
Abstract
Background
The clinical presentation of severe acute respiratory syndrome coronavirus-2 infection is highly variable from asymptomatic infection to fulminant disease. The reasons for the variation are only starting to unravel, with risk factors including age and certain comorbidities as well as genetic defects causing immunological perturbations in the interferon pathways.
Case presentation
We report the case of an otherwise healthy Caucasian man, who at ages 60 and 64 years suffered from severe H1N1 influenza virus infection and severe acute respiratory syndrome coronavirus-2 infections, respectively. In both cases, there were acute kidney impairment and the need for intensive care unit admission as well as mechanical ventilation. Fortunately, after both infections there was full clinical recovery. The severity of the infections indicates an underlying impairment in the ability to control these kinds of infections. Challenge of patient peripheral blood mononuclear cells showed impaired type I and III antiviral interferon responses and reduced interferon-stimulated gene expression. However, despite investigation of patient samples by whole exome sequencing and enzyme-linked immunosorbent assay, no known disease-causing genetic variants related to interferon pathways were found, nor were interferon autoantibodies demonstrated. Thus, any underlying immunological cause of this unusual susceptibility to severe viral infections remains unresolved.
Conclusion
The patient experienced very similar severe clinical pictures triggered by H1N1 and severe acute respiratory syndrome coronavirus-2 infections, indicating an underlying inability to contain these infections. We were unable to show that the patient had any of the currently known types of immune incompetence but identified genetic changes possibly contributing to the severe course of both infections. Further analyses to delineate contribution factors are needed.
Similar content being viewed by others

Introduction
The clinical course of coronavirus disease 2019 (COVID-19) is highly variable. The reasons behind this are only now starting to be elucidated. Several clinical risk factors for severe COVID-19 have been identified, including older age, male sex, and various comorbidities [1, 2]. Furthermore, it has recently been shown that genetic defects in the toll-like receptor (TLR)-3 and -7 dependent type I interferon (IFN) pathway as well as IFN autoantibodies are highly enriched among patients with life-threatening COVID-19 [3,4,5]. Here, we report an otherwise healthy man who, with an interval of 4 years, suffered first fulminant H1N1 and later severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infections, with full recovery after both, indicating an underlying weakness in the defense against these types of viral infections. Severe COVID-19 among individuals with prior fulminant viral infection(s) has, to our knowledge, not previously been reported.
Case description
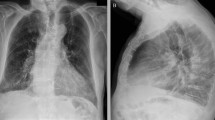
A 64-year-old Caucasian man was admitted to a hospital in Copenhagen, Denmark with hematuria on 7 March 2020. Within one day of admission, he became febrile with respiratory symptoms interpreted as nosocomial pneumonia. Four days later, the patient was discharged with moxifloxacin scheduled for a follow-up transurethral prostatectomy. The patient was, however, readmitted with worsening respiratory symptoms on 13 March. Only then was he tested for SARS-CoV-2 by polymerase chain reaction (PCR) and tested positive. At admission, the patient had elevated inflammation markers: C-reactive protein of 231 mg/L (< 10 mg/L), slight neutrophilic leukocytosis of 8,7/10,3 × 109/L (3.5–8.8 × 109/L), and elevated lactate dehydrogenase of 526 U/L (105–205 U/L). Procalcitonin was not measured at admission but was elevated at 2.31 (< 50 μg/L) 4 days later. X-ray on 14 March showed bilateral infiltrates (Fig. 1). Due to respiratory deterioration, the patient was soon after transferred to the intensive care unit (ICU) with a sequential organ failure assessment (SOFA) score of 4 and an O2 requirement of 15 L. He rapidly deteriorated with hypoxic respiratory failure and needed mechanical ventilation (Drager V500, PS). One day later, extracorporeal membrane oxygenation (ECMO) was initiated and maintained for 6 days; during ECMO treatment the patient was heparinized. Subsequently, the patient was again placed on mechanical ventilation (highest p plateau 14 cm H2O, PS) for 3 days. After extubation, the O2 requirement steadily decreased, and no oxygen treatment was needed for the last 3 days of hospitalization. Neither steroids nor tocilizumab were given, as these treatments were not implemented as part of COVID-19 treatment at the time. Total duration of stay at the ICU and hospital was 12 and 24 days, respectively. During the COVID-19 infection, the patient developed kidney impairment with a maximum creatinine of 302 (reference 60–105) µmol/L. Within the following months, the patient recovered completely and was able to resume work full-time. Five months post-COVID-19, the lung diffusion capacity for carbon monoxide (DLCO) was mildly impaired (64% of predicted), while ventilation capacity was normal (forced expiratory volume in the first second 120% predicted, forced vital capacity 112% predicted, and total lung capacity 90% predicted) and respiratory muscle strength was normal (maximum expiratory and inspiratory pressure 72% and 100% predicted, respectively).

