. Microstructure of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. No septal symmetry or generations. Septa not connected to each other, but some may be connected to the columella. Septal lateral face with pennulae. Pali absent. Costae present, confluent. Synapticulae not very common, mainly in the space between the corallites. Columella styliform and rather large. Endotheca absent. Budding intracalicinal, polystomodeal and complete.
Remarks The present specimen compares well in the distance between corallite rows and the septal counts to the (only illustrated) syntype of Microsolena ornata Koby, 1887 (MGL GEOLREG 4044). Dimorphastrea deickei Bölsche, 1877, Thamnasteria fromenteli Tomes, 1878, and Microsolena roemeri Bölsche, 1866 have comparable dimensions but much higher septal counts.
Other material BSGP 2003 XX 5804 (Lower Aptian from Greece, Viotía, Levadia).
Other occurrences Callovian to Lower Aptian of the Central Tethys (Switzerland, Germany), Lower Aptian of the Central Tethys (Greece), Aptian to Lower Albian of the Western Atlantic (Mexico) and the Western Tethys (France).
Dimorpharaea sp.
Fig. 5 4–6

1–3 Dimorpharaea williamsonensis (Wells, 1944), FPJ 435. 1 transversal thin section. 2 transversal thin section, detail. 3 longitudinal thin section. 4–6 Dimorpharaea sp., FPJ 454. 4 transversal thin section. 5 transversal thin section, detail. 6 longitudinal thin section. 7–9 Eocomoseris sp., FPJ 374. 7 transversal thin section. 8 transversal thin section, detail. 9 longitudinal thin section. Scale bar 1 mm
Material FPJ 454; 2 thin sections.
Dimensions (FPJ 454).
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 5 | 2.58–4.10 | 3.28 | 0.54 | 16.6 | 2.74–3.83 |
septa | 6 | 13–16 | 14.2 | 0.98 | 6.9 | 13–15 |
Description Thamnasterioid colony with corallites arranged in irregular rows. Septa very low in number and regularly perforated. Microstructure of large trabeculae. The thickness of septa in cross section does not vary throughout the whole septum. No septal symmetry or generations. Septa not connected to each other, but in places connected to the columella. Septal lateral face with pennulae. Pali absent. Synapticulae not very common, mainly in the space between the corallites. Columella styliform, but very small. Endotheca with few thin tabulae. Budding intracalicinal, polystomodeal and complete.
Remarks The present specimen cannot be compared to available type data. It compares to the Early Cretaceous D. williamsonensis Wells, 1944, but still has a lower number of septa. Morphometric data for D. williamsonensis Wells, 1944, as obtained from the holotype NMNH M-547426, are as follows: crd, 3.2–3.8; septa 16–26. These values are minimum–maximum values because the thin section from the holotype is very small and yielded only a few values.
Dimorpharaea cf. tosaensis Eguchi, 1951
Fig. 4 7–9
Material FPJ 377; 2 thin sections.
Dimensions (FPJ 377).
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 5 | 7.11–11.0 | 9.11 | 1.80 | 19.8 | 7.30–10.9 |
cdw | 10 | 3.34–7.17 | 4.62 | 1.19 | 25.9 | 3.42–5.82 |
septa | 10 | 20–23 | 21.5 | 0.97 | 4.5 | 21–22 |
Description Thamnasterioid colony with corallites arranged in rows with a large distance between them. The rows are regular. Septa regularly perforated. Microstructure of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. No septal symmetry or generations. Septa not connected to each other. Septal lateral face with pennulae. Pali absent. Synapticulae uncommon. Columella substyliform and small. Endotheca consists of thin tabulae. Budding intracalicinal, polystomodeal and complete.
Remarks The holotype of Dimorpharaea tosaensis (TUM 65362) has slightly larger dimensions and a higher number of septa: crd 11–12; cdw, 6; septa, 26–30. These values are minimum–maximum values because the thin section is very small and yielded only a few values.
Other material BSPG 2003 XX 5489 (Lower Aptian from Greece, Viotía, Arachova).
Other occurrences Aptian of the Central Tethys (Greece).
Dimorpharaea williamsonensis (Wells, 1944)
Fig. 5 1–3
*v 1944Microsolena williamsonensis Wells, n.sp.: Wells, p. 100, Pl. 20, Figs. 1–4
Material FPJ 435; 2 thin sections.
Dimensions (FPJ 435).
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 8 | 3.28–4.70 | 3.98 | 0.50 | 12.6 | 3.48–4.49 |
cdw | 8 | 2.95–4.02 | 3.59 | 0.39 | 10.9 | 3.19–3.98 |
septa | 10 | 17–29 | 21.7 | 3.30 | 15.2 | 18–25 |
Description Thamnasterioid colony with corallites arranged in very irregular rows. Septa regularly perforated. Microstructure of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. No septal symmetry or septal generations. Septa not connected to each other. Septal lateral face with pennulae, inner margin smooth. Pali absent. Costae present, confluent. Synapticulae fairly common. The columella varies from absent, to substyliform or styliform and thick by septal fusion. Endotheca consists of very few thin tabulae. Budding intracalicinal, polystomodeal and complete.
Remarks Although the present specimen has slightly higher septal counts and slightly higher distances of corallite rows, it still compares well to the holotype of Microsolena williamsonensis (NMNH M-547426). The present specimen also compares well to the corallite dimensions and septal counts given for Dimorpharaea japonica Eguchi, 1951, but the type of this species seems to be lost; it could not be found in the collections of the Tohoku University Museum in Sendai, Japan (where the collection of Motoki Eguchi is nearly completely stored; Löser et al., 2002) by the first author in 1999 and 2007.
Other material BSPG 2003 XX 6317 (lower upper Aptian from Spain, Cataluña, Montsec de Rubies), ERNO L-4906 (lower Albian of Mexico, Sonora, Cerro de Oro).
Other occurrences Upper Aptian of the Western Tethys (Spain), Lower Albian of the Western Atlantic (Mexico), Lower Cenomanian of the Western Atlantic (USA).
Eocomoseris Melnikova et al., 1993
Type species Eocomoseris gurumdyensis Roniewicz, 2011
Description Astreoid colony. Corallite distance varies. Septa irregularly perforated. Microstructure of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. Symmetry of septa irregular, but two size orders can be distinguished. Septal generations differ in length. Septa occasionally connected to each other. Septal lateral face with pennulae. Pali absent. Some septa may be attached to the columella. Costae present, sub-confluent to non-confluent. Synapticulae present. Columella styliform. Endotheca varies. Wall absent. Coenosteum narrow, consists of costae. Budding extracalicinal.
