
Abstract
Background
In Italy, malaria was endemic until the 1970s, when it was declared eradicated by WHO. Nowadays, with the persistence of competent mosquito populations, the effect of climate change, and increased possibility of importing malaria parasites from endemic counties due to growing migration, a malaria resurgence in Italy has become more likely. Hence, enhancing the understanding of the current distribution of the Anopheles maculipennis complex and the factors that influence the presence of this malaria vector is crucial, especially in Northern Italy, characterised by a high density of both human population and livestock.
Methods
To assess the presence and abundance of malaria vectors, a 4-year field survey in the plain areas of Lombardy and Emilia-Romagna region in Italy was conducted. Every sampling point was characterised in space by the land use in a 500-m radius and in time considering meteorological data collected in the short and long time periods before sampling. We combined the results of a linear regression model with a random forest analysis to understand the relative importance of the investigated niche dimensions in determining Anopheles mosquito presence and abundance.
Results
The estimated normalised variable importance indicates that rice fields were the most important land use class explaining the presence of Anopheles, followed by transitional woodlands and shrubland. Farm buildings were the third variable in terms of importance, likely because of the presence of animal shelters, followed by urbanised land. The two most important meteorological variables influencing the abundance of Anopheles in our study area were mean temperature in the 24 h before the sampling date and the sum of degree-days with temperature between 18 °C and 30 °C in the 14 days before the sampling date.
Conclusions
The results obtained in this study could be helpful in predicting the risk of autochthonous malaria transmission, based on local information on land cover classes that might facilitate the presence of malaria vectors and presence of short- and medium-term meteorological conditions favourable to mosquito development and activity. The results can support the design of vector control measures through environmental management.
Graphical Abstract

Similar content being viewed by others


Background
Climate change affects the transmission dynamics and geographic distribution of vector-borne diseases. Insect vector species are ectotherms, and the temperature increase affects their abundance, survival, and feeding activity as well as the pathogen development within the vector itself, reducing the duration of the incubation period [1]. The rapid expansion of global travel and trade increases the probability of travel by infected persons, who can be responsible for local outbreaks of severe diseases in non-endemic areas; the outbreaks of chikungunya in Italy in 2007 and 2017 [2, 3], dengue in Croatia [4], and West Nile virus in Romania in 1996 [5] are examples of the spreading capacity of these vector-borne diseases.
The increase of travellers and migratory flows to and from endemic areas make malaria the predominant imported infectious disease in non-endemic countries [6]. This is the case, for example, in Europe [7], which has fought malaria for centuries. However, malaria is still endemic in the coastal and inland plains of Asian areas of Turkey [8], and cases of local transmission in Greece are still reported [9]. Italy was one of the last countries where malaria was eradicated through a massive campaign treating infected human hosts and using effective insecticides. The combination of these efforts led to WHO's 1970 declaration that Italy is a malaria-free country [7]. Despite this, mosquito species of the genus Anopheles (which includes the main vectors of Plasmodium species responsible for human malaria) are still present in many suitable areas in Italy [8, 10,11,12,13]. In northern Italy, the Anophelinae of the Maculipennis complex is a group comprising both primary malaria vectors and species of low/negligible epidemiological importance. Recent studies have shown that the most abundant species of this complex in Pianura Padana, the main lowland of Italy, are Anopheles messeae, followed by An. maculipennis s.s., An. atroparvus, and An. melanoon [14].
Italian health institutions constantly monitor the possibility that an imported pathogen has activated a chain of local infections. The last episode of local transmission in Italy dates back to 1997 [15], but a local cluster of nosocomial malaria was reported in 2017 [6]. Malaria occurrence and epidemiological patterns are greatly influenced by environmental variables and land use patterns [16]. Multiple factors, including precipitation, temperature, altitude, and human population density, can act as risk components influencing the transmission process of malaria through sources of infection, transmission routes, and susceptible individuals [17]. Currently, only a few studies have been conducted to identify the broader environmental conditions influencing An. maculipennis complex vector presence and abundance in Italy as a risk factor for malaria transmission in this county [18]. For example, to evaluate the risk of malaria transmission in central Italy, Romi et al. [19] combined a multifactorial approach with climatic parameters. However, as pointed out by Boccolini et al. [6], there is a serious knowledge gap in an accurate and updated distribution map of Italian anopheline mosquitoes; this is especially relevant because of the recent climate and environmental changes, extending their geographical range beyond previous that of records.
