
Abstract
Background
Rheumatoid arthritis (RA) is a systemic chronic inflammatory disease that leads to joint destruction and functional disability due to the targeting of self-antigens present in the synovium, cartilage, and bone. RA is caused by a number of complex factors, including genetics, environment, dietary habits, and altered intestinal microbial flora. Microorganisms in the gut bind to nod-like receptors and Toll-like receptors to regulate the immune system and produce various metabolites, such as short-chain fatty acids (SCFAs) that interact directly with the host. Faecalibacterium prausnitzii is a representative bacterium that produces butyrate, a well-known immunomodulatory agent in the body, and this microbe exerts anti-inflammatory effects in autoimmune diseases.
Methods
In this study, F. prausnitzii was administered in a mouse model of RA, to investigate RA pathology and changes in the intestinal microbial flora. Using collagen-induced arthritic mice, which is a representative animal model of RA, we administered F. prausnitzii orally for 7 weeks.
Results
The arthritis score and joint tissue damage were decreased in the mice administered F. prausnitzii compared with the vehicle-treated group. In addition, administration of F. prausnitzii reduced the abundance of systemic immune cells that secrete the pro-inflammatory cytokine IL-17 and induced changes in SCFA concentrations and the intestinal microbial flora composition. It also resulted in decreased lactate and acetate concentrations, an increased butyrate concentration, and altered compositions of bacteria known to exacerbate or improve RA.
Conclusion
These results suggest that F. prausnitzii exerts a therapeutic effect on RA by regulation of IL-17 producing cells. In addition, F. prausnitzii modify the microbial flora composition and short chain fatty acids in experimental RA mouse model.
Similar content being viewed by others

Background
Rheumatoid arthritis (RA) is a systemic inflammatory autoimmune disease characterized by inflammation and the proliferation of synovial cells, as well as osteoporosis and erosion of bone and joints [1, 2]. Various types of T lymphocytes, such as activated T helper (Th) 1, Th17, and autoreactive CD4+ T cells, and the secretion of pro-inflammatory cytokines such as interleukin (IL)-17, IL-23, IL-6, and TNF-α, are mainly responsible for the development of RA [3,4,5,6].
Although many targeted treatments have been developed for RA alleviation, none can achieve a complete cure because the pathogenesis of RA is not fully understood [7, 8]. Numerous genetic and environmental factors are associated with an increased risk of RA [9]. Factors such as smoking, hormones, microorganisms, and infection may be involved in RA induction [10,11,12].
Dysbiosis of the commensal microbiota is among the most important factors associated with RA development and progression [13,14,15]. In cooperation with intestinal lymphoid tissue, the intestinal microbiota helps maintain immune homeostasis and serves as a marker of the host’s health status [16]. For example, bacterial cells can pass through mucus and stimulate epithelial cells to regulate immune function and enhance the intestinal barrier [17]. The intestinal microbiota can also interact directly with the host via microbial metabolites, such as short-chain fatty acids (SCFAs), to regulate the immune system [18, 19].
Faecalibacterium prausnitzii (formerly known as Fusobacterium prausnitzii) is an anaerobic gram-positive, non-spore-forming, acetate-consuming, butyrate-producing, and extremely oxygen-sensitive bacterium [20, 21]. F. prausnitzii is less abundant in diseased patients with inflammatory diseases such as RA [22], Sjögren’s syndrome [23], and multiple sclerosis [24], whereas it comprises 5–15% of the gut bacteria in healthy adults, suggesting that the relative distribution of F. prausnitzii is a key biomarker of disease [25, 26].
The anti-inflammatory effect of F. prausnitzii, which is well known in Crohn’s disease [27], asthma [28], and multiple sclerosis [29], also supports the importance of F. prausnitzii in various inflammatory diseases [30]. As a butyrate producer, the anti-inflammatory effect of F. prausnitzii has been attributed to butyrate production [31, 32]. Specifically, F. prausnitzii attenuates colitis by producing butyrate to reduce Th17 cell differentiation, and inhibit NF-κB activation and histone deacetylase (HDAC) 3 expression in T cells [33, 34]. The supernatant from F. prausnitzii cells attenuated colitis of mice model and in vitro human cells by inhibiting the differentiation of pro-inflammatory Th17 cells [35]. In addition, F. prausnitzii-derived butyrate ameliorated colitis via HDAC3 inhibition [36]. Butyrate relieved inflammatory arthritis in a collagen-induced arthritis (CIA) mouse model by inhibiting HDAC2 in osteoclasts and HDAC8 in T cells [37]. However, the mechanism of action of F. prausnitzii in RA is still unknown. In this study, we show that the anti-inflammatory effect of F. prausnitzii does not appear to be limited to butyrate.
We investigated the anti-inflammatory and immune-cell-modulatory effects, SCFA levels, and intestinal microbial flora changes induced by F. prausnitzii in a CIA mouse model, which is the most widely used animal model in RA research.
