
Abstract
Background
Tau protein serves a pro-inflammatory function in neuroinflammation. However, the role of tau in other inflammatory disorders such as rheumatoid arthritis (RA) is less explored. This study is to investigate the role of endogenous tau and the potential mechanisms in the pathogenesis of inflammatory arthritis.
Methods
We established collagen-induced arthritis (CIA) model in wild-type and Tau-/- mice to compare the clinical score and arthritis incidence. Micro-CT analysis was used to evaluate bone erosion of ankle joints. Histological analysis was performed to assess inflammatory cell infiltration, cartilage damage, and osteoclast activity in the ankle joints. Serum levels of pro-inflammatory cytokines were measured by ELISA. The expression levels of macrophage markers were determined by immunohistochemistry staining and quantitative real-time PCR.
Results
Tau expression was upregulated in joints under inflammatory condition. Tau deletion in mice exhibited milder inflammation and protected against the progression of CIA, evidenced by reduced serum levels of pro-inflammatory cytokines and attenuated bone loss, inflammatory cell infiltration, cartilage damage, and osteoclast activity in the ankle joints. Furthermore, tau deficiency led to the inhibition of classically activated type 1 (M1) macrophage polarization in the synovium.
Conclusion
Tau is a previously unrecognized critical regulator in the pathogenesis of RA and may provide a potential therapeutic target for autoimmune and inflammatory joint diseases.
Similar content being viewed by others

Introduction
Rheumatoid arthritis (RA) is one of the most autoimmune and inflammatory disorders affecting approximately 2% world population, characterized by joint swelling and destruction accompanied by bone erosion and cartilage damage due to synovial inflammation [1]. A range of infiltrating immune cells are implicated in the pathogenesis of RA, mainly comprising macrophages, T cells, B cells, and mast cells [2, 3]. Among them, the macrophage is regarded as the core of mediating inflammation in the development of RA because of its two main distinct phenotypes: classically activated type 1 (M1) and alternatively activated type 2 (M2) macrophages [4, 5]. Distinct phenotypic macrophages play opposing roles in inflammatory arthritis [6]. M1 macrophages produce pro-inflammatory cytokines such as tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β), and IL-6, which could activate fibroblasts to further generate pro-inflammatory factors, resulting in the acceleration of inflammation process [7]. Conversely, M2 macrophages are involved in the resolution of inflammation by the release of anti-inflammatory cytokines such as IL-10 [8].
Tau is defined as a microtubule-associated protein that functions in the regulation of microtubule assembly and stabilization [9, 10]. Compelling evidence have established the physiological and pathological function of tau in diverse neurodegenerative disorders, including Alzheimer’s disease (AD), frontotemporal dementia (FTD), Huntington disease (HD) [11,12,13]. Increasing evidence indicates that tau is implicated in various biological processes inside and outside of neurons by mediating multiple signaling pathways due to its diverse post-translational modifications [14, 15]. Emerging evidence from recent experimental studies indicate that tau plays a significant role in activating microglia and the aggregation of tau can directly induce microglia towards M1 phenotypes [16,17,18], manifesting that tau plays a pro-inflammatory role in neuroinflammation [19]. A significant finding was that tau-transgenic mice exhibited a greater susceptibility to collagen-induced arthritis (CIA), with an earlier onset and higher incidence than the control group [20]. This evidence suggests that tau may exert pro-inflammatory activity in other autoimmune and inflammatory diseases beyond neuroinflammation.
In the current study, we established CIA, an experimental model of human rheumatoid arthritis (RA) [21], in WT and Tau-/- mice to characterize the role of endogenous tau in inflammatory arthritis as well as the potential mechanism involved. Our findings demonstrate that tau deficiency could attenuate the progression of CIA by inhibiting M1 macrophage polarization, and targeting tau may provide a potential therapeutic strategy for treating autoimmune inflammatory joint diseases.
Materials and methods
Mice
C57BL/6 (000664) and Tau-/- (007251) mice were purchased form The Jackson Laboratory (Bar Harbor, ME, USA). Tau-/- mice were on the C57BL/6 background. All mice were housed on a 12-h light/dark cycle with free access to food and water in a specific pathogen‑free facility. All animal protocols were approved by the Institutional Animal Care and Use Committee of New York University.