This study aims to investigate the environmental factors (land use and meteorological variables) that mainly influence local presence and abundance of An. maculipennis complex in the Lombardy and Emilia-Romagna regions (Northern Italy) using data from the West Nile disease surveillance plan from 2017 to 2020. The entomological surveillance systems in the two Italian regions were based on the same attraction trap model and used the same sampling periodicity in summer. The Lombardy and Emilia-Romagna regions were characterised in the past by a high population abundance of Anopheles mosquitoes, and the environmental conditions are still locally suitable for the malaria vectors [14]. For example, Lombardy has 60% of Italy's rice fields, while Emilia-Romagna has a wide wetland area associated with the Po River Delta. Moreover, 24% of the Italian population (14.4 million people in an area of 46,354 km2) lives in these two regions. The results of our study can help to understand the population distribution of competent malaria vectors in northern Italy to plan better prevention and control activities for managing the risk of local transmission of malaria imported from endemic areas.
Methods
Study area
Samples of mosquitoes belonging to the Anopheles genus were retrieved from an entomological monitoring programme for the epidemiological surveillance of West Nile disease (WND) in Lombardy and Emilia-Romagna regions (Northern Italy). The study area covers approximately 28,000 km2 and involves a large part of the Po Valley, the most important Italian floodplain, characterised by intensive agriculture and animal husbandry. Annual crops are the most important component in the farming systems; vineyards, orchards, and horticulture are locally abundant. The two monitored regions are densely populated, with many cities, villages, and industrial areas. The eastern part of the Emilia-Romagna is located on the Adriatic Sea and is characterised by the presence of large natural wetlands (Po River Delta and Valli di Comacchio). The Po Valley is delimited by two mountain chains, the Apennines in the south and the Alps in the north, representing unsuitable areas for mosquitoes due to the altitude, although the valleys can act as important places for the mosquitoes to spread. The climate of the northern and southwestern part of the study area (Lombardy) is classified as humid continental; in the southeastern part (Emilia-Romagna), the climate is typically sub-continental, gradually becoming sub-Mediterranean towards the coastal part of the region.
Sampling design and mosquito collections
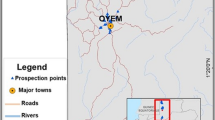
Mosquitoes were collected in a 4-year period from 2017 to 2020. The sampling area covers the whole plain area of the two regions, with some exceptions as found by traps placed in the province of Sondrio (Fig. 1a). The sampling area was subdivided in two regular grids: in 44 cells of 400 km2 each in Lombardy (Fig. 1a) and 95 cells of 110 km2 each in Emilia-Romagna (Fig. 1b). Traps were placed in sites considered of local risk (i.e. abundance of potential breeding sites) or in which previous West Nile virus circulation had been reported. Sampling sites were selected mainly in rural areas in Lombardy and semi-natural, rural, or peri-urban areas in Emilia-Romagna. Mosquitoes were trapped during the night from 17:00 until 9:00 every 2 weeks from June to September, using CDC attractive traps baited with dry ice as a source of carbon dioxide at fixed stations. In the trap catches Anopheles specimens were separated from other mosquitoes according to morphological taxonomic keys [10, 20].

Distribution of the sampling sites for the West Nile virus surveillance programme in A Lombardy and B Emilia-Romagna. Sampling points are represented by black crosses in Lombardy (above) and by orange dots in Emilia-Romagna (below)
The meteorological variables considered in the analysis were air temperature, dew point, and rainfall. Meteorological variables can impact mosquitoes’ life history parameters (e.g. development, survival, and fertility rates) and behavioural parameters (e.g. flight pattern, biting rate) influencing population abundance and activity. Therefore, we used two different time periods for the meteorological variables. To account for the influence of meteorological variables on life history parameters, a period of 14 days before the sampling date was considered (hereafter, ‘long term’). The meteorological variables in the 24-h period before the sampling (hereafter, ‘short term’) were considered to influence the mosquitoes' behaviour. Two derived meteorological variables were considered for their influence on mosquito life history. The first is defined as Tw and accounts for the suitable heat accumulation for the development and is calculated as the cumulative degree-days in the 14 days before the sampling event setting with the lower thermal threshold at 18 ℃ and the upper thermal threshold at 30 °C. The second variable is defined as Th and accounts for the thermal stress due to high temperature; it is calculated in terms of the cumulated degree-days equal to or above a threshold of 32 ℃ [21]. The meteorological and derived variables used in the study are detailed in Table 1.
