
Abstract
Degenerative disorders of joints, especially osteoarthritis (OA), result in persistent pain and disability and high costs to society. Nevertheless, the molecular mechanisms of OA have not yet been fully explained. OA is characterized by destruction of cartilage and loss of extracellular matrix (ECM). It is generally agreed that there is an association between pro-inflammatory cytokines and the development of OA. There is increased expression of matrix metalloproteinase (MMP) and “a disintegrin and metalloproteinase with thrombospondin motifs” (ADAMTS). Mesenchymal stem cells (MSCs) have been explored as a new treatment for OA during the last decade. It has been suggested that paracrine secretion of trophic factors, in which exosomes have a crucial role, contributes to the mechanism of MSC-based treatment of OA. The paracrine secretion of exosomes may play a role in the repair of joint tissue as well as MSC-based treatments for other disorders. Exosomes isolated from various stem cells may contribute to tissue regeneration in the heart, limbs, skin, and other tissues. Recent studies have indicated that exosomes (or similar particles) derived from MSCs may suppress OA development. Herein, for first time, we summarize the recent findings of studies on various exosomes derived from MSCs and their effectiveness in the treatment of OA. Moreover, we highlight the likely mechanisms of actions of exosomes in OA.
Similar content being viewed by others


Introduction
Osteoarthritis (OA), one of the most common musculoskeletal diseases, is characterized by sub-chondral bone sclerosis, synovial inflammation, cartilage degradation, ligament calcification, and osteophyte formation [1, 2]. Damage to joints following trauma, obesity, age, and genetic background are the major risk factors for OA [3]. OA leads to extensive public health costs for treatment and prevention, and a considerable loss of economic productivity due to worker incapacity [4, 5]. The direct cost of OA in the Canada, USA, UK, Australia, and France accounts for 1–2.5% of the gross national product of country; however, the indirect cost of OA are much higher, and it seems the true cost of OA is underestimated [5]. Direct costs are accounted for hospital stays, surgery, diagnosis, health professional visits, and treatments [5]. The indirect costs are those paid and unpaid activities, like employment, schooling, and homemaking, that result from disability associated with the health status [5]. Since the prevalence of OA rising with age, in an aging population, the OA is a growing source of indirect costs [5].
Currently, the treatments are mainly directed to relieving the symptoms of OA using non-steroid anti-inflammatory drugs (NSAID) and painkillers [6]. Because the mentioned drugs are not very effective, cannot ameliorate the disease and its symptoms for long periods of time, and also have many side effects [7], the search for novel therapeutic strategies is an important issue [8].
Mesenchymal stem/stromal cell (MSC)-based therapy is a promising new approach for OA, which has been further developed in recent years [9,10,11,12,13]. Preclinical studies have indicated that the cartilage of the joint can be protected from degeneration, and the development of OA can be delayed through intra-articular injection of MSCs isolated either from adipose tissue or from bone marrow [14,15,16,17]. Furthermore, some clinical trials have evaluated MSC-based treatments for OA and have shown decreases in inflammation and pain [12]. In a phase I–II clinical trial, Soler and colleagues have utilized ex vivo expanded autologous mesenchymal stromal cells for the treatment of osteoarthritis of the knee aiming to assess their safety and cartilage regeneration capacity. Their results showed that this cell-based therapy was well tolerated, though some adverse effect events (e.g., mild arthralgia and low back pain) were reported. There was a significant decrease in the intensity of pain since day 8 after the cell product administration and that was lasted after 12 months. The SF-36 QoL assay exhibited improvement of factors including bodily pain and physical functioning at month 12. The health assessment questionnaire showed a substantial decrease of disability. T2 mapping of cartilage also revealed signs of tissue regeneration in all patients at 12 months post-therapy [18]. In another study, Matas et al. have evaluated the safety and therapeutic efficacy of the single or repeated intraarticular administration of umbilical cord-derived (UC) MSCs in the knee of OA patients(a phase I/II trial) [19]. No severe adverse events were observed. MSC-treated patients showed more significant pain and greater function improvement from baseline. At 12 months, based on Western Ontario and Mc Master Universities Arthritis Index, MSC-treated patients (MSC-2 group) significantly experienced lower levels of pain versus the hyaluronic acid (HA) group. Visual analog scale for the pain and total Western Ontario and McMaster Universities Osteoarthritis Index (WOMAC) were also considerably lower in the MSC-2 group compared with the HA group at 12 months post-therapy [19].
MSCs may secrete mediators with chondro-protective and anti-inflammatory activities contributing to therapeutic impacts [20, 21].
MSC-derived extracellular vesicles (EVs) are a mixed population of heterogeneous membranous vesicles with distinct content which are critical mediators of intercellularcommunication. These vesicles establish a vesicle-mediated transport system to regulate a wide range of biological and pathological processes [22]. They are involved in cell-to-cell communication pathways [23]. The three major categories of EVs are (a) apoptotic bodies, (b) microparticles or microvesicles, and (c) exosomes or nanovesicles [24]. EVs are specified by their dimensions, expression of membrane markers, and their biogenesis. The endosomal compartments in multivesicular bodies are involved in producing exosomes. Endosomal markers (CD9, CD61, CD83, ALIX, TSG101) are expressed by exosomes, whereas microparticles (MPs) are released by a cell membrane budding process. It should be noted that MPs express markers according to their parental cells [2].
The crucial role of MSC-derived exosomes for the regulation of cell migration, proliferation, differentiation, and extracellular matrix synthesis has been increasingly supported by recent findings [25,26,27]. A report in 2010 showed that exosomes are secreted as active factors by MSCs responding to damage caused by myocardial ischemia reperfusion (I/R) [28]. Moreover, reports have demonstrated that MSC exosomes contribute to the repair and regeneration of cartilage via regulating immune reactivity, diminishing apoptosis, and increasing proliferation [29,30,31]. Exosomes, which function as intercellular communication vehicles, are small lipid-bilayer membrane vesicles between 50 and 150 nm in diameter [32]. Exosomes are able to transfer cargos of nucleic acids (mRNAs and microRNAs), proteins, and bioactive lipids [32]. Exosomes can produce biological responses in recipient cells [28]. Some findings have shown ambiguous effects of exosomes on the immune response or possible tumorigenicity, which may be considered unfavorable properties of exosomes [33, 34]. Nevertheless, there have been few studies conducted to investigate the precise molecular mechanisms by which MSC exosomes can promote cartilage chondrogenesis [27, 31, 35,36,37].
Recently, numerous investigations have been carried out to evaluate the potential role of exosomes derived from stem cells isolated from different sources. An in vivo study revealed that symptoms of OA could be decreased by exosomes isolated from iPS-derived MSCs, or from synovial MSCs; however, the highest efficiency was seen for exosomes from iPS-derived MSCs [38]. However in some studies, stem cells did not have a good therapeutic benefit in the treatment of OA. Tao and colleagues showed that the anabolic activity of chondrocytes was decreased by MSC-derived exosomes, unless the MSCs had been engineered to express miR-140-5p [39]. Also, the beneficial effects of embryonic stem cell-derived exosomes have been reported in a model involving destabilizing the medial meniscus (DMM) [36]. As yet, there is no information about the effect of other kinds of EVs on OA. In this review, we summarize the pathogenic roles of exosomes in OA, and the evidence for exosomes derived from MSCs in the treatment of OA.
