
Abstract
Background
Aedes japonicus japonicus (Theobald, 1901) and Aedes koreicus (Edwards, 1917) have rapidly spread in Europe over the last decades. Both species are very closely related and occur in sympatry. Females and males are difficult to distinguish. However, the accurate species discrimination is important as both species may differ in their vectorial capacity and spreading behaviour. In this study, we assessed the potential of geometric wing morphometrics as alternative to distinguish the two species.
Methods
A total of 147 Ae. j. japonicus specimens (77 females and 70 males) and 124 Ae. koreicus specimens (67 females and 57 males) were collected in southwest Germany. The left wing of each specimen was removed, mounted and photographed. The coordinates of 18 landmarks on the vein crosses were digitalised by a single observer. The resulting two-dimensional dataset was used to analyse the differences in the wing size (i.e. centroid size) and wing shape between Ae. j. japonicus and Ae. koreicus using geometric morphometrics. To analyse the reproducibility of the analysis, the landmark collection was repeated for 20 specimens per sex and species by two additional observers.
Results
The wing size in female Ae. koreicus was significantly greater than in Ae. j. japonicus but did not differ significantly for males. However, the strong overlap in wing size also for the females would not allow to discriminate the two species. In contrast, the wing shape clustering was species specific and a leave-one-out validation resulted in a reclassification accuracy of 96.5% for the females and 91.3% for the males. The data collected by different observers resulted in a similar accuracy, indicating a low observer bias for the landmark collection.
Conclusions
Geometric wing morphometrics provide a reliable and robust tool to distinguish female and male specimens of Ae. j. japonicus and Ae. koreicus.
Graphical Abstract

Similar content being viewed by others

Background
The global spread of mosquito species poses a serious risk for public health, including nuisance and transmission of pathogens [1,2,3]. The most prominent representative in Europe is the Asian tiger mosquito (Aedes albopictus Skuse, 1984), which is an aggressive biter and potential vector of several pathogens. Establishment of the species in the Mediterranean region allowed the local circulation of chikungunya virus (CHIKV) and small outbreaks of dengue virus (DENV) and Zika virus (ZIKV) over the last 15 years [4,5,6,7,8,9,10,11,12,13,14].
Besides the Asian tiger mosquito, there are two further established exotic Aedes species in Europe: the Japanese bush mosquito (Aedes japonicus japonicus) and Korean bush mosquito (Aedes koreicus). The first established population of Ae. j. japonicus was detected in Belgium in 2002 [15]. In 2008, it was confirmed for Switzerland and bordering Germany [16], where it was later confirmed to be widespread [17]. Within 2 decades, the species is now present in wide parts of Germany [18]. The species is also considered to be established in Austria [19], Slovenia [20], The Netherlands [21], Hungary [22], France [23], Croatia [24], Bosnia-Herzegovina [25], Serbia [25], Italy [19], Liechtenstein [22] and Spain [26]. Aedes koreicus was first detected outside its native range in Belgium in 2008, where it established and overwintered, but did not seem to spread further [27, 28]. In contrast, the population detected in northeastern Italy in 2011 was observed to rapidly expand its distribution in Italy [29, 30] and towards Switzerland [31]. Established small populations have also been observed in Germany [32, 33], Hungary [34] and the north coast of the Black Sea in Ukraine and Russia [35].
Analysis of the host-feeding patterns demonstrated that Ae. j. japonicus and Ae. koreicus show a high prevalence of mammalian blood meals, including humans, while birds only play a minor role [36, 37]. The vector competence of both species is not well studied. For Ae. j. japonicus, experimental studies confirmed vector competence for several arboviruses, for example, Japanese encephalitis virus (JEV), West Nile virus, Saint Louis encephalitis virus, La Crosse virus and CHIKV [38,39,40,41,42,43]. The vector competence of Ae. koreicus is even less studied, but the species was experimentally proven to be a competent vector for Dirofilaria immitis, CHIKV and ZIKV [44,45,46]. Moreover, the species is considered to play a role as a vector of the JEV in Asia [47].
