
Abstract
Background
Lyme borreliosis is the most common tick-borne disease in Europe and is often caused by Borrelia afzelii, which is transmitted by Ixodes ricinus ticks. The prevalence and abundance of infected ticks fluctuate in time and space, influencing human infection risk. Rodents are reservoir hosts for B. afzelii and important feeding hosts for larval ticks. In the study reported here, we examined how variation in rodent abundance is associated with B. afzelii infection prevalence in ticks, the density of nymphs (DON) and the density of infected nymphs (DIN) in the following year. We further analysed the relationships between the abundance of infected rodents and nymphal infection prevalence (NIP) and DIN.
Methods
We conducted a study that combined experimental and observational approaches on 15 islands (10 small islands and 5 large islands) in Finland. On all of the islands, ticks and rodents were monitored and sampled during the summer of 2019, with the monitoring of tick abundance and sampling continuing into the spring of 2020. On five of the 10 small islands, captured rodents were removed from the island (“removal” islands), and on the other five small islands, captured rodents were released back to the trapping site after marking and sampling (“control” islands). On the five large islands, captured rodents were released back to the trapping site after marking and sampling. The presence of B. afzelii from nymph and rodent samples was examined.
Results
The results of the experimental study showed that neither treatment (removal), rodent abundance index nor abundance index of infected rodents in 2019 was associated with DON, NIP or DIN in 2020. Based on data from the observational study, the NIP in 2020 decreased with increasing rodent abundance index and abundance index of infected rodents in 2019. However, the DIN in 2020 was not associated with the rodent abundance index or the abundance index of infected rodents in 2019. In addition, in the observational study, DON in 2020 increased with increasing rodent abundance index.
Conclusions
Our results suggest that low rodent abundance during the tick activity period is not sufficient for reducing the disease hazard and, hence, rodent removal may not be a feasible control measure in natural ecosystems.
Graphical abstract

Similar content being viewed by others


Background
Tick-borne diseases are a source of serious health concerns in Europe [1,2,3]. The most common tick-borne disease in the Northern Hemisphere is Lyme borreliosis (LB) [4, 5]. LB is caused by spirochete bacteria belonging to the Borrelia burgdorferi sensu lato (B. burgdorferi s.l.) complex that are transmitted among vertebrate hosts by ixodid ticks [6]. Ixodes ricinus is the main vector of B. burgdorferi s.l. in Europe [7]. The life stages of this tick differ in terms of host preferences, with rodents being common hosts for larvae, nymphs parasitizing a wider range of hosts and adults preferring large hosts, such as deer [8,9,10]. The nymphal stage is considered to be the life stage posing the highest risk to humans due to its small size of nymphs, their potentially high abundance and the potential for nymphs to carry pathogens [11]. Therefore, the local abundance of infected nymphs is considered to be a primary environmental risk for humans [12, 13].
Rodents not only provide a blood-meal source for immature I. ricinus ticks but also contribute to the propagation of horizontally transmitted tick-borne pathogens; uninfected larval ticks may acquire pathogens from infected reservoir rodents that acquire the infection from infected nymphs [14,15,16]. In Europe, one of the most common etiological agents of Lyme borreliosis is Borrelia afzelii (a genospecies of B. burgdorferi s.l.), which is maintained by rodent hosts such as mice and voles [15, 17,18,19,20,21,22,23,24]. The role of rodent abundance in the prevalence of B. burgdorferi s.l. infection in nymphs has been examined in several studies carried out in North America [25,26,27] and Central Europe [28, 29], with the results suggesting that a higher abundance of reservoir rodents translates into a higher infection prevalence in nymphs the following year. However, the Lyme disease system in North America and Central Europe differs from the system in Northern Fennoscandia in many ways. For example, occasional masting increases rodent densities in temperate Europe, whereas in northern Fennoscandia, vole-dominated rodent populations show seasonal and multiannual density fluctuations characterized by very low densities over extended periods (over the breeding season) [30, 31]. It remains to be quantified whether these very low rodent densities affect the dynamics of rodent-associated tick-borne pathogens and infection prevalence in ticks, as is expected because the rodent density fluctuations have been shown to translate into human Lyme disease incidence with time delays in northern Europe [32, 33].
Most studies addressing the role of rodent density in determining the prevalence of B. burgdorferi s.l. in ticks are observational or theoretical in design [26, 27, 33, 34]. Only recently has an experimental study, conducted in small outdoor enclosures in temperate Europe [29], shown that an increase in rodent density increases the prevalence of rodent-associated tick-borne pathogens in ticks the following year. Our study aimed to quantify whether rodent density variations, especially low rodent abundance, are associated with changes in B. afzelii infection prevalence in ticks the following year. In our study, we utilized a natural island system and incorporated both experimental and observational study designs. We investigated the associations between rodent abundance and the abundance of infected rodents in year one (t) and the density of nymphs (DON), the prevalence of B. afzelii in nymphs (referred to as nymph infection prevalence [NIP]) and the density of infected nymphs (DIN) in the subsequent year (t+1). We hypothesized that: (i) rodent removal decreases NIP the following year (NIPt+1) and (ii) the higher the rodent abundance index or abundance index of infected rodents, the higher NIPt+1. We also predicted that: (iii) a higher rodent abundance index will also result in a higher density of nymphs in the following year (DONt+1) because rodents are considered to be important feeding hosts of larvae. Consequently, we expect that: (iv) the rodent abundance index would increase the DIN the following year (DINt+1), which in turn determines the level of B. afzelii hazard to humans.
