
Abstract
Background
Epigenetic aberrations play a central role in the pathophysiology of acute myeloid leukemia (AML). It has been shown that molecular signatures based on DNA-methylation (DNAm) patterns can be used for classification of the disease. In this study, we followed the hypothesis that DNAm at a single CpG site might support risk stratification in AML.
Findings
Using DNAm profiles of 194 patients from The Cancer Genome Atlas (TCGA), we identified a CpG site in complement component 1 subcomponent R (C1R) as best suited biomarker: patients with higher methylation at this CpG site (>27 % DNAm) reveal significantly longer overall survival (53 versus 11 months; P < 0.0001). This finding was validated in an independent set of 62 DNAm profiles of cytogenetically normal AML patients (P = 0.009) and with a region-specific pyrosequencing assay in 84 AML samples (P = 0.012). DNAm of C1R correlated with genomic DNAm and gene expression patterns, whereas there was only moderate association with gene expression levels of C1R. These results indicate that DNAm of C1R is a biomarker reflecting chromatin reorganization rather than being of pathophysiological relevance per se. Notably, DNAm of C1R was associated with occurrence of specific genomic mutations that are traditionally used for risk stratification in AML. Furthermore, DNAm of C1R correlates also with overall survival in several other types of cancer, but the prognostic relevance was less pronounced than in AML.
Conclusions
Analysis of DNAm at C1R provides a simple, robust, and cost-effective biomarker to further complement risk assessment in AML.
Similar content being viewed by others


Findings
Risk assessment is relevant for the choice of therapeutic regimen in acute myeloid leukemia (AML). It is usually based on many parameters including age, white blood cell count, cytogenetic abnormalities, and specific mutations [1]. Epigenetic modifications, such as DNA-methylation (DNAm) changes, seem to play a critical role in pathogenesis of AML [2]. Various studies demonstrated that DNAm patterns can discriminate subgroups of patients with different clinical outcomes [3–7]. So far, these approaches utilized a combination of many differentially methylated regions and therefore require DNAm profiles based on microarray or sequencing technology. In contrast, a region-specific assay would be much faster, economic, and easier to interpret. We have recently described that aberrant hypermethylation at a specific region of the de novo methyltransferase DNMT3A is associated with poor prognosis in AML, but this association was only significant for patients without genomic mutations in DNMT3A because both modifications seem to have a similar molecular and clinical sequel [8]. This exemplifies that identification of simple epigenetic markers is hampered by the high molecular and clinical heterogeneity of the disease. Despite these hurdles, we followed the hypothesis that DNAm at a unique CpG site might provide a robust biomarker to further support risk assessment of AML.
Selection of CpG sites that are indicative for overall survival in AML
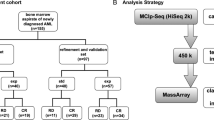
For initial selection of prognostic relevant CpGs, we utilized 194 DNAm profiles of AML patients from The Cancer Genome Atlas (TCGA) [9]. For each CpG site, the samples were stratified by its median DNAm level to estimate association with overall survival (OS) using the Kaplan-Meier (K-M) method (Fig. 1a). Only 60 CpGs revealed significant association upon adjustment for multiple testing (adjusted P < 0.05; Fig. 1b). Alternatively, we selected 418 CpGs with significant association with OS by COX-regression analysis (adjusted P < 0.05; Additional file 1: Figure S1). Only 26 CpGs were in the overlap of K-M and COX analysis (Additional file 1: Table S1). To narrow down to the best suited CpGs, we used the following additional criteria: (1) Sites should reveal relatively high variation in DNAm across different AML samples (as a higher DNAm range facilitates more reliable discrimination); (2) DNAm levels should be similar in different types of blood cells (to avoid bias by blood counts) [10]; and (3) there should be no correlation with age. In fact, many CpGs reveal age-associated DNAm changes [11, 12] and age is a major confounding factor for OS. Therefore, we excluded CpGs with clear DNAm changes upon aging. Using these parameters, we identified four candidate CpGs (Additional file 1: Table S1; Fig. 1c, d; Additional file 1: Figures S1–S3), which were tested in an independent dataset of Qu et al. [13]. Only one CpG site, positioned in the 5’UTR of complement component 1 subcomponent R (C1R, cg08799922), revealed significant association with OS (K-M analysis: P = 0.009; COX analysis: P = 0.012; Fig. 1e, f). We then designed a pyrosequencing assay for site-specific analysis at this genomic location (Additional file 1: Figure S4). Pyrosequencing results of 84 AML patients were stratified by the threshold of 27 % DNAm at cg08799922 (C1R), corresponding to the median beta-value in the training set (TCGA). Again, there was a clear association with OS (K-M analysis: P = 0.012; COX analysis: P = 0.035; Fig. 1g, h).