CIA induction and assessment
Twelve-week-old and weight-matched WT and Tau-/- male mice were randomly divided into two groups: the control group and the CIA group, respectively. CIA group mice were immunized intradermally with 100μL emulsion containing an equal volume of chicken type II collagen (20,012, Chondrex) and complete Freund’s adjuvant (7001, Chondrex) at the base of tail (day 0) [22,23,24]. Booster injections were performed with chicken type II collagen emulsified in incomplete Freund’s adjuvant (7002, Chondrex) on day 19 [25, 26]. Due to the mice on the C57BL/6 background having up to 60% arthritis incidence, the mice with CIA phenotype were included for further study. The mice without CIA phenotype were excluded from the study but could be used for the calculation of arthritis incidence. There were at least 6 control mice or 6 mice with CIA phenotype in the control group or the CIA group, respectively. After the booster injection, the clinical score was assessed on each paw every other day for five weeks using the following system: 0 = no erythema and swelling; 1 = erythema and mild swelling confined to the tarsals or ankle joint; 2 = erythema and mild swelling extending from the ankle to the tarsals; 3 = erythema and moderate swelling extending from the ankle to metatarsal joints; 4 = erythema and severe swelling encompass the ankle, foot and digits, or ankylosis of the limb [27]. Each paw was scored and scores of four paws were summed to give a total clinical score with a maximum possible score of 16 for each mouse. Arthritis incidence was defined as the ratio of the number of mice with CIA phenotype to a total number of immunized mice, multiplied by 100. After Five weeks after boost injections, the mice were sacrificed (day 55), hind paws and sera were harvested for further study.
Micro-CT analysis
Hind paws from WT and Tau-/- mice were fixed in 4% paraformaldehyde (PFA) for 24 h and then stored in 70% ethanol. After fixation, hand paws were scanned using a Scanco vivaCT40 cone-beam scanner (SCANCO Medical, Switzerland) with a source voltage of 55 kV, a source current of 145 μA, and a resolution of 10.5 μm. After reconstruction, the parameters of trabecular bone of distal tibia, including trabecular bone mineral density (Tb.BMD, g/cm3), bone volume/tissue volume (BV/TV, %), trabecular thickness (Tb.Th μm), and trabecular number (Tb.N, 1/mm), were analyzed using CT Analyser (CTan) v.1.18.8.0 (Bruker). The micro-CT images were obtained by CTvox v.3.3.1 software (Bruker).
Histological analysis
Hind paws were decalcified with 10% EDTA for four weeks and embedded in paraffin. Serial paraffin Sects. (5 μm) were prepared and stained with hematoxylin and eosin (H&E) to evaluate the inflammation score and bone destruction score. Inflammation scores were obtained based on the following criteria: 0 = no inflammation; 1 = slight thickening of the lining layer or some infiltrating cells in the underlying layer; 2 = slight thickening of the lining layer plus some infiltrating cells in the underlying layer; 3 = thickening of the lining layer, an influx of cells in the underlying layer, and presence of cells in the synovial space; 4 = synovium highly infiltrated with many inflammatory cells [6]. Safranin O staining was performed for cartilage damage assessment, which was based on the following score system: 0 = normal; 1 = minimal erosion limited to single spots; 2 = slight to moderate erosion in a limited area; 3 = more extensive erosions; 4 = general destruction [28]. Sections were stained for tartrate-resistant acid phosphatase (TRAP) to identify the osteoclasts. All images were captured using the Zeiss microscope (Axio Scope A.1, Carl Zeiss, LLC). The quantification of osteoclasts was performed with ImageJ software.
Immunohistochemistry staining
Paraffin sections were deparaffinized and rehydrated through xylenes and graded ethanol series. Then sections were pre-treated with 0.1% trypsin for antigen retrieval (30 min, 37 °C), followed by incubation with 3% H2O2 (30 min, 4 °C). After incubation with blocking buffer (3% BSA + 20% goat serum) for 60 min at 37 °C, sections were treated with primary antibodies against tau (1:250, PA5-29,610, Invitrogen), iNos (1:100, ab15323, Abcam), CD206 (1:8000, ab64693, Abcam) and F4/80 (1:100, ab6640, Abcam) overnight at 4 °C. The next day, the sections were incubated with biotinylated secondary antibodies for 60 min at room temperature. After washing with PBS, the Vectastain Elite ABC kit (PK-6100, Vector) was used to amplify signals. Sections were then incubated with 0.5 mg/ml 3,3-diaminobenzidine (DAB) in 50 mM Tris–Cl substrate (Sigma-Aldrich) until desired stain intensity developed. After this, sections were counterstained with 1% methyl green, cleared, and mounted. All images were obtained using the Zeiss microscope. The quantification analysis was performed with ImageJ software.