Methods
Study area and design
The study was conducted on 15 islands in the Porvoo archipelago in southern Finland, of which 10 islands were small (0.5–4 ha) and five were large (63–966 ha) (Fig. 1). An experimental study was conducted on the 10 small islands which consisted of: (i) the removal of all captured rodents from five of the islands (treatment: removal; designated “removal” islands); and (ii) the release of all captured rodents back to the trapping site (approximately) after sampling and marking on the remaining five small islands (designated “control” islands). Each pair of small islands was randomly allocated to removal and control treatments before the study. The observational study was carried out on the five small “control” islands and the five large islands where the rodents were released after the sampling and marking. Tick collections were also carried out on all islands during the same time period and also during the following year in the spring. An overview of the study design is shown in Fig. 2. Island size, study area and tick collection information are available in Additional file 1: Table S1 and Additional file 1: Table S2. Dung surveys were also carried out at the same time as rodent trappings to estimate the abundance of cervids (Additional file 2: Text S1); however, these surveys had no significant association with the study design, and the dung data are addressed only in the additional files (Additional file 1: Table S3; Additional file 1: Table S4) and the dung index is not included in other models (see section Statistical Analyses).

Map of the study sites showing rodent removal, control and large islands. Grey islands have not been included in the study

Study design including a collection of ticks and rodents in the experimental (yellow) and observational (green) studies to examine the associations between rodents, ticks and Borrelia afzelii. The crossover indicates that no rodent trapping was done in May 2020. The distance in meters indicates the tick collection distance on the islands
Rodent trapping
Rodents were trapped in May, June, July and late August/early September in 2019 using Ugglan live traps (Grahnab AB, Gnosjö, Sweden). The number of trapping squares (4 traps per square, with an inter-trap distance of 20 m) used depended on the island size. On the small islands, we used between two and seven trapping squares (ntraps = 8–28 per island) to cover the whole island area, while on large islands we used four trapping squares (ntraps = 16 per island) placed 50–100 m apart from each other; see Additional file 1: Table S1 for more details. Traps baited with sunflower seeds and potatoes were set in the late afternoon at pre-marked locations on each island and checked on two consecutive mornings. All trapping was done within approximately 1 week (May–July: 6 days; August/September: 10 days). In total, we captured three rodent species: bank vole (Clethrionomys glareolus), field vole (Microtus agrestis) and yellow-necked mouse (Apodemus flavicollis). We calculated the rodent abundance index per 100 traps as the sum of bank voles and field voles relative to the trapping effort on the islands (the number of traps per island multiplied by the 2 trapping nights) × 100 to control for the differences in trapping effort between islands. Bank voles and field voles were chosen because they were the most common rodent species on the islands and as their presence varied on different islands, with some islands not having any field voles or bank voles. Yellow-necked mice were not included in the analysis due to the low number of individuals trapped across all islands (n = 3).
We determined the species of each rodent and recorded the individual’s characteristics and presence of ectoparasites. An ear biopsy (diameter: 2 mm), taken from one ear of each animal for B. afzelii screening, was stored in 70% ethanol at - 20 °C until further processing. Animals captured on the control and large islands were marked with a microchip (Trovan Unique™; Microchips Australia, Keysborough, VIC, Australia) at the first capture and released near their trapping location after sampling. The same data and samples were taken from recaptured animals in the following trapping months (sessions). All animals captured on removal islands were euthanized using cervical dislocation after data collection and sampling.
Tick collection
Ticks were collected during all rodent trapping sessions and in May 2020. In Finland, tick activity is absent during the winter frost and snow periods. In Finland, depending on the region, life stage and biotope, I. ricinus is mostly active from April/May to October/November [35,36,37,38]. Both I. ricinus larvae and nymphs have been found to be abundant in vegetation in May–June and in August–September in southwest and central Finland [35, 38]. Thus, most of the activity period of the larvae and nymphs was covered by the study period. However, this was the first study to observe the activity dynamics of I. ricinus on this island system.
Ticks were collected by dragging a 1-m2 cotton flannel flag through the vegetation. After every 10 m, the flag was checked for ticks, and all ticks on the flag were recorded, removed, placed in 70% ethanol and stored at − 20 °C until further processing. The dragging distance depended on the size of the island and the session. On the large islands, the dragging distance was 400 m in all sampling sessions and on all islands in May 2019 and 2020. On the small islands in other sessions (July–August/September), the dragging distance was 100 m to avoid the removal of too many ticks, thus potentially affecting tick abundance and tick-borne pathogen circulation (Fig. 2). Details on the tick collections and infection prevalence per session per island are available in Additional file 1: Table S2. All collected ticks were determined to be I. ricinus using a light microscope and morphological identification keys [39,40,41]. We estimated the average DON per 100 m2 per site per session as the number of nymphs collected/dragging distance × 100.
Detection of B. afzelii in ticks and rodents
DNA from nymphs (n = 1709) was extracted using the ammonium hydroxide (NH4OH) (Sigma Aldrich, St. Louis, MO, USA) method [42] with slight modifications. Each nymph was taken from the 70% ethanol in which it was stored, air dried, and placed in a 2-ml safe lock microtube (Sarstedt, Numbrecht, Germany) containing 200 µl of 1.25% NH4OH solution. Each sample was heated for 3 min at 100 °C, crushed using the Qiagen TissueLyser (Qiagen, Hilden, Germany) for 2 min and then incubated at 100 °C for 20 min. After quick cooling, the microtubes were left open and incubated at 100 °C for 15 min to evaporate the ammonia (approx. 50% of the starting volume). Negative controls (containing only 1.25% NH4OH solution) were included after every five samples. Total DNA was extracted from rodent ear biopsy samples using the protocol of Laird et al. [43] with negative controls included after every five samples. All extracted DNA was stored at − 20 °C until further analysis.
The DNA samples from the ticks and rodents were screened for B. burgdorferi s.l. using real-time PCR (Bio-Rad CFX96 Touch Deep Well Real-Time PCR Detection System; Bio-Rad Laboratories, Hercules, CA, USA). Ticks were screened for B. burgdorferi s.l. by targeting 23S ribosomal RNA (rRNA) [44] in most cases, but the flagellin gene [45] was targeted for ticks captured from specific islands in May and June 2019 (Additional file 1: Table S2) as these samples were also included in another study (in preparation). All tick and ear biopsy samples positive for B. burgdorferi s.l. were screened using B. afzelii-specific primers [46]. The list of primers and probes used is available in Additional file 1: Table S5, while detailed protocols for the PCR assays are provided in Additional file 2: Text S2.