DNA-methylation at C1R has prognostic relevance in AML. a Scheme for selection of the relevant CpG site. b For Kaplan-Meier (K-M) estimation of overall survival (OS) in 194 AML patients, [9] the DNAm levels for each of the 390,000 CpGs were stratified by their median DNAm level. The Manhattan plot demonstrates that the 60 CpGs with adjusted P < 0.05 (dashed line) are distributed over different chromosomes. c Beta-value distribution at the CpG site cg08799922 (C1R) in 656 healthy controls (GSE40279) [19] and 194 AML samples (TCGA) [9]. d K-M plot of TCGA samples classified by median DNAm level at cg08799922 (C1R; median DNAm level 27 %; P < 0.0001). e Beta-value distribution of DNAm in C1R of 10 healthy controls and 62 cytogenetic normal AML (CN-AML) samples (GSE58477) [13]. f Validation of prognostic relevance of cg08799922 for OS in this dataset (P < 0.009; classified by DNAm level of 27 %). g Beta-value distribution of DNAm in C1R of 40 healthy controls and 84 AML samples analyzed by pyrosequencing. h K-M plot of these AML samples upon classification by 27 % DNAm level at the relevant CpG site in C1R (P < 0.012)
Epigenetic co-regulation of C1R with other genomic regions and correlation with gene expression
The serine protease C1R catalyzes the initial event of the classical complement pathway, which is crucial for innate defense mechanisms against pathogens and altered-self cells [14]. The CpG site is located in the 5’UTR region of C1R and may interfere with a CCCTC-binding factor (CTCF) and cohesin-binding site (Fig. 2a) [15]. Differential DNAm in the neighboring genomic region is of prognostic relevance too, as another upstream site of cg08799922 was also among the initial 60 CpGs from the K-M estimate (Fig. 2b; cg06915053, adjusted P = 0.047). Furthermore, neighboring CpGs in the pyrosequencing assay revealed similar correlation with OS as well (Additional file 1: Figure S5). To determine whether or not DNAm at C1R (cg08799922) was co-regulated with other genomic regions, we performed linear correlation of DNAm with all CpGs from the 450K BeadChip: 1448 CpGs correlated (Pearson’s R >0.5), and these were particularly located in homeobox genes (Additional file 1: Figure S6). This co-regulation of C1R and HOX regions might be mediated by chromatin loops via CTCF and cohesin [16]. Furthermore, we analyzed if higher methylation of C1R entails down-regulation of the corresponding gene, but there was only a moderate association from RNA-seq data (R = 0.35, Fig. 2c). However, expression of 82 genes throughout the genome correlated with DNAm in C1R and were linked to developmental processes (R >0.5; Additional file 1: Figure S7). Taken together, DNAm of C1R does not seem to be relevant for functional changes in AML per se; it rather resembles a biomarker that reflects reorganization of chromatin in a subset of AML patients.