Exosomes: biogenesis, cargos, and different subtypes
Biogenesis of exosomes
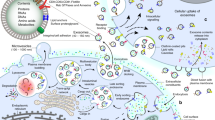
Exosomes have dimensions ranging between 30 and 150 nm. They are nanoscale EVs that are produced by nearly all types of cells [40, 41]. Exosomes are produced by the endosomal network, via intra-luminal vesicles (ILV, also known as pre-exosomes) formed by inward budding of the membranes of multivesicular bodies [40, 42]. Numerous different pathways [43], such as ESCRT (endosomal sorting complexes required for transport), and ESCRT-independent pathways are involved in exosome biogenesis (Fig. 1). Exosome biogenesis requires four separate multiprotein sub-complexes (ESCRT-0, ESCRT-I, ESCRT-II, and ESCRT-III) that compose the ESCRT membrane-scission machinery. Ubiquitinated proteins are bound in initial ESCRT complexes (ESCRT-0, ESCRT-I, and ESCRT-III) via their ubiquitin-binding sub-units, which result in the formation of strongly bound complexes with proteins in the cytoplasm. The ESCRT-III complex temporarily assembles on the endosome membrane and undergoes vesicle scission [44]. It has been found that additional components, such as apoptosis-linked gene 2 (ALG-2)-interacting protein X (ALIX) (a cytoplasmic protein concentrated and expressed in exosomes and phagosomes), ATPase, and vacuolar protein sorting-associated protein (VPS4) are involved in regulating the ESCRT membrane-scission machinery [44]. Moreover, lipids such as the sphingolipids, ceramide [45, 46], and sphingosine 1-phosphate function in the ESCRT-independent pathway to regulate the release of exosomes [47].

Exosome biogenesis and its relationship with osteoarthritis. A clathrin-dependent pathway or a clathrin-independent pathway initially mediates endocytosis, at a lipid raft. The endocytic vesicles contain signaling proteins, growth factor receptors, oncoproteins, combined with normal membrane proteins, including tetraspanins (e.g., CD9, CD63, and CD81), MHC I and II, and adhesion molecules (e.g., cadherins, integrins). Exosome biogenesis occurs via the endosomal network in the endosomal sorting complexes needed for ESCRT-independent or ESCRT-dependent pathways. Inward budding of MVB produces intra-luminal vesicles (exosomes). Several cytoplasmic molecules (e.g., heat shock proteins, ubiquitin-related proteins, mRNAs, microRNAs [miRNAs], cytoskeleton proteins) and nuclear molecules (e.g., long-noncoding RNAs [lncRNAs], transcriptional factors, DNAs) can be loaded into MVB by stage-specific pathways, some of which are osteoarthritis type-specific. Moreover, plasma membrane fusion of multi-vesicular bodies leads to release of exosomes by exocytosis. Numerous Rab GTPases (such as Rab11/35, Rab7, and Rab27) are present in secreted exosomes. Eventually, MSC-derived exosomes are transported to the osteoarthritis micro-environment where they modulate osteoarthritis. ESCRT, endosomal sorting complexes required for transport; MHC, major histocompatibility complex; MSCs, mesenchymal stem cells; MVB, multi-vesicular bodies; rER, rough endoplasmic reticulum; sER, smooth endoplasmic reticulum; Rab, Ras-associated binding
Cargos of exosomes
The molecular components of exosomes, including lipids, nucleic acids, metabolites, and proteins, differ from each other, based on the cellular origin, environment, developmental phase, epigenetic modification, and the precise biogenesis mechanism. Moreover, studies have identified a variety of RNA types, such as microRNAs (miRNAs), messenger RNAs (mRNAs), transfer RNAs (tRNAs), long noncoding RNAs (lncRNAs), or ribosomal RNAs (rRNAs) that all can be present in exosomes [48]. Different types of RNA cargo may be involved in epigenetic modification of cells and altering biological activities. Recent studies have indicated that double-stranded DNAs (dsDNAs) found in tumor-derived exosomes could reflect the mutational status of their parental tumors [49]. Studies have shown that > 10 kb pieces of cellular genomic DNA are indicative of mutations in oncogenes or tumor suppressor genes in tumor-derived exosomes [50]. Furthermore, various bioactive proteins, which originate from the plasma membrane and the cytoplasm, exist within exosomes. Certain proteins, including Ras-related GTP-binding protein (Rab), ALIX, or ESCRT complexes that participate in exosome biogenesis, have been found in exosomes [44]. Additional proteins, including tetraspanins (i.e., CD9, CD63, and CD81), heat shock proteins (e.g., HSP70 or HSP 90), and integrins, can be packaged into exosomes, which are involved in intracellular assemblage or trafficking [40]. The expression patterns of proteins which reflect the pathophysiological status of the parental cells can be determined by analysis of exosomal proteins. The composition of the membranes of exosomes is characteristic of lipid rafts. They contain larger amounts of particular lipid species, such as sphingomyelin, phosphatidylcholine, cholesterol, ceramide, and diacylglycerol in comparison with their parental cells [51]. The release and formation of exosomes are regulated by lipid-metabolizing enzymes, such as phospholipase D2 (PLD2) or neutral sphingomyelinase (nSMase) [45].
Subtypes of exosomes
Most researchers, who have investigated exosome-mediated cell functions, have used heterogeneous exosome preparations with different molecular cargos, and varying dimensions and morphology, all resulting in different biophysical properties. These have included large exosome vesicles (90 to 120 nm), small exosome vesicles (60 to 80 nm), and non-membranous nanoparticles (also known as exomeres, size about 35 nm) [52]. Glycomic, lipidomic, and proteomic analyses have shown different molecular profiles inside exosomes and exomeres. Proteomic analysis has shown a higher expression of enzymes associated with metabolism, hypoxia, and microtubule and coagulation-related proteins in exomeres, in comparison with large or small membranous exosomal vesicles. On the other hand, large and small exosomal vesicles contain higher amounts of signaling proteins associated with the mitotic spindle, interleukin (IL)-2/STAT5, and proteins associated with endosome secretion pathways [52]. Differences between exomeres and exosomes also involve sialylated glycoproteins (i.e., galectin-3-binding protein) [52]. Because there are differences in the lipid composition between different exosomes (and also exomeres) [52], density gradient centrifugation can separate them into two distinguishable populations: higher density exosomes (HD-Exo) and lower density exosomes (LD-Exo) [53]. An increase in “solute carrier family 38 member 1” (SLC38A1) was demonstrated in LD-Exo-treated endothelial cells in comparison with HD-Exo-treated endothelial cells [53]. Overall, studies have indicated that different biological effects can be triggered on recipient cells by using different exosomal sub-populations.
Exosomes and osteoarthritis
mRNAs, proteins, long noncoding RNAs (lncRNA, cirRNA), and miRNAs are all found in the exosomal contents. Changes in gene expression, downstream function, and processes related to physiology or pathology may be caused by the action of exosomes on recipient cells [54].
Recently, some studies have been carried out to evaluate the various effects of exosomes on different cells involved in the joint diseases [23, 42, 55, 56]. Cell-derived EVs have been isolated from synovial fibroblasts (SF) extracted from the inflamed joints of OA and RA patients and investigated their role in the cellular processes such as inflammation and cartilage degeneration which are implicated in the disease progression [56,57,58]. Domenis et al. explored the immune regulatory properties of SF-derived exosomes from end-stage OA patients, on macrophages differentiated from human peripheral blood mononuclear cells (PBMCs) [59]. When patient cells were treated by exosomes, it was demonstrated that the macrophages generated a spectrum of chemokines and pro-inflammatory cytokines, such as CCL8, IL-1β, MMP12, CCL15, MMP7, and CCL20, which would result in cartilage degradation and inflammation in joints [59]. Kolhe et al. performed similar experiments and showed a significant decrease in cell survival and the expression of anabolic genes (COL2A1, ACAN), and an increase in the expression of catabolic and inflammatory genes (IL-6, TNF-α) using articular chondrocytes treated with exosomes derived from SF from OA patients [60]. Kato et al. [61] investigated whether exosomes mediated the interaction between articular chondrocytes and inflammatory synovial fibroblasts (SFBs). Exosomes were isolated from untreated similar fragment pairs (SFBs) and from similar fragment blocks (SFPs) that had been treated or not with IL-1β, and were then added to normal articular chondrocytes. These workers showed upregulation of the expression of MMP-13 and ADAMTS-5 and downregulation of ACAN and COL2A1 in articular chondrocytes when treated with IL-1β-treated SFB-derived exosomes, compared to exosomes from untreated SFBs. Additionally, exosomes from IL-1β-treated SFBs produced OA-like changes in both in vitro and in vivo models.