Materials and methods
Preparation of animals and F. prausnitzii
All animal experimental designs and procedures were approved by the animal research ethics committee of the Catholic University of Korea (approval number CUMC-2019–0242-01). Male DBA/1 J mice (Orient Bio, Seongnam, Korea) were maintained in a specific pathogen-free, temperature-controlled environment (22 ± 2 °C) at a relative humidity of 50 ± 10% under a 12 h light/dark cycle. The mice had ad libitum access to standard mouse chow (Ralston Purina, Gray Summit, MO, USA) and water. The fecal isolates F. prausnitzii A2-165 was used. Commercial strains of F. prausnitzii A2-165 was purchased from DSMZ-German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany). F. prausnitzii A2-165 were cultured anaerobically for 24 h at 37 °C in modified reinforced clostridial (MRC) broth in Hungate tubes. The bacteria were harvested by centrifugation at 8000 × g for 10 min at 4 °C, washed in phosphate-buffered saline, and re-suspended in sterile saline. The cells were administered orally (1 × 109 CFU) in 200 μL sterile saline. To assess the anti-inflammatory effect of F. prausnitzii on the development of RA, 10-week-old DBA/1 J mice were orally administered F. prausnitzii (Faecali group) or sterile saline (Vehicle group) for 7 weeks after observation of RA symptoms induced by collagen injection.
Induction of arthritis and F. prausnitzii administration
DBA/1 J mice (6–7 weeks old) were randomly divided into three groups (n = 5 per group): Faecali group, Vehicle group, and Heat-killed Faecali group. Mice were immunized with 100 µg chicken type II collagen (Chondrex Inc., Redmond, WA, USA) dissolved overnight in 0.1 N acetic acid (4 mg/mL) in complete Freund’s adjuvant. The immunizations were given intradermally via the base of the tail. Two weeks after the primary immunization, mice were boosted with 100 µg type II collagen in incomplete Freund’s adjuvant (Chondrex Inc., Redmond, WA, USA). CIA mice were administered 109 CFU F. prausnitzii daily, in a sterile saline suspension in a total volume of 200 µl.
Clinical scoring of arthritis
The onset, duration, and severity of joint inflammation were evaluated twice weekly for 10 weeks after the primary immunization. Arthritis severity was scored using the mean arthritis index on a scale of 0–4, as follows: 0, no swelling; 1, mild swelling confined to the toes; 2, erythema and mild swelling extending from the ankle to the midfoot; 3, moderate swelling extending from the ankle to the metatarsal joints; and 4, severe swelling encompassing the ankle, foot, and digits. Arthritis severity was determined as the sum of the scores for all legs, as assessed by two independent observers blinded to the experimental groups.
Histopathological analysis
Histological analysis was performed to determine the extent of joint damage. The joint tissues of mice were fixed in 4% paraformaldehyde, decalcified in a histological decalcifying agent (Calci-Clear Rapid; National Diagnostics, Atlanta, GA, USA), embedded in paraffin, and sectioned. The sections were deparaffinized using xylene and dehydrated using an alcohol gradient. Sections were stained with hematoxylin and eosin and safranin O.
Immunohistochemical analysis
Paraffin-embedded sections were incubated at 4 °C with the following primary monoclonal antibodies: anti-IL-1β (ab9722; Abcam, Cambridge, UK), anti-TNF-α (ab6671; Abcam), and anti-IL-17 (ab79056; Abcam). Then, the samples were incubated with the respective secondary biotinylated antibodies, followed by incubation for 30 min with streptavidin–peroxidase complex. The reaction product was developed using 3,3-diaminobenzidine chromogen (K3468; Dako, Santa Clara, CA, USA).
Flow cytometry
The cells used for analysis of the CD4+ IFNγ + (Th1), CD4+ IL-4+ (Th2), CD4+ IL-17+ (Th17), and IL-17-secreting CD19+ B cell (B17) populations were stimulated with PMA and ionomycin for 4 h using GolgiStop (BD Biosciences, Franklin Lake, NJ, USA). To quantify the Th1, Th2, Th17, and B17 cell populations, mouse splenocytes were immunostained using a PerCp5.5-conjugated anti-CD4 antibody (eBioscience, San Diego, CA, USA) for T cells and a PerCP5.5-conjugated anti-CD19 antibody for B cells. The cells were fixed and permeabilized using the Cytofix/Cytoperm Plus kit (BD Biosciences) following the manufacturer’s instructions, and then stained with APC-conjugated anti-IFNγ, PE-conjugated anti-IL-4, APC-conjugated anti-IL-10, and FITC-conjugated anti-IL-17 antibodies (eBioscience), respectively. All samples were analyzed using the Attune NxT Flow cytometer (Thermo Fisher Scientific, Waltham, MA, USA). The flow cytometry data were analyzed using FlowJo™ software (FlowJo, Ashland, OR, USA).
Analysis of short-chain fatty acids
The cecum and serum samples were homogenized in 400 µl DW and then acidified using 25% metaphosphoric acid (Sigma Aldrich, St. Louis, MO, USA), at a 1:5 ratio of acid to sample, on ice for 30 min. The samples were centrifuged at 15,000 × g for 15 min, and the supernatants were stored at − 80 °C until further processing. SCFAs such as acetate, lactate, and butyrate were analyzed by high-performance liquid chromatography, using the YL9100 system (Young-lin, Anyang, Korea) equipped with a refractive index detector and Aminex HPX-87H column (300 × 7.8 mm; Bio-Rad Laboratories, Hercules, CA, USA). The liquid culture medium was centrifuged at 18,000 × g for 15 min, and the supernatant was filtered through a 0.2-µm filter. For the analysis, 5 mM sulfuric acid (J.T. Baker, Phillipsburg, MA, USA) was used as the mobile phase, with a flow rate of 0.6 mL/min. The concentration of each organic acid (mM) was normalized to the corresponding concentration of the external SCFA standard.