DNA-methylation in C1R is associated with clinical parameters. a The CpG site cg08799922 is positioned in a regulatory region in the C1R gene, with binding sites of the architectural protein CTCF and cohesin components SMC1 and RAD21 in close proximity [20]. b Beta-value range of 194 AML patients [9] (yellow) and 656 healthy individuals [19] (brown) at 11 CpGs that are associated with C1R. Particularly, our selected CpG cg08799922 (in 5’UTR; red) and another site cg06915053 (first upstream in TSS200) are associated with OS. c Overall, AML patients [9] with lower DNAm (<27 % DNAm level) at cg08799922 (C1R) have higher gene expression of C1R. Correlation of DNAm at cg08799922 (C1R) with bone marrow blasts (d), gender (e), cytogenetic risk (f), and AML FAB-subtype classification (g). *P < 0.01, **P < 0.001, and ***P < 0.0001 by Mann-Whitney test. h Enrichment of specific mutations according to DNAm level at C1R (P values: hypergeometric distribution)
Association of DNA-methylation at C1R with clinical parameters
DNAm levels in C1R were subsequently compared with clinical parameters of the TCGA dataset [9]: there was no clear association with blast counts (R = 0.0046; Fig. 2d) and no gender difference (Mann-Whitney P = 0.82; Fig. 2e). AML samples with favorable cytogenetic risk score (Fig. 2f) and AML subtype M3 (Fig. 2g) revealed significantly higher DNAm in C1R. Multivariate analysis of OS considering age, gender, bone marrow blast count, and FAB classification demonstrated that DNAm at C1R can support risk stratification (Additional file 1: Table S2). If alternatively only DNAm in C1R, cytogenetic risk score, and molecular risk score were included as parameters for multivariate analysis, the relevance of C1R was also significant (Additional file 1: Table S3). However, if all parameters were combined into one multivariate model, only age and molecular risk score were classified as relevant parameters (Additional file 1: Table S4). To further evaluate if the prognostic relevance of DNAm in C1R is independent from cytogenetic risk groups, we performed K-M analysis within individual cytogenetic groups. DNAm at C1R revealed significant association with OS in the intermediate group of TCGA (K-M analysis: P = 0.015), and the same trend was also observed in the other groups (in TCGA and pyrosequencing datasets; Additional file 1: Figure S8). These results indicate that DNAm in C1R might be of independent prognostic relevance, but this needs to be further validated in larger cohorts. Furthermore, higher methylation at C1R was associated with significantly less mutations in TP53, RUNX1, TET2, and DNMT3A, whereas mutations in CEBPA and WT1 and the translocations PML-PARA and MYH11-CBFB were significantly enriched (Fig. 2h; Additional file 1: Figure S9). Thus, it can be speculated that either DNAm at C1R is influenced by these mutations or vice versa [12, 17]. Because of the strong interaction of DNAm at C1R with prognostic relevant mutations, we performed an additional Kaplan-Meier analysis for patients without the above mentioned mutations. In this subset of patients, DNAm of C1R was also indicative for OS (P = 0.036; Additional file 1: Figure S10).
Analysis of DNA-methylation at C1R in various other types of cancer
To estimate if DNAm in C1R might also be a suitable biomarker for OS in other types of cancer, we utilized 5699 DNAm profiles of 25 different types of tumor from TCGA. In fact, significant association in COX analysis was also observed for kidney renal papillary carcinoma (P = 0.00009), low-grade glioma (P = 0.0002), skin cutaneous melanoma (P = 0.002), hepatocellular carcinoma (P = 0.026), and glioblastoma (P = 0.042; Additional file 1: Table S5). These results indicate similar association of DNAm in C1R with OS in other tumors, but the prognostic relevance was found to be particularly predominant in patients with AML.