Two studies carried out comparisons between exosomes from various sources for discovering arthritis mechanisms. SF exosomes from patients with RA and OA were compared by Zhang et al. They observed a membrane-bound form of TNF-α in exosomes generated from SF of patients with RA; however, they did not see this phenomenon in the case of OA. Moreover, another research found that this kind of TNF-α enhances SFB exosome generation in RA and forms a positive amplification loop in RA pathogenesis [62].
Kolhe et al. compared SF-derived exosomes isolated from females and males with OA and non-OA in order to examine sex differences in OA pathophysiology. They observed a difference in the miRNA contents of EVs between groups with OA and non-OA, as well as between the two sexes [60]. The “Kyoto Encyclopedia of Genes and Genomes” (KEGG) represents a comprehensive database which is used to map associated genes to their respective pathways [63]. Though pathway maps, the KEGG pathway database provides pathway diagrams in both healthy and diseased states [64]. KEGG annotation analysis was used to assess the relationship between glycan degradation, cell adhesion molecules, and mucin type O-glycan biosynthesis and downregulated miRNAs in OA for each sex. Moreover, the possible involvement of thyroid hormone synthesis, biotin metabolism, and signaling related to amphetamine addiction was investigated along with upregulated miRNAs. Contributions from ovarian steroidogenesis and estrogen signaling pathways were also detected using KEGG annotation analysis in female OA patients [60]. Table 1 shows exosomes and their cargos involved in arthritis.
MSC-derived exosomes and OA
There has been ever-increasing interest in the clinical application of MSCs for a variety of diseases in recent years. MSCs have been widely studied for their potential to treat joint damage and OA [27, 79,80,81,82,83]. The MSCs have usually been isolated from synovium [84], bone marrow [85], and adipose tissue [21]. Researchers have assessed the effectiveness of MSCs in restoration of damaged tissue function or alleviate disease symptoms in OA or cartilage damage [86, 87]. For example, the safety and tolerability over the medium-term (~ 5 years) of intra-articular injection of MSCs has been shown for the treatment of knee in the OA in phase I and II clinical trials. This success has led to the reduction of pain and enhancement of joint performance and improved quality of cartilage repair [88,89,90].
It has been revealed that MSC-derived exosomes could protect cartilage and bone from degradation in OA by increasing the expression of chondrocyte markers like type II collagen and aggrecan, reducing catabolic markers such as MMP-13 and ADAMTS5, decreasing inflammatory markers (iNOS), protecting chondrocytes from apoptosis, and blocking of macrophage activation [2]. Furthermore, it has been exhibited that MSC-derived exosomes could attenuate OA by stimulation of chondrocyte migration and proliferation [38].
The motivation for transplantation of MSC for treatment of joint diseases is based on their potential for multi-lineage differentiation into mesenchymal cell types (e.g., bone, cartilage, and fat tissues) which are all essential for musculoskeletal repair. Of course, treatment with allogeneic MSC is relied on the lack of evoking an immune response [90]. Nevertheless, numerous researchers have found that in spite of the functional enhancement (or even the regeneration of joint tissue) which was observed following transplantation of MSCs into diseased joints, their engraftment and subsequent differentiation into the desirable cell types only occurred only rarely [91]. The paracrine function of the MSCs has been suggested to explain these observations, with the hypothesis that MSCs principally execute their therapeutic effect through the secretion of trophic factors that improve regeneration and decrease inflammation [92]. In the next section, the promising role of MSC-derived exosomes in the mitigation of inflammation in the OA is discussed in detail.
MSC-derived exosomes and their anti-inflammatory effects in OA
Although OA is commonly regarded as a degenerative disease, recent work has proposed that low-grade inflammatory processes are able to induce disease symptoms and promote the progression of disease [93]. Hence, immunomodulatory function of MSCs should be further considered. MSCs are able to promote the regeneration of joint components through their two secretory functions as follows:
-
a.
Anti-inflammatory factors. MSCs may downregulate inflammatory signals in osteoarthritic cartilage, by secretion of interleukin (IL)-1β, IL-6, IL-8, matrix metalloproteinase (MMP)-1, and MMP-13 [94, 95].
-
b.
Trophic factors. These are molecules that give rise to cell proliferation, decrease formation of scar tissue, and trigger the repair of endogenous cartilage, such as epithelial growth factor (EGF), insulin-like growth factor (IGF)-1, basic fibroblast growth factor (bFGF), transforming growth factor (TGF)-β, and vascular endothelial growth factor (VEGF) [94, 96].
Although there is much evidence for the ability of MSCs to heal damage to joints, and to treat OA, there are several problems with the approach of direct cell transplantation, such as the poor survival of the cells after injection, the inability to predict lasting improvements in cell behavior and cell-cell interactions, and problems in maintaining an adequate storage bank of cells to allow off-the-shelf treatment [97]. The suitability of donors may be another important challenge, because it was found that MSCs isolated from old or otherwise unhealthy donors led to decreased performance and proliferation [98]. Moreover, the induction of senescence, loss of proliferative potential, and reduced capacity for differentiation (particularly beyond 10–20 population doublings) have been attributed to prolonged ex vivo cell expansion of MSCs before transplantation [98]. Because of their genetic programming to undergo calcification after chondrogenic induction as part of the normal process of endochondral ossification, there are problems in maintaining the cartilage phenotype in differentiated MSCs and preventing them from expanding towards the osteogenic phenotype [99]. Moreover, MSCs are sensitive to certain “environmentally responsive” factors and undergo specific behavioral modification in response to minute traces of unknown substances in the microenvironment [96]. Although this responsiveness is frequently utilized in regenerative medicine, it can have a negative impact on the MSC response in a diseased joint environment. For example, reports have demonstrated that human adipose tissue-derived MSCs can switch to a pro-inflammatory secretome, when treated with tumor necrosis factor (TNF), and can then play a role in augmenting the inflammatory response [100]. Therefore, current studies have focused on testing the secretory products of MSCs, including MSC-derived exosomes in experimental models of joint damage and OA, rather than transplanting the cells themselves.
There are many similarities between the biological effects of stem cell-derived EVs and the intact stem cells themselves. However, stem cell-derived EVs have advantages (e.g., small dimensions, low immunogenicity, and elimination of procedural issues related to direct cell injection). EVs were first described as “platelet dust” in the 1960s [101] and have been intensely investigated by researchers during the past decades. EVs are now realized to be powerful intercellular messengers that not only can mediate pathological processes, but can also maintain tissue homeostasis, and can modulate physiological functions in a variety of therapeutic approaches [102, 103]. They have been studied as anti-tumor treatments, to improve vaccination against pathogens, as immunomodulatory treatments, as drug delivery vectors, and to improve regenerative therapies [104]. Although several studies have been conducted on the use of stem cell-derived EVs as a treatment for joint damage and OA, there is still a lack of hard evidence [23, 58]. However, there are a few studies on this emerging and novel topic, suggesting the powerful ability of stem cell-derived EVs to improve joint repair and protect joints from degeneration after damage (Table 2).
Zhu et al. carried out a comparative study between exosomes derived from synovial membrane MSCs (SMMSC-Exos) and exosomes derived from induced pluripotent stem cell-derived MSCs (iMSC-Exos) to treat OA [38]. These researchers showed that the diameter of both SMMSC-Exos and iMSC-Exos was between 50 and 150 nm. These exosomes expressed TSG101, CD9, and CD63. Injection of both SMMSC-Exos and iMSC-Exos into a murine OA model reduced symptoms of OA; however, a greater therapeutic impact was achieved with iMSC-Exos compared to SMMSC-Exos. Both SMMSC-Exos and iMSC-Exos triggered the proliferation and migration of chondrocytes. Overall, they showed a higher therapeutic impact of iMSC-Exos in comparison with SMMSC-Exos [38, 114]. Since autologous iMSCs can be more readily obtained, they may provide a new treatment procedure for OA [38].