Isolation of primary bone marrow derived macrophages (BMDMs) and M1 and M2 polarization
Bone marrow cells were extracted from twelve-week-old WT and Tau-/- mice and cultured in αMEM supplemented with 10% FBS plus M-CSF (10 ng/mL, Biolegend) for 5 days. After bone marrow cells were differentiated into macrophages, the cells were treated with LPS (100 ng/mL, Sigma-Aldrich) plus IFN-γ (20 ng/mL, Peprotech) or IL-4 (20 ng/mL, Peprotech) for 18 h to polarize cells to M1 or M2 macrophages, respectively.
Quantitative real-time PCR
Total RNA was extracted from cells using TRIzol reagent according to the manufacturer’s instructions. cDNA was prepared using 1 μg RNA with SuperScript® Reverse Transcriptase (M314C, Promega Corporation). Quantitative real-time PCR (qRT-PCR) was performed using SYBR® Green PCR Master Mix (4,309,155, Applied Biosystems). The prepared reaction was conducted using StepOnePlusTM real-time PCR Systems (Applied Biosystems). The following primers were used: Tau (forward: CCTGAGCAAAGTGACCTCCAAG; reverse: CAAGGAGCCAATCTTCGACTGG); IL-6 (forward: TTCCATCCAGTTGCCTTCTTG; reverse: AGGTCTGTTGGGAGTGGTATC); Nos2 (forward: TGTTAGAGACACTTCTGAGGCTC; reverse: ACTTTGGATGGATTTGACTTTGAAG); Arg1 (forward: TGCCAAAGACATCGT GTACATTG; reverse: CTTCCCAGCAGGTAGCTGAAG); Mgl1 (forward: CAGATCCGTATCTGTCTGGATC; reverse: AGGTGGGTCCAAGAGAGGATG). Target gene mRNA levels were analyzed using ΔΔCT method. Relative mRNA fold changes were normalized to Gapdh.
Western blot
Raw264.7 cells were treated with PBS or TNF-α (30 ng/mL, PHC3015, Invitrogen) for 48 h and total protein was then extracted. The protein samples were separated using SDS-PAGE and then transferred a nitrocellulose membrane (1,620,097, Bio-Rad) using a wet transfer system. 5% (w/v) non-fat milk in TBST was used to block the membrane for 1 h at room temperature. The membrane was incubated with anti-Tau antibody (1:1000, MA5-12,808, Invitrogen) or GAPDH (1:1000, 60,004-I-Ig, Proteintech) at 4 °C overnight. After three times washing, horseradish peroxidase conjugated anti-mouse IgG (1:10,000, 115–035-003, Jackson ImmunoResearch Laboratories) was added for 1 h at room temperature. After incubated with chemiluminescent (ECL) substrate (NEL104001, Thermo Fisher Scientific), the bands on the membrane were detected using ChemiDoc Imaging System (17,001,401, Bio-Rad). The quantification of bands was performed using ImageJ software.
ELISA
The concentrations of IL-1β and IL-6 were measured in serum collected from mouse models using ELISA kits, according to product specifications (IL-1β: 88–7013-88, Invitrogen; IL-6: 88–7064-88, Invitrogen).
Statistical analysis
Statistical analysis was performed using GraphPad Prism 8 statistic software (GraphPad). All data were presented as mean ± standard errors (SD). Statistical significance between two groups was analyzed using two-tailed, unpaired Student’s t-tests. Comparison of multiple groups was analyzed using two-way ANOVA. A value of P was considered statistically significant if P < 0.05.
Results
The expression of tau is increased in macrophages and synovium of joints under inflammation conditions
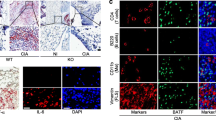
To examine whether tau expression altered under inflammatory conditions, we first measured tau mRNA and protein levels in Raw264.7 cells after TNF-α stimulation. Tau protein level was significantly elevated following TNF-α stimulation while mRNA level remained unchanged (Fig. 1A-C), suggesting inflammation could regulate tau expression at posttranscriptional level. To further determine the alteration of tau expression level in vivo, we performed immunohistochemical staining on ankle joint sections from wild-type (WT) mice with or without CIA. The results revealed that tau expression was dramatically increased in the synovial tissues of CIA mice compared with those of the control group (Fig. 1D, E). Besides, osteoarthritis (OA) is also considered to be a mild chronic inflammatory disease [29], we also determined whether tau expression changed in the synovium of surgically induced destabilization of medial meniscus OA mice. In line with the results obtained from CIA mice, tau expression was also enhanced in the synovium of knee joints from OA mice compared with that from the control group (Fig. 1F, G). Taken together, the expression of tau is increased in macrophages and synovium of joints under inflammation conditions.