Conclusions
In this study, we demonstrate that DNAm at a single CpG site in C1R is indicative for overall survival in AML. In contrast to previously described epigenetic signatures [3, 7, 18], our method is based on only one CpG site. It is unclear if C1R plays a functional role for leukemia development or if it is differentially methylated in leukemia initiating cells, but this is not a prerequisite for biomarkers. Our data indicate that DNAm of C1R may reflect global chromatin reorganization in a subset of AML patients, and this may contribute to specific genomic mutations or vice versa [12, 17]. We have demonstrated that DNAm of C1R was also of prognostic relevance in cytogenetic normal AMLs, in subsets with defined cytogenetic risk score, and in subsets without specific genomic mutations. These results suggest that DNAm in C1R is of independent prognostic significance, but further validation with larger datasets is required to ultimately substantiate the clinical potential. Many cytogenetic aberrations are routinely analyzed in AML diagnostics, but the importance of epigenetic biomarkers should not be neglected, considering that AML develops by means of genetic and epigenetic changes as well. Complementation of epigenetic biomarkers to existing genetic biomarkers could provide a balanced and more accurate diagnostic approach. Furthermore, it is conceivable that DNAm in C1R is also indicative for treatment response to specific drugs, particularly for demethylating agents, and this needs to be addressed in future studies. Either way, analysis of DNAm at a unique CpG site provides a very simple and robust biomarker to complement risk assessment in AML.
Study design
DNAm profiles and bioinformatics analysis
We used publically available DNAm profiles (all based on HumanMethylation450K BeadChip) of AML patients from The Cancer Genome Atlas (TCGA) [9] and a study of Qu et al. (GSE58477) [13], of healthy individuals (GSE40279, GSE35069) [10, 19], and of 25 other types of cancer (TCGA). For Kaplan-Meier estimation of overall survival, we stratified samples by median DNAm levels and adjusted the results for multiple testing (log-rank test calculated in R). Alternatively, we calculated COX regression with OS for each individual CpG site in R.
Blood samples and pyrosequencing
Blood samples were taken from 40 healthy donors and 84 AML patients after written consent according to the guidelines of the ethics committee of the Medical Faculty of the RWTH Aachen (Permit Number: EK206/09). Patient characteristics are summarized in Additional file 1: Table S6. Genomic DNA was isolated from blood, bisulfite converted, and analyzed by pyrosequencing (primers are provided in Additional file 1: Table S7). More detailed methods are described in the supplemental document.
Availability of supporting data
The data sets supporting the results of this article are available at The Cancer Genome Atlas portal (https://tcga-data.nci.nih.gov/tcga/) [9] and at Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo/) under the accession numbers GSE58477 [13], GSE40279 [19], and GSE35069 [10].
Abbreviations
- AML:
-
Acute myeloid leukemia
- C1R:
-
Complement component 1 subcomponent R
- CTCF:
-
CCCTC-binding factor
- DNAm:
-
DNA-methylation
- FAB:
-
French-American-British classification of AML
- K-M:
-
Kaplan-Meier
- OS:
-
Overall survival
- PSQ:
-
pyrosequencing
- TCGA:
-
The Cancer Genome Atlas
References
Patel JP, Gonen M, Figueroa ME, Fernandez H, Sun Z, Racevskis J, et al. Prognostic relevance of integrated genetic profiling in acute myeloid leukemia. N Engl J Med. 2012;366:1079–89.
Shih AH, Abdel-Wahab O, Patel JP, Levine RL. The role of mutations in epigenetic regulators in myeloid malignancies. Nat Rev Cancer. 2012;12:599–612.
Bartholdy B, Christopeit M, Will B, Mo Y, Barreyro L, Yu Y, et al. HSC commitment-associated epigenetic signature is prognostic in acute myeloid leukemia. J Clin Invest. 2014;124:1158–67.
Christopeit M, Bartholdy B. Epigenetic signatures as prognostic tools in acute myeloid leukemia and myelodysplastic syndromes. Epigenomics. 2014;6:371–4.
Figueroa ME, Lugthart S, Li Y, Erpelinck-Verschueren C, Deng X, Christos PJ, et al. DNA methylation signatures identify biologically distinct subtypes in acute myeloid leukemia. Cancer Cell. 2010;17:13–27.
Deneberg S, Guardiola P, Lennartsson A, Qu Y, Gaidzik V, Blanchet O, et al. Prognostic DNA methylation patterns in cytogenetically normal acute myeloid leukemia are predefined by stem cell chromatin marks. Blood. 2011;118:5573–82.