DNA extraction and 16S rRNA gene amplification and sequencing
DNA from cecum samples (100 mg) was extracted using the QIAamp Fast DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The V3–V4 region of the bacterial 16S rRNA gene was amplified using 16S amplicon PCR forward (5′‑TCG TCG GCA GCG TCA GAT GTG TAT AAG AGA CAG CCT ACG GGN GGC WGC AG‑3′) and reverse (5′‑GTC TCG TGG GCT CGG AGA TGT GTA TAA GAG ACA GGA CTA CHV GGG TAT CTA ATC C‑3′) primers. The resulting products were purified, quantified, and pooled together at equal concentrations (5 ng/µl), as described previously [38]. The purified products were sequenced using the MiSeq platform (Illumina, San Diego, CA, USA).
Bioinformatic analysis
Data analysis was performed using Quantitative Insights Into Microbial Ecology 2 (version 2020.3; http://qiime2.org), as described previously [38]. Paired–end sequences were joined using fastq-join, demultiplexed, and quality-filtered. The filter parameters for trimming and truncating using the DADA2 plugin were 0 and 280, respectively, to remove low-quality sequence regions. Chimeric sequences were removed, and reads were clustered into operational taxonomic units (OTUs) using the Greengenes 13_8 database, with a 99% sequence identity cutoff. Analyses were performed using a rarefied table of 11,000 sequences per sample. In addition, beta diversity was analyzed using the Bray–Curtis, Jaccard, and UniFrac indexes. Differential OTUs among groups and cladograms were identified based on the linear discriminant analysis effect size (LEfSe).
Statistical analysis
Cecal and serum SCFA data were compared by ordinary one-way ANOVA, and Dunnett’s multiple comparison test was used to compare the experimental and control groups. The number of reads from the feature/OTU table was normalized using the cumulative sum method to generate relative abundances. Non-parametric statistical analyses, including the Kruskal–Wallis test, were performed to compare the relative abundances of taxa and alpha diversity in each group. All statistical analyses were performed using Prism 8 (GraphPad Software, Inc., San Diego, CA, USA).
Results
F. prausnitzii attenuates the progression of RA in CIA mice
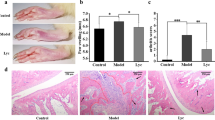
To investigate the anti-inflammatory effects of F. prausnitzii, we induced arthritis in mice via two immunizations at 2-week intervals. Starting at 1 week after the second immunization, 109 CFU F. prausnitzii (Faecali group) and saline (Vehicle group) were administered to the mice, daily for 7 weeks (Fig. 1A). The arthritis score was decreased significantly at week 8, and the incidence of arthritis was notably decreased at week 10, in Faecali mice (Fig. 1B). After 10 weeks of F. prausnitzii administration, the mice were sacrificed and analyzed. Quantitative PCR was performed to detect F. prausnitzii in mouse feces, and the results showed significantly increased abundance of F. prausnitzii in the Faecali group (Fig. 1C). The level of damage in joint tissue was confirmed by hematoxylin and eosin and safranin O staining. Bone erosion and cartilage damage were significantly reduced in the Faecali group compared with the Vehicle group (Fig. 1D). These results suggest that administration of F. prausnitzii prevents arthritis progression and damage to joint tissues.

RA pathogenesis is prevented by oral administration of F. prausnitzii in CIA mice. A Timeline of the immunization and F. prausnitzii administration procedure. B The arthritis clinical score and incidence were reduced after oral F. prausnitzii administration in CIA mice. Arthritis was observed twice a week from week 3 to week 10, which is indicated on the X-axis (w = week). C Mice were sacrificed after 10 weeks F. prausnitzii administration and feces were collected. Quantitative PCR results showing elevated F. prausnitzii abundance in the feces of F. prausnitzii-treated mice. D Microscopy images of histologic sections of hind leg joints of Vehicle and Faecali group mice stained with hematoxylin and eosin (H&E) and safranin O. H&E images were displayed at × 40 magnification and safranin O images at × 200 magnification, respectively. The data are means ± standard deviation (n = 5). *p < 0.05 and **p < 0.01
F. prausnitzii regulates systemic immune cell populations and inhibits IL-17 secretion
As systemic immune cells are closely involved in the progression of autoimmune arthritis and tissue damage, we stained mouse splenocytes and analyzed systemic immune cells by flow cytometry. The proportion of IL-17+ cells among CD4+ cells (Th17) was decreased in the Faecali group. Among CD19+ cells, the proportion of IL-17-secreting cells (B17) was significantly reduced in the Faecali group, while there was no significant change in the abundance of IL-10-secreting cells (data not shown). The abundances of other subtypes of helper T cells, i.e., CD4+ IFNγ+ (Th1) and CD4+ IL-4+ (Th2) cells, were also decreased by F. prausnitzii administration (Fig. 2). These results suggest that F. prausnitzii inhibits IL-17 secretion in both CD4+ and CD19+ cells and induces an anti-inflammatory effect by regulating systemic immune cell populations.

Immunomodulatory effect of F. prausnitzii administration on mouse splenocytes. Th17 (CD4+ IL-17+), B17 (CD19+ IL-17+), and Th1 (CD4+ IFN-γ+)/Th2 (CD4+ IL-4+) cell populations were analyzed by flow cytometry. The data are means ± standard deviation (n = 5). * p < 0.05 and *** p < 0.001
F. prausnitzii decreases pro-inflammatory cytokine levels in joint tissues
To identify the local inflammatory response and immune cell infiltration, we performed immunohistochemical analysis of the pro-inflammatory cytokines TNF-α, IL-1β, and IL-17 in mouse synovial tissue. The levels of TNF-α, IL-1β, and IL-17 were decreased in the Faecali group. In addition, the Faecali group mice were similar to normal mice in term of adipose tissue. In the Vehicle group, fibrosis with immune cell infiltration was observed (Fig. 3). These results suggest that F. prausnitzii reduces immune cell infiltration, pro-inflammatory cytokine expression and, by extension, fibrosis in joint tissue.