Remarks For more details see Melnikova et al. (1993).
Eocomoseris sp.
Fig. 5 7–9
Material FPJ 374; 2 thin sections.
Dimensions (FPJ 374)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
cmin | 36 | 0.85–1.51 | 1.16 | 0.19 | 16.9 | 0.96–1.36 |
cmax | 21 | 1.19–2.10 | 1.56 | 0.25 | 16.0 | 1.31–1.81 |
ccd | 25 | 1.37–2.72 | 1.99 | 0.36 | 18.4 | 1.63–2.36 |
septa | 15 | 18–27 | 21.9 | 2.52 | 11.4 | 19–24 |
Description Astreoid colony. Corallite over a large distance. Septa irregularly perforated. Microstructure of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. Symmetry of septa irregular. Two irregular size orders can be distinguished, that differ in length, but not in thickness. Septa occasionally connected to each other in the centre of the corallite, and to the columella. Septal lateral face with pennulae. Pali absent. Some septa may be attached to the columella. Costae present, sub-confluent to non-confluent, and much more regularly perforated than septa. Synapticulae present and fairly common. Columella styliform and well marked. The endotheca has thin tabulae and extended thin tabulae. Wall absent. Coenosteum extended, consists of costae. Budding extracalicinal.
Remarks The problem of separating the numerous and mostly unnamed Eocomoseris species was already discussed in Löser et al. (2021). Only very few species are established but the genus is common in the Jurassic and Cretaceous. Because the corallite dimensions vary much within one colony (as well as being recognisable in the high values for the coefficient of variation for the corallite dimensions in the table above), it is difficult to limit well different species.
Superfamily Misistelloidea Eliášová, 1976
Family Misistellidae Eliášová, 1976
Miscellosmilia Eliášová, 1976
Type species. Miscellosmilia famosa Eliášová, 1976
Description Solitary turbinate or cylindrical coral with a circular outline. Septa compact. Microstructure of small-sized trabeculae, septa with a median dark line. Septa in cross section externally slightly thicker, becoming slightly thinner towards the centre. Symmetry of septa radial and regularly hexameral. Cycles of septa subregular. Septal cycles differ in length and thickness. First two or three septal cycles extend to the corallite centre, later cycles are shorter. Septa occasionally connected to each other close to the corallite centre. Septal distal margin unknown, lateral face with numerous thorns, inner margin smooth. Pali absent. Some septa may be attached to the columella. Short costae are present. Synapticulae absent. Columella lamellar. Endotheca consists of numerous thin tabulae. Wall absent. Epitheca present.
Remarks The genus was thoroughly described by Eliášová (1976).
Miscellosmilia famosa Eliášová, 1976
Fig. 6 1–2

1, 2 Miscellosmilia famosa Eliášová, 1976, FPJ 446. 1 transversal thin section. 2 transversal thin section, detail. 3, 4 Trochophyllia cf. melania (Fromentel, 1861), FPJ 384. 3 transversal thin section. 4 transversal thin section, detail. 5, 6 Montlivaltia icaunensis Orbigny, 1850, FPJ 440, 5 transversal thin section. 6 transversal thin section, detail. 7–9 Clausastrea superficialis Alloiteau, 1960, FPJ 439. 7 transversal thin section. 8 transversal thin section, detail. 9 longitudinal thin section. Scale bar 1 mm
*v 1976Miscellosmilia famosa: Eliášová, p. 342, Pl. 5, Fig. 1-3
Material FPJ 445, 446; 2 thin sections.
Dimensions (FPJ 445).
c 16.4 × 17.8 mm.
cl 15.2 × 15.8 mm.
septa 48.
sk12.
(FPJ 446)
C 20.6 × 20.9 mm.
cl 19.4 × 19.7 mm.
septa56.
sk12.
Description Solitary cylindrical corals with a circular outline. Septa compact. Microstructure not preserved. Septa in cross section externally slightly thicker, becoming slightly thinner towards the centre. Symmetry of septa radial and regularly hexameral. Cycles of septa subregular; there are three complete septal cycles and the beginning of a fourth cycle. Septal cycles differ in length and thickness. The first two cycles cannot be distinguished; they extend to the corallite centre, the third cycle reaches halfway around the corallite radius. The septa of the beginning fourth cycle are still shorter than those of the third cycle. Septa not connected to each other. Septal distal margin unknown, lateral face with thorns, inner margin smooth. Pali absent. Some septa of the first or second cycle are attached to the columella. Costae not visible. Synapticulae absent. Columella lamellar. Endotheca consists of thin tabulae. Wall absent. Epitheca present.
Remarks The genus is monospecific and the present two specimens compare well in corallite dimensions and septal counts to the holotype of Miscellosmilia famosa.
Other material ZPAL Hiii1629 (Lower Kimmeridgian of Poland, Swietokrzyskie: Holy Cross Mts).
Other occurrences Lower Kimmeridgian of the European Boreal (Poland), Tithonian to Lower Berriasian of the European Boreal (Czech Republic).
Family Rayasmiliidae Löser, 2022
Description Solitary and phaceloid corals. The septa are always free. A lamellar columella is present in some genera. One or two septa may be connected to the columella.
Trochophyllia Alloiteau, 1952
Type species Montlivaltia melania Fromentel, 1861
Description. Solitary turbinate coral. Corallite outline circular, corallite pit depressed. Septa compact. Microstructure of septa unknown. Septa in cross section externally slightly thicker, becoming slightly thinner towards the centre. Symmetry of septa radial and regularly hexameral or decameral. Cycles of septa regular. Septal cycles differ in length and thickness. First septal cycles extend to the corallite centre, later cycles are shorter and thinner. Septa not connected to each other. Septal distal margin smooth, lateral face with granules, inner margin smooth. Pali absent. Costae present, smooth on their surface. Synapticulae absent. Columella absent. Endotheca consists of dissepiments. Wall absent. Epitheca present.
Remarks. The genus has not received much attention after its creation. The description of the characteristics is based on the lectotype of Montlivaltia melania. Thin sections are not available from the type specimen, therefore the septal microstructure can only be inferred from the ornamentation of the upper septal margin.
