
Abstract
Objective
A centrifugation-assisted peptide-mediated gene transfer (CAPT) method was recently developed as an efficient system for gene delivery into plant cells. However, the gene transfer efficiency of CAPT into plant cells was not entirely satisfactory for detecting transient expression of a transgene driven into mitochondria. Here, we report a new gene delivery system using a method called particle bombardment-assisted peptide-mediated gene transfer (PBPT).
Results
We investigated various parameters of the PBPT method to increase transient gene expression efficiency in Brassica campestris. The optimal conditions for PBPT were a single bombardment with gold particles coated with a DNA‒peptide complex (6 µg of DNA and 2 µg of peptide) at an acceleration pressure of 5 kg/cm2 and a target distance of 12.5 cm. Moreover, bombardment under the optimal conditions successfully transferred the transgene into the cells of other plant species, namely B. juncea and tomato. Thus, we developed a PBPT method for highly efficient delivery of a DNA‒peptide complex into plant mitochondria.
Similar content being viewed by others
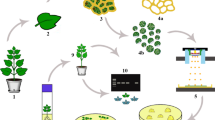


Introduction
Mitochondrion is an essential cellular organelle for catalyzing ATP synthesis by oxidative phosphorylation in eukaryotes [1]. Compared to canonically mammalian mitochondrial genomes, higher plant mitochondria have larger genomes and greater numbers of mitochondrial genes [2]. In several plant species, the cytoplasmic male sterility (CMS)-associated gene is located on the mitochondrial genome, and the CMS trait is valuable in crop breeding [3]. In conventional plant breeding, the CMS trait can be introduced only from crossable plant species with CMS-associated genes. Thus, theoretically, the introduction of an exogenous CMS-associated gene could be accomplished by transmitochondrial technology in all plant species.
On the other hand, transmitochondrial technologies have not been established in higher organisms [4]. Recently, we developed a system for gene delivery to plant mitochondria by using a fusion peptide composed of polycationic peptides with mitochondrial targeting signals (cytocox-KH) and successfully integrated a transgene into a plant mitochondrial genome [5, 6]. We also developed the centrifugation-assisted peptide-mediated gene transfer (CAPT) method as a high-throughput means of transferring DNA‒peptide complexes to plant organelles [7].
It has been reported that many mitochondria exist in plant cells; for example, a single onion epidermal cell contains more than 15,000 mitochondria [8]. Since mitochondria are more numerous than nuclei and chloroplasts (80 to 120 chloroplasts per Arabidopsis mesophyll cell [9]) in a plant cell, it is necessary to introduce many complexes into plant cells. In the present study, we developed a method called particle bombardment-associated peptide-mediated gene transfer (PBPT) that introduces complexes into plant cells more efficiently than the CAPT method.
Materials and methods
Plant materials and growth conditions
B. campestris (cv. Kyoto Fushimi Kanzaki Hanana), B. juncea (cv. Coral Reef Plume), and tomato (cv. Momotaro) seeds (all purchased from Takii Seed, Kyoto, Japan) were treated with 70% EtOH for 1 min, followed by 20% bleach for 15 min, then rinsed 3 times with sterile distilled water. The sterilized seeds were placed on germination medium (GM) containing half-strength Murashige and Skoog medium [10] with 10 g/L sucrose and 2.5 g/L Phytagel (Sigma-Aldrich, St. Louis, MO, USA) at pH 5.8. Seeds were germinated under a 16 h light/8 h dark condition at 30 °C (B. campestris) or 25 °C (B. juncea and tomato). After 7 days, the cotyledons of Brassica spp. and the hypocotyls of tomato were used as explants.
Plasmid construction
pGWB-Prrn18: Nluc was constructed in the 35S-Nluc-TNOS backbone [11]. This plasmid contains the NLuc gene [12] driven by Prrn18. Prrn18 is the promoter region of the gene coding for mitochondrial 18S ribosomal RNA (rrn18) and probably activates strong transcription [13]. The DNA fragments containing Prrn18 was amplified using the primers, SacI-rrn18 F (5’-GCGAGCTCCGGAAGTAGCGCGACAAAGAG-3’, SacI site is underlined) and KpnI-rrn18 R (5’- GCGGTACCAACTCTTCTTTTGAGTATGAT-3’, KpnI site is underlined) from Nicotiana tabacum mitochondrial DNA as a template. The PCR product of Prrn18 was inserted into the SacI-KpnI region of the 35 S-Nluc-TNOS vector to replace CaMV 35 S promoter (Fig. S1).