Administration of F. prausnitzii leads to a decrease in pro-inflammatory cytokine levels in mouse joint tissues. The levels of TNF-α, IL-1β, and IL-17, which are pro-inflammatory cytokines, were evaluated by immunohistochemistry. The data are means ± standard deviation (n = 5). *p < 0.05 and **p < 0.01
F. prausnitzii regulates SCFA concentrations and the phylum-level microbial flora
To determine the effect of F. prausnitzii administration on the microbial flora of CIA mice, blood was drawn by retro-orbital blood collection immediately before sacrifice after the end of the experiment. Cecal samples were collected after sacrifice. We also injected heat-killed F. prausnitzii as a positive control for F. prausnitzii administration (data not shown). In the mice administered heat-killed F. prausnitzii, there was a slight reduction in the arthritis score but no significant changes in the SCFA concentrations or intestinal microbial flora. We evaluated beta diversity using the Bray–Curtis index. The Vehicle and positive control groups had similar Bray–Curtis index values, which were different from that of the Faecali group (Fig. 4A). SCFA concentrations were measured in the cecum and serum. The lactate level in the cecum was significantly decreased (Fig. 4B), and the butyrate and acetate levels in serum were increased and decreased, respectively, by F. prausnitzii administration (Fig. 4C). Next, we investigated whether F. prausnitzii affects the microbial flora composition at the phylum level. The abundances of Firmicutes and Bacteroides, which accounted for the majority of the phyla, were increased and decreased, albeit not significantly, by F. prausnitzii treatment, respectively. The abundance of Proteobacteria was notably elevated in the Faecali group (Fig. 4D). These results suggest that F. prausnitzii administration leads to changes in SCFA and intestinal microbial flora compositions.

SCFA concentrations and fecal microbial abundances in mice with RA. A Effect of F. prausnitzii administration on bacterial beta diversity based on the Bray–Curtis distance in mice with RA. B The concentrations of SCFAs in cecum (lactate) and C serum (butyrate and acetate). D The average relative abundances of phyla in the Vehicle and Faecali groups. The data are means ± standard deviation (n = 5). *p < 0.05, **p < 0.01, and ***p < 0.001
F. prausnitzii modifies the RA-related intestinal microbial flora at the genus level
To determine whether F. prausnitzii administration prevents the progression of RA and inhibits the production of IL-17 via modification of the microbiota, we investigated changes in the microbial flora at the genus level (Fig. 5A). LEfSe analysis is used to identify high-dimensional biomarkers. The contributions of microbial flora changes to the differences between the groups were assessed using the linear discriminant analysis score. LEfSe analysis revealed the top 10 most abundant taxa in the Faecali group and the top 5 most abundant in the Vehicle group (Fig. 5B). Based on LEfSe analysis, the abundances of Bilophila and Akkermansia, which are negatively associated with pro-inflammatory cytokines production, were increased in the Faecali group (Fig. 5C), while those of Desulfovibrio and Bacteroides, which are positively associated with pro-inflammatory cytokines secretion, were decreased in the Faecali group (Fig. 5D) [39]. In addition, we investigated whether F. prausnitzii administration affects the abundances of other butyrate-producing bacteria, i.e., Roseburia, Coprococcus, Oscillospira, Ruminococcus, and Clostridium. The abundances of butyrate-producing bacteria tended to be increased by F. prausnitzii administration, but only that of Roseburia was significantly increased. These data demonstrated that F. prausnitzii administration exerts a therapeutic effect on RA by modifying the composition of the intestinal microbial flora (Fig. 5E).

Effect of F. prausnitzii administration on the fecal microbial composition at the genus level. A The average relative abundances of microbial genera after F. prausnitzii administration (taxonomic analysis with relative abundance > 0.5%). B Significantly different taxon abundances between the Faecali (green) and Vehicle (red) groups identified by LEfSe analysis (threshold > 2.0). C The relative abundances of the genera Bilophila and D Akkermansia and Desulfovibrio and Bacteroides. E The relative abundances of the genera of Roseburia, Coprococcus, Oscillospira, Ruminococcus, and Clostridium were presented. The data are means ± standard deviation (n = 5). p < 0.05, **p < 0.01, and ***p < 0.001
Discussion
The CIA mouse model, created by injecting DBA/1 J mice with an emulsion of complete Freund’s adjuvant and type II collagen, which induces arthritis in 80–100% of animals, is commonly used to evaluate RA [40, 41]. The CIA mouse model shares immunological and pathological features with human RA [42] and is suitable for investigating the pathogenesis and mechanisms of RA [43]. In this study, we used the CIA mouse model to investigate the protective effects of F. prausnitzii on inflammation and the composition of the intestinal microbial flora.
The mechanism of RA is complex and diverse; it involves the genetic background, environment, and microbiome, and complete cure has not yet been achieved [44, 45]. Previous studies showed that butyrate has powerful anti-inflammatory effects in autoimmune disease model mice or human [37, 46, 47]. Hence, this study investigated the global effect of the administration of F. prausnitzii, which is a well-known butyrate producer, and the underlying mechanism.