A reliable differentiation of exotic mosquito species is important to monitor the spread, initiate early control measurements or understanding the local risk of pathogen transmission. Eradication of exotic mosquito species was only demonstrated to be effective if the populations were detected in an early stage of establishment [48, 49]. From the established exotic mosquito species in Europe, Ae. j. japonicus and Ae. koreicus have the highest morphological similarity. The coloration of the hind femur, the pale basal scales on hind tarsomere IV and the subspiracular patch are considered distinctive characters to morphologically differentiate Ae. koreicus and Ae. j. japonicus [32, 50, 51]. However, these morphological differences are very subtle, and their accuracy for species identification varied between 91 and 100% depending on the respective studies [32, 50, 51]. In addition, the conditions of the samples can make morphological identification even for specialised entomologists difficult. Molecular assays for the differentiation of both species are well established with barcoding, e.g. the nad4 gene [52]. However, PCRs are still expensive, requiring specialised equipment and trained personal [53]. Geometric morphometric analysis of wings is an alternative low-cost method, proven to be suitable to analyse the evolution and population structure and for the correct species identification of mosquitoes. The method showed similar accuracy to molecular barcoding [54] and is even suitable to differentiate cryptic mosquito species, e.g. Culex pipiens pipiens biotype pipiens Linnaeus, 1758 (Cx. p. pipiens), and Cx. torrentium Martini, 1925 [55]. Martinet et al. [56] successfully used wing morphometry to differentiate Ae. j. japonicus and Ae. albopictus. However, their study did not include Ae. koreicus and was only focused on males. Thus, the aim of the present study was to evaluate whether geometric wing morphometrics represents a reliable tool to differentiate Ae. koreicus and Ae. j. japonicus female and male specimens.
Methods
Field work and rearing
Ovitraps were used to collect eggs of Ae. j. japonicus and Ae. koreicus from different sites in the southwestern region of Germany (see Table 1 for the coordinates of the sampling sites) during the summer of 2021, following the field method described by Pfitzner et al. [32]. The sticks with eggs were submerged in water for 3 days, air-dried for 2 days and then submerged again for 3 days to allow the hatching of delayed eggs that had not hatched during initial immersion. The mosquitoes were reared in a controlled environment with a temperature of 26 °C, a relative humidity of 70% and a light:dark cycle of 12:12 h, including a 30-min twilight period. Larvae were fed every 2–3 days with Catfish Chips Nature (Sera, Heinsberg, Germany). Mosquitoes were fed ad libitum using cotton pads soaked with an 8% fructose solution (Carl Roth, Karlsruhe, Germany), which were refreshed every 2–3 days.
Molecular identification
Identification of all specimens was confirmed by DNA barcoding. DNA isolation was performed from one leg per specimen adapting the protocol described by Blattner et al. [57]. Individual legs were placed into 2-ml tubes, and 180 µl ATL buffer (Qiagen) and 20 µl (20 mg/ml) Proteinase K (Qiagen) were added followed by incubation overnight at 56 °C on a shaking thermomixer (400 rpm). Extraction with the DNeasy Blood & Tissue kit (Qiagen) was performed according to the manufacturer’s protocol. Polymerase chain reaction (PCR) amplification of the COI (Ae. j. japonicus) and nad4 gene region (Ae. koreicus) was conducted with the protocol published by Fang et al. [58] and Fonseca et al. [52]. All amplicons were further processed with Sanger sequencing (LGC Genomics, Berlin, Germany), pre-processed with Geneious 7.1.9 (https://www.geneious.com) and compared to GenBank sequences (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Representative sequences for both species have been submitted to GenBank (accession no. OR699056, OR699057, OR723972, OR723973, OR723974).
Wing preparation and landmark collection
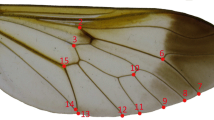
In total, 147 Ae. j. japonicus specimens (77 females and 70 males) and 124 Ae. koreicus specimens (67 females and 57 males) were included in the study. The left wing of each mosquito was removed and mounted in Euparal (Carl Roth, Karlsruhe, Germany) on microscopic slides and dried. Subsequently, the mounted wings were photographed (Olympus DP23, Olympus GmbH, Tokyo, Japan) under 20 × magnification with a stereomicroscope (Olympus SZ61, Olympus GmbH, Tokyo, Japan). The collection of landmark coordinates for 18 wing vein crosses was performed with the multi-point tool in Fiji [59] as bioscience bundle of imageJ [60]. The selected landmarks are consistent with a variety of studies analysing the interspecific wing shape variation of mosquitoes, e.g [54, 61,62,63]. All landmark coordinates were collected by a single observer (author KK). To assess the degree of observer bias in the landmark collection, the measurements were repeated for 20 randomly selected images per species and sex by two observers (authors FGS and LR).