Trochophyllia cf. melania (Fromentel, 1861)
Fig. 6 3–4
Material FPJ 384; 1 thin section.
Dimensions (FPJ 384)
cmin33.8
septa120
Description Solitary turbinate coral. Corallite outline elliptical. Septa compact. Microstructure of septa unknown. Septa in cross section externally slightly thicker, becoming slightly thinner towards the centre. Symmetry of septa radial and regularly hexameral. Cycles of septa regular. Septal cycles differ in length and thickness. Three septal cycles extend to the corallite centre, later cycles are shorter and thinner. Septa not connected to each other. Septal distal margin unknown, lateral face smooth (probably because of the poor state of conservation). The inner margin is slightly swollen. Pali absent. Costae present. Synapticulae absent. Columella absent. Endotheca consists of very few dissepiments. Wall absent. A very thin epitheca is present.
Remarks The present specimen is only a fragment. It can only be compared to T. melania because the lectotype of T. melania (MNHN M03534) has only 98 septa. This species was originally described from the Upper Oxfordian of Champlitten (France, Haute-Saône). The present specimen also compares to T. tourtiensis (Bölsche, 1871) but that species has much smaller corallite dimensions.
Superfamily Montlivaltioidea Felix, 1900
Family Montlivaltiidae Felix, 1900
Clausastrea Orbigny, 1849
Type species Clausastrea tessellata Orbigny, 1849
Description Astreoid or thamnasterioid colonies with corallites that can be arranged in rows. Corallite centres slightly depressed. Septa compact. Microstructure of septa of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. Symmetry of septa irregular, but size orders can be present. Septal generations differ in length. Half of all septa extend close to the corallite centre. Septa not connected to each other. Septal distal margin coarsely granulated, lateral face with vertical keels. Pali absent. Costae present, confluent to non-confluent. Synapticulae absent. Columella absent. Endotheca consists of tabulae. Wall absent. Coenosteum narrow, consists of costae and exothecal dissepiments. Budding extracalicinal.
Remarks The genus was the subject of an extended study by Alloiteau (1960).
Clausastrea sp.
Fig. 7 1–3

1–3 Clausastrea sp., FPJ 434. 1 transversal thin section. 2 transversal thin section, detail. 3 longitudinal thin section. 4, 5 Thamnasteria sp., FPJ 379. 4 transversal thin section. 5 transversal thin section, detail. Scale bar 1 mm
v 1998Clausastrea saltensis Alloiteau, 1960: Morycowa & Masse, p. 747, Figs. 14.1–4
Material FPJ 434; 2 thin sections.
Dimensions (FPJ 434)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 8 | 4.36–6.18 | 5.00 | 0.64 | 12.8 | 4.35–5.64 |
cdw | 12 | 2.53–5.56 | 3.81 | 1.05 | 27.6 | 2.76–4.87 |
septa | 10 | 14–18 | 16.5 | 1.26 | 7.6 | 15–18 |
Description Thamnasterioid colony with corallites arranged in irregular rows. Septa compact. Microstructure of septa of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. No symmetry of septa, but two size orders can be distinguished, that differ in length (best observable in the centre of the corallite). Septa not connected to each other. Septal distal margin unknown, lateral face with vertical keels. Pali absent. Costae present, mostly confluent. Synapticulae absent. Columella absent. Endotheca consists of numerous very thin tabulae and exothecal dissepiments. Wall absent. Coenosteum narrow, consists of costae and exothecal dissepiments. Budding extracalicinal.
Remarks The present specimen cannot be well compared to available type material. It is similar to the Late Jurassic to early Late Cretaceous Clausastrea alnisensis Alloiteau, 1960, but it has slightly smaller dimensions and higher septal counts. The holotype of C. alnisensis (MNHN R10859) has the following dimensions and septal counts: crd, 6–8, cdw, 3.5–6; septa, 13–13. These are minimum–maximum values taken from the complete specimen because thin sections are not available.
Other material BSPG 2003 XX 5474 (Lower Aptian of Greece, Viotía, Arachova), CGS HF 416 (Tithonian to lower Berriasian of the Czech Republic, Morava, Štramberk), TUM 65963 (Tithonian of Japan, Wakayama prefecture, Nabaenohana Peninsula).
Other occurrence Tithonian to Lower Berriasian of the European Boreal (Czech Republic), Tithonian of the Western Pacific (Japan), Lower Aptian of the Central Tethys (Greece) and Western Tethys (France).
Clausastrea superficialis Alloiteau, 1960
Fig. 6 7–9
*v 1960Clausastrea superficialis: Alloiteau, p. 21, Pl. 3, Fig. 5
v 1998Paraclausastrea pulchra nov. sp.: Morycowa & Masse, p. 750, Figs. 14.7, 15.4–6, 17
Material FPJ 439; 2 thin sections.
Dimensions (FPJ 439)
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 7 | 3.89–5.62 | 4.77 | 0.71 | 14.9 | 4.06–5.49 |
cdw | 8 | 3.18–6.01 | 4.45 | 0.89 | 20.1 | 3.55–5.34 |
septa | 10 | 19–27 | 22.7 | 2.79 | 12.2 | 20–25 |
Description Thamnasterioid, in places astreoid colony with corallites arranged in irregular rows. Septa compact. Microstructure of septa of large trabeculae. Septa in cross section equal in thickness throughout the whole septum. No symmetry of septa, but two size orders can be distinguished, that differ in length (best observable in the centre of the corallite). Septa not connected to each other. Septal distal margin unknown, lateral face with vertical keels. Pali absent. Costae present, non-confluent to confluent. Synapticulae absent. Columella absent. Endotheca consists of numerous very thin tabulae and exothecal dissepiments. Wall absent. Coenosteum narrow, consists of costae and exothecal dissepiments. Budding extracalicinal.
Remarks The distance between corallite rows and the septal counts compare well to the holotype of Clausastrea superficialis. Similar dimensions are present in C. eliasovae Ricci et al., 2018 but that species has much higher septal counts. All other species present larger distances of corallite rows.
Other material BSPG 2003 XX 5473 (Lower Aptian of Greece, Viotía, Arachova), BUFGG Do080 (Upper Oxfordian to lower Kimmeridgian of Romania, Tulcea, Dobrogea), CGS HF 348, 371, 376, 3244 (Tithonian to lower Berriasian of the Czech Republic, Morava, Štramberk).