Chest X-ray obtained 1 day after readmission and diagnosis of SARS CoV-2 infection
Interestingly, 4 years prior, the patient had experienced a very similar clinical and radiological picture (Table 2, Fig. 2a) due to an H1N1 infection requiring 5 weeks of hospital admission including ICU stay, mechanical ventilation, and dialysis. The patient had in the intervening period been without significant health problems except for known prostate hypertrophy and palpitations and marginally increased blood pressure treated with beta blockers. A CT scan had been performed at a follow-up post-H1N1 outpatient visit (Table 1). The findings included ground-glass opacities (GGO), reticulations, and traction bronchiectasis with an upper region dominance (Fig. 2A). No further scans were available prior to the patient being diagnosed with COVID-19. A CT scan 1 month after onset of COVID-19 revealed a large extent of GGO in both upper and lower regions. There was volume loss and bronchiectasis, all suggesting ongoing infection and inflammation with a component of organizing pneumonia (Fig. 2B). Five months after onset of COVID-19 symptoms, there was marked improvement, almost to the level of post-H1N1 infection (Fig. 2C).

A CT scan at the end of the H1N1 infection with ground-glass opacities, reticulations, and traction bronchiectasis with upper region dominance. B CT scan 1 month after onset of COVID-19 infection showing a large extent of ground-glass opacities in both upper and lower regions, with a sparring of the subpleural regions. There is volume loss and bronchiectasis, all suggesting ongoing infection and inflammation with a component of organizing pneumonia. C CT scan 5 months after COVID-19 onset with almost normalization to the level of post-H1N1 infection
Severe infection with H1N1 and SARS-CoV-2 have both been associated with genetic variants impairing the signaling pathways inducing type I IFN [3, 4]. We therefore investigated the patient’s ability to mount an innate immune response upon viral infection. PBMCs from P1 and healthy controls were infected with SARS-CoV-2 at a multiplicity of infection (MOI) of 0.5 for 24 hours, and the induction of IFNs and proinflammatory cytokines was measured in the supernatants. PBMCs from P1 demonstrated significantly impaired production of both type I and III IFNs (Fig. 3A–C), whereas the production of IFNγ was significantly increased compared with healthy controls (Fig. 3D). Patient PBMCs also exhibited significantly decreased CXCL10 levels in response to infection (Fig. 3E). Finally, of the three proinflammatory cytokines measured, only tumor necrosis factor alpha (TNFα) was significantly increased, as both interleukin 6 (IL-6) and IL-1β were expressed at levels comparable to the controls (Fig. 3F–H).

Cytokine responses in patient PBMCs upon SARS-CoV-2 infection. A–H Patient (P1) and control PBMCs (C1–5) were left untreated (UT) or infected with SARS-CoV-2 at a MOI of 0.5 for 24 hours. Induction of IFNs and proinflammatory cytokines was measured in supernatants by U-PLEX Meso Scale Technology. The experiment was performed once, but all stimulations were done in triplicate. Statistical differences were calculated using Mann–Whitney test. *p < 0.05, **p < 0.01, ns nonsignificant
Despite the deficient type I and III IFN responses, genetic testing by whole exome sequencing for the recently reported disease-causing genetic mutations within the IFN signaling pathway [3, 4] was negative (see below). Additional rare gene variants with a high prediction for deleteriousness/disease-causing potential are presented in Table 2.
Finally, presence of IFN autoantibodies has been demonstrated in plasma from up to 10% of cases with severe COVID-19 [4], thereby functionally impairing IFN activities, in the absence of disease-causing genetic variants. However, investigation for autoantibodies against IFNα or IFNω in patient plasma by ELISA did not reveal the presence of such autoantibodies (data not shown).
Discussion
We report the case of a man who, at ages 60 and 64 years, suffered severe H1N1 and SARS-CoV-2 infections, respectively, in both cases with acute kidney injury and requirement for mechanical ventilation. Both prior to the H1N1 infection and pre- and post-COVID-19, the patient was without respiratory complaints and, if any, only marginal, renal impairment. Lung function test 5 months post-COVID-19 was with slightly reduced DLCO at 64% of expected, while FEV1 and FVC were at 120% and 112% of predicted, respectively. Among the post-COVID-19 lung function test impairments most often reported is reduced DLCO [6, 7], in line with the findings in the current case. The radiological findings were also in line with those reported in the literature [8, 9].