Based on the tick data we calculated the percentage NIP as (the number of quantitative PCR-positive nymphs/total number of screened nymphs) × 100, and the density of infected nymph (DIN per 100 m2) as DON × NIP per site per session. For rodents, we calculated the abundance index of infected rodents as (the number of infected animals captured per island divided by trapping effort) × 100 (Additional file 1: Table S1).
Statistical analyses
First, we evaluated whether rodent removal was successful at controlling rodent abundance. We fitted generalized linear mixed models (GLMMs) with a Poisson distribution using the glmer function in the R package lme4 [47] to examine whether rodent abundance depended on sessions and treatment. The model included the number of rodents captured as the response variable; the trapping effort as an offset; and session, treatment and their interaction as explanatory variables. Island identity was included in the model as a random effect to control for the potential correlation among observations from the same island. An equivalent model was used to examine the role of treatment and session on the abundance of infected rodents in the experimental study. We further studied the response of rodent abundance and the abundance of infected rodents to session and the size category (small vs large) of the island in an observational study. The level of tick infestation of rodents in 2019 was examined in relation to the treatment of the island (removal, control, large), trapping session (May, June, July and August/September) and rodent species (bank voles vs field voles) with individual level rodent trapping data. A GLMM approach with negative binomial distribution and trapping island as the random effect was performed using the glmmTMB package in R [48]. An overview of all the tested models is presented in Additional file 1: Table S6.
On the experimental islands, we investigated whether tick-related measures, i.e. NIPt+1, DONt+1 and DINt+1, in May 2020 were associated with the rodent removal treatment (removal vs control) or rodent abundance index or abundance index of infected rodents in 2019. We used generalized linear models (GLMs; NIPt+1) with binomial error distributions or linear models (LMs; DONt+1 and DINt+1). With respect to the NIPt+1 and DINt+1 models, we used either rodent removal versus control treatment; the rodent abundance index in 2019 (rodent abundance index of each session tested separately); or the abundance index of infected rodents (each session tested separately). The DONt+1 model included the rodent removal treatment versus control treatment, or the rodent abundance index in 2019 (the rodent abundance index with each session tested separately). We tested several NIPt+1, DONt+1 and DINt+1 models using the rodent abundance index in each trapping session separately, as the rodent abundance index estimates at different sessions were correlated and, thus, cannot be included in the same model. To take into account multiple testing for each response variable (rodent abundance index at four sessions using the same data), the threshold significance level was determined based on Bonferroni correction (0.05/4 = 0.0125), i.e. P-value ≤ 0.0125 was considered to indicate a significant result. We also examined DON, NIP and DIN in 2019 to ensure that the treatment (removal vs control) did not differ among the islands before treatment (Additional file 2: Text S3; Additional file 1: Table S6).
On observational islands, we used the same approach as with the experimental islands. We used GLMs and the LM model for testing NIPt+1 and DINt+1, respectively, by including the size category (small vs large) of the islands, either the rodent abundance index in 2019 (tested each session separately) or the abundance index of infected rodents in 2019 (each session tested separately) as fixed factors. DONt+1 models were analysed using LM including the rodent abundance index in 2019 (tested each session separately) as a fixed factor (Additional file 1: Table S6). To take into account the multiple testing for each response variable, the Bonferroni corrected P-value was used in the interpretation of the results (threshold P-value = 0.0125).
All data analyses and model building were performed in R (4.2.2) [49].
Results
Rodents and tick infestation on the experimental and observational islands in 2019
Rodent abundance did not differ between the control and removal islands at the beginning of the study (May 2019) (Fig. 3; Additional file 1: Table S7). Rodent abundance increased on the control islands during the breeding season, being significantly higher in the August/September trapping session than in the May trapping session (Fig. 3, Additional file 1: Table S7). Rodent abundance was lower on the removal islands than on the control islands, but the difference was statistically significant only in July 2019 (P = 0.04; Additional file 1: Table S7). Additionally, the abundance of infected rodents did not differ between treatments (Additional file 1: Table S7).

Mean (± standard deviation) rodent abundance index (a) and abundance index of infected rodents (b) on the removal islands, control islands and large islands during the trapping sessions in 2019
Neither rodent abundance nor the abundance of infected rodents differed between the small (control) and large islands at the beginning of the study (May 2019) (Fig. 3; Additional file 1: Table S8). However, the abundances were higher on the large islands than on the small control islands in the other trapping sessions (Fig. 3, P < 0.001; Additional file 1: Table S8).
The mean tick infestations on the rodents were 7.6 larvae and 0.56 nymphs per rodent. Of the 281 rodents examined, 246 were infested by ticks (87.5%) and 74 of these (30%) were infested with both larvae and nymphs. Larval tick infestation load on the rodents did not differ between the treatments or the trapping session, but was significantly higher in bank voles than in field voles (Additional file 1: Table S9).
Ticks and B. afzelii in ticks in 2020
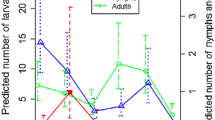
In 2020, a total of 550 nymphal ticks were collected. The estimated mean (± standard deviation [SD]) density of nymphs (DONt+1) per 100 m2 was 6.1 ± 1.79 on the rodent removal islands, 8.09 ± 2.77 on the small control islands and 22.9 ± 7.19 on the large islands (Fig. 4).

Mean (+/- standard deviation) (a) density of nymphs (DON), (b) nymph infection prevelance (NIP) and (c) density of infected nymphs (DIN) in May 2020 in study islands (removal, control and large). (a) mean DONt+1 per 100 m2, (b) NIPt+1 per 100 m2 (c) and DINt+1 per 100 m2 in the subsequent year (2020) in removal, control and large islands
In total, 414 of the collected nymphs from May 2020 were tested for B. afzelii. In May 2020, the infection prevalence in nymphs (NIPt+1) was 15.4% (95% confidence interval [CI]: 3–34%) on the rodent removal islands, 21.2% (95% CI: 11–32%) on the small control islands and 12.7% (95% CI: 4–21%) on the large islands (Fig. 4).