Meteorological variables were extracted from the ERA5-Land database, which provides a reanalysed dataset at a high spatial (9 km grid) and temporal (hourly) resolution [22]. To obtain meteorological data at the specific sampling point and time, the ERA5-Land gridded data were interpolated on the sampling points using bilinear interpolation through Climate Data Operators (CDO) [23].
Land use
For each sampling site, we characterised the land use in a circular area with a 500-m radius [24], according to Taconet [25]. Data on land use were retrieved from two geodatabases, the DUSAF 6 (‘Destinazione d’Uso dei Suoli Agricoli e Forestali’) for the Lombardy region [26] and the ‘Coperture vettoriali uso del suolo di dettaglio’ for the Emilia-Romagna region [27]. Both geodatabases have a hierarchical structure with five nested classification levels. The first three levels are mainly based on the Corine Land Cover classification [28]; the fourth and fifth levels, those with greater detail, have been differently specified by the two regional systems. To proceed with the analysis, we created a common classification of land use types by combining the two regional classifications. We obtained 25 types of land use, detailed in the Additional materials (see Additional file 1: Table S1) (Table 2).
Statistical analysis
To investigate relationships between the abundance of Anopheles maculipennis complex and environmental (i.e. meteorological and land use) variables, we combined the simplicity and easiness of interpretation of linear regression (LR) with the high accuracy of the random forest analysis (RF) [29]. In both models, to account for the skewness of data, we considered the logarithm base 10 of the abundance of An. maculipennis complex in the site per sampling data as the dependent variable. Before fitting the regression models, we preliminary investigated the linear correlations between the environmental regressors and excluded variables presenting Pearson correlation coefficient S > 0.7. In the LR analysis, we applied the stepwise approach to select the environmental regressors according to the Akaike information criterion (AIC). We studied multicollinearity among regressors using variance inflation factor (VIF), excluding variables with VIF value > 5 to eradicate strong dependence among regressors.
The variable importance measures, i.e. the relative influence of a regressor in predicting the response variable [30], of LR and RF are used to merge the results of the two models. First, we normalised the variable importance estimated within each model. The highest value was set equal to 100, and other values were proportionally rescaled, with variable importance for not used regressors set equal to 0. Then, we considered the average between LR and RF values for each regressor. All the statistical analyses were performed in the R environment [31], using functions lm for LR and randomForest (package randomForest) [32] for RF.
Results
In the 4-year of survey period, a total of 1,365,958 mosquitoes of different species were collected (264,453 in Lombardy and 1,101,505 in Emilia-Romagna), of which 18,637 females belonged to the An. maculipennis complex (16,289 in Lombardy and 2348 in Emilia-Romagna). Only samplings with presence of at least a single An. maculipennis complex mosquito (positive samplings) were included in the analysis. The original dataset is related to the West Nile disease surveillance plan; unfortunately, in this context, not all the observations with a 0 abundance of Anopheles were reported in the dataset. Consequently, we had to remove the few available observations for consistency. In total, for 3263 total samplings, there were 859 positive samples (26.3%).
The number of Anopheles caught was much higher in Lombardy than in Emilia-Romagna; this could be explained by the large representation of farms in the sampling locations in Lombardy where the typical farm combines crop area and animal husbandry, which are particularly attractive for An. maculipennis complex.
The descriptive statistics of the land use surrounding the sampling sites and the meteorological variables are reported in Tables 3 and 4. Based on the analysis of the correlation matrix, we (i) removed four meteorological variables (Hs, Tl, Hl, Th), (ii) merged ‘broad-leaf forest’, ‘coniferous forest’, and ‘mixed forests are joined’ in the new land use category ‘forest’, and (iii) merged ‘inland marshes’ and ‘salt marshes’ in the new land use category ‘wetlands’. Therefore, we finally obtained 25 regressors (4 meteorological variables and 21 land use categories) for the LRM analysis and the RF.
The linear regression model with stepwise procedure selected 13 regressors (Table 5) as those statistically significant to explain the variability of the log abundance of An. maculipennis complex females in the samplings (the adjusted R2 is 0.2096).
Four regressors had a negative estimated coefficient (‘road and rail networks and associated land’, water courses, pastures, and Rs), indicating a negative relationship between the regressor and average expected abundance of An. maculipennis complex. The p-values are all < 0.1, with eight p-values < 0.01 and three p-values < 0.05, so we can consider all estimated coefficients to be statistically different from 0. According to the normalised variable importance (VI norm), the most important regressor is ‘rice fields’.