Zhang et al. evaluated the contribution of MSC exosomes to regulate the inflammatory response, nociceptive behavior, condylar cartilage, and subchondral bone preservation in an immunocompetent rat model of temporomandibular joint osteoarthritis (TMJ-OA) [109]. These researchers showed that there was an initial inhibition of pain and degeneration with decreased inflammation, accompanied by a strong proliferation and gradual enhancement in matrix expression and the architecture of the subchondral bone, which demonstrated an overall joint repair and reconstruction. Chondrocyte culture can be used to assess cellular activities during exosome-mediated joint repair, measuring adenosine signaling and the activity of AKT, ERK, and AMPK. MSC-derived exosomes increased s-GAG synthesis that had been inhibited by IL-1β, and reduced IL-1β-induced nitric oxide and MMP13 production. Inhibitors of adenosine receptor activation and AKT, ERK, and AMPK phosphorylation partly reduced these effects. Overall, Zhang et al. observed increased TMJ restoration and reconstruction in OA, via a mechanism mediated by MSC exosomes. This mechanism involved several cellular processes involving repair of the matrix and general joint homeostasis. The researchers suggested this was a cell-free, ready-to-use exosome-based therapy to treat TMJ pain and degeneration [109].
Therapeutic potentials of MSC-derived exosomes from different cell origins in OA
The cargo of exosomes derived from various cell types is highly heterogeneous and comprised of different biological molecules such as proteins, nucleic acids, and lipids that not only show the cell origin of exosomes, but also reflect the pathological or physiological conditions of the cell of origin [115, 116].
MSC-derived exosomes take part in intercellular communications and also deliver various miRNAs, mRNAs, and proteins into recipient cells, in a similar manner to exosomes derived from other cells [97]. Up to now, more than 150 miRNAs and 850 unique gene products have been recognized in MSC-derived exosomes [117, 118]. Prior studies have shown that, depending on the MSC source, the function and phenotype of MSC-derived exosomes may vary [119]. By utilizing RNA sequencing and comparative analysis, it has been shown that the content of exosomes derived from MSCs of human adipose tissue and bone marrow has significant differences in their composition, like certain tRNA species (Nanog, POU5F1A/B, and Sox2 expression), which seems to be correlated with the differentiation status of MSCs and their cell of origin [119, 120]. The therapeutic properties of human MSCs derived from adipose tissues, bone marrow, and endometrium have been recently compared in an experimental model of myocardial infarction [121]. The findings revealed the superior cardioprotection with endometrial MSCs compared to adipose and bone marrow-derived MSCs [121]. These data suggest that intrinsic differences among exosomes derived from MSCs isolated from different cellular sources should be considered as these differences may have profound impact on therapeutic outcomes [121].
The degeneration of cartilage tissue during OA progression is caused by chronic inflammation. Adipose tissue-derived MSCs (AD-MSCs) have the potential to blunt degenerative and inflammatory processes in OA. These cells also showed a paracrine effect on chondrocytes. Tofiño-Vian et al. have revealed that EVs isolated from human AD-MSCs exerted chondroprotective functions through multiple mechanisms such as reduced the production of inflammatory mediators (e.g., TNF-α, IL-6, PGE2 and NO), decreased the release of MMP activity, and enhanced the production of the anti-inflammatory cytokine IL-10 [110]. The authors concluded that AD-MSC extracellular vesicles can be employed for the development of new therapeutic approaches in joint conditions.
Researchers have comparatively evaluated the function of exosomes or microvesicles/microparticles (MPs) in OA [2]. These researchers showed that there is no significant difference between function of exosomes and MPs derived from BM-MSCs. They also found that treatment with both exosomes and MPs derived from BM-MSCs can not only restore the expression of mature articular chondrocytes markers (e.g., type II collagen and aggrecan), but also decrease the expression of catabolic (e.g., MMP-13, ADAMTS5) and inflammatory (e.g., iNOS) markers in a dose-dependent manner. Intriguingly, the highest dose of BM-MSC-derived MPs or exosomes could not only reverse the OA phenotype of chondrocytes but also could modulate anabolic and catabolic chondrocyte markers to a similar extent as BM-MSCs. Another important finding was that preactivation of BM-MSCs with TGFβ3 can improve the effectiveness of both exosomes and MPs. Although the differences were relatively small in comparison with non-preactivated BM-MSCs, the gene expression pattern was significantly different in the MPs or exosomes derived from pre-activated BM-MSCs [2]. In addition, researchers have shown that BM-MSCs may contribute to mediate an anti-fibrotic function [122]. They speculated that TGFβ3 preactivation of BM-MSCs could trigger their anti-fibroblastic and pro-chondrogenic function via the release of factors which could be transferred by exosomes and MPs. They proved the beneficial effects of MPs and exosomes pathogenic proliferation of fibroblast and skin fibrosis. In aggregate, these studies indicate that the therapeutic potential of MSCs and their derivative exosomes. These studies further emphasized that therapeutic potential of MSC-derived exosomes could largely vary based on their content composition and cell of origin in different pathological and physiological conditions and should be considered for the therapeutic applications. Recompositioning of exosome content through ex vivo pretreatment and/or preactivation of MSCs with different compounds (e.g., TGFβ3) would be also another interesting therapeutic approach to maximize anti-OA potential of exosomes.
Conclusions
Exosomes carry out many different functions in organisms that include repair of tissue injuries, regulation of immune response, and inhibition of inflammation. Due to their ability to repair damaged tissues, MSC-derived exosomes have been widely studied in regenerative medicine. The improvement in tissue homeostasis caused by MSCs can be produced by cell-to-cell direct interaction and also by secretion of soluble factors. Exosomes are a kind of soluble biological mediator isolated from MSCs culture media in vitro. MSC-derived exosomes are generated under both pathological and physiological conditions. They are primary mediators of intercellular communications by transferring mRNAs, lipids, siRNA, proteins, miRNAs, and ribosomal RNAs to adjacent cells or remote cells. Different disease models have been studied in MSC-derived exosome experiments. Findings have demonstrated a similarity of function between MSC-derived exosomes and intact MSCs themselves. This review has presented the evidence for MSC-derived exosomes as a new approach to cell-free treatment of OA and joint damage. It is clear that numerous additional investigations must be performed to prove the effectiveness and feasibility of MSC-derived exosomes in the therapy of OA in patients.
Availability of data and materials
The primary data for this study is available from the authors on direct request.
Abbreviations
- AD-MSCs:
-
Adipose tissue-derived mesenchymal stem cells
- ALG-2:
-
Apoptosis-linked gene 2
- ALIX:
-
ALG-2-interacting protein X
- bFGF:
-
Basic fibroblast growth factor
- BM-MSCs:
-
Bone marrow-derived mesenchymal stem cells
- DMM:
-
Destabilizing a medial meniscus
- dsDNAs:
-
Ribosomal RNAs
- EGF:
-
Epithelial growth factor
- ESCRT:
-
Endosomal sorting complexes required for transport
- EVs:
-
Extracellular vesicles
- HEK:
-
Human embryonic kidney cell line
- IGF-1:
-
Insulin-like growth factor1
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- LD-Exo:
-
Lower density exosomes
- LncRNA:
-
Long noncoding RNAs
- MMP:
-
Matrix metalloproteinase
- MPs:
-
Microparticles
- MSCs:
-
Mesenchymal stem cells
- MVB:
-
Multi-vesicular bodies
- NO:
-
Nitric oxide
- NSAID:
-
Non-steroid anti-inflammatory drugs
- OA:
-
Osteoarthritis
- PBMCs:
-
Peripheral blood mononuclear cells
- PGE2:
-
Prostaglandin E2
- PLD2:
-
Phospholipase D2
- Rab:
-
Ras-related gtp-binding protein
- rRNAs:
-
Ribosomal RNAs
- SLC38A1: SFBs:
-
Synovial fibroblasts; solute carrier family 38 member 1
- TGF- β:
-
Transforming growth factor-beta
- TMJ-OA:
-
Temporomandibular joint osteoarthritis
- TNF:
-
Tumor necrosis factor
- tRNAs:
-
Transfer RNAs
- VEGF:
-
Vascular endothelial growth factor
- VPS4:
-
Vacuolar protein sorting-associated protein
References
Kraus VB, Blanco FJ, Englund M, Karsdal MA, Lohmander LS. Call for standardized definitions of osteoarthritis and risk stratification for clinical trials and clinical use. Osteoarthr Cartil. 2015;23(8):1233–41.