Marcucci G, Yan P, Maharry K, Frankhouser D, Nicolet D, Metzeler KH, et al. Epigenetics meets genetics in acute myeloid leukemia: clinical impact of a novel seven-gene score. J Clin Oncol. 2014;32:548–56.
Jost E, Lin Q, Ingrid WC, Wilop S, Hoffmann M, Walenda T, et al. Epimutations mimic genomic mutations of DNMT3A in acute myeloid leukemia. Leukemia. 2014;28:1227–34.
Cancer Genome Atlas Research Network. Genomic and epigenomic landscapes of adult de novo acute myeloid leukemia. N Engl J Med. 2013;368:2059–74.
Reinius LE, Acevedo N, Joerink M, Pershagen G, Dahlen SE, Greco D, et al. Differential DNA methylation in purified human blood cells: implications for cell lineage and studies on disease susceptibility. PLoS ONE. 2012;7:e41361.
Weidner CI, Lin Q, Koch CM, Eisele L, Beier F, Ziegler P, et al. Aging of blood can be tracked by DNA methylation changes at just three CpG sites. Genome Biol. 2014;15:R24.
Lin Q, Wagner W. Epigenetic aging signatures are coherently modified in cancer. PLoS Genet. 2015;11:e1005334.
Qu Y, Lennartsson A, Gaidzik VI, Deneberg S, Karimi M, Bengtzen S. Differential methylation in CN-AML preferentially targets non-CGI regions and is dictated by DNMT3A mutational status and associated with predominant hypomethylation of HOX genes. Epigenetics. 2014;9:1108–19.
Wijeyewickrema LC, Yongqing T, Tran TP, Thompson PE, Viljoen JE, Coetzer TH, et al. Molecular determinants of the substrate specificity of the complement-initiating protease, C1r. J Biol Chem. 2013;288:15571–80.
Shukla S, Kavak E, Gregory M, Imashimizu M, Shutinoski B, Kashlev M, et al. CTCF-promoted RNA polymerase II pausing links DNA methylation to splicing. Nature. 2011;479:74–9.
Ong CT, Corces VG. CTCF: an architectural protein bridging genome topology and function. Nat Rev Genet. 2014;15:234–46.
Wagner W, Weidner CI, Lin Q. Do age-associated DNA methylation changes increase the risk of malignant transformation? Bioessays. 2015;37:20–4.
Bullinger L, Ehrich M, Dohner K, Schlenk RF, Dohner H, Nelson MR, et al. Quantitative DNA methylation predicts survival in adult acute myeloid leukemia. Blood. 2010;115:636–42.
Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell. 2013;49:459–367.
Ernst J, Kellis M. Discovery and characterization of chromatin states for systematic annotation of the human genome. Nat Biotechnol. 2010;28:817–25.
Acknowledgements
We thank researchers of TCGA and the Karolinska Institute for making their DNAm and sequencing data of AML patients available. This work was supported by the Else Kröner-Fresenius Stiftung (2014_A193), the German Research Foundation (WA/1706/2-1), and the Interdisciplinary Center for Clinical Research (IZKF) within the Faculty of Medicine at the RWTH Aachen University (O1-1).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
WW is involved in the company Cygenia GmbH that may provide service for this method (www.cygenia.com). Apart from this, the authors have nothing to disclose.
Authors’ contributions
TB and QL carried out the experiments. JF and MH supported pyrosequencing. EJ and THB provided AML samples. SW and CM-T supported analysis. TB, QL, EJ, and WW conceived the study and wrote the manuscript. All authors read and approved the final manuscript.
Tanja Božić and Qiong Lin contributed equally to this work.
Additional file
Additional file 1:
PDF with all supplemental data. This file contains additional details on methods, Tables S1–S7, and Figures S1–S10. (PDF 1718 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Božić, T., Lin, Q., Frobel, J. et al. DNA-methylation in C1R is a prognostic biomarker for acute myeloid leukemia. Clin Epigenet 7, 116 (2015). https://doi.org/10.1186/s13148-015-0153-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13148-015-0153-6