The random forest model showed a better fitting to the data than the linear regression model, with an explained variance equal to 42.89%. The normalised variable importance for the random forest model is reported in Table 6. ‘Rice fields’ is the most important variable in explaining the variability of the target variable (in accordance with the linear model), followed by ‘transitional woodland shrub environment’ and two meteorological regressors, Ts and ‘Tw. Only three regressors (‘beaches, dunes, sand plains’, ‘airports’, and ‘port areas’) have a normalised variable importance equal to 0, indicating that they are not selected by the model to explain the variability in the abundance of An. maculipennis complex mosquitoes.
To integrate the results of the linear regression and the non-linear regression (random forest) model, the overall variable importance is calculated (Fig. 2). The overall variable importance is set equal to 0 for regressors not used by both models (‘beaches, dunes, sand plains’ and ‘airports’) and equal to 100 for the most important regressor selected by both models (‘rice fields’). ‘Transitional woodland shrub environment’ was the second most important variable. Then, we identified three groups of regressors: the medium importance (6 variables with overall variable importance between 35 and 50), low importance (5 variables with overall variable importance between 12 and 25), and very low importance (9 variables with overall variable importance between 1 and 10). Results are reported in Fig. 2.

Overall variable importance estimates for Anopheles abundance in the study area combining results of the linear regression and random forest for the set of variables investigated. The orange rectangles cluster variables with similar importance
The two single-model rankings of variable importance showed good accordance, except for ‘farm buildings’, classified as the eighth most influential regressor in the random forest analysis (normalised VI = 14) and second in the linear regression model (normalised V = 84), and for ‘roads and rail networks and associated land’, which had a high normalised importance in the linear regression analysis and a low one in the random forest analysis (68 and 8, respectively).
Discussion
Our analysis identified a clear ranking in the importance of the variable investigated and allowed grouping them in four classes: high, medium, low, and very low importance. There was also a fifth class grouping the variables with no importance in our study area.
In the high importance class, we reported the presence of rice fields in the surroundings of the sampling site as the most critical factor positively influencing the abundance of An. maculipennis complex collected. This result is consistent with several studies considering rice fields as the main breeding site for Anopheles in Italy [33,34,35], and it supports previous findings demonstrating that the abundance of the Maculipennis complex is particularly high in sites characterised by extensive breeding grounds, such as rice fields and wetlands [14]. Both regression models highlighted the great importance of transitional woodland and shrub habitats in positively influencing the presence of Anopheles. This result is consistent with similar findings, as the one reported in Mali, where removing a flowered invasive shrub resulted in a 69.4% drop in mosquito population density [36]. Multiple factors could be involved in explaining our observations. In the study area, transitional woodlands and shrubs often represent the interface between the rural and urban environments. These habitats can guarantee good resting sites to adult mosquitoes and are frequently and rapidly colonised by plants rich in sugars, such as nectar and gland exudates (e.g. Ailanthus altissima and Robinia pseudoacacia), which can serve as a trophic source for the Anopheles mosquito.
In the medium importance class, we observed a positive relation between the abundance of Anopheles and the presence of ‘farm buildings’, in line with the well-known attractiveness of bovine herds for malaria vectors. In Ethiopia [37] and Malawi [38], it has been observed that proximity of cattle increases the abundance of Anopheles. In Europe, the importance of bovines was confirmed by a survey in a former endemic area in Romania, which found that 85.5% of captured mosquitoes fed on cattle [39]. Similar results (82.4% cattle, 5.1% human) have been obtained in a study performed in Corsica (France) [40]. Animal shelters are also used as resting sites. In previous studies, over a hundred females [40], up to 500 [41], were found in animal shelters. Although our results are in line with the literature, the importance attributed to ‘farm buildings’ in our study could be partially overestimated by the location of sampling points in the Lombardy region that were close to animal shelters.
In our study, thermal sum in the optimal range for development Tw (the sum of degree-days with temperature between 18 ℃ and 30 ℃ in the 14-day period before the sampling date) shows high variable importance. This is in accordance with literature where the importance of the thermal sum in the optimal range temperatures is confirmed for the development of eggs [42] and larval stages [43]. The mean temperature in the 24 h before the sampling, Ts, is also a positive factor in our study since higher temperatures cause an increase in mosquito activity. The cumulative rain in the 24 h before the sampling (Rs) was a negative factor affecting the probability of trapping mosquitoes. Although mosquitoes can fly during rain [44], some studies show that as rainfall increases the number of mosquitoes sampled decreases [45].