We found that the arthritis score, arthritis incidence, and tissue damage were decreased by F. prausnitzii administration. Also, the proportions of IL-17-secreting cells, including Th17 and B17 cells in the spleen, were significantly decreased in the Faecali group compared with the Vehicle group. This result was consistent with a previous study using a DSS-induced colitis mouse model, in which the plasma IL-17A level and percentage of Th17 splenocytes were significantly reduced in the mice treated with F. prausnitzii supernatant [35]. Our previous studies reported that butyrate inhibits RA progression by regulating HDAC 8 in T cells and suppresses Sjögren’s syndrome by regulating IL-10-secreting B cells [37, 46]. In addition, Th1 and Th2 cells were reduced in the Faecali group in this study. Immunohistochemistry showed that fibrosis in joint tissue and pro-inflammatory cytokine expression were decreased in the Faecali group compared with the Vehicle group. These data suggests that F. prausnitzii may inhibit arthritis in a CIA model by regulating the function of inflammatory immune cells.
We conducted additional experiments to determine how F. prausnitzii induces changes in the gut microbial flora and SCFA production. Analysis of the gut microbiota revealed different remodeling patterns among the treatment groups. In the cecum, the lactate level was decreased in the Faecali group. Lactate promotes chronic inflammation and exacerbates RA pathology. SCFAs are the major end products of bacterial fermentation in the colon [48]. Bacterial strains break down complex polysaccharides into monosaccharides, which are further fermented into SCFAs, including acetate, butyrate, and propionate [49, 50]. Butyrate is the key energy source for colonocytes, and plays important roles in the maintenance of colonic mucosal health, inhibition of inflammation and oxidative stress, improvement of barrier function, and promotion of satiety [51]. The serum butyrate level was higher in our Faecali than Vehicle group; however, there was no difference in the cecal butyrate level between the groups (data not shown). In this study, the propionate level was not significantly different between the Faecali and Vehicle groups (data not shown). However, the acetate level was decreased, and the butyrate level increased, in the serum of F. prausnitzii-treated mice. F. prausnitzii consumes acetate and produces butyrate, and butyrate promotes the differentiation of regulatory T cells [52, 53]. However, in this study, the elevated butyrate level induced by F. prausnitzii treatment modulated the abundance of IL-17-secreting immune cells but did not lead to a significant increase in regulatory T cell abundance. This suggests that oral administration of F. prausnitzii regulates the composition of SCFAs in the body. In addition, the lactate level in the cecum was decreased by F. prausnitzii administration. Lactate activates the tricarboxylic acid cycle via pyruvate in T cells [54]. The resulting fatty acid synthesis not only reduces cell motility but also promotes activation of the transcription factor STAT3 [55]. Activated STAT3 is a well-known inducer of RORγt, which promotes IL-17 production [56, 57].
Interestingly, administration of a single F. prausnitzii bacterium elicited changes in the intestinal microbial flora in the RA mouse model. The ratio of Firmicutes to Bacteroidetes, the dominant microbial communities at the phylum level, tended to be elevated in the Faecali group, but the differences between the groups were not significant. Proteobacteria, which has a low abundance in the microbiota, was increased in the Faecali group. Previous studies showed that the Firmicutes to Bacteroidetes ratio and Proteobacteria abundance were decreased in RA patients compared with healthy controls in East Asia [58]. These results suggest that F. prausnitzii administration improves the health of the RA mouse model.
In the Faecali group, the abundances of Bilophila and Akkermansia, which promote RA, were increased. The abundance of Bilophila is negatively correlated with TNF-α production [39]. The abundance of Akkermansia muciniphila, a mucus-colonizing member of microbiota, was increased by F. prausnitzii treatment. The mucin degradation activity of Akkermansia muciniphila leads to the production of propionate and acetate [59, 60]. On the other hand, Desulfovibrio and Bacteroides abundances were reduced by F. prausnitzii administration. Desulfovibrio spp. are anaerobic sulfate-reducing bacteria that colonize the human gut; they are associated with gastrointestinal diseases and stimulation of the epithelial immune response, including nitric oxide production in macrophages [61]. In addition, Bacteroides fragilis leads to arthritis and is present in autoimmune disease patients [62,63,64]. Although the relationship between RA and the intestinal microbial flora is not well understood, this study showed that the administration of F. prausnitzii can induce positive changes in the microbial flora for RA.
In summary, this is the first study to evaluate the effects of F. prausnitzii on RA in an animal model. F. prausnitzii altered gut microbial composition and exerted an anti-inflammatory effect associated with the regulation of IL-17-producing immune cells.
Conclusion
The aim of this study was to investigate the effect of F. prausnitzii administration on SCFA metabolism, immunomodulatory activity, and gut microbial composition. The oral administration of F. prausnitzii prevents disease progression and regulates SCFA, proportion of immune cells and intestinal microbial flora on RA mouse model. Hence, this study suggests that F. prausnitzii administration is a novel strategy for RA treatment and symptom relief, although it is difficult to cultivate because of its extreme oxygen sensitivity.
Availability of data and materials
All data are available in the manuscript or upon request to the authors.
Abbreviations
- RA:
-
Rheumatoid arthritis
- SCFAs:
-
Short-chain fatty acids
- Th:
-
T helper
- IL:
-
Interleukin
- HDAC:
-
Histone deacetylase
- CIA:
-
Collagen-induced arthritis
- MRC:
-
Modified reinforced clostridial
- IFN:
-
Interferon
- OTU:
-
Operational taxonomic units
- TNF:
-
Tumor necrosis factor
References
Smolen JS, Aletaha D, Barton A, Burmester GR, Emery P, Firestein GS, Kavanaugh A, McInnes IB, Solomon DH, Strand V, Yamamoto K. Rheumatoid arthritis. Nat Rev Dis Primers. 2018;4:18001.