Particle bombardment of DNA‒peptide complex
To prepare the DNA‒peptide complex, a plasmid, pGWB-Prrn18: Nluc, and a peptide, cytocox-KH (amino acid sequence: MLSLRQSIRFFKKHKHKHKHKHKHKHKHKH), were mixed by the method described previously [14, 15]. The DNA‒peptide/gold coating procedure was carried out as followings: 10 µL aliquots of gold particles (Tanaka Kikinzoku Kogyo, Tokyo, Japan; 0.3–2 μm in diameter, 60 mg/mL) were coated with the complex mixture (6 µg of DNA and 2 µg of peptide, 10 µL of 2.5 M CaCl2, and 4 µL of 0.1 M spermidine) by vortex vigorously. The mixture was incubated at room temperature for 60 min and centrifuged to collect the gold particles, which were then washed with 100 µL of 70% EtOH and resuspended in 15 µL of 99.5% EtOH. Fifteen microliter solution was used per bombardment.
The particle inflow gun and general bombardment methods were described previously [16]. Seven cotyledons of B. campestris, 10 cotyledons of B. juncea, or 14 hypocotyl segments of tomato, respectively, were placed in the center of a 9 cm petri dish containing 20 mL of 1.5% agar plate. The cotyledons of B. compestris was bombarded at various target distances, DNA‒peptide complex loads per bombardment, numbers of bombardments, and acceleration pressures by nitrogen gas (Table 1). B. campestris was suitable for sample preparation and was used for optimize the parameters of PBPT method. The cotyledons of B. juncea or the hypocotyl segments of tomato was bombarded using the optimal conditions determined by B. campestris. The explants were transferred to GM and were incubated in the dark at 30 °C. After 1 day, the bombarded samples of 4 cotyledons of B. campestris, 6 cotyledons of B. juncea, or 7 hypocotyl segments of tomato were homogenized in GUS/LUC buffer [17] with 5.0 mm Stainless Beads (Biomedical Science, Tokyo, Japan) at 1,100 rpm for 30 s using Shake Master (Biomedical Science). The homogenized solutions were centrifuged at 10,000 xg for 10 min, and the luciferase activity of the supernatants was measured with the Nano-Glo Luciferase Assay System (Promega, Madison, WI, USA) analyzed on a GM2000 GloMax Navigator Microplate Luminometer (Promega). The total protein in each sample was quantified by the Bradford assay (XL-Bradford KY-1040, Apro Science, Tokushima, Japan). The luciferase activities were evaluated in relative light units (RLU) per milligram of protein.
Transfer of DNA‒peptide complex by CAPT method
The CAPT method was performed as described previously [7]. Four cotyledons of B. campestris were added to 500 µL of the solution containing the complex mixed with 15 µg of DNA and 5 µg of peptide, followed by centrifugation at 10, 000 xg for 60 s. The luciferase activities were evaluated by the methods described above.
Statistical analyses
All statistical analyses were performed by EZR software [18]. Statistical significance was determined using Mann–Whitney U-test. All values are expressed as mean ± standard error (SE).
Results and discussion
PBPT method compared to CAPT inB. campestris.