Other occurrences Upper Oxfordian to Lower Berriasian of the Central Tethys (Romania, Czech Republic), Tithonian of the Western Tethys (Spain), Upper Barremian to Lower Aptian of the Western Tethys (France), Lower Aptian of the Central Tethys (Greece).
Montlivaltia Lamouroux, 1821
Type species Montlivaltia caryophyllata Lamouroux, 1821
Remarks The specimen MNHN A32924, marked as syntype of the type species, barely compares to the illustration in Lamouroux (1821) and it is questionable whether it belongs to the original type series. The genus Montlivaltia is conceptual and is generally applied in the sense of Gill and Lafuste (1971).
Montlivaltia icaunensis Orbigny, 1850
Fig. 6 5–6
*v 1850Montlivaltia icaunensis: Orbigny, (2), p. 90
v 1863Montlivaultia Icaunensis: Fromentel, p. 315, Pl. 42, Fig. 2, 2 a-b, Pl. 78, Fig. 3, 3 a, Pl. 81, Fig. 1, 1 a, 2, 2 a
vp 1889 Montlivaltia multiformis nov. spec.: Toula, p. 87, Pl. 6, Fig. 8
v 1946Montlivaltia burckhardti Wells, n.sp.: Wells, p. 5, Pl. 2, Fig. 7-14
v 2003Montlivaltia sp.: Baron-Szabo, Hamedani & Senowbari-Daryan, p. 204, Pl. 36, Fig. 8, Pl. 39, Fig. 6
Material FPJ 440; 1 thin section.
Dimensions (FPJ 440).
c 29 × 31.8 mm.
septa 96.
Description Solitary turbinate coral. Corallite outline elliptical. Septa compact. Microstructure of septa of large trabeculae. Septa in cross section externally slightly thicker, becoming slightly thinner towards the centre. Symmetry of septa radial and regularly hexameral. Cycles of septa regular. Septal cycles differ in length and thickness. Three septal cycles extend to the corallite centre, later cycles are shorter and thinner. Septa not connected to each other. Septal distal margin unknown, lateral face with vertical keels. The inner margin of the septa of the first two cycles is swollen. Pali absent. Costae present. Synapticulae absent. Columella absent. Endotheca consists of dissepiments. Wall absent. A very thin epitheca is present.
Remarks The genus Montlivaltia is one of the most species-rich genera in the Jurassic and Cretaceous with 224 species in the Jurassic and 74 in the Cretaceous (Lathuilière, 1989; Löser, 2000). It seemed to be always easier to establish a new species in place to compare own material to the vast number of existing taxa (Lathuilière, 1988). A correct taxonomic comparison is therefore difficult and must restrict to species where the type material was available for study. The present specimen compares well to the two syntypes of Montlivaltia icaunensis. Similar septal counts have also Montlivaltia cavali Achiardi, 1880, Montlivaltia coaluilensis Wells, 1946 but both differ in the dimensions of the corallite: Montlivaltia cavali has a much smaller diameter and Montlivaltia coaluilensis has a much larger corallite diameter.
Other material FLH 4452 (Kimmeridgian of Germany, Bayern, Saal an der Donau), UPV 220303 (Lower Albian of Spain, Cantabria, Cala de Islares).
Other occurrence. Kimmeridgian of the European Boreal (Germany), lower Hauterivian of the Western Atlantic (Mexico) and the European Boreal (France), Barremian of the Central Tethys (Bulgaria), Upper Aptian to Albian of the Eastern Tethys (Iran), Lower Albian of the Western Tethys (Spain).
Superfamily Rhizangioidea Orbigny, 1851
Family Thamnasteriidae Reuss, 1864
Thamnasteria Lesauvage, 1823
Type species. Thamnasteria lamourouxi Lesauvage, 1823
Description Thamnasterioid colony with corallites often arranged in rows. Septa compact. Microstructure of septa of medium-sized trabeculae. Septa in cross section externally slightly thicker, becoming slightly thinner towards the centre. No septal symmetry, but size orders can be distinguished. Septal generations differ in length. Approximately half of all septa extend to the calicular centre. Septa of younger generations often connected to the septa of preceding generations. Septal distal margin finely granulated, lateral face with fine thorns, inner margin smooth. Pali absent. All septa of the first generation are attached to the columella. Costae present, confluent. Occasional synapticulae present, mainly in the space between corallites. Columella styliform. Endotheca consists of numerous dissepiments. Wall absent. Budding extracalicinal.
Thamnasteria sp.
Fig. 7 4–5
Material FPJ 379, 383; 2 thin sections.
Dimensions (FPJ 379).
n | min–max | µ | s | cv | µ ± s | |
|---|---|---|---|---|---|---|
crd | 15 | 1.43–2.17 | 1.79 | 0.22 | 12.4 | 1.56–2.01 |
cdw | 30 | 1.44–2.28 | 1.87 | 0.22 | 11.9 | 1.64–2.09 |
septa | 10 | 20–25 | 22.8 | 1.75 | 7.6 | 21–25 |
Description Thamnasterioid colony with corallites loosely arranged in rows. Septa compact. Microstructure of septa of medium-sized trabeculae. Septa in cross section externally slightly thicker, becoming slightly thinner towards the centre. No septal symmetry, but size orders can be distinguished. Septal generations differ in length and slightly in thickness. Less than half of all septa extend to the calicular centre. Septa of younger generations often connected to the septa of preceding generations. Septal distal margin not preserved, lateral face with fine thorns, inner margin smooth. Pali absent. All septa of the first generation are attached to the columella. Costae present, confluent. Occasional synapticulae present, but only in the space between corallites. Columella styliform, thickened by the septa connected to it. Endotheca not preserved. Wall absent. Budding extracalicinal.
Remarks The present specimen cannot be compared to available type material. It shares some similarities with the syntype MNHN A32299 of Thamnaraea digitalis Etallon, 1864 from the middle Oxfordian of the Swiss Jura. This syntype has the following corallite dimensions and septal counts: ccd, 1.76–2.19; septa, 20–23 (first interval for both). Whereas in the present specimen the corallites are arranged in rows, they are irregularly distributed in the syntype MNHN A32299, which makes a comparison even more complicated. Other two syntypes (MJSN S184) belong to the genus Dendraraea Orbigny, 1849 (according to an additional specimen label signed by Bernard Lathuilière, Nancy, dating from 2010). A lectotype has so far not been designated. Therefore, it is unclear to which genus belongs the species Thamnaraea digitalis.