The presence of urban areas in the surroundings of the sampling points increases the probability of catching An. maculipennis complex. This is consistent with other studies in which a high abundance of Anopheles has been found even in cities, as in a survey in The Netherlands where An. maculipennis occurred in similar proportions in urban and agricultural areas [46].
The study identifies rails and railroads as disturbing elements that reduce the expected abundance of Anopheles. In other studies, they have been found to be a positive factor since roads usually have drainage areas along their margins that become transient water bodies during and after rain. The possible use of these areas as breeding sites was investigated in Ecuador, and they hosted species of the Anopheles genus such as An. punctimacula and An. albimanus [47]. This positive effect was not recorded in our study area.
Rivers can be used as breeding nesting sites by Anopheles, particularly in areas with slow water flow and the presence of riparian vegetation [43], as reported for example in surveys in Ecuador [47] and in Italy by Romi et al. [33], which found different species of Anopheles in riverine areas. These favourable riparian habitats are present in the study area; however, rivers are characterised by a negative coefficient in the regression analysis. A possible explanation for this result is the level of the pollutants characterising many rivers in the study area, which show low to moderate concentrations of heavy metals and organic micropollutants and high contents of nutrients causing eutrophication, as reported in Provini and Binelli [48]. Although in the literature Anopheles seems to show an increasing tolerance for organic pollutants [49], the contamination level of the river in the study area can be higher than its tolerance threshold.
The low importance attributed to port areas, in our study area fluvial ports, and the positive regression coefficient, in contrast to the negative coefficient of riverine areas, can be attributed to the fact that port areas are only present in a small fraction of our study area (< 0.01%).
In the low importance class, we observed a positive relationship between permanent crops and Anopheles presence, in agreement with other studies [41]. An example is the research conducted in Ghana [50], which found many suitable larval habitats in farmlands and pastures with abundant ditches, furrows, and drinking troughs. Similar results were found in a survey in The Netherlands [46]. In our study, pastures were a negative coefficient, which can be attributed to the small spatial extension of this area since in our study area intensive animal husbandry is mainly practised in shelters.
Our results show a positive moderate variable importance for graveyards in which there are constant artificial water reservoirs such as flower pots [51]. A review found that cemeteries are perfect breeding grounds for artificial container-breeding mosquitoes [52], which is not the case for Anopheles, even though An. maculipennis s.s. is considered more adapted to artificial environments than other species in the group [46].
In the very low importance class, we observed a cluster of very different variables. ‘Green urban areas’ belong to this class, although a meta-analysis on the abundance of mosquitoes comparing urban environments and green urban areas showed no significant difference between the two [53].
The very low importance of wetlands, although considered one of the most suitable habitats for Anopheles mosquitoes, can be attributed to the small presence of wetlands in our study and to the salinity of coastal wetlands, which characterises most of the wetlands close to the sampling points and can be above the tolerance threshold for An. maculipennis [40].
Airports, beach dunes, and plains were land use classes of no importance in our study because of their high artificial or natural draining capacity.
Conclusions
Serious knowledge gaps were mainly found in entomological risk assessment due to (i) lack of accurate and up-to-date distribution maps of Italian Anopheline mosquitoes, which have most likely changed their geographic range in recent decades because of environmental and climatic changes; (ii) limited knowledge on the susceptibility of current Anopheline populations to infection by different species and geographic populations of imported Plasmodium [6].
Our study provides data on the distribution and abundance of Anopheline species in two extensive regions of northern Italy, contributing to filling the knowledge gaps on malaria vectors in this area. The results we obtained allowed us to characterise environmental drivers influencing the presence and the abundance of the An. maculipennis complex. Quantitative information could help with the development of models predicting the risk of transmission of possible pathogens carried by this group of insects, including malaria, by knowing land cover types and the current status and past trends of meteorological variables. The results could also be applied to the design and implementation of vector control measures through environmental management methods to reduce the risk of human-mosquito contact.
Availability of data and materials
The data supporting the findings of the study must be available within the article and/or its supplementary materials, or deposited in a publicly available database.
References
Rocklöv J, Dubrow R. Climate change: an enduring challenge for vector-borne disease prevention and control. Nat Immunol. 2020;21:479–83.