The estimated mean (± SD) density of infected nymphs (DINt+1 = NIPt+1*DONt+1) per 100 m2 in May 2020 was 1.36 ± 0.75 on the rodent removal islands, 1.70 ± 0.63 on the small control islands and 2.95 ± 1.55 on the large islands (Fig. 4).
In 2019, DON, NIP and DIN did not differ between the rodent removal and control islands in any of the sessions, i.e. in May, June, July, and August/September (Additional file 1: Table S10).
Experimental study: effects of rodents on DONt+1, NIPt+1 or DINt+1
On the experimental study islands (control and removal islands), there was no effect of rodent removal treatment in the first study year (2019) on DONt+1, NIPt+1 and DINt+1 (May 2020) (P > 0.5; Additional file 1: Table S11). Within the experimental islands, neither DONt+1 (P > 0.5; Additional file 1: Table S12) nor NIPt+1 (P > 0.5; Additional file 1: Table S13) nor DINt+1 (P > 0.5; Additional file 1: Table S14) was associated with the rodent abundance index in any of the trapping sessions in 2019. The abundance index of infected rodents in any of the trapping sessions in 2019 were associated with NIPt+1 on the experimental study islands (Additional file 1: Table S13).
Observational study: effects of rodents and island size on, DONt+1, NIPt+1 or DINt+1
On the observational islands (small control and large islands), DONt+1 showed a positive association with the rodent abundance index in September (Fig. 5; Additional file 1: Table S12), whereas no association with the rodent abundance index was observed in the other study sessions (Additional file 1: Table S12).

Predicted (line with 95% confidence interval in shading) and observed (points, with different colours indicating the small and large islands, respectively) density of nymphs (DONt+1) per 100 m2 in May 2020 in relation to the rodent abundance index in August/September 2019 in the observational study. The figure show the relationship between the rodent abundance index in year t (2019) and DONt+1 in year t+1 (2020)
Moreover, NIPt+1 was negatively associated with the rodent abundance index and the abundance index of infected rodents in July 2019 (P = 0.011; Fig. 6; Additional file 1: Table S15), but no significant associations were observed during the other trapping sessions. However, DINt+1 was not associated with the rodent abundance index in any trapping session in 2019 (P > 0.5; Additional file 1: Table S16).

Predicted (line with 95% confidence interval) and observed (points, with different colours indicating the small and large islands, respectively) nymph infection prevalence (NIPt+1) in May 2020 in relation to the rodent abundance index in July 2019 in the observational study. The figure show the relationship between the rodent abundance index in year t (2019) and DONt+1 in year t+1 (2020)
Neither DONt+1 nor DINt+1 was associated with the island type (Table 1). NIPt+1 was associated with island type, with a lower NIPt+1 value on large islands (P = 0.046; Table 1).
Discussion
In this study, we examined the association between the abundance of the reservoir rodent host and the abundance of nymphal I. ricinus ticks and a rodent-associated tick-borne pathogen, B. afzelii, in ticks. We observed that rodent removal in the experimental study did not translate into changes in the abundance of ticks, tick infection prevalence or the abundance of infected ticks in the following year. Neither NIPt+1 nor DINt+1 was associated with the abundance index of rodents or infected rodents on the small experimental islands. In the observational component of this study, we aimed to test whether NIPt+1, DONt+1, or DINt+1 was associated with rodent density and island type. Contrary to our hypothesis, we found a negative association between NIPt+1 and the abundance index of rodents and infected rodents. However, we found a positive association between rodent abundance index and DONt+1.
NIPt+1 in relation to preceding rodent abundances
Our experimental study revealed no association between rodent abundance at t (2019) and NIPt+1 (2020). This finding contrasts with the findings of an earlier experimental study by Krawczyk et al. [29] which showed that high rodent abundance was associated with higher infection prevalence of rodent-associated tick-borne pathogens (TBPs) in nymphs. A possible explanation for these differing results is the difference in rodent abundance between the two study systems. Krawczyk and coworkers operated in small (0.25 ha) enclosures, where the highest rodent density was > 40 individuals in an enclosure per year in the removal treatment [29]. Our study was carried out on natural islands that varied in size and had overall lower rodent abundance, with the highest number of captured rodents being up to 15 individuals on a 2-ha removal island. In addition, our tick density was relatively low (on average, < 10 nymphs per 100 m2 on removal islands). The low rodent density together with low tick density might decrease the likelihood of larval and nymphal ticks feeding on the same reservoir rodent host individuals, or rodents in general, potentially leading to a lack of association between rodent abundance and NIPt+1. However, approximately one third of the rodent individuals were infested with both larvae and nymph(s), which does not provide support to the assumption that low host density and low tick densities decrease aggregation on the host individuals. Nevertheless, our results suggest that even a very low rodent density is sufficient to support the circulation of B. afzelii. Alternatively, B. afzelii might have been supported by other reservoir species, such as shrews or squirrels [15, 50,51,52,53,54], although the abundance of these species was not estimated reliably in this study. Irrespective of the mechanism supporting pathogen persistence, our results suggest that the removal of rodents even on small islands may not limit the transmission cycle of B. afzelii as: (i) we were not able to detect any effect on NIPt+1 and (ii) B. afzelii was detected on all islands with ticks in May 2020 and rodents 2019. Moreover, although the removal of rodents is unlikely to be a realistic management action for controlling the health hazard caused by B. afzelii, it may cause deleterious effects on ecosystems though trophic cascades, such as those affecting predators that prey on rodents.