Tau protein level is increased in inflammation conditions. A qRT-PCR analysis of Tau mRNA level in Raw264.7 cells treated with PBS or TNF-α (30 ng/mL) for 24 h. B Western blot analysis of tau protein in Raw264.7 cells treated with PBS or TNF-α (30 ng/mL) for 48 h. C Densitometry analysis of Western blot results shown in (B). D Representative images of immunohistochemistry staining of tau in the synovium of ankle joints from WT mice with or without CIA at 7 weeks after immunization. Scale bar: 100 µm. E Quantification of tau positive cells in the synovium of ankle joints. n = 6 mice for each group. F Representative images of immunohistochemistry staining of tau in the synovium of knee joints from sham or DMM operated WT mice at 12 weeks after surgery. Scale bar: 50 µm. G Quantification of tau positive cells in the synovium of knee joints. n = 4 mice for each group. **p < 0.01; ***p < 0.001; ****p < 0.0001. CIA, collagen-induced arthritis; OA, osteoarthritis; DMM, destabilization of the medial meniscus
Tau deficiency reduces arthritis severity
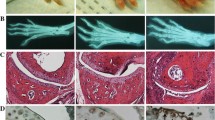
To determine the role of endogenous tau in the pathogenesis of RA, we established CIA model in WT and Tau-/- mice [30, 31]. The absence of tau led to a reduction in hind paw swelling compared to WT mice following CIA induction (Fig. 2A). Accordingly, tau deletion reduced clinical score, delayed the onset of disease, and decreased disease incidence (Fig. 2B, C).

Tau deficiency attenuates the onset and progression of CIA. A Representative images of hind paws of WT and Tau-/- mice with CIA. B, C Clinical score (B) and arthritis incidence (C) of WT and Tau-/- mice with CIA model. n = 6 mice for each group. **p < 0.01
To further investigate the effect of tau deletion on arthritis associated bone erosion, we performed Micro-CT analysis on ankle joints from WT and Tau-/- mice on day 55 of CIA. Micro-CT images demonstrated that ankle joints of WT mice exhibited considerable bone erosion associated with arthritis, while ankle joints of Tau-/- mice showed significantly less bone erosion (Fig. 3A). The reduced bone erosion in Tau-/- CIA mice was also quantitatively evidenced by the increase in bone mineral density (BMD), bone volume/tissue volume (BV/TV), trabecular thickness (Tb. Th), and trabecular number (Tb. N) in the distal tibias compared to those in WT CIA mice (Fig. 3B). Collectively, these findings clearly demonstrate that tau deficiency in mice leads to decreased susceptibility to CIA.

Ankle joint destruction is alleviated in Tau-/- mice with CIA. A Representative micro-CT images of hind paws of WT and Tau-/- mice. Scale bar: 500 µm. B Bone mineral density (BMD), bone volume/tissue volume (BV/TV), trabecular thickness (Tb. Th), and trabecular number (Tb. N) in the distal tibia were analyzed by micro-CT. n = 6 mice for each group. *p < 0.05
Tau deletion alleviates inflammation and bone loss
Enhanced inflammatory cell infiltration, cartilage damage, and osteoclast activity in the ankle joints are typical hallmarks during the progression of CIA [32, 33]. Histopathological and quantitative analysis on the H&E stained ankle joints showed that WT CIA mice have abundant inflammatory cell infiltration, whereas Tau-/- CIA mice exhibited a significant reduction of inflammatory cell inflammation (Fig. 4A, D). Safranin O staining demonstrated that Tau-/- mice were markedly protected from arthritis-associated cartilage loss observed in WT mice (Fig. 4B, E). In addition, tartrate-resistant acid phosphatase (TRAP) staining revealed abundant osteoclasts in WT CIA mice, and tau deletion could markedly protect against arthritis-associated osteoclasts formation and bone destruction (Fig. 4C, F). Accordingly, the serum levels of pro-inflammatory cytokines, such as IL-1β and IL-6, significantly declined in Tau-/- mice with CIA (Fig. 4G, H). The results demonstrate that Tau-/- mice have milder inflammatory arthritis phenotypes and tau deletion significantly protects against the development of inflammatory arthritis.