To develop the PBPT method, a reporter construct designated as pGWB-Prrn18: Nlu encodes Nano luciferase (Nluc) [12] under the control of tobacco mitochondrial rrn18 promoter (Prrn18). The cotyledons of B. campestris were bombarded once with gold particles coated with either pGWB-Prrn18: Nluc only or a complex of pGWB-Prrn18: Nluc and cytocox-KH at an acceleration pressure of 2 kg/cm2 and with a distance of 12.5 cm between the syringe filter and the target plate. These parameters were used as the standard conditions (Table 1) to evaluate each parameter separately (n = 4 × 12). The gene transfer efficiency by bombardments with the DNA‒peptide complex showed a 5.4-fold higher median in comparison with that of DNA (P < 0.0001, Fig. 1). This result suggested that the particle bombardment method transferred a small amount of exogenous DNA solely to mitochondria. Using the complex with cytocox-KH, the median gene transfer efficiency of the PBPT method (P = 0.0009) was 3.5-fold that of the CAPT method (Fig. 1) [7]. Because Cytocox-KH does not contain a cell-penetrating peptide, it has a poor ability to permeate plasma membranes [5]. In the CAPT method, the DNA‒peptide complex can accumulate only on the plant cell surface and not actively penetrate plant cells [7]. Using particle bombardment, gold particles coated with the DNA‒peptide complex can easily penetrate plasma membranes [19]. Because their enables high permeability of plasma membranes, the PBPT method provides highly efficient delivery of complexes into plant cells.

Transient expression of the Nluc gene in B. campestris transformed by CAPT and PBPT.
Effect of target distance in B. campestris
Our inflow gun can be adjusted to three target distances: 6.2, 9.6, and 12.5 cm. We optimized the target distance based on the standard conditions (Table 1). The target distance is an important parameter for spreading gold particles with the DNA‒peptide complex over a target area [20]. Nluc expression bombarding a target distance of 12.5 cm is significantly greater than that of 6.2 cm (P = 0.0153, Fig. S2). No significant difference in Nluc expression was observed between 6.2 and 9.6 cm or between 9.6 and 12.5 cm (Fig. S2). Thus, the target distance of 12.5 cm is optimal.
Effect of DNA‒peptide complex load per bombardment in B. campestris
To identify the optimal conditions, the amount of DNA‒peptide complex is also an important bombardment parameter. We evaluated the DNA‒peptide complex load per bombardment. To prepare 8 µg of DNA‒peptide complex under standard conditions (Table 1), 6 µg DNA was mixed with 2 µg peptide. A total of 8 µg of the complex per bombardment achieved the highest Nluc expression (Fig. S3). In previous studies, particles coated with large amounts of DNA had low membrane permeability due to particle aggregation [21], and a high concentration of cationic peptide induced cytotoxicity in plant cells [14]. These results suggested that the expression levels induced by 16 and 24 µg of DNA‒peptide complex per bombardment were remarkably decreased.
Effect of the number of bombardments in B. campestris
The cotyledons of B. campestris were bombarded once or twice to analyze the efficiency of Nluc expression. Several reports indicated that multiple bombardments affected gene transfer efficiency in various plant species [21,22,23]. Nluc expression induced by two bombardments was significantly lower than that induced by one bombardment (P = 0.0100, Fig. S4). Multiple bombardments of target tissues may cause cytotoxicity induced by cationic peptide [14] and mechanical damage by gold particles [24]. Thus, a single bombardment is optimal.
Effect of acceleration pressure in B. campestris
Acceleration pressure is an important parameter for estimating mechanical damage to target plants. To achieve high gene transfer efficiency without mechanical damage by gold particles, we optimized acceleration pressure by nitrogen gas (1, 2, 3, 5, and 7 kg/cm2). The highest Nluc expression was observed at 5 kg/cm2 nitrogen pressure, and the 1 kg/cm2 nitrogen pressure had the lowest gene transfer efficiency (Fig. S5). Each explant has different characteristics, such as size, shape, thickness, and surface structure. Therefore, the acceleration pressure might be necessary to optimize depending on each explant [25]. These results demonstrated the optimal conditions for the bombardment of B. campestris cotyledons.
Optimized bombardment conditions in B. juncea and tomato
To validate the PBPT method for use with other plant species, DNA‒peptide complexes were introduced into cells of B. juncea and tomato using the optimal conditions determined above. The Nluc expression levels in both plants were significantly higher than those in the respective controls (2.4-fold, P = 0.0293; Figs. 1 and 2A.7-fold, P = 0.0028; Fig. 2B. Customizing the parameters (e.g., acceleration pressure) for each plant species might enhance the PBPT method’s efficiency at introducing DNA‒peptide complexes into plant cells.