Yang Y, He X, Zhao R, Guo W, Zhu M, Xing W, Jiang D, Liu C, Xu X. Serum IFN-gamma levels predict the therapeutic effect of mesenchymal stem cell transplantation in active rheumatoid arthritis. J Transl Med. 2018;16:165.
Chemin K, Gerstner C, Malmstrom V. Effector functions of CD4+ T cells at the site of local autoimmune inflammation-lessons from rheumatoid arthritis. Front Immunol. 2019;10:353.
Chen Z, Bozec A, Ramming A, Schett G. Anti-inflammatory and immune-regulatory cytokines in rheumatoid arthritis. Nat Rev Rheumatol. 2019;15:9–17.
Jacobs JP, Wu HJ, Benoist C, Mathis D. IL-17-producing T cells can augment autoantibody-induced arthritis. Proc Natl Acad Sci U S A. 2009;106:21789–94.
Wang J, He L, Li W, Lv S. A role of IL-17 in rheumatoid arthritis patients complicated with atherosclerosis. Front Pharmacol. 2022;13: 828933.
Smolen JS. Insights into the treatment of rheumatoid arthritis: a paradigm in medicine. J Autoimmun. 2020;110: 102425.
Zuo J, Tang J, Lu M, Zhou Z, Li Y, Tian H, Liu E, Gao B, Liu T, Shao P. Glycolysis rate-limiting enzymes: novel potential regulators of rheumatoid arthritis pathogenesis. Front Immunol. 2021;12: 779787.
Gabriel SE, Crowson CS. Risk factors for cardiovascular disease in rheumatoid arthritis. Curr Opin Rheumatol. 2012;24:171–6.
Yang H, Bian S, Chen H, Wang L, Zhao L, Zhang X, Zhao Y, Zeng X, Zhang F. Clinical characteristics and risk factors for overlapping rheumatoid arthritis and Sjogren’s syndrome. Sci Rep. 2018;8:6180.
Burggraaf B, van Breukelen-van der Stoep DF, de Vries MA, Klop B, Liem AH, van de Geijn GM, van der Meulen N, Birnie E, van der Zwan EM, van Zeben J, Castro Cabezas M. Effect of a treat-to-target intervention of cardiovascular risk factors on subclinical and clinical atherosclerosis in rheumatoid arthritis: a randomised clinical trial. Ann Rheum Dis. 2019;78:335-341.
Scherer HU, Haupl T, Burmester GR. The etiology of rheumatoid arthritis. J Autoimmun. 2020;110: 102400.
Zaiss MM, Joyce Wu HJ, Mauro D, Schett G, Ciccia F. The gut-joint axis in rheumatoid arthritis. Nat Rev Rheumatol. 2021;17:224–37.
Maeda Y, Takeda K. Host-microbiota interactions in rheumatoid arthritis. Exp Mol Med. 2019;51:1–6.
Zhang X, Zhang D, Jia H, Feng Q, Wang D, Liang D, Wu X, Li J, Tang L, Li Y, et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat Med. 2015;21:895–905.
Wells PM, Adebayo AS, Bowyer RCE, Freidin MB, Finckh A, Strowig T, Lesker TR, Alpizar-Rodriguez D, Gilbert B, Kirkham B, et al. Associations between gut microbiota and genetic risk for rheumatoid arthritis in the absence of disease: a cross-sectional study. Lancet Rheumatol. 2020;2:e418–27.
Yang W, Cong Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol Immunol. 2021;18:866–77.
Zheng D, Liwinski T, Elinav E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020;30:492–506.
Parada Venegas D, De la Fuente MK, Landskron G, Gonzalez MJ, Quera R, Dijkstra G, Harmsen HJM, Faber KN, Hermoso MA. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front Immunol. 2019;10:277.
Song H, Yoo Y, Hwang J, Na YC, Kim HS. Faecalibacterium prausnitzii subspecies-level dysbiosis in the human gut microbiome underlying atopic dermatitis. J Allergy Clin Immunol. 2016;137:852–60.
Martin R, Bermudez-Humaran LG, Langella P. Searching for the bacterial effector: the example of the multi-skilled commensal bacterium Faecalibacterium prausnitzii. Front Microbiol. 2018;9:346.
Bodkhe R, Balakrishnan B, Taneja V. The role of microbiome in rheumatoid arthritis treatment. Ther Adv Musculoskelet Dis. 2019;11:1759720X19844632.
de Paiva CS, Jones DB, Stern ME, Bian F, Moore QL, Corbiere S, Streckfus CF, Hutchinson DS, Ajami NJ, Petrosino JF, Pflugfelder SC. Altered mucosal microbiome diversity and disease severity in Sjogren syndrome. Sci Rep. 2016;6:23561.
Cantarel BL, Waubant E, Chehoud C, Kuczynski J, DeSantis TZ, Warrington J, Venkatesan A, Fraser CM, Mowry EM. Gut microbiota in multiple sclerosis: possible influence of immunomodulators. J Investig Med. 2015;63:729–34.