Interestingly, on the observational study islands, we found a negative association between rodent abundance in the summer and NIP in the following spring. This result contrasts with those from previous studies that showed a positive association between high rodent abundance and nymphal infection prevalence [25,26,27,28,29]. There are several possible reasons for our result. First, it may be that high rodent density could lead to a decrease in tick infection burden on rodents [55, 56], thereby possibly resulting in a decrease infection prevalence in rodents and, ultimately, in decreased NIPt+1 in the following year. However, this would be likely only in very low tick densities and very high rodent densities, which were not observed in our study. Although the mean tick infestation load on rodents was considerably lower in our study system than that reported in another northern ecosystem in Norway [57], the rodent density was also low and thus unlikely to decrease tick aggregation on rodents. A second possible explanation is that high rodent abundance in the summer/early autumn likely results from an influx of young, uninfected and/or maternal antibody-protected [58] rodents in the population. Consequently, the high proportion of uninfected rodents potentially limits the transmission of pathogens from rodents to larval ticks. However, the proportion of infected rodents was high throughout our study period. A third potential explanation is that the success of host-tick transmission by B. afzelii is limited, as the bank vole, which is the most common rodent in the study system, becomes immune against tick infestation [59, 60].
Lastly, the type of the island in our study was shown to have a significant association with NIPt+1, with large islands having lower NIPt+1. A previous study conducted in the USA showed that NIP is negatively associated with increasing forest fragment size, indicating the importance of a diverse host community, which can play an important role in tick control and transmission of tick-borne pathogens [61]. As simultaneously large islands had higher rodent abundance, it might be that the negative association between NIPt+1 and rodents is caused by some other factor associated with the size of the island. For example, DON was higher on large islands, suggesting that the abundance of hosts that support tick breeding, such as deer, is higher and/or environmental conditions are more suitable for an abundant tick population. Hence, on the larger islands with high tick abundance, a higher proportion of larvae might feed on deer that do not support pathogen transmission, resulting in a decrease in pathogens according to the dilution effect [62,63,64,65]. However, we monitored the abundance of dung piles of cervids and found no association either with island type or with DON. Nevertheless, our study highlights the complexity of this disease system and emphasizes the need for further studies to quantify the interplay between rodents, ticks and pathogens, especially in northern Europe.
Associations of rodent abundance with DONt+1 and DINt+1
A positive association between the rodent abundance index and DONt+1 was found only in the observational study. Our results are in accordance with those of a previous study that showed a positive association between high rodent abundance and the density of nymphs [27, 35, 66]. This positive relationship might be due to the role that rodents play in the tick life-cycle, acting as a main host for larval feeding and providing good engorgement success for larvae [67], thereby driving nymphal abundance in the following year. An alternative explanation for this positive association is that although cervids were observed on all of our study islands, they may be more persistent on large islands. This persistence could contribute to higher tick density on larger islands, despite the lack of association between DONt+1 or the size category of the island, and the abundance of cervid dung piles.
The DIN is a key measure of entomological risk associated with I. ricinus because nymphs are the major vectors of human infection. Nymphs are more abundant in nature and more difficult to detect when attached to the host body than adults, while simultaneously carrying several TBPs at relatively high prevalence [11, 13]. Therefore, DIN provides an indication of the risk of human infection with TBPs transmitted by I. ricinus nymphs. We expect that the factors influencing DON and NIP can also influence DINt+1, which is a product of DON and NIP [27]. In our observational study on the islands, we found no association between DIN and rodent abundance. This could be because the rodent abundance index has a positive effect on DON but a negative effect on NIP; together, these two opposing effects resulted in a non-significant difference in DIN. Thus, our findings highlight the need for further studies on the fluctuations in the abundance of other vertebrate hosts affecting the density of nymphs and infected nymphs.
Conclusions
The results of our field experiment study demonstrated that the expected positive associations between rodents and rodent-associated TBPs are not always found. We demonstrated that a reduction in rodent population density may not lead to a corresponding decrease in tick infection prevalence, suggesting that only a few rodents or alternative rodent reservoirs may maintain B. afzelii in nature. Consequently, artificial rodent removal may not be sufficient for reducing zoonotic hazards in natural systems. However, our experimental rodent removal and observational rodent trappings were carried out for only 1 year, whereas longer-term studies on islands with larger variations in tick and rodent abundances could provide a better understanding of their relationships in a natural ecosystem. In addition, detailed information on the biodiversity of vertebrate hosts for ticks and pathogens in the ecosystem is required to determine the role of cyclic rodent populations in driving tick and tick-borne pathogens.
Availability of data and materials
All population-level data are provided in Additional file 1.
Abbreviations
- DIN:
-
Density of infected nymphs
- DON:
-
Density of nymphs
- NIP:
-
Nymphal infection prevalence
References
Parola P, Raoult D. Ticks and tickborne bacterial diseases in humans: an emerging infectious threat. Clin Infect Dis. 2001;32:897–928.
Rochlin I, Toledo A. Emerging tick-borne pathogens of public health importance: a mini-review. J Med Microbiol. 2020;69:781.
Sprong H, Azagi T, Hoornstra D, Nijhof AM, Knorr S, Baarsma ME, et al. Control of Lyme borreliosis and other Ixodes ricinus-borne diseases. Parasit Vectors. 2018;11:1–16.
Rosenberg R, Lindsey NP, Fischer M, Gregory CJ, Hinckley AF, Mead PS, et al. Vital signs: trends in reported vectorborne disease cases—United States and Territories, 2004–2016. Morb Mortal Wkly Rep. 2018;67:496.
Sykes RA, Makiello P. An estimate of Lyme borreliosis incidence in Western Europe. J Public Health. 2017;39:74–81.
Gern L, Humair P. Ecology of Borrelia burgdorferi sensu lato. In: Gray J, Kahl O, Lane RS, Stanek G, editors. Lyme borreliosis. Biology, epidemiology and control. CABI International: Wallingford; 2002. p. 149–74.