Cosenza S, Ruiz M, Toupet K, Jorgensen C, Noël D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci Rep. 2017;7(1):16214.
Palazzo C, Ravaud J-F, Papelard A, Ravaud P, Poiraudeau S. The burden of musculoskeletal conditions. PLoS One. 2014;9(3):e90633.
Puig-Junoy J, Ruiz Zamora A. Socio-economic costs of osteoarthritis: a systematic review of cost-of-illness studies. Semin Arthritis Rheum. 2015;44(5):531–41.
Iban MAR, Benavides J, Forero JP, Bittelman S, Martinez R, Mite MA, Heredia JD, Ulloa S, Ferrand MML. Use of strong opioids for chronic pain in osteoarthritis: an insight into the Latin American reality; 2017.
Rannou F, Pelletier JP, Martel-Pelletier J. Efficacy and safety of topical NSAIDs in the management of osteoarthritis: evidence from real-life setting trials and surveys. Semin Arthritis Rheum. 2016;45(4 Suppl):S18–21.
Silverstein FE, Faich G, Goldstein JL, Simon LS, Pincus T, Whelton A, Makuch R, Eisen G, Agrawal NM, Stenson WF, et al. Gastrointestinal toxicity with celecoxib vs nonsteroidal anti-inflammatory drugs for osteoarthritis and rheumatoid arthritis: the CLASS study: a randomized controlled trial. Celecoxib Long-term Arthritis Safety Study. JAMA. 2000;284(10):1247–55.
Jordan KM, Arden NK, Doherty M, Bannwarth B, Bijlsma JW, Dieppe P, Gunther K, Hauselmann H, Herrero-Beaumont G, Kaklamanis P, et al. EULAR recommendations 2003: an evidence based approach to the management of knee osteoarthritis: report of a Task Force of the Standing Committee for International Clinical Studies Including Therapeutic Trials (ESCISIT). Ann Rheum Dis. 2003;62(12):1145–55.
Al-Najar M, Khalil H, Al-Ajlouni J, Al-Antary E, Hamdan M, Rahmeh R, Alhattab D, Samara O, Yasin M, Abdullah AA, et al. Intra-articular injection of expanded autologous bone marrow mesenchymal cells in moderate and severe knee osteoarthritis is safe: a phase I/II study. J Orthop Surg Res. 2017;12(1):190.
Bastos R, Mathias M, Andrade R, Bastos R, Balduino A, Schott V, Rodeo S, Espregueira-Mendes J. Intra-articular injections of expanded mesenchymal stem cells with and without addition of platelet-rich plasma are safe and effective for knee osteoarthritis. Knee Surg Sports Traumatol Arthroscopy. 2018;26(11):3342–50.
Lamo-Espinosa JM, Mora G, Blanco JF, Granero-Molto F, Nunez-Cordoba JM, Lopez-Elio S, Andreu E, Sanchez-Guijo F, Aquerreta JD, Bondia JM, et al. Intra-articular injection of two different doses of autologous bone marrow mesenchymal stem cells versus hyaluronic acid in the treatment of knee osteoarthritis: long-term follow up of a multicenter randomized controlled clinical trial (phase I/II). J Transl Med. 2018;16(1):213.
Pers YM, Rackwitz L, Ferreira R, Pullig O, Delfour C, Barry F, Sensebe L, Casteilla L, Fleury S, Bourin P, et al. Adipose mesenchymal stromal cell-based therapy for severe osteoarthritis of the knee: a phase I dose-escalation trial. Stem Cells Transl Med. 2016;5(7):847–56.
Wang Y, Jin W, Liu H, Cui Y, Mao Q, Fei Z, Xiang C. Curative effect of human umbilical cord mesenchymal stem cells by intra-articular injection for degenerative knee osteoarthritis. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2016;30(12):1472–7.
ter Huurne M, Schelbergen R, Blattes R, Blom A, de Munter W, Grevers LC, Jeanson J, Noël D, Casteilla L, Jorgensen C. Antiinflammatory and chondroprotective effects of intraarticular injection of adipose-derived stem cells in experimental osteoarthritis. Arthritis Rheumatism. 2012;64(11):3604–13.
Toupet K, Maumus M, Luz-Crawford P, Lombardo E, Lopez-Belmonte J, Van Lent P, Garin MI, Van den Berg W, Dalemans W, Jorgensen C. Survival and biodistribution of xenogenic adipose mesenchymal stem cells is not affected by the degree of inflammation in arthritis. PLoS One. 2015;10(1):e0114962.
Murphy JM, Fink DJ, Hunziker EB, Barry FP. Stem cell therapy in a caprine model of osteoarthritis. Arthritis Rheumatism. 2003;48(12):3464–74.
Desando G, Cavallo C, Sartoni F, Martini L, Parrilli A, Veronesi F, Fini M, Giardino R, Facchini A, Grigolo B. Intra-articular delivery of adipose derived stromal cells attenuates osteoarthritis progression in an experimental rabbit model. Arthritis Res Ther. 2013;15(1):R22.
Soler R, Orozco L, Munar A, Huguet M, Lopez R, Vives J, Coll R, Codinach M, Garcia-Lopez J. Final results of a phase I-II trial using ex vivo expanded autologous mesenchymal stromal cells for the treatment of osteoarthritis of the knee confirming safety and suggesting cartilage regeneration. Knee. 2016;23(4):647–54.
Matas J, Orrego M, Amenabar D, Infante C, Tapia-Limonchi R, Cadiz MI, Alcayaga-Miranda F, Gonzalez PL, Muse E, Khoury M, et al. Umbilical cord-derived mesenchymal stromal cells (MSCs) for knee osteoarthritis: repeated MSC dosing is superior to a single MSC dose and to hyaluronic acid in a controlled randomized phase I/II trial. Stem Cells Transl Med. 2019;8(3):215-224.
Maumus M, Manferdini C, Toupet K, Peyrafitte J-A, Ferreira R, Facchini A, Gabusi E, Bourin P, Jorgensen C, Lisignoli G. Adipose mesenchymal stem cells protect chondrocytes from degeneration associated with osteoarthritis. Stem Cell Res. 2013;11(2):834–44.
Manferdini C, Maumus M, Gabusi E, Piacentini A, Filardo G, Peyrafitte JA, Jorgensen C, Bourin P, Fleury-Cappellesso S, Facchini A. Adipose-derived mesenchymal stem cells exert antiinflammatory effects on chondrocytes and synoviocytes from osteoarthritis patients through prostaglandin E2. Arthritis Rheumatism. 2013;65(5):1271–81.
Patel JM, Saleh KS, Burdick JA, Mauck RL. Bioactive factors for cartilage repair and regeneration: improving delivery, retention, and activity. Acta Biomater. 2019;93:222–38.
Anderson HC, Mulhall D, Garimella R. Role of extracellular membrane vesicles in the pathogenesis of various diseases, including cancer, renal diseases, atherosclerosis, and arthritis. Lab Investig. 2010;90(11):1549.
Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neuro-Oncol. 2013;113(1):1–11.
Toh WS, Foldager CB, Pei M, Hui JHP. Advances in mesenchymal stem cell-based strategies for cartilage repair and regeneration. Stem Cell Rev Rep. 2014;10(5):686–96.
da Silva ML, Fontes AM, Covas DT, Caplan AI. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009;20(5–6):419–27.
Toh WS, Lai RC, Hui JHP, Lim SK. MSC exosome as a cell-free MSC therapy for cartilage regeneration: implications for osteoarthritis treatment. Semin Cell Dev Biol. 2017;67:56-64.
Lai RC, Arslan F, Lee MM, Sze NSK, Choo A, Chen TS, Salto-Tellez M, Timmers L, Lee CN, El Oakley RM. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010;4(3):214–22.
Zhang S, Chu W, Lai R, Lim S, Hui J, Toh W. Exosomes derived from human embryonic mesenchymal stem cells promote osteochondral regeneration. Osteoarthr Cartil. 2016;24(12):2135–40.