Other material GZG 611-74 (Upper Oxfordian of Germany, Niedersachsen, Hannover), MNHN A3299 (Middle Oxfordian of Switzerland, Jura, Calabri).
Other occurrence Middle Oxfordian of the Central Tethys (Switzerland), Upper Oxfordian of the European Boreal (Germany).
Discussion
The overall environmental conditions for the corals were not optimal because they rarely occur in the Herznach Member. The studied corals are represented by large solitary forms or massive platy colonies. Platy corals are considered to be an adaptation to low-level light conditions, because of either water depth or turbidity (Kolodziej & Bucur, 2020; Rosen et al., 2002; and literature cited therein). Five out of the eleven species belong to the superfamily Cyclolitoidea. This superfamily is characterised by having balcony-like septal ornamentation (pennulae). These pennulae are considered to support a heterotroph form of alimentation (Löser & Bilotte, 2017; Schlichter, 1991) being, therefore, better adapted to low-level light conditions. Comparable associations from the Cretaceous consist predominantly of members of the superfamily Cyclolitoidea (Löser & Bilotte, 2017; Löser & Callapez, 2022).
The fauna consists of twelve species in seven genera. This number is too low to allow any palaeobiogeographic analysis. The seven coral genera are already known from the Callovian, except for Miscellosmilia, that was hitherto only known from the Kimmeridgian onwards (Fig. 8). All genera reach into the Cretaceous, but only two occur beyond the Cenomanian. The distribution of species (Fig. 9) suggests that most species have their first occurrence in the study area, but the data available for these Middle Jurassic corals is too scarce to confirm this pattern with certainty. The distinction of species is here based on morphometric data that have a certain statistical significance (see Löser, 2020), achieved by systematic measurements in colonial corals. Such data are hardly available from the literature. Therefore, a comparison based on the literature alone is difficult. When not taking into account the occurrences in the Cretaceous, the coral species from the present fauna are also found in Middle to Upper Jurassic coral faunas of the Czech Republic (Moravia), England (Wiltshire), Germany (Bayern, Niedersachsen, Württemberg), Japan (Wakayama), Poland (Swietokrzyskie), Romania (Tulcea), and Spain (Aragón, Valencia).

Stratigraphical ranges of the genera of the study area. The blue bar indicates the age of the study area

Stratigraphic distribution and commonness of species (Middle Jurassic to Hauterivian). The thickness of the bars indicates the number of localities in which the concerned species was found. The blue bar indicates the age of the study area
Availability of data and materials
Not applicable.
References
Achiardi, A. (1880). Coralli giurassici dell’ Italia settentrionale. Atti Della Società Toscana di Scienze Naturali, 4, 233–310.
Agassiz, L. (1839). Description des Echinodermes fossiles de la Suisse. 1. Spatangoides et Clypeasteroides. Nouveau Mémoire de la Société Helvétique des Sciences Naturelles, 3, 1–101.
Agassiz, L. (1840). Description des Echinodermes fossiles de la Suisse. 2. Cidarides. Nouveau Mémoire de la Société Helvétique des Sciences Naturelles, 4, 1–107.
Allenbach, R. P. (2001). Synsedimentary tectonics in an epicontinental sea: A new interpretation of the Oxfordian basins of northern Switzerland. Eclogae Geologicae Helvetiae, 94, 265–287.
Allenbach, R., & Wetzel, A. (2006). Spatial patterns of Mesozoic facies relationships and the age of the Rhenish Lineament: A compilation. International Journal of Earth Sciences, 95, 803–813. https://doi.org/10.1007/s00531-006-0071-0
Alloiteau, J. (1952). Madréporaires post-paléozoïques. In J. Piveteau (Ed.), Traité de Paléontologie (Vol. 1, pp. 539–684). Masson.
Alloiteau, J. (1957). Contribution à la systématique des Madréporaires fossiles (pp. 1–462). Centre National de la Recherche Scientifique.
Alloiteau, J. (1960). Sur le genre Clausastrea. Annales de Paléontologie, (Invertébrés), 46, 3–46.
Arthaud, F., & Matte, P. H. (1977). Late Paleozoic strike-slip faulting in southern Europe and northern Africa. Geological Society of America Bulletin, 88, 1305–1320.
Baron-Szabo, R. C. (1997). Die Korallenfazies der ostalpinen Kreide (Helvetikum: Allgäuer Schrattenkalk; Nördliche Kalkalpen: Brandenberger Gosau) Taxonomie, Palökologie. Zitteliana, 21, 3–97.
Baron-Szabo, R. C., & González León, C. M. (2003). Late Aptian-Early Albian corals from the Mural Limestone of the Bisbee Group (Tuape and Cerro de Oro areas), Sonora, Mexico. In R. W. Scott (Ed.), Bob F. Perkins memorial volume (pp. 187–225). Special Publications in Geology.
Baron-Szabo, R. C., Hamedani, A., & Senowbari-Daryan, B. (2003). Scleractinian corals from Lower Cretaceous deposits north of Esfahan (Central Iran). Facies, 48, 199–216. https://doi.org/10.1007/bf02667539
Beauvais, L. (1963). Sur quelques genres de madréporaires peu connus de l’Argovien supérieur suisse. Bulletin de la Société Géologique de France, 7, 147–153.
Beauvais, L. (1966). Révision des madréporaires du Dogger de la collection Koby. Eclogae Geologicae Helvetiae, 59(2), 989–1024.
Beauvais, L. (1970). Madréporaires du Dogger: étude des types de la collection Milne-Edwards et J. Haime. Annales de Paléontologie. (Invertébrés), 56, 39–74.
Birenheide, R. (1969). Der Holotypus von Latusastrea valvata (Scleractinia, Oberer Jura). Senckenbergiana Lethaea, 50, 57–66.
Bitterli-Dreher, P. (2012). Die Ifenthal-Formation im nördlichen Jura. Bulletin für Angewandte Geologie, 17, 93–117.
Bölsche, W. (1866). Die Korallen des norddeutschen Jura- und Kreide-Gebirges. Zeitschrift der Deutschen Geologischen Gesellschaft, 18, 439–486.