Transient expression of the Nluc gene in other plant species bombarded by the optimal conditions
In the PBPT method, a majority of the DNA‒peptide complexes hit to cytosol are subsequently delivered to mitochondria by a function of mitochondrial targeting signals of Cytocox-KH [5, 6]. By conventional particle bombardment method, the plastid transformation of several crops has been reported [26], however, the mitochondrial transformation has not [27]. Despite the success of DNA delivery to mitochondria, the transmitochondrial plants have not been produced using Cytocox-KH [5, 6]. The CAPT method was developed to enhance DNA delivery using an artificial peptide fused with cell-penetrating peptide (CPP) for nuclear transformation [7]. We expected that a combination of Cytocox-KH and the CAPT method deliver a large amount of DNA to mitochondria. However, the CAPT method was insufficient for DNA delivery with Cytocox-KH lacking CPP to penetrate the cell membrane (Fig. 1). The PBPT method overcame the problem of Cytocox-KH showing less penetration of the cell membrane. Similar to transplastomic plants, homoplasmic transmitochondrial plants may produce more recombinant proteins than heteroplasmic plants [28]. Therefore, the efficient transformation of plant mitochondria by the PBPT method will require increased efficiency of the introduction of DNA‒peptide complexes into plant mitochondria.
Limitation
Although we introduced transgenes into the mitochondrial genome by the optimized PBPT method and homologous recombination, we could not detect the transgene integration by Southern blot analysis. Since a selectable marker for mitochondrial genome transformation has not been developed, a transmitochondrial plant has not been produced yet. It is unclear whether Nluc expression in this study depends only the expression of Prrn18: Nluc in mitochondria, as we have not demonstrated that no cytocox-KH is delivered to the nucleus or chloroplast. In addition, it is unknown whether the PBPT method is applicable to monocots, such as rice, wheat, and barley.
Data Availability
The data of the current study are available from the corresponding author (M. K.) on reasonable request.
Abbreviations
- PBPT:
-
Particle Bombardment-assisted Peptide-mediated gene Transfer
- Nluc:
-
Nanoluc
- CAPT:
-
Centrifugation-Assisted Peptide-mediated gene Transfer
References
van der Bliek AM, Sedensky MM, Morgan PG. Cell Biology of the Mitochondrion Genetics. 2017;207(3):843–71.
Møller IM, Rasmusson AG, Van Aken O. Plant mitochondria – past, present and future. Plant J. 2021;108(4):912–59.
Kim YJ, Zhang D. Molecular Control of male fertility for crop hybrid breeding. Trends Plant Sci. 2018;23(1):53–65.
Li S, Chang L, Zhang J. Advancing organelle genome transformation and editing for crop improvement. Plant Commun. 2021;2(2):100141.
Chuah J-A, Yoshizumi T, Kodama Y, Numata K. Gene introduction into the mitochondria of Arabidopsis thaliana via peptide-based carriers. Sci Rep. 2015;5(1):7751.
Yoshizumi T, Oikawa K, Chuah J-A, Kodama Y, Numata K. Selective gene delivery for integrating exogenous DNA into Plastid and mitochondrial genomes using peptide – DNA complexes. Biomacromolecules. 2018;19(5):1582–91.
Kimura M, Yoshizumi T, Numata K. A centrifugation-assisted peptide-mediated gene transfer method for high-throughput analyses. Plant Biotechnol. 2019;36(1):49–52.
Arimura S, Tsutsumi N. Plant mitochondrial fission and fusion. Plant Biotechnol. 2005;22(5):415–8.
Crumpton-Taylor M, Grandison S, Png KM, Bushby AJ, Smith AM. Control of starch granule numbers in Arabidopsis chloroplasts. Plant Physiol. 2012;158(2):905–16.
Murashige T, Skoog F. A revised medium for Rapid Growth and Bio assays with Tobacco tissue cultures. Physiol Plant. 1962;15(3):473–97.
Midorikawa K, Kodama Y, Numata K. Vacuum/Compression infiltration-mediated permeation pathway of a Peptide-pDNA complex as a non-viral carrier for Gene Delivery in Planta. Sci Rep. 2019;9(1):1–10.