Fitzgerald CB, Shkoporov AN, Sutton TDS, Chaplin AV, Velayudhan V, Ross RP, Hill C. Comparative analysis of Faecalibacterium prausnitzii genomes shows a high level of genome plasticity and warrants separation into new species-level taxa. BMC Genomics. 2018;19:931.
Lopez-Siles M, Duncan SH, Garcia-Gil LJ, Martinez-Medina M. Faecalibacterium prausnitzii: from microbiology to diagnostics and prognostics. ISME J. 2017;11:841–52.
Sokol H, Pigneur B, Watterlot L, Lakhdari O, Bermudez-Humaran LG, Gratadoux JJ, Blugeon S, Bridonneau C, Furet JP, Corthier G, et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci U S A. 2008;105:16731–6.
Hu W, Lu W, Li L, Zhang H, Lee YK, Chen W, Zhao J. Both living and dead Faecalibacterium prausnitzii alleviate house dust mite-induced allergic asthma through the modulation of gut microbiota and short-chain fatty acid production. J Sci Food Agric. 2021;101:5563–73.
Cantoni C, Lin Q, Dorsett Y, Ghezzi L, Liu Z, Pan Y, Chen K, Han Y, Li Z, Xiao H, et al. Alterations of host-gut microbiome interactions in multiple sclerosis. EBioMedicine. 2022;76: 103798.
Quevrain E, Maubert MA, Michon C, Chain F, Marquant R, Tailhades J, Miquel S, Carlier L, Bermudez-Humaran LG, Pigneur B, et al. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut. 2016;65:415–25.
Gebrayel P, Nicco C, Al Khodor S, Bilinski J, Caselli E, Comelli EM, Egert M, Giaroni C, Karpinski TM, Loniewski I, et al. Microbiota medicine: towards clinical revolution. J Transl Med. 2022;20:111.
Lenoir M, Martin R, Torres-Maravilla E, Chadi S, Gonzalez-Davila P, Sokol H, Langella P, Chain F, Bermudez-Humaran LG. Butyrate mediates anti-inflammatory effects of Faecalibacterium prausnitzii in intestinal epithelial cells through Dact3. Gut Microbes. 2020;12:1–16.
Rossi O, van Berkel LA, Chain F, Tanweer Khan M, Taverne N, Sokol H, Duncan SH, Flint HJ, Harmsen HJ, Langella P, et al. Faecalibacterium prausnitzii A2–165 has a high capacity to induce IL-10 in human and murine dendritic cells and modulates T cell responses. Sci Rep. 2016;6:18507.
Zhang J, Huang YJ, Yoon JY, Kemmitt J, Wright C, Schneider K, Sphabmixay P, Hernandez-Gordillo V, Holcomb SJ, Bhushan B, et al. Primary human colonic mucosal barrier crosstalk with super oxygen-sensitive Faecalibacterium prausnitzii in continuous culture. Med (N Y). 2021;2(74–98): e79.
Huang XL, Zhang X, Fei XY, Chen ZG, Hao YP, Zhang S, Zhang MM, Yu YQ, Yu CG. Faecalibacterium prausnitzii supernatant ameliorates dextran sulfate sodium induced colitis by regulating Th17 cell differentiation. World J Gastroenterol. 2016;22:5201–10.
Wu SE, Hashimoto-Hill S, Woo V, Eshleman EM, Whitt J, Engleman L, Karns R, Denson LA, Haslam DB, Alenghat T. Microbiota-derived metabolite promotes HDAC3 activity in the gut. Nature. 2020;586:108–12.
Kim DS, Kwon JE, Lee SH, Kim EK, Ryu JG, Jung KA, Choi JW, Park MJ, Moon YM, Park SH, et al. Attenuation of rheumatoid inflammation by sodium butyrate through reciprocal targeting of HDAC2 in osteoclasts and HDAC8 in T cells. Front Immunol. 2018;9:1525.
Kang S, You HJ, Lee YG, Jeong Y, Johnston TV, Baek NI, Ku S, Ji GE. Production, structural characterization, and in vitro assessment of the prebiotic potential of butyl-fructooligosaccharides. Int J Mol Sci. 2020;21(2):445.
Schirmer M, Smeekens SP, Vlamakis H, Jaeger M, Oosting M, Franzosa EA, Ter Horst R, Jansen T, Jacobs L, Bonder MJ, et al. Linking the Human gut microbiome to inflammatory cytokine production capacity. Cell. 2016;167(1125–1136): e1128.
Pietrosimone KM, Jin M, Poston B, Liu P. Collagen-induced arthritis: a model for murine autoimmune arthritis. Bio Protoc. 2015;5(20):e1626.
Brand DD, Latham KA, Rosloniec EF. Collagen-induced arthritis. Nat Protoc. 2007;2:1269–75.
Asquith DL, Miller AM, McInnes IB, Liew FY. Animal models of rheumatoid arthritis. Eur J Immunol. 2009;39:2040–4.
Inglis JJ, Notley CA, Essex D, Wilson AW, Feldmann M, Anand P, Williams R. Collagen-induced arthritis as a model of hyperalgesia: functional and cellular analysis of the analgesic actions of tumor necrosis factor blockade. Arthritis Rheum. 2007;56:4015–23.
Firestein GS, McInnes IB. Immunopathogenesis of rheumatoid arthritis. Immunity. 2017;46:183–96.
van Delft MAM, Huizinga TWJ. An overview of autoantibodies in rheumatoid arthritis. J Autoimmun. 2020;110: 102392.