Statistics
The two-dimensional landmark coordinates were used to calculate the centroid size and the superimposed shape coordinates of each specimen with the “gpagen” function in the R package “geomorph”, version 4.0.1 [64]. The centroid size is considered a proxy for wing size and was used to statistically compare the mean wing size of Ae. j. japonicus and Ae. koreicus through an analysis of variance (ANOVA). As mosquitoes are well known for sex-specific differences in their wing size [65], ANOVA was applied separately for the females and males. The allometric effects of the centroid size on the wing shape were assessed with the “procD.lm” function using 1000 permutations [64]. The variability in the superimposed wing shape coordinates between the specimens was visualised by principal component analyses (PCA). In addition, the wing shape coordinates were analysed by linear discriminant analyses (LDA) with the R package “MASS”, version 7.3.58.2 [66] to classify Ae. j. japonicus and Ae. koreicus. Subsequently, the obtained species classification from the LDA was cross-validated (leave-one-out method) to test the classification accuracy. The LDA and cross-validation were also done separately for both sexes since mosquitoes have sex-specific wing shape differences and should not be mixed in the same analyses when researchers are interested in species-specific differences [67]. The mean shape configuration of the 18 landmarks was calculated to visualise differences between female and male Ae. j. japonicus and Ae. koreicus. In addition, the superimposed shape coordinates of each landmark were plotted individually to visually inspect their importance for species identification.
A potential observer effect on the centroid size was assessed with an ANOVA by means of the “prcoD.lm” function in gemorph using 500 permutations [67]. Two ANOVAs were conducted for the wing size of females and males with the three observers as categorial covariate. The effect of different observers on the wing shape coordinates was assessed through the “morphol.disparity” function in gemorph using 500 [67]. Thereby, the morphological disparity, i.e. mean Procrustes variance, was calculated for the three measurements per wing and for the two species. This was also done separately for males and females. Based the resulting morphological disparity only, it is difficult to interpret the influence of disparity due to different observers on the actual species classification accuracy. Therefore, we conducted an additional LDA and reclassified the results with a leave-one-out cross-validation for all specimens, which were measured by three different observers. These analyses involved a dataset with replicated measurements of individual wings and should therefore not be interpreted as final accuracy. Instead, it was conducted to analyse the effect of different observers on the species classification accuracy to get deeper insight into the robustness of geometric morphometrics to differentiate the target species. All statisitical analyses and visulisation were conducted in R, version 4.2.3 [68], including the package ggplot2, version 3.4.0 [69].
Results
The mean centroid size of the female Ae. koreicus specimens was significantly greater than for female Ae. j. japonicus specimens (F1,142 = 5.82, P = 0.017), but no significant difference was observed between the males of both species (F1,125 = 0.22, P = 0.641) (Fig. 1). A low but statistically significant allometric effect on the wing shape could be registered in males (F1,125 = 5.55, R2 = 0.043, P < 0.001) and females (F1,142 = 3.9, R2 = 0.027, P = 0.006). The first two principal components of the PCAs explained 50.3% of the wing shape variation in the females (Fig. 2) and 42.1% of the wing shape variation in the males (Fig. 3). In both sexes, an overlap between the two species-specific clusters was observed (Figs. 2, 3).

Centroid size of Aedes japonicus japonicus and Aedes koreicus for female and male specimens. Grey dots represent the centroid size of one specimen. The black dots mark the mean centroid size with 95% confidence interval as error bars

Principal component analysis of the wing shape variation of female Aedes japonicus japonicus and Aedes koreicus

Principal component analysis of the wing shape variation of male Aedes japonicus japonicus and Aedes koreicus
The leave-one-out cross-validation based on the classification of the LDA revealed a total accuracy of 96.5% for the species identification of the females. Three of 77 female Ae. j. japonicus specimens were misclassified as Ae. koreicus and two of 67 female Ae. koreicus specimens were misclassified as Ae. j. japonicus. The species identification accuracy (leave-one-out method) for the males was 91.3%. Thereby, seven of 70 Ae. j. japonicus specimens and four of 57 Ae. koreicus specimens were falsely classified. The highest mean shape variation between both species and both sexes was observed for landmark 18 (Fig. 4). However, the variation of all superimposed shape coordinates, including landmark 18, showed a strong overlap between both species, indicating that none of the landmarks alone provide enough information for an accurate species identification (Additional file 1: Figs. S1 and S2).

Comparison of the mean shape variation of the superimposed landmarks between Aedes japonicus japonicus (red) and Aedes koreicus (blue)
Observer effect
The centroid size did not significantly differ among the three observers for both, females (linear model, F = 0.276, Z = − 0.751, R2 = 0.005, P = 0.776) and males (linear model, F = 0.219, Z = − 0.848, R2 = 0.004, P = 0.784). The mean Procrustes variance for the repeated measurements per specimen was 0.000439 for the females and 0.00034 for the males and thereby considerably smaller than the mean Procrustes variance observed within the two species, Ae. j. japonicus (females: 0.001686, males: 0.001494) and Ae. koreicus (females: 0.00094, males: 0.001639). This indicates a relatively low observer effect and was also confirmed by the leave-one-out classification with the wing data set of the repeated measurements, which yielded a similar accuracy as the dataset produced by a single observer for the females (95.8%) and an even a higher accuracy for the males (97.5%).