Liumbruno GM, Calteri D, Petropulacos K, Mattivi A, Po C, Macini P, et al. The chikungunya epidemic in Italy and its repercussion on the blood system. Blood Transfus. 2008;6:199–210.
Rezza G. Chikungunya is back in Italy: 2007–2017. J Travel Med. 2018. https://doi.org/10.1093/jtm/tay004.
Gjenero-Margan I, Aleraj B, Krajcar D, Lesnikar V, Klobučar A, Pem-Novosel I, et al. Autochthonous dengue fever in Croatia, August-September 2010. Euro Surveill Bull Eur Sur Mal Transm Eur Commun Dis Bull. 2011;16:19805.
Romanca C, Vladimirescu A, Tsai TF, Ceianu C, Karabatsos N, Lanciotti R, et al. Entomologic and avian investigations of an epidemic of West Nile fever in Romania in 1996, with serologic and molecular characterization of a virus isolate from mosquitoes. Am J Trop Med Hyg. 1999;61:600–11.
Boccolini D, Menegon M, Di Luca M, Toma L, Severini F, Marucci G, et al. Non-imported malaria in Italy: paradigmatic approaches and public health implications following an unusual cluster of cases in 2017. BMC Public Health. 2020;20:857.
Martini M, Angheben A, Riccardi N, Orsini D. Fifty years after the eradication of Malaria in Italy. The long pathway toward this great goal and the current health risks of imported malaria. Pathog Glob Health. 2021;115:215–23.
Alten S, Kampen H, Fontenille D. Malaria in Southern Europe: resurgence from the past? Emerg pests vector-borne Dis Eur. 2007. Disponibile su: https://avesis.hacettepe.edu.tr/yayin/b8aabc59-ef48-441b-900e-89ac43a9e035/malaria-in-southern-europe-resurgence-from-the-past. Accessed 22 Mar 2023.
European Centre for Disease Prevention and Control. Update on autochthonous Plasmodium vivax malaria in Greece. 2011. https://www.ecdc.europa.eu/sites/default/files/media/en/publications/Publications/131003_TER_Malaria_Greece_Risk_Assessment.pdf
Becker N, Petric D, Zgomba M, Boease C, Dahl C, Lane J, et al. Mosquitoes and their control. New York, Boston, Dordrecht, London, Moscow: Kluwer Academics/Plenum Publishers. 2003. https://doi.org/10.1007/978-3-540-92874-4. Accessed 22 Mar 2023.
Ramsdale CD, Alten B, Caglar SS, Ozer N. A revised, annotated checklist of the mosquitoes (Diptera, Culicidae) of Turkey. Eur Mosq Bull. 2001;9:18–27.
Ramsdale C, Snow K. Distribution of the genus Anopheles in Europe. Eur Mosq Bull. 2000. https://www.semanticscholar.org/paper/Distribution-of-the-genus-Anopheles-in-Europe.-Ramsdale-Snow/bc2461c1b4a14074d8eb497ac473f7da00e2306e. Accessed 22 Mar 2023.
Schaffner F, Angel G, Geoffroy B, Hervey J, Rhaiem A, Brunhes J. The mosquitoes of Europe / Les moustiques d’Europe: an identification and training programme. IRD Editions & EID Mediterranee, Montpellier, France; 2001. https://www.nhbs.com/the-mosquitoes-of-europe-les-moustiques-deurope. Accessed 22 Mar 2023.
Calzolari M, Desiato R, Albieri A, Bellavia V, Bertola M, Bonilauri P, et al. Mosquitoes of the Maculipennis complex in Northern Italy. Sci Rep. 2021;11:6421.
Baldari M, Tamburro A, Sabatinelli G, Romi R, Severini C, Cuccagna G, et al. Malaria in Maremma, Italy. Lancet. 1998;351:1246–7.
Ren Z, Wang D, Ma A, Hwang J, Bennett A, Sturrock HJW, et al. Predicting malaria vector distribution under climate change scenarios in China: challenges for malaria elimination. Sci Rep. 2016;6:20604.
Song Y, Ge Y, Wang J, Ren Z, Liao Y, Peng J. Spatial distribution estimation of malaria in northern China and its scenarios in 2020, 2030, 2040 and 2050. Malar J. 2016;15:345.
Lee EH, Miller RH, Masuoka P, Schiffman E, Wanduragala DM, DeFraites R, et al. Predicting risk of imported disease with demographics: geospatial analysis of imported malaria in Minnesota, 2010–2014. Am J Trop Med Hyg. 2018;99:978–86.