Bölsche, W. (1871). Die Korallen des unteren Pläner im Sächsischen Elbthale. Palaeontographica, 20, 46–57.
Bölsche, W. (1877). Über einige Korallen aus der westphälischen Kreide. Jahresbericht des Naturwissenschaftlichen Vereins zu Osnabrück, 3, 68–71.
Boigk, H., & Schöneich, H. (1974). Perm, Trias und älterer Jura im Bereich der Mittelmeer-Mjösen-Zone und des Rheingrabens. In J. H. Illies & K. Fuchs (Eds.), Approaches to Taphrogenesis, Inter-Union Commission on Geodynamics, Scientific Report (pp. 60–71). Schweizerbart.
Bourne, G. C. (1900). The anthozoa. In R. Lankester (Ed.), Treatise on zoology. Part 2: Porifera and coelenterata (pp. 59–79). A. & C Black.
Brockkamp, O., Schlegel, A., & Clauer, N. (2011). Mesozoic hydrothermal impact on Rotliegende and Bunter immature sandstones of the High Rhine trough and its adjacent eastern area (southern Black Forest). Sedimentary Geology, 234, 76–88. https://doi.org/10.1016/j.sedgeo.2010.12.001
Burkhalter, R. M. (1996). Die Passwang-Alloformation (unteres Aalénien bis unteres Bajocien) im zentralen und nördlichen Schweizer Jura. Eclogae Geologicae Helvetiae, 89, 875–934.
Eguchi, M. (1951). Mesozoic hexacorals from Japan. Science Reports of the Tohoku Imperial University, (2: Geology), 24, 1–96.
Eliášová, H. (1976). Familles Placosmiliidae Alloiteau, 1952 et Misistellidae nov.fam. (Hexacorallia) des calcaires de Stramberk. Casopis pro Mineralogii a Geologii, 21, 337–347.
Etallon, A. (1864). Classe des Polypes. Neue Denkschriften der allgemeinen schweizerischen Gesellschaft für die gesammten Naturwissenschaften, 20, 357–412
Færseth, R. B. (1996). Interaction of Permo-Triassic and Jurassic extensional fault-blocks during the development of the northern North Sea. Journal of the Geological Society London, 153, 931–944.
Felix, J. (1900). Über die Gruppe der Montlivaltiaceae. Sitzungsberichte der Naturforschenden Gesellschaft zu Leipzig, 6(1900), 20–24.
Fromentel, E. (1861). Introduction à l’étude des polypiers fossiles. Mémoires de la Société D’émulation du Doubs, 3(5), 1–357.
Fromentel, E. (1863). Zoophytes, terrain crétacé (4). Paléontologie française (A.d' ;Orbigny ed.), 8, 145–192.
Gill, G.A. & Lafuste, J. (1971). Madréporaires simples du Dogger d’Afghanistan: Étude sur les structures de type <Montlivaltia>. Mémoires de la Société géologique de France, (n.s.), 50, 1–40.
Gonzalez, R. (1993). Die Hauptrogenstein-Formation der Nordwestschweiz (mittleres Bajocien bis unteres Bathonien). Dissertationen aus dem Geologisch-Paläontologischen Institut der Universität Basel, 2, 191.
Gonzalez, R., & Wetzel, A. (1996). Stratigraphy and paleogeography of the Hauptrogenstein and Klingnau Formations (middle Bajocian to late Bathonian), northern Switzerland. Eclogae Geologicae Helvetiae, 89, 695–720. https://doi.org/10.5169/seals-167921
Gründel, J., & Hostettler, B. (2012). Gastropoden aus dem Callovium von Liesberg Andil, Kanton Basellandschaft, und Les Grangettes, Kanton Jura. Freiberger Forschungshefte, C542, 65–90.
Gygi, R. A. (2000). Integrated stratigraphy of the Oxfordian and Kimmeridgian (Late Jurassic) in northern Switzerland and adjacent southern Germany. Denkschriften der Schweizerischen Akademie der Naturwissenschaften, 104, 1–151.
Gygi, R., & Persoz, F. (1986). Mineralostratigraphy, litho- and biostratigraphy combined in correlation of the Oxfordian (Late Jurassic) formations of the Swiss Jura range. Eclogae Geologicae Helvetiae, 79, 385–454.
Hostettler, B. (2014). Lithostratigraphie und Biostratigraphie im Callovien und Oxfordien des zentralen Nordwestschweizer Jura. Dissertation Universität Bern, 201
Koby, F. L. (1880–1889). Monographie des polypiers jurassiques de la suisse. Mémoires de Société Paleontologique, 7–16, 1–582.
Koechlin, E. (1933). Über das Vorkommen von Bajocienkorallen im Kanton Baselland. Verhandlungen der Naturforschenden Gesellschaft in Basel, 43, 4–11.
Koechlin, E. (1950). Demonstration einiger neuer oder wenig bekannter Fossilien aus dem Jura der Umgebung Basels. Eclogae Geologicae Helvetiae, 43, 217–220.
Kolodziej, B., & Bucur, I. I. (2020). An Early Cretaceous mesophotic coral ecosystem built by platy corals (middle Aptian, Southern Carpathians, Romania). Cretaceous Research, 109(104374), 1–12. https://doi.org/10.1016/j.cretres.2020.104374
Kutz, M., Löser, H., Franz, M., & Blattmann, E. (2020). Fossilien aus dem Breisgauer Hauptrogenstein (Bajocium, Mitteljura) unter besonderer Berücksichtigung der Korallen. Steinkern, 41, 14–33.
Lamouroux, J. V. F. (1821). Exposition méthodique des genres de l’ordre des polypiers (pp. 1–115). Agasse.
Lathuilière, B. (1988). Analyse de populations d’Isastrées bajociennes (Scléractiniaires jurassiques de France). Geobios, 21, 269–305.
Lathuilière, B. (1989). Répertoire objectif des coraux jurassiques (pp. 1–76). Presses Universitaires.
Lesauvage, M. (1823). Mémoire sur un genre nouveau de polypier fossile. Mémoires De La Société D’histoire Naturelle De Paris, 1, 241–244.
Löser, H. (2000). Repertoire of species. Catalogue of Cretaceous Corals, 1, 1–137.