Strnad M, Hönig V, Růžek D, Grubhoffer L, Rego ROM. Europe-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus ticks. Appl Environ Microbiol. 2017;83:e00609-e617.
Matuschka F-R, Fischer P, Musgrave K, Richter D, Spielman A. Hosts on which nymphal Ixodes ricinus most abundantly feed. Am J Trop Med Hyg. 1991;44:100–7.
Mysterud A, Hügli C, Viljugrein H. Tick infestation on medium–large-sized mammalian hosts: are all equally suitable to Ixodes ricinus adults? Parasit Vectors. 2021;14:1–6.
Talleklint L, Jaenson TGT. Infestation of mammals by Ixodes ricinus ticks (Acari: Ixodidae) in south-central Sweden. Exp Appl Acarol. 1997;21:755–71.
Fish D. Environmental risk and prevention of Lyme disease. Am J Med. 1995;98:2S-9S.
Barbour AG, Fish D. The biological and social phenomenon of Lyme disease. Science. 1993;260:1610–6.
Falco RC, McKenna DF, Daniels TJ, Nadelman RB, Nowakowski J, Fish D, et al. Temporal relation between Ixodes scapularis abundance and risk for Lyme disease associated with erythema migrans. Am J Epidemiol. 1999;149:771–6.
Helble JD, McCarthy JE, Hu LT. Interactions between Borrelia burgdorferi and its hosts across the enzootic cycle. Parasite Immunol. 2021;43:e12816.
Humair P-F, Gern L. Relationship between Borrelia burgdorferi sensu lato species, red squirrels (Sciurus vulgaris) and Ixodes ricinus in enzootic areas in Switzerland. Acta Trop. 1998;69:213–27.
Radolf JD, Caimano MJ, Stevenson B, Hu LT. Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes. Nat Rev Microbiol. 2012;10:87–99. https://doi.org/10.1038/nrmicro2714.
Hanincova K, Schäfer SM, Etti S, Sewell H-S, Taragelova V, Ziak D, et al. Association of Borrelia afzelii with rodents in Europe. Parasitology. 2003;126:11–20.
Hu CM, Humair P-F, Wallich R, Gern L. Apodemus sp. rodents, reservoir hosts for Borrelia afzelii in an endemic area in Switzerland. Zentralbl Bakteriol. 1997;285:558–64.
Humair P-F, Peter O, Wallich R, Gern L. Strain variation of Lyme disease spirochetes isolated from Ixodes ricinus ticks and rodents collected in two endemic areas in Switzerland. J Med Entomol. 1995;32:433–8.
Gern L, Estrada-Pena A, Frandsen F, Gray J, Jaenson T, Jongejan F, et al. European reservoir hosts of Borrelia burgdorferi sensu lato. Zentralbl Bakteriol. 1998;287:196–204.
Kybicova K, Kurzova Z, Hulinska D. Molecular and serological evidence of Borrelia burgdorferi sensu lato in wild rodents in the Czech Republic. Vector Borne Zoonotic Dis. 2008;8:645–52.
Król N, Obiegala A, Imholt C, Arz C, Schmidt E, Jeske K, et al. Diversity of Borrelia burgdorferi sensu lato in ticks and small mammals from different habitats. Parasit Vectors. 2022;15:1–11.
Kalmár Z, Sándor AD, Matei IA, Ionică A, D’Amico G, Gherman CM, et al. Borrelia spp. in small mammals in Romania. Parasit Vectors. 2019;12:1–6.
Mysterud A, Stigum VM, Jaarsma RI, Sprong H. Genospecies of Borrelia burgdorferi sensu lato detected in 16 mammal species and questing ticks from northern Europe. Sci Rep. 2019;9:5088.
Jones CG, Ostfeld RS, Richard MP, Schauber EM, Wolff JO. Chain reactions linking acorns to gypsy moth outbreaks and Lyme disease risk. Science. 1998;279:1023–6.
LoGiudice K, Ostfeld RS, Schmidt KA, Keesing F. The ecology of infectious disease: effects of host diversity and community composition on Lyme disease risk. Proc Natl Acad Sci USA. 2003;100:567–71.
Ostfeld RS, Canham CD, Oggenfuss K, Winchcombe RJ, Keesing F. Climate, deer, rodents, and acorns as determinants of variation in Lyme-disease risk. PLoS Biol. 2006;4:e145.
Duijvendijk GV, Sprong H, Takken W. Multi-trophic interactions driving the transmission cycle of Borrelia afzelii between Ixodes ricinus and rodents: a review. Parasit Vectors. 2015;8:1–11.
Krawczyk AI, van Duijvendijk GLA, Swart A, Heylen D, Jaarsma RI, Jacobs FHH, et al. Effect of rodent density on tick and tick-borne pathogen populations: consequences for infectious disease risk. Parasit Vectors. 2020;13:1–17.
Henttonen H, Oksanen T, Jortikka A, Haukisalmi V. How much do weasels shape microtine cycles in the northern Fennoscandian taiga? Oikos. 1987;50:353–65.
Hansson L, Henttonen H. Rodent dynamics as community processes. Trends Ecol Evol. 1988;3:195-200. https://doi.org/10.1016/0169-5347(88)90006-7.
Aminikhah M, Forsman JT, Koskela E, Mappes T, Sane J, Ollgren J, et al. Rodent host population dynamics drive zoonotic Lyme Borreliosis and orthohantavirus infections in humans in Northern Europe. Sci Rep. 2021;11:16128.
Tkadlec E, Václavík T, Široký P. Rodent host abundance and climate variability as predictors of tickborne disease risk 1 year in advance. Emerg Infect Dis. 2019;25:1738.
Schauber EM, Ostfeld RS, Andrew SE. What is the best predictor of annual Lyme disease incidence: weather, mice, or acorns? Ecol Appl. 2005;15:575–86.
Cayol C, Koskela E, Mappes T, Siukkola A, Kallio ER. Temporal dynamics of the tick Ixodes ricinus in northern Europe: epidemiological implications. Parasit Vectors. 2017;10:1–11.