Tau deletion decreases the inflammation of ankle joints in CIA mice. A-C Representative images of H&E (A), Safranin O (B), and TRAP staining (C) in ankle joints from WT and Tau-/- mice with or without CIA. Scale bar: 100 µm. D, E Inflammation score (D) and cartilage damage score (E) were evaluated according to the scoring system. F Histomorphometric analysis of osteoclast surface per bone surface (Oc.S/BS). G, H Serum levels of IL-1β (G) and IL-6 (H) were measured by ELISA. n = 6 mice for each group. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001
Loss of tau inhibits M1 macrophage polarization
To understand the mechanisms underlying the protective effects of tau deficiency against inflammatory arthritis, we sought to determine whether tau deletion could have an impact on synovial macrophages infiltration and polarization, which are the most abundant cells in the synovium and strongly associated with the pathogenesis of RA [34, 35]. The immunohistochemistry staining for F4/80, a marker of general macrophage, revealed that the total number of macrophages in the synovium of ankle joints from WT mice with CIA was much higher than that of Tau-/- CIA mice (Fig. 5A, B). We next explored the effects of tau deletion on macrophage polarization. The results suggested that the expression of iNOS, a marker for M1 macrophage, in Tau-/- CIA mice was obviously lower than that in WT CIA group (Fig. 5A, C). However, tau deletion appeared to have no effect on M2 macrophage, as evidenced by unchanged number of CD206 positive cells between WT and Tau-/- mice with CIA (Fig. 5A, D). Notably, the deletion of tau did not alter the macrophage infiltration and macrophage polarization under steady condition (Fig. 5A-D).

Tau deletion inhibits macrophage infiltration and M1 polarization in CIA. A Representative images of immunohistochemical staining of F4/80, iNOS and CD206 in the synovium of ankle joints from WT and Tau-/- mice with or without CIA. Scale bar: 100 µm. B-D Quantification of F4/80 positive (B), iNos positive (C) and CD206 positive (D) macrophages in the synovium of ankle joints from WT and Tau-/- mice with or without CIA. n = 6 mice for each group. ****p < 0.0001
To examine the role of tau in macrophage polarization in vitro, we isolated bone marrow–derived macrophages (BMDMs) from WT and Tau-/- mice, which were then polarized to M1 and M2 macrophages, respectively [6, 24]. As shown in Fig. 6A and B, tau deletion significantly reduced the expressions of M1-assocaited genes Il6 and Nos2, without affecting the expressions of M2-associated genes Arg1 and Mgl1 (Fig. 6A-D). In sum, the findings in this study indicate that tau deletion suppresses inflammation by inhibiting M1 macrophage polarization both in vitro and in vivo.

Tau deletion inhibits macrophage skew towards M1 in vitro. A, B mRNA levels of Il6 (A) and Nos2 (B) in WT and Tau-/- BMDMs polarized to M1. C, D mRNA levels of Arg1 (C) and Mgl1 (D) in WT and Tau-/- BMDMs polarized to M2. n = 3 biological replicates. *p < 0.05
Discussion
Tau is well recognized as a microtubule-related protein, abundantly found in brain cells (neurons), and closely associated with the pathogenesis of various neurodegenerative diseases [15, 36]. Numerous studies regarding the function of tau protein mainly focus on neuroinflammation [37,38,39]. However, the role of tau in autoimmune and inflammatory diseases remains largely unknown. In the current study, we demonstrated a previously unrecognized role of tau in CIA, a common model for human RA, by controlling macrophage infiltration and polarization. We observed that tau protein expression level was increased after inflammation stimulation in macrophage in vitro. Accordingly, its expression level was up-regulated in the synovium under inflammatory conditions in both OA and CIA. We further revealed that tau deletion mice were less sensitive to CIA. These results manifest that tau plays a critical role in inflammatory arthritis and suggest manipulation of tau expression and function could be a promising therapeutic target for autoimmune and inflammatory disorders.
RA, caused by autoimmune dysfunction, is defined as a chronic and inflammatory joint disease, characterized by infiltration of the synovial membrane with diverse immune cells as well as neovascularization, progressively leading to the destruction of articular cartilage and bone [40,41,42]. Besides, another prominent feature of RA is the substantial enhancement of osteoclast activity due to inflammatory responses, leading to intense bone loss [43], also known as inflammation-induced bone loss. There are three forms of bone loss in RA: (1) focal articular bone erosion; (2) periarticular osteopenia adjacent to inflamed joints; (3) generalized osteoporosis [44, 45]. The treatment strategies in RA have evolved dramatically in the past two decades, aiming at prevention of the onset of RA, inhibition of inflammation, prevention of disability to maintain normal quality of life [46, 47]. Here, tau deletion exhibited anti-inflammation effect in the course of CIA, including delayed disease onset, downregulation of the serum pro-inflammatory cytokines IL-1β and IL-6, and attenuated inflammatory cell infiltration, cartilage damage and bone erosion in the ankle joints. In particular, decreased number of osteoclasts caused by tau deletion contributed to milder bone loss in ankle joints of CIA mice compared with WT groups. These findings imply that tau is involved in the pathological process of RA and tau deficiency could attenuate the inflammation and bone loss during the progression of RA.