Löser, H. (2012). Intraspecific variation in the genus Stelidioseris (family Actinastraeidae, suborder Archeocaeniina, order Scleractinia; Jurassic-Cretaceous). Geologica Belgica, 15(4), 382–387.
Löser, H. (2013). An Early Albian shallow marine coral fauna from Southern France – insight into evolution and palaeobiogeography of Cretaceous corals. Palaeobiodiversity and Palaeoenvironments, 93(1), 1–43. https://doi.org/10.1007/s12549-012-0088-2
Löser, H. (2016). Systematic part. Catalogue of Cretaceous Corals, 4, 1–710.
Löser, H. (2020). Regional persistence of the extant coral genus Stephanocoenia in the Western Hemisphere since the Early Cretaceous. Paläontologische Zeitschrift, 94(1), 17–39. https://doi.org/10.1007/s12542-019-00457-y
Löser, H. (2022). A new coral family and three new genera (Scleractinia) from the Lower Cretaceous of Puebla and Sonora, Mexico. Revista Mexicana de Ciencias Geológicas, 39, 220–229. https://doi.org/10.22201/cgeo.20072902e.2022.3.1698
Löser, H., & Bilotte, M. (2017). Taxonomy of a platy coral association from the Late Cenomanian of the southern Corbières (Aude, France). Annales De Paléontologie, 103, 3–17. https://doi.org/10.1016/j.annpal.2016.10.005
Löser, H., & Callapez, P. M. (2022). Upper Cenomanian and lower Turonian (Cretaceous) corals from the Tethyan West Portuguese Carbonate Platform. Journal of Iberian Geology, 48, 141–162. https://doi.org/10.1007/s41513-022-00186-3
Löser, H., Nieto, L. M., Castro, J. M., & Reolid, M. (2021). A Lower Valanginian coral fauna from the South Iberian Palaeomargin (Internal Prebetic, SE Spain). Palaeontologia Electronica, 24, 1–57. https://doi.org/10.26879/1030
Löser, H., Sugiyama, T., & Mori, K. (2002). Catalog of the Mesozoic corals at the Tohoku University Museum. Bulletin of the Tohoku University Museum, 2, 47–76.
Löser, H., Steuber, T., & Löser, C. (2018). Early Cenomanian coral faunas from Nea Nikopoli (Kozani, Greece; Cretaceous). Carnets de Géologie/Notebooks on Geology, 18, 23–121. https://doi.org/10.4267/2042/66094
Madritsch, H., Naef, H., Meier, B., Franzke, H. J., & Schreurs, G. (2018). Architecture and kinematics of the Constance-Frick Trough (Northern Switzerland): Implications for the formation of Post-Variscan basins in the foreland of the Alps and scenarios of their Neogene reactivation. Tectonics, 37, 2197–2220. https://doi.org/10.1029/2017TC004945
Maystrenko, Y., Bayer, U., Brink, H.-J., & Littke, R. (2008). The Central European Basin system—an overview. In R. Littke, U. Bayer, D. Gajewski, & S. Nelskamp (Eds.), Dynamics of complex intracontinental basins: the Central European Basin System (pp. 15–34). Berlin: Springer.
Matthews, S. C. (1973). Notes on open nomenclature and on synonymy lists. Palaeontology, 16, 713–719.
McCann, T., Pascal, C., Timmerman, M. J., Krzywiec, P., López-Gómez, J., Wetzel, A., Krawczyk, C. M., Rieke, H., & Lamarche, J. (2006). Post-Variscan (end Carboniferous-Early Permian) basin evolution in Western and Central Europe. In D. G. Gee & R. A. Stephenson (Eds.), European lithosphere dynamics (Vol. 32, pp. 355–388). Geological Society London, Memoirs.
Melnikova, G. K., Roniewicz, E., & Löser, H. (1993). New microsolenid coral genus Eocomoseris (Scleractinia, Early Lias-Cenomanian). Annales Societatis Geologorum Poloniae, 63, 3–12.
Ménard, G., & Molnar, P. (1988). Collapse of a Hercynian Tibetan Plateau into a late Palaeozoic European Basin and Range province. Nature, 334, 235–237.
Meyer, G. (1888). Die Korallen des Doggers von Elsass-Lothringen. Abhandlungen zur Geologischen Specialkarte von Elsass-Lothringen, 4, 1–44.
Milne Edwards, H. (1860). Histoire naturelle des coralliaires ou polypes proprement dits (3, Atlas) (pp. 1–560). Librairie encyclopédique de Roret.
Milne Edwards, H., & Haime, J. (1849). Mémoire sur les polypiers appartenant à la famille des oculinides, au groupe intermédiaire des Pseudoastréides et à la famille des Fongides. Comptes Rendus Hebdomadaires Des Séances De L’académie Des Sciences, 29, 67–73.
Milne Edwards, H., & Haime, J. (1851). A monograph of the British fossil corals (2:) Corals from the oolitic formations. Palaeontographical Society Monographs, 5, 74–146.
Morycowa, E., & Masse, J. P. (1998). Les Scléractiniaires du Barrémien-Aptien inférieur de Provence (SE de la France). Geobios, 31, 725–766.
NAGRA. (2002). Projekt Opalinuston—Synthese der geowissenschaftlichen Untersuchungsergebnisse. Nagra Technischer Bericht, 02–03, 1–659.
NAGRA. (2008). Vorschlag geologischer Standortgebiete für das SMA- und das HAA-Lager: Geologische Grundlagen (Textband + Beilagenband). Nagra Technischer Bericht, 08–04, 1–439.
Orbigny, A. (1849). Note sur les polypiers fossiles (pp. 1–12). Masson.
Orbigny, A. (1850). Prodrôme de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés (1–2). (pp. 1–394, 1–428). Masson
Orbigny, A. (1851). Cours élémentaire de Paléontologie (3:) Polypiers ou Zoophytes (pp. 151–189). Masson.
Pandey, D. K., McRoberts, C. A., & Pandit, M. K. (1999). Dimorpharaea de Fromentel, 1861 (Scleractinia, Anthozoa) from the Middle Jurassic of Kachchh, India. Journal of Paleontology, 73, 1015–1028.