Romi R, Boccolini D, Vallorani R, Severini F, Toma L, Cocchi M, et al. Assessment of the risk of malaria re-introduction in the Maremma plain (Central Italy) using a multi-factorial approach. Malar J. 2012;11:98.
Severini F, Toma L, Di Luca M, Romi R. Italian mosquitoes: general information and identification of adults (Diptera, Culicidae)/Le Zanzare Italiane: Generalità E Identificazione Degli Adulti (Diptera, Culicidae). Fragm Entomol. 2009; 213–372
Bayoh MN, Lindsay SW. Effect of temperature on the development of the aquatic stages of Anopheles gambiae sensu stricto (Diptera: Culicidae). Bull Entomol Res. 2003;93:375–81.
Muñoz-Sabater J, Dutra E, Agustí-Panareda A, Albergel C, Arduini G, Balsamo G, et al. ERA5-Land: a state-of-the-art global reanalysis dataset for land applications. Earth Syst Sci Data. 2021;13:4349–83.
Schulzweida U. CDO User Guide. 2019. https://zenodo.org/record/3539275. Accessed 22 Mar 2023.
Dambach P, Sié A, Lacaux JP, Vignolles C, Machault V, Sauerborn R. Using high spatial resolution remote sensing for risk mapping of malaria occurrence in the Nouna district, Burkina Faso. Glob Health Action. 2009;2:2094.
Taconet P, Porciani A, Soma DD, Mouline K, Simard F, Koffi AA, et al. Data-driven and interpretable machine-learning modeling to explore the fine-scale environmental determinants of malaria vectors biting rates in rural Burkina Faso. Parasit Vectors. 2021;14:345.
Regione Lombardia. Destinazione Uso e copertura del suolo 2018 (DUSAF 6.0). 2018.
Geoportale Regione Emilia Romagna. Coperture Vettoriali Uso Del Suolo Di Dettaglio—Edizione 2020. 2020.
Büttner G. CORINE land cover and land cover change products. In: Manakos I, Braun M, editors. Land use and land cover mapping in Europe: practices & trends. Dordrecht: Springer Netherlands; 2014 [citato 28 maggio 2022]. p. 55–74. (Remote sensing and digital image processing). https://doi.org/10.1007/978-94-007-7969-3_5
Breiman L. Random forests. Mach Learn. 2001;45:5–32.
Wei P, Lu Z, Song J. Variable importance analysis: a comprehensive review. Reliab Eng Syst Saf. 2015;142:399–432.
R Core Team. R: a language and environment for statistical computing. Vienna: R foundation for Statistical Computing. 2022. https://www.R-project.org/. Accessed 26 Dec 2020.
Liaw A, Wiener M. Classification and regression by random forest. R News. 2002;218–22.
Romi R, Pierdominici G, Severini C, Tamburro A, Cocchi M, Menichetti D, et al. Status of malaria vectors in Italy. J Med Entomol. 1997;34:263–71.
Zhao TY, Xue RD. Integrated mosquito management in rice field in China. Wetl Ecol Manag. 2022;30:963–73.
Bettini S, Gradoni L, Cocchi M, Tamburro A. Rice culture and Anopheles labranchiae in Central Italy. Geneva: World Health Organisation; 1978. Report No.: WHO/VBC series, 78.686, WHO/MAL/ 78.897. WHO.
Muller GC, Junnila A, Traore MM, Traore SF, Doumbia S, Sissoko F, et al. The invasive shrub Prosopis juliflora enhances the malaria parasite transmission capacity of Anopheles mosquitoes: a habitat manipulation experiment. Malar J. 2017;16:237.
Zeru MA, Shibru S, Massebo F. Exploring the impact of cattle on human exposure to malaria mosquitoes in the Arba Minch area district of southwest Ethiopia. Parasit Vectors. 2020;13:322.
Mburu MM, Zembere K, Mzilahowa T, Terlouw AD, Malenga T, Van Den Berg H, et al. Impact of cattle on the abundance of indoor and outdoor resting malaria vectors in southern Malawi. Malar J. 2021;20:353.
Fălcuţă E, Prioteasa FL, Teodorescu I, Nicolescu G. Anopheles maculipennis complex (Diptera: Culicidae) in Comana area (Giurgiu county, Romania). Romanian J Biol. 2011.