Macrophages, belonging to myeloid immune cells, play an essential role in coordinating the inflammatory process of RA by interacting with other immune cells, including T cells, B cells, neutrophils, synovial fibroblasts, and so on [48,49,50]. Macrophages and functionally heterogeneity and extracellular cues regulate the polarization of macrophages, which comprise two distinguishing phenotypes: classically activated macrophages (M1) and alternatively activated macrophages (M2) [51, 52]. M1 polarization can be induced by some pro-inflammatory stimuli, such as granulocyte–macrophage colony stimulating factor (GM-CSF) and lipopolysaccharide (LPS) [53]. M1 macrophages are considered as pro-inflammatory cells due to the high production of pro-inflammatory cytokines through the activation of nuclear factor kappa B (NF-κB) signaling pathway, including TNF-α, IL-1β, and IL-6 [54], which play an important role in aggravating the inflammatory response. Nevertheless, M2 macrophages serve as anti-inflammatory cells and are generally activated by macrophage colony stimulating factor (M-CSF) and IL-4 [55]. M2 macrophages are responsible for shifting inflammation activation toward inflammation inhibition by high expression levels of IL-10 and transforming growth factor-β (TGF-β), therefore alleviating inflammatory responses [56, 57]. Inducible nitric oxide synthase (iNOS) and cluster of differentiation 206 (CD206), highly expressed in M1 and M2 macrophages, are thought as specific markers for M1 and M2 phenotypes, respectively [50]. Increasing evidence demonstrates that the balance between M1 and M2 is particularly important for the pathogenesis of RA. In the current study, we revealed that tau deletion inhibited M1 macrophage polarization in vitro and in vivo, which is in line with the previous studies that tau induces microglia into M1 phenotype in neurodegenerative disorders [17, 58, 59]. Thus, tau exerts pro-inflammatory effects via skewing macrophages towards M1 macrophages in autoimmune and inflammatory joint disorders. In the current study, we mainly focus on the role of tau protein in M1 polarization and its impact on CIA. One limitation of this study is that the mechanism behind the effect of tau deficiency has not been fully investigated. The exploration of accurate mechanism of M1 macrophage polarization inhibition caused by tau deficiency will be addressed in our future research endeavors.
Conclusions
Taken together, these findings unveil the important function of tau in the inflammatory arthritis and indicate that tau deficiency could reduce inflammatory reactions by inhibiting the polarization of macrophages into M1 phenotype, thereby preventing bone loss and protecting against the progression of CIA. Our work offers the evidence for better understanding the pathogenesis of RA and provides a potential therapeutic strategy for autoimmune and inflammatory diseases.
Availability of data and materials
All data needed to evaluate the conclusions in the paper are present in the paper. Additional data related to this paper may be requested from the authors.
Abbreviations
- RA:
-
Rheumatoid arthritis
- CIA:
-
Collagen-induced arthritis
- M1:
-
Type 1 macrophages
- M2:
-
Type 2 macrophages
- TNF-α:
-
Tumor necrosis factor α
- IL-1β:
-
Interleukin-1β
- AD:
-
Alzheimer’s disease
- FTD:
-
Frontotemporal dementia
- HD:
-
Huntington disease
- Tb.BMD:
-
Trabecular bone mineral density
- BV/TV:
-
Bone volume/tissue volume
- Tb.Th:
-
Trabecular thickness
- Tb.N:
-
Trabecular number
- TRAP:
-
Tartrate-resistant acid phosphatase
- BMDMs:
-
Bone marrow derived macrophages
- qRT-PCR:
-
Quantitative real-time PCR
- WT:
-
Wild-type
- OA:
-
Osteoarthritis
- iNOS:
-
Inducible nitric oxide synthase
- CD206:
-
Cluster of differentiation 206
References
Smolen JS, Aletaha D, McInnes IB. Rheumatoid arthritis. Lancet. 2016;388(10055):2023–38.
Smolen JS, et al. New therapies for treatment of rheumatoid arthritis. The Lancet. 2007;370(9602):1861–74.
Tu J, et al. A Tale of Two Immune Cells in Rheumatoid Arthritis: The Crosstalk Between Macrophages and T Cells in the Synovium. Front Immunol. 2021;12:655477.
Ma Y, Pope RM. The role of macrophages in rheumatoid arthritis. Curr Pharm Des. 2005;11(5):569–80.
Tardito S, et al. Macrophage M1/M2 polarization and rheumatoid arthritis: A systematic review. Autoimmun Rev. 2019;18(11):102397.