The reason why this otherwise healthy man was so severely affected by these viral infections is currently undetermined. Infection of patient PBMCs with SARS-CoV-2 in vitro demonstrated significantly impaired type I and III IFN as well as CXCL10 responses, which may have resulted in impaired ability to clear the infection. Unfortunately, we were not able to couple the impaired responses with genetic variants in any of the IFN pathway susceptibility genes previously reported, including IRF3, IRF7, IRF9, TLR3, TRIF, TRAF3, UNC93, TBK1, IFNAR1, STAT1, and STAT2 [3]. We did, however, identify variants in other genes that could potentially be of relevance (Table 2), although these were not functionally validated, therefore leaving a potential link to the disease presentation uncertain.
Most likely, further inborn errors of immunity and/or acquired traits will be identified as explanations for the highly variable clinical course among individuals infected with SARS CoV-2.
The contribution to COVID-19 case severity impacted by various immunodeficiencies is largely unresolved. It has been debated whether certain immunodeficiencies might paradoxically protect against severe disease [10]. A British study found an overall negative effect of immunodeficiencies, with secondary immunodeficiencies resulting in worse clinical courses relative to primary immunodeficiencies [11]. Other studies have also found certain immunodeficiencies predisposing to more severe disease [12]. Further unraveling of the interplay between the various types of immune incompetence and clinical course of COVID-19 will most likely further our understanding of both the pathogenesis of SARS CoV-2 and the functioning of the various components of our immune system.
It is reassuring that even patients suffering from full-blown infections (in casu to H1N1 and SARS CoV-2) can fully recover. However, the requirement for ICU treatment for these individuals, for which access is limited or lacking in many countries highly affected by the current COVID-19 pandemic [13, 14], is worrisome. Hopefully, potent vaccines [15, 16] and treatments will soon be widely available, minimizing the need for ICU treatment.
Patient perspective
The patient reported satisfaction with the treatment received during admission for COVID-19 and during follow-up.
Conclusion
We report a case where a patient experienced very similar severe clinical pictures triggered by H1N1 and SARS-CoV-2 infections, indicating an underlying inability to contain these infections. We were unable to show that the patient had any of the currently known types of immune incompetence but identified genetic changes possibly contributing to the severe course of both infections.
Materials and methods
PBMC purification
PBMCs were purified from heparinized blood by Ficoll density centrifugation. Isolated PBMCs were stored in liquid nitrogen until further use. PBMCs were cultured in Roswell Park Memorial Institute (RPMI) medium with 10% heat-inactivated fetal bovine serum and 1% penicillin–streptomycin.
DNA isolation
DNA was isolated from ethylenediaminetetraacetic acid (EDTA)–blood using the QIAamp DNA Blood Mini Kit (Qiagen, 51104).
Whole exome sequencing analysis
Whole exome sequencing was performed as previously described [17]. Variants were kept only if minor allele frequency < 0.1%, CADD > 15. Loci associated with severe COVID-19 described by Zhang et al. [3] were included in the variant filtering.
SARS-CoV-2 infection
Patient and control PBMCs were seeded in 96-well plates (500,000 per well), rested overnight, and subsequently infected with SARS-CoV-2, strain FR-4286, at an MOI of 0.5 for 24 hours. Supernatants were harvested, inactivated with 0.4% Triton-X-100 at a ratio of 1:1 for 30 minutes, and stored at − 80 °C until further use.
Meso Scale
Cytokine expression was measured in supernatants from SARS-CoV-2-infected PBMCs by U-PLEX Meso Scale assays detecting IFNα2a, IFNβ, IFNγ, IFNλ1, TNFα, IL-1β, L-6, and CXCL10 (K15067L-2, MSD) according to the manufacturer’s instructions.
IFN autoantibodies
IFN autoantibodies were measured in plasma as previously described [4]. Briefly, ELISA plates were coated with 1 μg/mL IFNα (130-093-874, Miltenyi Biotec) or IFNω (BMS304, Invitrogen) overnight at 4 °C followed by blocking in 5% skimmed milk. Plasma samples were diluted 50× in high-performance ELISA (HPE) buffer (M1940, Sanquin) before incubation on the plates. Bound autoantibodies were detected with horseradish peroxidase (HRP)-conjugated goat anti-human IgG IgA IgM (GAHu/Ig(Fc/PO, Nordic-MUbio) and HRP substrate SureBlue KPL (5120-0077, SeraCare).
Statistics
Experiments were performed once due to limited patient material, but all stimulations were done in triplicate. Statistics were calculated using Mann–Whitney test.
Availability of data and materials
Data available to researchers upon request.