Philippe, Y., Coletta, B., Deville, E. & Mascle, A. (1996). The Jura fold-and-thrust belt: a kinematic model based on map-balancing. In: P. Ziegler, & F. Horvath (Eds) Peri-Tethys Memoir 2: Structure and Prospects of Alpine Basins and Forelands, Mémoires du Muséum National d'Histoire Naturelle, 170. Éditions du Muséum Paris, Paris, (pp. 235–261)
Pieñkowski, G., Schudack, M. E., Bosák, P., Enay, R., Feldman-Olszewska, A., Golonka, J., Gutowski, J., Herngreen, G. F. W., Jordan, P., Krobicki, M., Lathuiliere, B., Leinfelder, R., Michalík, J., Mönnig, E., Noe-Nygaard, N., Pálfy, J., Pint, A., Rasser, M. W., Reisdorf, A. G., … Wong, T. E. (2008). Jurassic. In T. McCann (Ed.), The Geology of Central Europe (Volume 2: Mesozoic and Cenozoic) (pp. 823–922). The Geological Society.
Reisdorf, A. G., & Wetzel, A. (2018). Evidence for synsedimentary differential tectonic movements in a low-subsidence setting: Early Jurassic in northwestern Switzerland. Swiss Journal of Geosciences, 111, 417–444. https://doi.org/10.1007/s00015-018-0318-3
Reuss, A.E. (1864). Die fossilen Foraminiferen, Anthozoen und Bryozoen von Oberburg in Steiermark. Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse, 23, 1–38.
Ricci, C., Lathuilière, B., & Rusciadelli, G. (2018). Coral communities, zonation and paleoecology of an Upper Jurassic reef complex (Ellipsactinia limestones, central Apennines, Italy). Rivista Italiana Di Paleontologia e Stratigrafia, 124, 433–508.
Robin, C., Guillocheau, F., & Gaulier, J.-M. (1998). Discriminating between tectonic and eustatic controls on the stratigraphic record in the Paris basin. Terra Nova, 10, 323–329.
Roniewicz, E. (2011). Early Norian (Triassic) corals from the Northern Calcareous Alps, Austria, and the intra-Norian faunal turnover. Acta Palaeontologica Polonica, 56, 401–428. https://doi.org/10.4202/app.2009.0092
Rosen, B.R., Aillud, G.S., Bosellini, F.R., Clack, N., Insalaco, E., Valldeperas, X.F., & Wilson, M.E.J. (2002). Platy coral assemblages: 200 million years of functional stability in response to the limiting effects of light and turbidity. In: Moosa, M.K., Soemodihardjo, S., & Soegiarto, A. (Eds.): Proceedings of the ninth International coral reef symposium. Bali, 23–27 October 2000, (pp. 255–265)
Schlichter, D. (1991). A perforated gastrovascular cavity in Leptoseris fragilis: A new strategy to improve heterotrophic nutrition in corals. Naturwissenschaften, 78, 467–467.
Stäuble, A. (1959). Zur Stratigraphie des Callovian im zentralen Schweizer Jura. Eclogae Geologicae Helvetiae, 52, 57–176.
Thurmann, J., & Etallon, A. (1864). Lethaea bruntrutana. Études paléontologiques et stratigraphiques sur le Jura bernois et en particulier sur les environs de Porrentruy (3). Neue Denkschriften der Allgemeinen Schweizerischen Gesellschaft für die Gesammten Naturwissenschaften, 10, 356–580.
Timar-Geng, Z., Fügenschuh, B., Wetzel, A., & Dresmann, H. (2006). The low-temperature thermal history of northern Switzerland as revealed by fission track analysis and inverse thermal modelling. Eclogae Geologicae Helvetiae, 99, 255–270.
Tomes, R. F. (1878). A list of the Madreporaria of Crickley Hill, Gloucestershire, with descriptions of some new species. Geological Magazine, 5, 297–305.
Vaughan, T. W., & Wells, J. W. (1943). Revision of the suborders, families and genera of Scleractinia. Special Papers Geological Society of America, 44, 1–363.
von Raumer, J. F. (1998). The Paleozoic evolution in the Alps: From Gondwana to Pangea. Geologische Rundschau, 87, 407–435.
Wells, J. W. (1944). A new coral from the Buda Limestone (Cenomanian) of Texas. Journal of Paleontology, 18, 100.
Wells, J. W. (1946). Some Jurassic and Cretaceous corals from northern Mexico. Journal of Paleontology, 20(1), 1–7.
Wells, J. W. (1956). Scleractinia. In R. C. Moore (Ed.), Treatise on invertebrate paleontology, part F (pp. 328–444). Geological Society/University Kansas Press.
Wetzel, A., Allenbach, R., & Allia, V. (2003). Reactivated basement structures affecting the sedimentary facies in a tectonically “quiescent” epicontinental basin: An example from NW Switzerland. Sedimentary Geology, 157, 153–172. https://doi.org/10.1016/S0037-0738(02)00230-0
Wetzel, A., & Allia, V. (2000). The significance of hiatus beds in shallow-water mudstones: An example from the Middle Jurassic of Switzerland. Journal of Sedimentary Research, 70, 170–180. https://doi.org/10.1306/2DC40908-0E47-11D7-8643000102C1865D
Wetzel, A., & Meyer, C. A. (2006). The dangers of high-rise living on a muddy seafloor: An example of crinoids from shallow-water mudstones (Aalenian, northern Switzerland). Palaios, 21, 155–167. https://doi.org/10.2110/palo.2005.p05-132
Ziegler, P. A. (1990). Geological Atlas of Western and Central Europe (pp. 1–239). Shell Internationale Petroleum Maatschappij.
Acknowledgements
The preparation of thin sections in the ERNO laboratory by Aimée Orcí (Hermosillo, Sonora, Mexico) is gratefully acknowledged. A language check was carried out by Matthew Copley (Barcelona). We are grateful to two anonymous reviewers and the handling editor for critical comments on our contribution.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
HL: preparation of the fossil corals, measurements, identification, text (introduction, methods, systematic description, discussion; Figs. 1, 4, 5, 6, 7); AW: regional geology, text (introduction, geological setting, discussion; Fig. 2); BH: material collection and cleaning, local geology, text (introduction, geological setting; Fig. 3). All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Editorial handling: Michael Hautmann
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Löser, H., Wetzel, A. & Hostettler, B. Callovian corals from the Swiss Jura. Swiss J Palaeontol 143, 3 (2024). https://doi.org/10.1186/s13358-023-00298-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13358-023-00298-y