Fu W, et al. TNFR2/14–3–3epsilon signaling complex instructs macrophage plasticity in inflammation and autoimmunity. J Clin Invest. 2021;131(16):e144016.
McInnes IB, Schett G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat Rev Immunol. 2007;7(6):429–42.
Siouti E, Andreakos E. The many facets of macrophages in rheumatoid arthritis. Biochem Pharmacol. 2019;165:152–69.
Wang Y, Mandelkow E. Tau in physiology and pathology. Nat Rev Neurosci. 2016;17(1):5–21.
Avila J, et al. Role of tau protein in both physiological and pathological conditions. Physiol Rev. 2004;84(2):361–84.
Brunello CA, et al. Mechanisms of secretion and spreading of pathological tau protein. Cell Mol Life Sci. 2020;77(9):1721–44.
Lee VM, Goedert M, Trojanowski JQ. Neurodegenerative tauopathies. Annu Rev Neurosci. 2001;24:1121–59.
Teravskis PJ, Ashe KH, Liao D. The Accumulation of Tau in Postsynaptic Structures: A Common Feature in Multiple Neurodegenerative Diseases? Neuroscientist. 2020;26(5–6):503–20.
Chang CW, Shao E, Mucke L. Tau: Enabler of diverse brain disorders and target of rapidly evolving therapeutic strategies. Science. 2021;371(6532):eabb8255.
Guo T, Noble W, Hanger DP. Roles of tau protein in health and disease. Acta Neuropathol. 2017;133(5):665–704.
Ising C, et al. NLRP3 inflammasome activation drives tau pathology. Nature. 2019;575(7784):669–73.
Morales I, et al. Tau oligomers and fibrils induce activation of microglial cells. J Alzheimers Dis. 2013;37(4):849–56.
Glass CK, et al. Mechanisms underlying inflammation in neurodegeneration. Cell. 2010;140(6):918–34.
Das R, Balmik AA, Chinnathambi S. Phagocytosis of full-length Tau oligomers by Actin-remodeling of activated microglia. J Neuroinflammation. 2020;17(1):10.
Lang SC, et al. Neurodegeneration Enhances the Development of Arthritis. J Immunol. 2017;198(6):2394–402.
Cho YG, et al. Type II collagen autoimmunity in a mouse model of human rheumatoid arthritis. Autoimmun Rev. 2007;7(1):65–70.
Wei JL, Liu CJ. Establishment of a Modified Collagen-Induced Arthritis Mouse Model to Investigate the Anti-inflammatory Activity of Progranulin in Inflammatory Arthritis. Methods Mol Biol. 2018;1806:305–13.
Fu W, et al. Foxo4- and Stat3-dependent IL-10 production by progranulin in regulatory T cells restrains inflammatory arthritis. FASEB J. 2017;31(4):1354–67.
Liu R, et al. Fexofenadine inhibits TNF signaling through targeting to cytosolic phospholipase A2 and is therapeutic against inflammatory arthritis. Ann Rheum Dis. 2019;78(11):1524–35.
Chen YH, et al. Penfluridol targets acid sphingomyelinase to inhibit TNF signaling and is therapeutic against inflammatory autoimmune diseases. Arthritis Res Ther. 2022;24(1):27.
Zhang Y, Wei F, Liu CJ. Overexpression of ADAMTS-7 leads to accelerated initiation and progression of collagen-induced arthritis in mice. Mol Cell Biochem. 2015;404(1–2):171–9.
Brand DD, Latham KA, Rosloniec EF. Collagen-induced arthritis. Nat Protoc. 2007;2(5):1269–75.
Camps M, et al. Blockade of PI3Kgamma suppresses joint inflammation and damage in mouse models of rheumatoid arthritis. Nat Med. 2005;11(9):936–43.
Scanzello CR, Loeser RF. Editorial: inflammatory activity in symptomatic knee osteoarthritis: not all inflammation is local. Arthritis Rheumatol. 2015;67(11):2797–800.
Tang W, et al. The growth factor progranulin binds to TNF receptors and is therapeutic against inflammatory arthritis in mice. Science. 2011;332(6028):478–84.
Wei JL, et al. Role of ADAMTS-12 in Protecting Against Inflammatory Arthritis in Mice By Interacting With and Inactivating Proinflammatory Connective Tissue Growth Factor. Arthritis Rheumatol. 2018;70(11):1745–56.
Luross JA, Williams NA. The genetic and immunopathological processes underlying collagen-induced arthritis. Immunology. 2001;103(4):407–16.
De Klerck B, et al. Enhanced osteoclast development in collagen-induced arthritis in interferon-gamma receptor knock-out mice as related to increased splenic CD11b+ myelopoiesis. Arthritis Res Ther. 2004;6(3):R220–31.