Vonk LA, van Dooremalen SF, Liv N, Klumperman J, Coffer PJ, Saris DB, Lorenowicz MJ. Mesenchymal stromal/stem cell-derived extracellular vesicles promote human cartilage regeneration in vitro. Theranostics. 2018;8(4):906.
Zhang S, Chuah SJ, Lai RC, Hui JHP, Lim SK, Toh WS. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials. 2018;156:16–27.
Andaloussi SE, Mäger I, Breakefield XO, Wood MJ. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov. 2013;12(5):347.
Xin H, Li Y, Chopp M. Exosomes/miRNAs as mediating cell-based therapy of stroke. Front Cell Neurosci. 2014;8:377.
Burger D, Viñas JL, Akbari S, Dehak H, Knoll W, Gutsol A, Carter A, Touyz RM, Allan DS, Burns KD. Human endothelial colony-forming cells protect against acute kidney injury: role of exosomes. Am J Pathol. 2015;185(8):2309–23.
Mancuso P, Raman S, Glynn A, Barry F, Murphy JM. Mesenchymal stem cell therapy for osteoarthritis: the critical role of the cell secretome. Front Bioeng Biotechnol. 2019;7:9.
Wang Y, Yu D, Liu Z, Zhou F, Dai J, Wu B, Zhou J, Heng BC, Zou XH, Ouyang H, et al. Exosomes from embryonic mesenchymal stem cells alleviate osteoarthritis through balancing synthesis and degradation of cartilage extracellular matrix. Stem Cell Res Ther. 2017;8(1):189.
Mao G, Zhang Z, Hu S, Zhang Z, Chang Z, Huang Z, Liao W, Kang Y. Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res Ther. 2018;9(1):247.
Zhu Y, Wang Y, Zhao B, Niu X, Hu B, Li Q, Zhang J, Ding J, Chen Y, Wang Y. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res Ther. 2017;8(1):64.
Tao S-C, Yuan T, Zhang Y-L, Yin W-J, Guo S-C, Zhang C-Q. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics. 2017;7(1):180.
Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–83.
Becker A, Thakur BK, Weiss JM, Kim HS, Peinado H, Lyden D. Extracellular vesicles in cancer: cell-to-cell mediators of metastasis. Cancer Cell. 2016;30(6):836–48.
Chang Y-H, Wu K-C, Harn H-J, Lin S-Z, Ding D-C. Exosomes and stem cells in degenerative disease diagnosis and therapy. Cell Transplant. 2018;27(3):349–63.
Kowal J, Tkach M, Théry C. Biogenesis and secretion of exosomes. Curr Opin Cell Biol. 2014;29:116–25.
Christ L, Raiborg C, Wenzel EM, Campsteijn C, Stenmark H. Cellular functions and molecular mechanisms of the ESCRT membrane-scission machinery. Trends Biochem Sci. 2017;42(1):42–56.
Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, Wieland F, Schwille P, Brügger B, Simons M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319(5867):1244–7.
Mercier V, Laporte MH, Destaing O, Blot B, Blouin CM, Pernet-Gallay K, Chatellard C, Saoudi Y, Albiges-Rizo C, Lamaze C. ALG-2 interacting protein-X (Alix) is essential for clathrin-independent endocytosis and signaling. Sci Rep. 2016;6:26986.
Kajimoto T, Okada T, Miya S, Zhang L, Nakamura S-I. Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes. Nat Commun. 2013;4:2712.
Pefanis E, Wang J, Rothschild G, Lim J, Kazadi D, Sun J, Federation A, Chao J, Elliott O, Liu Z-P. RNA exosome-regulated long non-coding RNA transcription controls super-enhancer activity. Cell. 2015;161(4):774–89.
Thakur BK, Zhang H, Becker A, Matei I, Huang Y, Costa-Silva B, Zheng Y, Hoshino A, Brazier H, Xiang J. Double-stranded DNA in exosomes: a novel biomarker in cancer detection. Cell Res. 2014;24(6):766.
Kahlert C, Melo SA, Protopopov A, Tang J, Seth S, Koch M, Zhang J, Weitz J, Chin L, Futreal A. Identification of double-stranded genomic DNA spanning all chromosomes with mutated KRAS and p53 DNA in the serum exosomes of patients with pancreatic cancer. J Biol Chem. 2014;289(7):3869–75.
Skotland T, Sandvig K, Llorente A. Lipids in exosomes: current knowledge and the way forward. Prog Lipid Res. 2017;66:30–41.
Zhang H, Freitas D, Kim HS, Fabijanic K, Li Z, Chen H, Mark MT, Molina H, Martin AB, Bojmar L. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat Cell Biol. 2018;20(3):332.
Willms E, Johansson HJ, Mäger I, Lee Y, Blomberg KEM, Sadik M, Alaarg A, Smith CE, Lehtiö J, Andaloussi SE. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci Rep. 2016;6:22519.
Tkach M, Thery C. Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016;164(6):1226–32.
Li JJ, Hosseini-Beheshti E, Grau GE, Zreiqat H, Little CB. Stem cell-derived extracellular vesicles for treating joint injury and osteoarthritis. Nanomaterials (Basel). 2019;9(2). https://doi.org/10.3390/nano9020261.
Withrow J, Murphy C, Liu Y, Hunter M, Fulzele S, Hamrick MW. Extracellular vesicles in the pathogenesis of rheumatoid arthritis and osteoarthritis. Arthritis Res Ther. 2016;18(1):286.
Li Z, Wang Y, Xiao K, Xiang S, Li Z, Weng X. Emerging role of exosomes in the joint diseases. Cell Physiol Biochem. 2018;47(5):2008–17.
Maumus M, Jorgensen C, Noel D. Mesenchymal stem cells in regenerative medicine applied to rheumatic diseases: role of secretome and exosomes. Biochimie. 2013;95(12):2229–34.
Domenis R, Zanutel R, Caponnetto F, Toffoletto B, Cifù A, Pistis C, Di Benedetto P, Causero A, Pozzi M, Bassini F. Characterization of the proinflammatory profile of synovial fluid-derived exosomes of patients with osteoarthritis. Mediators Inflamm. 2017;2017:4814987.
Kolhe R, Hunter M, Liu S, Jadeja RN, Pundkar C, Mondal AK, Mendhe B, Drewry M, Rojiani MV, Liu Y. Gender-specific differential expression of exosomal miRNA in synovial fluid of patients with osteoarthritis. Sci Rep. 2017;7(1):2029.
Kato T, Miyaki S, Ishitobi H, Nakamura Y, Nakasa T, Lotz MK, Ochi M. Exosomes from IL-1β stimulated synovial fibroblasts induce osteoarthritic changes in articular chondrocytes. Arthritis Res Ther. 2014;16(4):R163.
Zhang H-G, Liu C, Su K, Yu S, Zhang L, Zhang S, Wang J, Cao X, Grizzle W, Kimberly RP. A membrane form of TNF-α presented by exosomes delays T cell activation-induced cell death. J Immunol. 2006;176(12):7385–93.
Kanehisa M, Goto S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000;28(1):27–30.
Kanehisa M, Goto S, Furumichi M, Tanabe M, Hirakawa M. KEGG for representation and analysis of molecular networks involving diseases and drugs. Nucleic Acids Res. 2009;38(suppl_1):D355–60.
Skriner K, Adolph K, Jungblut PR, Burmester GR. Association of citrullinated proteins with synovial exosomes. Arthritis Rheumatism. 2006;54(12):3809–14.
Cloutier N, Tan S, Boudreau LH, Cramb C, Subbaiah R, Lahey L, Albert A, Shnayder R, Gobezie R, Nigrovic PA, et al. The exposure of autoantigens by microparticles underlies the formation of potent inflammatory components: the microparticle-associated immune complexes. EMBO Mol Med. 2013;5(2):235–49.
Yoo J, Lee SK, Lim M, Sheen D, Choi E-H, Kim SA. Exosomal amyloid A and lymphatic vessel endothelial hyaluronic acid receptor-1 proteins are associated with disease activity in rheumatoid arthritis. Arthritis Res Ther. 2017;19(1):119.