England CG, Ehlerding EB, Cai W. NanoLuc: a small luciferase is brightening up the field of Bioluminescence. Bioconjug chem. 2016;27(5):1175–87.
Hameed MW, Juszczak I, Bock R, van Dongen JT. Comparison of mitochondrial gene expression and polysome loading in different tobacco tissues. Plant Methods. 2017;13(1):112.
Lakshmanan M, Kodama Y, Yoshizumi T, Sudesh K, Numata K. Rapid and efficient gene delivery into plant cells using designed peptide carriers. Biomacromolecules. 2013;14(1):10–6.
Lakshmanan M, Yoshizumi T, Sudesh K, Kodama Y, Numata K. Double-stranded DNA introduction into intact plants using peptide-DNA complexes. Plant Biotechnol. 2015;32:39–45.
Akashi R, Yuge C, Gondo T, Kawamura O, Hoffmann F. Bialaphos-resistant cells of Dallisgrass(Paspalum dilatatum Poir.)Through particle bombardment with a simple self-built Inflow Gun. Grassl Sci. 2002;47(6):588–93.
Yamamoto YY, Deng XW. A New Vector Set for GAL4-Dependent transactivation assay in plants. Plant Biotechnol. 1998;15(4):217–20.
Kanda Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplantat. 2013;48(3):452–8.
Stephens DJ, Pepperkok R. The many ways to cross the plasma membrane. Proc Natl Acad Sci U S A. 2001;98(8):4295–8.
O’Brien JA, Lummis SCR. Biolistic transfection of neuronal cultures using a hand-held gene gun. Nat protocols. 2006;1(2):977.
Mookkan M. Particle bombardment – mediated gene transfer and GFP transient expression in Seteria viridis. Plant Signal Behav. 2018;13(4):e1441657–e.
Daniels D, Kosky RG, Posada-Perez L. Genetic Transformation of the Banana Hybrid Cultivar ‘FHIA-18’ (Musa sp.) via particle bombardment using argon gas. Int J Adv Sci Res Eng. 2018;4(9):42–8.
Zhao Y, Zhang Y, Su P, Yang J, Huang L, Gao W. Genetic transformation system for woody plant Tripterygium wilfordii and its application to product natural celastrol. Front Plant Sci. 2018;8:2221.
Dhir S, Knowles K, Pagan CL, Mann J, Dhir S. Optimization and transformation of Arundo donax L. using particle bombardment. Afr J Biotechnol. 2010;9(39):6460–9.
Helenius E, Boije M, Niklander-Teeri V, Palva ET, Teeri TH. Gene delivery into intact plants using the Helios™ Gene Gun. Plant Mol Biol Report. 2000;18(3):287–8.
Yu Y, Yu PC, Chang WJ, Yu K, Lin CS. Plastid Transformation: how does it work? Can it be Applied to crops? What can it offer? Int J Mol Sci. 2020;21(14):1–21.
Lacroix B, Citovsky V. Biolistic Approach for transient gene expression studies in plants. Methods Mol Biol. 2020;2124:125–39.
Okuzaki A, Tsuda M, Konagaya KI, Tabei Y. A novel strategy for promoting homoplasmic plastid transformant production using the barnase-barstar system. Plant Biotechnol. 2020;37(2):223–32.
Acknowledgements
We would like to thank Ms. Ayumi Nagai (Takasaki University of Health and Welfare) for her technical supports, Prof. Keiji Numata (Kyoto University) for providing cytocox-KH, and Dr. Sachiko Isobe (Kazusa DNA Research Institute) for providing an experimental apparatus.
Funding
This work was supported by JSPS KAKENHI Grant Number 18K05638 and 22K19140 (M. K.), 20K21319 (T. Y), JST-Mirai Program Grant Number JPMJMI21C3 (T. Y.), and in part, New Energy and Industrial Technology Development Organization (NEDO) Grant (T. Y.).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kimura, M., Endo, A., Nagira, Y. et al. Particle bombardment-assisted peptide-mediated gene transfer for highly efficient transient assay. BMC Res Notes 16, 46 (2023). https://doi.org/10.1186/s13104-023-06320-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13104-023-06320-3