Kinne RW, Stuhlmuller B, Burmester GR. Cells of the synovium in rheumatoid arthritis. Macrophages Arthritis Res Ther. 2007;9(6):224.
Kinne RW, et al. Macrophages in rheumatoid arthritis. Arthritis Res. 2000;2(3):189–202.
Michalicova A, Majerova P, Kovac A. Tau Protein and Its Role in Blood-Brain Barrier Dysfunction. Front Mol Neurosci. 2020;13:570045.
Naseri NN, et al. The complexity of tau in Alzheimer’s disease. Neurosci Lett. 2019;705:183–94.
Perea JR, Bolos M, Avila J. Microglia in Alzheimer's Disease in the Context of Tau Pathology. Biomolecules. 2020;10(10):1439.
Leng F, Edison P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat Rev Neurol. 2021;17(3):157–72.
McInnes IB, Schett G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet. 2017;389(10086):2328–37.
Aletaha D, Smolen JS. Diagnosis and Management of Rheumatoid Arthritis: A Review. JAMA. 2018;320(13):1360–72.
Zhao J, et al. Molecular and Cellular Heterogeneity in Rheumatoid Arthritis: Mechanisms and Clinical Implications. Front Immunol. 2021;12:790122.
Choi Y, Arron JR, Townsend MJ. Promising bone-related therapeutic targets for rheumatoid arthritis. Nat Rev Rheumatol. 2009;5(10):543–8.
Goldring SR, Gravallese EM. Mechanisms of bone loss in inflammatory arthritis: diagnosis and therapeutic implications. Arthritis Res. 2000;2(1):33–7.
Romas E. Bone loss in inflammatory arthritis: mechanisms and therapeutic approaches with bisphosphonates. Best Pract Res Clin Rheumatol. 2005;19(6):1065–79.
Demoruelle MK, Deane KD. Treatment strategies in early rheumatoid arthritis and prevention of rheumatoid arthritis. Curr Rheumatol Rep. 2012;14(5):472–80.
Burmester GR, Pope JE. Novel treatment strategies in rheumatoid arthritis. Lancet. 2017;389(10086):2338–48.
Udalova IA, Mantovani A, Feldmann M. Macrophage heterogeneity in the context of rheumatoid arthritis. Nat Rev Rheumatol. 2016;12(8):472–85.
Wculek SK, et al. Metabolism of tissue macrophages in homeostasis and pathology. Cell Mol Immunol. 2022;19(3):384–408.
Boutet MA, et al. Novel insights into macrophage diversity in rheumatoid arthritis synovium. Autoimmun Rev. 2021;20(3):102758.
Wang Y, et al. Is macrophage polarization important in rheumatoid arthritis? Int Immunopharmacol. 2017;50:345–52.
Xu Y, et al. Role of macrophages in tumor progression and therapy (Review). Int J Oncol. 2022;60(5):57.
Mantovani A, et al. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004;25(12):677–86.
Cutolo M, et al. The Role of M1/M2 Macrophage Polarization in Rheumatoid Arthritis Synovitis. Front Immunol. 2022;13:867260.
Zhang YH, et al. Modulators of the Balance between M1 and M2 Macrophages during Pregnancy. Front Immunol. 2017;8:120.
Saradna A, et al. Macrophage polarization and allergic asthma. Transl Res. 2018;191:1–14.
Du Y, et al. Macrophage polarization: an effective approach to targeted therapy of inflammatory bowel disease. Expert Opin Ther Targets. 2021;25(3):191–209.
Tang Y, Le W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol Neurobiol. 2016;53(2):1181–94.
Wang Q, et al. Microglia Polarization in Alzheimer’s Disease: Mechanisms and a Potential Therapeutic Target. Front Aging Neurosci. 2021;13:772717.
Acknowledgements
Not applicable.
Funding
This work was supported partly by NIH research grants R01AR062207, R01AR061484, R01AR076900, R01AR078035 and R01NS070328.
Author information
Authors and Affiliations
Contributions
C.L. conceived and designed the project. M.C. and W.F. designed and performed experiments and analyzed the data. H.X. revised the manuscript. All authors contributed to the writing and editing of the manuscript and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All animal protocols were approved by the Institutional Animal Care and Use Committee of New York University.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Chen, M., Fu, W., Xu, H. et al. Tau deficiency inhibits classically activated macrophage polarization and protects against collagen-induced arthritis in mice. Arthritis Res Ther 25, 146 (2023). https://doi.org/10.1186/s13075-023-03133-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13075-023-03133-4