Kim SH, Lechman ER, Bianco N, Menon R, Keravala A, Nash J, Mi Z, Watkins SC, Gambotto A, Robbins PD. Exosomes derived from IL-10-treated dendritic cells can suppress inflammation and collagen-induced arthritis. J Immunol. 2005;174(10):6440–8.
Kim SH, Bianco NR, Shufesky WJ, Morelli AE, Robbins PD. Effective treatment of inflammatory disease models with exosomes derived from dendritic cells genetically modified to express IL-4. J Immunol. 2007;179(4):2242–9.
Bianco NR, Kim SH, Ruffner MA, Robbins PD. Therapeutic effect of exosomes from indoleamine 2,3-dioxygenase-positive dendritic cells in collagen-induced arthritis and delayed-type hypersensitivity disease models. Arthritis Rheum. 2009;60(2):380–9.
Marton N, Kovács OT, Baricza E, Kittel Á, Győri D, Mócsai A, Meier FM, Goodyear CS, McInnes IB, Buzás EI. Extracellular vesicles regulate the human osteoclastogenesis: divergent roles in discrete inflammatory arthropathies. Cell Mol Life Sci. 2017;74(19):3599–611.
Liu B, Wu P, Mei L, Luo Y, Li H, Mao X. Differential expression of exosomal miRNAs in osteoblasts in osteoarthritis. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2018;43(12):1294–300.
Wang L, Wang C, Jia X, Yu J. Circulating exosomal miR-17 inhibits the induction of regulatory T cells via suppressing TGFBR II expression in rheumatoid arthritis. Cell Physiol Biochem. 2018;50(5):1754–63.
Xu D, Song M, Chai C, Wang J, Jin C, Wang X, Cheng M, Yan S. Exosome-encapsulated miR-6089 regulates inflammatory response via targeting TLR4. J Cell Physiol. 2019;234(2):1502–11.
Wang Y, Zheng F, Gao G, Yan S, Zhang L, Wang L, Cai X, Wang X, Xu D, Wang J. MiR-548a-3p regulates inflammatory response via TLR4/NF-κB signaling pathway in rheumatoid arthritis. J Cell Biochem. 2019;120(2):1133–40.
Maeda Y, Farina NH, Matzelle MM, Fanning PJ, Lian JB, Gravallese EM. Synovium-derived micrornas regulate bone pathways in rheumatoid arthritis. J Bone Miner Res. 2017;32(3):461–72.
Zhao Y, Xu J. Synovial fluid-derived exosomal lncRNA PCGEM1 as biomarker for the different stages of osteoarthritis. Int Orthop. 2018;42(12):2865–72.
Song J, Kim D, Han J, Kim Y, Lee M, Jin E-J. PBMC and exosome-derived Hotair is a critical regulator and potent marker for rheumatoid arthritis. Clin Exp Med. 2015;15(1):121–6.
Davatchi F, Sadeghi Abdollahi B, Mohyeddin M, Nikbin B. Mesenchymal stem cell therapy for knee osteoarthritis: 5 years follow-up of three patients. Int J Rheum Dis. 2016;19(3):219–25.
Lamo-Espinosa JM, Mora G, Blanco JF, Granero-Molto F, Nunez-Cordoba JM, Sanchez-Echenique C, Bondia JM, Aquerreta JD, Andreu EJ, Ornilla E, et al. Intra-articular injection of two different doses of autologous bone marrow mesenchymal stem cells versus hyaluronic acid in the treatment of knee osteoarthritis: multicenter randomized controlled clinical trial (phase I/II). J Transl Med. 2016;14(1):246.
Vega A, Martin-Ferrero MA, Del Canto F, Alberca M, Garcia V, Munar A, Orozco L, Soler R, Fuertes JJ, Huguet M, et al. Treatment of knee osteoarthritis with allogeneic bone marrow mesenchymal stem cells: a randomized controlled trial. Transplantation. 2015;99(8):1681–90.
Ham O, Lee C, Kim R, Lee J, Oh S, Lee M, Kim J, Hwang K-C, Maeng L-S, Chang W. Therapeutic potential of differentiated mesenchymal stem cells for treatment of osteoarthritis. Int J Mol Sci. 2015;16(7):14961–78.
Qi Y, Feng G, Yan W. Mesenchymal stem cell-based treatment for cartilage defects in osteoarthritis. Mol Biol Rep. 2012;39(5):5683–9.
Koizumi K, Ebina K, Hart D, Hirao M, Noguchi T, Sugita N, Yasui Y, Chijimatsu R, Yoshikawa H, Nakamura N. Synovial mesenchymal stem cells from osteo-or rheumatoid arthritis joints exhibit good potential for cartilage repair using a scaffold-free tissue engineering approach. Osteoarthr Cartil. 2016;24(8):1413–22.
Van Buul G, Villafuertes E, Bos P, Waarsing J, Kops N, Narcisi R, Weinans H, Verhaar J, Bernsen M, Van Osch G. Mesenchymal stem cells secrete factors that inhibit inflammatory processes in short-term osteoarthritic synovium and cartilage explant culture. Osteoarthr Cartil. 2012;20(10):1186–96.
Mendicino M, Bailey AM, Wonnacott K, Puri RK, Bauer SR. MSC-based product characterization for clinical trials: an FDA perspective. Cell Stem Cell. 2014;14(2):141–5.
Lee WY-w, Wang B. Cartilage repair by mesenchymal stem cells: clinical trial update and perspectives. J Orthop Transl. 2017;9:76–88.
Chahla J, Piuzzi NS, Mitchell JJ, Dean CS, Pascual-Garrido C, LaPrade RF, Muschler GF. Intra-articular cellular therapy for osteoarthritis and focal cartilage defects of the knee: a systematic review of the literature and study quality analysis. JBJS. 2016;98(18):1511–21.
McIntyre JA, Jones IA, Han B, Vangsness CT Jr. Intra-articular mesenchymal stem cell therapy for the human joint: a systematic review. Am J Sports Med. 2018;46(14):3550–63.
Yubo M, Yanyan L, Li L, Tao S, Bo L, Lin C. Clinical efficacy and safety of mesenchymal stem cell transplantation for osteoarthritis treatment: a meta-analysis. PLoS One. 2017;12(4):e0175449.
Wyles CC, Houdek MT, Behfar A, Sierra RJ. Mesenchymal stem cell therapy for osteoarthritis: current perspectives. Stem Cells Cloning. 2015;8:117.
Fu Y, Karbaat L, Wu L, Leijten J, Both SK, Karperien M. Trophic effects of mesenchymal stem cells in tissue regeneration. Tissue Eng B Rev. 2017;23(6):515–28.
Liu-Bryan R, Terkeltaub R. Emerging regulators of the inflammatory process in osteoarthritis. Nat Rev Rheumatol. 2015;11(1):35–44.
Ruiz M, Cosenza S, Maumus M, Jorgensen C, Noël D. Therapeutic application of mesenchymal stem cells in osteoarthritis. Expert Opin Biol Ther. 2016;16(1):33–42.
Richards MM, Maxwell JS, Weng L, Angelos MG, Golzarian J. Intra-articular treatment of knee osteoarthritis: from anti-inflammatories to products of regenerative medicine. Phys Sportsmed. 2016;44(2):101–8.
Murphy MB, Moncivais K, Caplan AI. Mesenchymal stem cells: environmentally responsive therapeutics for regenerative medicine. Exp Mol Med. 2013;45(11):e54.
Heldring N, Mäger I, Wood MJ, Le Blanc K, Andaloussi SE. Therapeutic potential of multipotent mesenchymal stromal cells and their extracellular vesicles. Hum Gene Ther. 2015;26(8):506–17.
Siddappa R, Licht R, van Blitterswijk C, de Boer J. Donor variation and loss of multipotency during in vitro expansion of human mesenchymal stem cells for bone tissue engineering. J Orthop Res. 2007;25(8):1029–41.
Dickhut A, Pelttari K, Janicki P, Wagner W, Eckstein V, Egermann M, Richter W. Calcification or dedifferentiation: requirement to lock mesenchymal stem cells in a desired differentiation stage. J Cell Physiol. 2009;219(1):219–26.