Toty C, Barré H, Le Goff G, Larget-Thiéry I, Rahola N, Couret D, et al. Malaria risk in Corsica, former hot spot of malaria in France. Malar J. 2010;9:231.
European Centre for Disease Prevention and Control. Anopheles labranchiae—factsheet for experts. 2017. https://www.ecdc.europa.eu/en/disease-vectors/facts/mosquito-factsheets/anopheles-labranchiae
Kettle DS, Sellick G. The duration of the egg stage in the races of Anopheles maculipennis meigen (Diptera, Culicidae). J Anim Ecol. 1947;16:38.
Jetten TH, Takken W. Anophelism without malaria in Europe: a review of the ecology and distribution of the genus Anopheles in Europe. Wageningen, The Netherlands: Wageningen Agricultural University; 1994. 69 (Wageningen Agricultural University papers).
Dickerson AK, Shankles PG, Madhavan NM, Hu DL. Mosquitoes survive raindrop collisions by virtue of their low mass. Proc Natl Acad Sci. 2012;109:9822–7.
Okorie P, Popoola KOK, Awobifa O, Kolade I, Ademowo G. Species composition and temporal distribution of mosquito populations in Ibadan, Southwestern Nigeria. J Entomol Zool Stud. 2014;2:164–9.
Ibáñez-Justicia A, Smitz N, Blom R, Vanderheyden A, Jacobs F, Meganck K, et al. Anopheles maculipennis complex in The Netherlands: first record of Anopheles daciae (Diptera: Culicidae). Diversity. 2022;14:636.
Pinault LL, Hunter FF. Larval habitat associations with human land uses, roads, rivers, and land cover for Anopheles albimanus,A. pseudopunctipennis, and A. punctimacula (Diptera: Culicidae) in Coastal and Highland Ecuador. Front Physiol. 2012 [citato 18 maggio 2023]. https://doi.org/10.3389/fphys.2012.00059/abstract
Provini A, Binelli A. Environmental quality of the Po River Delta. In: Wangersky PJ, curatore. Estuaries. Berlin/Heidelberg: Springer-Verlag; 2006 [citato 18 maggio 2023]. p. 175–95. (The handbook of environmental chemistry; vol. 5H). https://doi.org/10.1007/698_5_028
Mattah PAD, Futagbi G, Amekudzi LK, Mattah MM, de Souza DK, Kartey-Attipoe WD, et al. Diversity in breeding sites and distribution of Anopheles mosquitoes in selected urban areas of southern Ghana. Parasit Vectors. 2017;10:25.
Hinne IA, Attah SK, Mensah BA, Forson AO, Afrane YA. Larval habitat diversity and Anopheles mosquito species distribution in different ecological zones in Ghana. Parasit Vectors. 2021;14:193.
Rydzanicz K, Czułowska A, Dyczko D, Kiewra D. Assessment of mosquito larvae (Diptera: Culicidae) productivity in urban cemeteries in Wroclaw (SW Poland). Int J Trop Insect Sci. 2021;41:3249–55.
Vezzani D. Review: artificial container-breeding mosquitoes and cemeteries: a perfect match: mosquitoes and cemeteries. Trop Med Int Health. 2007;12:299–313.
Rhodes CG, Scavo NA, Finney M, Fimbres-Macias JP, Lively MT, Strauss BH, et al. Meta-analysis of the relative abundance of nuisance and vector mosquitoes in Urban and blue-green spaces. Insects. 2022;13:271.
Acknowledgements
The authors thank Monica Cerioli of the Osservatorio Lombardia for the cartographical material.
Funding
This study was supported by the Ministry of Health, PRC 2018012.
Author information
Authors and Affiliations
Contributions
Conceptualisation, GG, AL, MC, FD and AS; methodology, SC. FD, MC; formal analysis, AS, GG, MM; data curation, SC FD, MC; writing—original draft preparation, AH, FD; writing—review and editing, AH, GG, MC, FD and AL; project administration, D.L. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study does not involve human participants, material, or data.
Consent for publication
This work contains no material related to individual persons.
Competing interests
The authors declares no competing interests for the publication of this research paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Classification of land use categories.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gilioli, G., Defilippo, F., Simonetto, A. et al. Characterization of environmental drivers influencing the abundance of Anopheles maculipennis complex in Northern Italy. Parasites Vectors 17, 109 (2024). https://doi.org/10.1186/s13071-024-06208-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06208-6