Laaksonen M, Sajanti E, Sormunen JJ, Penttinen R, Hänninen J, Ruohomäki K, et al. Crowdsourcing-based nationwide tick collection reveals the distribution of Ixodes ricinus and I. persulcatus and associated pathogens in Finland. Emerg Microbes Infect. 2017;6:1–7.
Sormunen JJ, Klemola T, Vesterinen EJ, Vuorinen I, Hytönen J, Hänninen J, et al. Assessing the abundance, seasonal questing activity, and Borrelia and tick-borne encephalitis virus (TBEV) prevalence of Ixodes ricinus ticks in a Lyme borreliosis endemic area in Southwest Finland. Ticks Tick-Borne Dis. 2016;7:208–15.
Sormunen JJ, Andersson T, Aspi J, Bäck J, Cederberg T, Haavisto N, et al. Monitoring of ticks and tick-borne pathogens through a nationwide research station network in Finland. Ticks Tick-Borne Dis. 2020;11:101449.
Arthur DR. British ticks. Butterworth & Co.: London; 1963.
Filippova NA. Arachnida class: ixodid ticks of the subfamily Ixodinae. In: Paukoobraznye Fauna SSSR, vol. 4. Leningrad: Nauka; 1977.
Snow KR. Identification of larval ticks found on small mammals in Britain. Berkshire: Mammal Society; 1978.
Schouls LM, Pol IVD, Rijpkema SGT, Schot CS. Detection and identification of Ehrlichia, Borrelia burgdorferi sensu lato, and Bartonella species in Dutch Ixodes ricinus ticks. J Clin Microbiol. 1999;37:2215–22.
Laird PW, Zijderveld A, Linders K, Rudnicki MA, Jaenisch R, Berns A. Simplified mammalian DNA isolation procedure. Nucleic Acids Res. 1991;19:4293. https://doi.org/10.1093/nar/19.15.4293.
Courtney JW, Kostelnik LM, Zeidner NS, Massung RF. Multiplex real-time PCR for detection of anaplasma phagocytophilum and Borrelia burgdorferi. J Clin Microbiol. 2004;42:3164–8. https://doi.org/10.1128/JCM.42.7.3164-3168.2004.
Schwaiger M, Peter O, Cassinotti P. Routine diagnosis of Borrelia burgdorferi (sensu lato) infections using a real-time PCR assay. Clin Microbiol Infect. 2001;7:461–9.
Michelet L, Delannoy S, Devillers E, Umhang G, Aspan A, Juremalm M, et al. High-throughput screening of tick-borne pathogens in Europe. Front Cell Infect Microbiol. 2014;4:103.
Bates D, Maechler M, Bolker B, Walker S. Mixed-effects models using lme4. J Stat Softw. 2015;67:1–48.
Brooks ME, Kristensen K, Van Benthem KJ, Magnusson A, Berg CW, Nielsen A, et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017;9:378–400.
R Core Team. A language and environment for statistical computing. Vienna: Foundation for Statistical Computing; 2022.
De Pelsmaeker N, Korslund L, Steifetten Ø. Host in reserve: The role of common shrews (Sorex araneus) as a supplementary source of tick hosts in small mammal communities influenced by rodent population cycles. Ecol Evol. 2022;12:e8776.
Mysterud A, Byrkjeland R, Qviller L, Viljugrein H. The generalist tick Ixodes ricinus and the specialist tick Ixodes trianguliceps on shrews and rodents in a northern forest ecosystem–a role of body size even among small hosts. Parasit Vectors. 2015;8:1–10.
Pisanu B, Chapuis J-L, Dozières A, Basset F, Poux V, Vourc’h G. High prevalence of Borrelia burgdorferi sl in the European red squirrel Sciurus vulgaris in France. Ticks Tick-Borne Dis. 2014;5:1–6.
Sormunen JJ, Mäkelä S, Klemola T, Alale TY, Vesterinen EJ. Voles, shrews and red squirrels as sources of tick blood meals and tick-borne pathogens on an island in southwestern Finland. Ticks Tick-Borne Dis. 2023;14:102134.
Talleklint L, Jaenson TG. Transmission of Borrelia burgdorferi sl from mammal reservoirs to the primary vector of Lyme borreliosis, Ixodes ricinus (Acari: Ixodidae), Sweden. J Med Entomol. 1994;31:880–6.
Kiffner C, Vor T, Hagedorn P, Niedrig M, Rühe F. Factors affecting patterns of tick parasitism on forest rodents in tick-borne encephalitis risk areas, Germany. Parasitol Res. 2011;108:323–35.
Krasnov BR, Stanko M, Morand S. Host community structure and infestation by ixodid ticks: repeatability, dilution effect and ecological specialization. Oecologia. 2007;154:185–94.
Lindsø LK, Anders JL, Viljugrein H, Herland A, Stigum VM, Easterday WR, et al. Individual heterogeneity in ixodid tick infestation and prevalence of Borrelia burgdorferi sensu lato in a northern community of small mammalian hosts. Oecologia. 2023;34:1–13.
Gomez-Chamorro A, Heinrich V, Sarr A, Roethlisberger O, Genné D, Bregnard C, et al. Maternal antibodies provide bank voles with strain-specific protection against infection by the lyme disease pathogen. Appl Environ Microbiol. 2019;85:e01887-19. https://doi.org/10.1128/AEM.01887-19.
Dizij A, Kurtenbach K. Clethrionomys glareolus, but not Apodemus flavicollis, acquires resistance to Ixodes ricinus L, the main European vector of Borrelia burgdorferi. Parasite Immunol. 1995;17:177–83.
Gomez-Chamorro A, Li Y, Herrera A, Rais O, Dautel H, Voordouw MJ. Borrelia afzelii does not suppress the development of anti-tick immunity in bank voles. bioRxiv. 2020;8:504.