Lee MJ, Kim J, Kim MY, Bae Y-S, Ryu SH, Lee TG, Kim JH. Proteomic analysis of tumor necrosis factor-α-induced secretome of human adipose tissue-derived mesenchymal stem cells. J Proteome Res. 2010;9(4):1754–62.
Wolf P. The nature and significance of platelet products in human plasma. Br J Haematol. 1967;13(3):269–88.
Yáñez-Mó M, Siljander PR-M, Andreu Z, Bedina Zavec A, Borràs FE, Buzas EI, Buzas K, Casal E, Cappello F, Carvalho J. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4(1):27066.
Hosseini-Beheshti E, Grau GE. Extracellular vesicles and microvascular pathology: Decoding the active dialogue. Microcirculation. 2019;26(2):e12485.
Lener T, Gimona M, Aigner L, Börger V, Buzas E, Camussi G, Chaput N, Chatterjee D, Court FA, HAd P. Applying extracellular vesicles based therapeutics in clinical trials–an ISEV position paper. J Extracell Vesicles. 2015;4(1):30087.
Sun H, Hu S, Zhang Z, Lun J, Liao W, Zhang Z. Expression of exosomal microRNAs during chondrogenic differentiation of human bone mesenchymal stem cells. J Cell Biochem. 2019;120(1):171–81.
Chen Z, Wang H, Xia Y. Therapeutic potential of mesenchymal cell-derived miRNA-150-5p-expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J Immunol. 2018;201(8):2472-2482.
Zhang X, Liu J, Zhang P, Dai L, Wu Z, Wang L, Cao M, Jiang J. Silibinin induces G1 arrest, apoptosis and JNK/SAPK upregulation in SW1990 human pancreatic cancer cells. Oncol Lett. 2018;15(6):9868–76.
Qi H, Liu DP, Xiao DW, Tian DC, Su YW, Jin SF. Exosomes derived from mesenchymal stem cells inhibit mitochondrial dysfunction-induced apoptosis of chondrocytes via p38, ERK, and Akt pathways. In vitro Cell Dev Biol Anim. 2019;55(3):203–10.
Zhang S, Teo KYW, Chuah SJ, Lai RC, Lim SK, Toh WS. MSC exosomes alleviate temporomandibular joint osteoarthritis by attenuating inflammation and restoring matrix homeostasis. Biomaterials. 2019;200:35–47.
Tofino-Vian M, Guillen MI, Perez Del Caz MD, Silvestre A, Alcaraz MJ. Microvesicles from human adipose tissue-derived mesenchymal stem cells as a new protective strategy in osteoarthritic chondrocytes. Cell Physiol Biochem. 2018;47(1):11–25.
Tofiño-Vian M, Guillén MI, del Caz P, Dolores M, Castejón MA, Alcaraz MJ. Extracellular vesicles from adipose-derived mesenchymal stem cells downregulate senescence features in osteoarthritic osteoblasts. Oxid Med Cell Longev. 2017;2017:7197598.
Cosenza S, Toupet K, Maumus M, Luz-Crawford P, Blanc-Brude O, Jorgensen C, Noel D. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics. 2018;8(5):1399–410.
Casado JG, Blázquez R, Vela FJ, Álvarez V, Tarazona R, Sánchez-Margallo FM. Mesenchymal stem cell-derived exosomes: immunomodulatory evaluation in an antigen-induced synovitis porcine model. Front Vet Sci. 2017;4:39.
He X. Exosomes and Regenerative Medicine: State of the art and Perspectives. Transl Res. 2018;196:1-16.
Reiner AT, Witwer KW, van Balkom BWM, de Beer J, Brodie C, Corteling RL, Gabrielsson S, Gimona M, Ibrahim AG, de Kleijn D, et al. Concise review: developing best-practice models for the therapeutic use of extracellular vesicles. Stem Cells Transl Med. 2017;6(8):1730–9.
Zhang B, Yeo RW, Tan KH, Lim SK. Focus on extracellular vesicles: therapeutic potential of stem cell-derived extracellular vesicles. Int J Mol Sci. 2016;17(2):174.
Chen TS, Lai RC, Lee MM, Choo AB, Lee CN, Lim SK. Mesenchymal stem cell secretes microparticles enriched in pre-microRNAs. Nucleic Acids Res. 2010;38(1):215–24.
Lai RC, Tan SS, Teh BJ, Sze SK, Arslan F, de Kleijn DP, Choo A, Lim SK. Proteolytic potential of the MSC exosome proteome: implications for an exosome-mediated delivery of therapeutic proteasome. Int J Proteomics. 2012;2012:971907.
Borger V, Bremer M, Ferrer-Tur R, Gockeln L, Stambouli O, Becic A, Giebel B. Mesenchymal stem/stromal cell-derived extracellular vesicles and their potential as novel immunomodulatory therapeutic agents. Int J Mol Sci. 2017;18(7). https://doi.org/10.3390/ijms18071450.
Baglio SR, Rooijers K, Koppers-Lalic D, Verweij FJ, Perez Lanzon M, Zini N, Naaijkens B, Perut F, Niessen HW, Baldini N, et al. Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res Ther. 2015;6:127.
Wang K, Jiang Z, Webster KA, Chen J, Hu H, Zhou Y, Zhao J, Wang L, Wang Y, Zhong Z. Enhanced cardioprotection by human endometrium mesenchymal stem cells driven by exosomal microRNA-21. Stem Cells Transl Med. 2017;6(1):209–22.
Wu Y, Peng Y, Gao D, Feng C, Yuan X, Li H, Wang Y, Yang L, Huang S, Fu X. Mesenchymal stem cells suppress fibroblast proliferation and reduce skin fibrosis through a TGF-beta3-dependent activation. Int J Low Extrem Wounds. 2015;14(1):50–62.
Acknowledgements
Not applicable.
Funding
MRH was supported by US NIH Grants R01AI050875 and R21AI121700.
Author information
Authors and Affiliations
Contributions
HM and MRH contributed in conception, design, statistical analysis, and drafting of the manuscript. EM, HRM, MMT, AR, RS, and M-HP contributed in data collection and manuscript drafting. All authors approved the final version for submission.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was considered exempt by the KAUMS Institutional Review Board.
Consent for publication
Not applicable.
Competing interests
MRH declares the following potential conflicts of interest. Scientific Advisory Boards: Transdermal Cap Inc., Cleveland, OH; BeWell Global Inc., Wan Chai, Hong Kong; Hologenix Inc. Santa Monica, CA; LumiThera Inc., Poulsbo, WA; Vielight, Toronto, Canada; Bright Photomedicine, Sao Paulo, Brazil; Quantum Dynamics LLC, Cambridge, MA; Global Photon Inc., Bee Cave, TX; Medical Coherence, Boston MA; NeuroThera, Newark DE; JOOVV Inc., Minneapolis-St. Paul MN; AIRx Medical, Pleasanton CA; FIR Industries, Inc. Ramsey, NJ; UVLRx Therapeutics, Oldsmar, FL; Ultralux UV Inc., Lansing MI; Illumiheal & Petthera, Shoreline, WA; MB Lasertherapy, Houston, TX; ARRC LED, San Clemente, CA; Varuna Biomedical Corp. Incline Village, NV; Niraxx Light Therapeutics, Inc., Boston, MA. Consulting; Lexington Int, Boca Raton, FL; USHIO Corp, Japan; Merck KGaA, Darmstadt, Germany; Philips Electronics Nederland B.V. Eindhoven, Netherlands; Johnson & Johnson Inc., Philadelphia, PA; Sanofi-Aventis Deutschland GmbH, Frankfurt am Main, Germany. Stockholdings: Global Photon Inc., Bee Cave, TX; Mitonix, Newark, DE.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Mianehsaz, E., Mirzaei, H.R., Mahjoubin-Tehran, M. et al. Mesenchymal stem cell-derived exosomes: a new therapeutic approach to osteoarthritis?. Stem Cell Res Ther 10, 340 (2019). https://doi.org/10.1186/s13287-019-1445-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13287-019-1445-0