Kim DS, Woo JS, Min HK, Choi JW, Moon JH, Park MJ, Kwok SK, Park SH, Cho ML. Short-chain fatty acid butyrate induces IL-10-producing B cells by regulating circadian-clock-related genes to ameliorate Sjogren’s syndrome. J Autoimmun. 2021;119: 102611.
Zhou W, Zhang, Li Z, Jiang H, Li J, Ren R, Gao X, Li J, Wang X, Wang W, Yang Y. The fecal microbiota of patients with pancreatic ductal adenocarcinoma and autoimmune pancreatitis characterized by metagenomic sequencing. J Transl Med. 2021;19:215.
den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud DJ, Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013;54:2325–40.
Hoek M, Merks RMH. Emergence of microbial diversity due to cross-feeding interactions in a spatial model of gut microbial metabolism. BMC Syst Biol. 2017;11:56.
Sharon G, Garg N, Debelius J, Knight R, Dorrestein PC, Mazmanian SK. Specialized metabolites from the microbiome in health and disease. Cell Metab. 2014;20:719–30.
Hamer HM, Jonkers D, Venema K, Vanhoutvin S, Troost FJ, Brummer RJ. Review article: the role of butyrate on colonic function. Aliment Pharmacol Ther. 2008;27:104–19.
Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature. 2013;504:446–50.
Kespohl M, Vachharajani N, Luu M, Harb H, Pautz S, Wolff S, Sillner N, Walker A, Schmitt-Kopplin P, Boettger T, et al. The microbial metabolite butyrate induces expression of Th1-associated factors in CD4(+) T cells. Front Immunol. 2017;8:1036.
Martinez-Reyes I, Chandel NS. Mitochondrial TCA cycle metabolites control physiology and disease. Nat Commun. 2020;11:102.
Wang T, Fahrmann JF, Lee H, Li YJ, Tripathi SC, Yue C, Zhang C, Lifshitz V, Song J, Yuan Y, et al. JAK/STAT3-regulated fatty acid beta-oxidation is critical for breast cancer stem cell self-renewal and chemoresistance. Cell Metab. 2018;27(136–150): e135.
Ivanov II, Zhou L, Littman DR. Transcriptional regulation of Th17 cell differentiation. Semin Immunol. 2007;19:409–17.
Xu S, Cao X. Interleukin-17 and its expanding biological functions. Cell Mol Immunol. 2010;7:164–74.
Sun Y, Chen Q, Lin P, Xu R, He D, Ji W, Bian Y, Shen Y, Li Q, Liu C, et al. Characteristics of gut microbiota in patients with rheumatoid arthritis in Shanghai China. Front Cell Infect Microbiol. 2019;9:369.
Lukovac S, Belzer C, Pellis L, Keijser BJ, de Vos WM, Montijn RC, Roeselers G: Differential modulation by Akkermansia muciniphila and Faecalibacterium prausnitzii of host peripheral lipid metabolism and histone acetylation in mouse gut organoids. mBio. 2014;5(4):e01438–14.
Ouwerkerk JP, van der Ark KCH, Davids M, Claassens NJ, Finestra TR, de Vos WM, Belzer C. Adaptation of Akkermansia muciniphila to the Oxic-Anoxic Interface of the Mucus Layer. Appl Environ Microbiol. 2016;82:6983–93.
Figueiredo MC, Lobo SA, Sousa SH, Pereira FP, Wall JD, Nobre LS, Saraiva LM. Hybrid cluster proteins and flavodiiron proteins afford protection to Desulfovibrio vulgaris upon macrophage infection. J Bacteriol. 2013;195:2684–90.
Hevia A, Milani C, Lopez P, Cuervo A, Arboleya S, Duranti S, Turroni F, Gonzalez S, Suarez A, Gueimonde M, et al. Intestinal dysbiosis associated with systemic lupus erythematosus. mBio. 2014;5:e01548–01514.
Nehra A, Mehta RN, Encarnacion C. Bacteroides fragilis arthritis in a patient with sickle cell disease. Am J Med. 2001;111:161.
Rosenkranz P, Lederman MM, Gopalakrishna KV, Ellner JJ. Septic arthritis caused by Bacteroides fragilis. Rev Infect Dis. 1990;12:20–30.
Acknowledgements
None.
Funding
This research was supported by a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number HI20C1496). This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (No. NRF-2017M3A9F3041045 and RS-2023–00208207).
Author information
Authors and Affiliations
Contributions
JM, ARL, HK, MSP, GEJ, MLC, and SHP participated in the study design and data interpretation. JM wrote the original draft of manuscript. JM and ARL were involved in the mouse experiments and analysis. HK produced the F. prausnitzii and analyzed data of SFCA and microbial flora. JWC produced tissue data. JM, ARL and JYJ proceeded the revision. JM, ARL, HK, JYJ, SYL, YJ, GEJ MLC, and SHP interpreted the data and were involved in the critical revisions of the manuscript for important intellectual content. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The protocol for the study involving human participants was approved by the Institutional Review Board of Seoul St. Mary’s Hospital. All patients/participants provided written informed consent to participate in this study. The animal study protocol was approved by the Animal Research Ethics Committee of the Catholic University of Korea.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Moon, J., Lee, A.R., Kim, H. et al. Faecalibacterium prausnitzii alleviates inflammatory arthritis and regulates IL-17 production, short chain fatty acids, and the intestinal microbial flora in experimental mouse model for rheumatoid arthritis. Arthritis Res Ther 25, 130 (2023). https://doi.org/10.1186/s13075-023-03118-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13075-023-03118-3