Allan BF, Keesing F, Ostfeld RS. Effect of forest fragmentation on Lyme disease risk. Conserv Biol. 2003;17:267–72.
Gandy S, Kilbride E, Biek R, Millins C, Gilbert L. Experimental evidence for opposing effects of high deer density on tick-borne pathogen prevalence and hazard. Parasit Vectors. 2021;14:1–12.
Gandy S, Kilbride E, Biek R, Millins C, Gilbert L. No net effect of host density on tick-borne disease hazard due to opposing roles of vector amplification and pathogen dilution. Ecol Evol. 2022;12:e9253.
Rosef O, Paulauskas A, Radzijevskaja J. Prevalence of Borrelia burgdorferi sensu lato and Anaplasma phagocytophilum in questing Ixodes ricinus ticks in relation to the density of wild cervids. Acta Vet Scand. 2009;51:1–8.
Schmidt KA, Ostfeld RS. Biodiversity and the dilution effect in disease ecology. Ecology. 2001;82:609–19.
Ostfeld RS, Schauber EM, Canham CD, Keesing F, Jones CG, Wolff JO. Effects of acorn production and mouse abundance on abundance and Borrelia burgdorferi infection prevalence of nymphal Ixodes scapularis ticks. Vector-Borne Zoonotic Dis. 2001;1:55–63.
Hofmeester TR, Coipan EC, Wieren SEV, Prins HHT, Takken W, Sprong H. Few vertebrate species dominate the Borrelia burgdorferi sl life cycle. Environ Res Lett. 2016;11:43001.
Acknowledgements
We are thankful to Johan Sundbäck, Mika Sundbäck and Camilla Sundbäck for the help in the field and recruitment on the islands, and to the landowners for the permission to carry out the study. We would like to thank Saija Vuorenmaa and Kata Tiainen for their efforts in the field. We thank also all “vole team” members, who gave constructive feedback for earlier versions of the manuscript. This research work was funded by the Academy of Finland (Grant numbers 329308, 329332, 335651, 329326 and 354988 to ERK) and the University of Jyväskylä Graduate School (NK). We extend our gratitude to the reviewers for their time and effort in reviewing the manuscript: we sincerely appreciate all your valuable comments and suggestions, which helped us improve the quality of the manuscript.
Funding
Open Access funding provided by University of Jyväskylä (JYU).
Author information
Authors and Affiliations
Contributions
ERK and TM, designed the methodology. ERK, EW and IB, collected the data. NK and SS analysed the samples. NK, YXGW and ERK performed the statistical analyses. NK led the writing of the manuscript. All authors contributed to the drafts and gave their feedback and approval for the final publication.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All rodent handling procedures were done following the protocols approved by the Finnish Animal Experiment Board the authorization ESAVI/3981/2018. The handling complied with the Finnish Act on the Use of Animals for Experimental Purposes (62/2006). All trappings were done with landowners' permission. Tick collections do not require ethical permissions.
Consent for publication
Not applicable.
Competing interests
No competing interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
The population-level data of rodent abundance index and abundance index of infected rodents, including information about trapping sessions and the number of traps. Table S2. Population-level data on ticks including information about the number of collected nymphs and infected nymphs during 2019 and 2020. Table S3. The estimated effects of the treatments (removal, control and large islands) on the mean density of cervid dung in 2019. Table S4. The estimated effects of the treatments (removal, control and large islands) and the average density of cervid dung in 2019 on the density of nymphs on the island (DONt+1) in 2020. Table S5. Detailed information on primer and probes used for screening of B. afzelii. Table S6. List of all tested models. Table S7. The estimated effects of rodent removal treatment and session (May, June, July and August/September) and their interactions on the rodent abundance and the abundance of infected rodents on the experimental study. Table S8. The estimated effects of island size category (small control vs large) and session (May, June, July and August/September) and their interactions on the rodent abundance and the abundance of infected rodents in the observational study. Table S9. The estimated effects of the trapping sessions (May, June, July, August/Septe,ber), treatment (removal, control and large islands), and the rodent species (bank vole vs field vole) the larval tick infestation load in 2019.Table S10. The estimated effects of the study session (May, June, July, August/September) and rodent removal treatment on the density of nymphs (DON), the density of infected nymphs (DIN) and NIP in 2019 on experimental islands. Table S11. The estimated effects of rodent removal treatment on DONt+1, NIPt+1 and DINt+1 (May 2020) on the experimental islands. Table S12. The estimated effects of the rodent abundance index in different trapping sessions (May, June, July and August/September) on DONt+1 (May 2020) (a) on the experimental and (b) on the observational study islands. Table S13. The estimated effects of the rodent abundance index and the abundance index of infected rodents in different trapping sessions (May, June, July and August/September) on NIPt+1 (2020) on the experimental islands. Table S14. The estimated effects of the rodent abundance index and the abundance index of infected rodents in different trapping sessions (May, June, July and August/September) on the density of infected nymphs (DINt+1) in 2020 on the experimental islands. Table S15. The estimated effects of the rodent abundance index and the abundance index of infected rodents in different trapping sessions (May, June, July and August/September) on NIPt+1 (2020) on the observational islands. Table S16. The estimated effects of the rodent abundance index and the abundance index of infected rodents in different trapping sessions (May, June, July and August/September) on the density of infected nymphs (DINt+1) in 2020 on the observational islands.
Additional file 2: Text S1.
Detailed information on the dung survey and the model used to examine how cervids differ in island treatments (removal, control, and large) and how they affect the density of nymphs (DONt+1 Text S2. Additional laboratory protocol for PCR. Text S3. D Model selection for the effect of treatment on the density of nymphs (DON), the density of infected nymphs (DIN) and nymphs infection prevalence (NIP) in 2019 on experimental islands.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kiran, N., Brila, I., Mappes, T. et al. Effects of rodent abundance on ticks and Borrelia: results from an experimental and observational study in an island system. Parasites Vectors 17, 157 (2024). https://doi.org/10.1186/s13071-024-06130-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06130-x