Abbreviations
- COVID-19:
-
Coronavirus disease 2019
- CT scan:
-
Computer tomography scan
- DLCO :
-
Lung diffusion capacity for carbon monoxide
- ECMO:
-
Extracorporeal membrane oxygenation
- FEV1:
-
Forced expiratory volume in the first second
- FVC:
-
Forced vital capacity
- GGO:
-
Ground-glass opacities
- H1N1:
-
Hemagglutinin 1, neuraminidase 1
- ICU:
-
Intensive care unit
- INF:
-
Interferon
- PBMC:
-
Peripheral blood mononuclear cells
- SARS-CoV-2:
-
Severe acute respiratory syndrome coronavirus 2
- TLR:
-
Toll-like receptor
References
Ruan Q, Yang K, Wang W, Jiang L, Song J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 2020;46(5):846–8.
Williamson EJ, Walker AJ, Bhaskaran K, Bacon S, Bates C, Morton CE, et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature. 2020;584(7821):430–6.
Zhang Q, Bastard P, Liu Z, Le PJ, Moncada-Velez M, Chen J, et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science. 2020;370:6515.
Bastard P, Rosen LB, Zhang Q, Michailidis E, Hoffmann HH, Zhang Y, et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science. 2020;370(6515):eabd4585.
van der Made CI, Simons A, Schuurs-Hoeijmakers J, van den Heuvel G, Mantere T, Kersten S, et al. Presence of genetic variants among young men with severe COVID-19. JAMA. 2020;324:663–73.
Daher A, Balfanz P, Cornelissen C, Muller A, Bergs I, Marx N, et al. Follow up of patients with severe coronavirus disease 2019 (COVID-19): pulmonary and extrapulmonary disease sequelae. Respir Med. 2020;174:106197.
Frija-Masson J, Debray MP, Gilbert M, Lescure FX, Travert F, Borie R, et al. Functional characteristics of patients with SARS-CoV-2 pneumonia at 30 days post-infection. Eur Respir J. 2020;56(2):167.
Zhao YM, Shang YM, Song WB, Li QQ, Xie H, Xu QF, et al. Follow-up study of the pulmonary function and related physiological characteristics of COVID-19 survivors 3 months after recovery. EClinicalMedicine. 2020;25:100463.
Xiong Y, Sun D, Liu Y, Fan Y, Zhao L, Li X, et al. Clinical and high-resolution CT features of the COVID-19 infection: comparison of the initial and follow-up changes. Invest Radiol. 2020;55(6):332–9.
Babaha F, Rezaei N. Primary immunodeficiency diseases in COVID-19 pandemic: a predisposing or protective factor? Am J Med Sci. 2020;360(6):740–1.
Shields AM, Burns SO, Savic S, Richter AG. COVID-19 in patients with primary and secondary immunodeficiency: the United Kingdom experience. J Allergy Clin Immunol. 2021;147(3):870–5.
Delavari S, Abolhassani H, Abolnezhadian F, Babaha F, Iranparast S, Ahanchian H, et al. Impact of SARS-CoV-2 pandemic on patients with primary immunodeficiency. J Clin Immunol. 2021;41(2):345–55.
Hopman J, Allegranzi B, Mehtar S. Managing COVID-19 in low- and middle-income countries. JAMA. 2020;323(16):1549–50.
Siow WT, Liew MF, Shrestha BR, Muchtar F, See KC. Managing COVID-19 in resource-limited settings: critical care considerations. Crit Care. 2020;24(1):167.
Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N Engl J Med. 2020;383:2603–15.
WHO. Coronavirus disease (COVID-19): vaccines. 2021.
Mardahl M, Jorgensen SE, Schneider A, Raaschou-Jensen K, Holm M, Veirum J, et al. Impaired immune responses to herpesviruses and microbial ligands in patients with MonoMAC. Br J Haematol. 2019;186(3):471–6.
Acknowledgements
No applicable.
Funding
No external funding was received.
Author information
Authors and Affiliations
Contributions
TLK and MH treated the patient for his COVID-19 and obtained data from the previous infection. TLK, MH, JM, AK, and THM decided on the analyses performed. AK and JM reviewed the CT scans. THM and SEJ performed the immunological analyses. TLK drafted the manuscript. All co-authors read and accepted the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The patients and healthy controls were included following oral and written consent and in accordance with the Helsinki Declaration and national ethics guidelines and after approval from the Danish National Committee on Health Ethics (1-10-72-80-20), the Data Protection Agency, and Institutional Review Board.
Consent for publication
Written informed consent was obtained from the patient for publication of this case report and any accompanying images. A copy of the written consent is available for review by the Editor-in-Chief of this journal.
Competing interests
None of the authors has any conflicts of interest to report.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Katzenstein, T.L., Jørgensen, S.E., Mortensen, J. et al. Fulminant H1N1 and severe acute respiratory syndrome coronavirus-2 infections with a 4-year interval without an identifiable underlying cause: a case report. J Med Case Reports 15, 505 (2021). https://doi.org/10.1186/s13256-021-03113-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13256-021-03113-9