Discussion
Aedes. japonicus japonicus and Ae. koreicus are invasive mosquitoes in Europe, which are established in different countries [15,16,17,18,19, 21,22,23,24,25,26,27,28,29,30,31,32,33,34,35]. Analysis of the vector competence [38,39,40,41,42,43,44,45,46] and host-feeding patterns [36, 37] of the two species indicate a vector capacity of a variety of pathogens for both, e.g. ZIKV and CHIKV. However, monitoring of the spread of both sympatric occurring species is difficult because of their morphological similarity.
The wing size was statistically significantly different between the females but not for the males of both species. On average, female Ae. koreicus showed a larger centroid size than Ae. j. japonicus. However, although statistically significant, the wing size showed a very strong overlap and is not a reliable feature to discriminate between Ae. j. japonicus and Ae. koreicus females. This is a common phenomenon, which was demonstrated for several taxonomic groups including mosquitoes [70,71,72]. The wing size is strongly affected by local environmental factors, e.g. temperature or food availability in the breeding sites [73], resulting in a high intraspecific wing size variability which does not allow a clear species identification based on size, only.
However, it has again been shown that the geometric morphometric analysis of the wing shape is a powerful tool for the identification of mosquitoes. Its use is of particular interest when molecular identification cannot be performed or when damaged mosquitoes hinder an accurate morphological identification. Our study provides the first morphometric information about Ae. koreicus. We demonstrated that Ae. j. japonicus and Ae. koreicus can be identified with a high classification accuracy (96.5% for females, 91.3% for males), which can otherwise morphologically only be distinguished by very subtle differences [51]. In both sexes, the strongest difference between Ae. j. japonicus and Ae. koreicus was observed for landmark 18, i.e. where the media bifurcates into M1+2 and M3+4. However, none of the landmarks alone showed enough divergence to clearly distinguish Ae. j. japonicus and Ae. koreicus. Hence, the full set of landmarks and geometric morphometric analysis is required to differentiate the two species.
We analysed the repeatability of landmark collection by three different observers with a subset of the mosquito wings. The results demonstrated that the observer bias plays no or only a minor role when studying in the centroid size of mosquito wings. However, an observer effect should be considered in the shape analysis. Thereby, the observer bias was lower for males than for females. Unlike most other Dipteran families, mosquitoes have scales on their wing veins, which can obstruct a clear view on the vein crosses. For Ae. j. japonicus and Ae. koreicus, these scales are less dense in males compared to females, i.e. the vein crosses are more easily visible. This likely increases the reproducibility of landmark collection for males. As demonstrated by Lorenz and Suesdek [74], the removal of the wing scales can improve the accuracy and reproducibility in landmark-based geometric morphometrics but also increases the effort for wing preparation. In our study, we did not remove wing scales, but still obtained a high accuracy to classify the two species, even when the landmarks were collected by different observers. This underpins the robustness of geometric wing morphometrics to distinguish Ae. j. japonicus and Ae. koreicus. Nevertheless, when interested in more subtle differences in the wing shape, e.g. intraspecific patterns, the removal of the wing scales and a single observer should still be considered to increase the accuracy of the landmark coordinates.
Conclusions
As previously demonstrated for the separation between Ae. j. japonicus and Ae. albopictus [56], Cx. p. pipiens and Cx. torrentium [55] or Anopheles species [75], our study again demonstrated that geometric wing morphometrics is a powerful tool for the identification of mosquito species. Future research should especially focus on the development of user-friendly tools for a quick landmark collection and subsequent species identification, e.g. using deep learning methods for automatic landmark detection [76].
Availability of data and materials
All data generated by this study and used is presented within this published article and supplementary files. All wing images are stored and available at: https://doi.org/10.5061/dryad.zcrjdfnjn.
References
Bonizzoni M, Gasperi G, Chen X, James AA. The invasive mosquito species Aedes albopictus: current knowledge and future perspectives. Trends Parasitol. 2013;29:460–8.
Medlock JM, Hansford KM, Schaffner F, Versteirt V, Hendrickx G, Zeller H, et al. A review of the invasive mosquitoes in Europe: ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012;12:435–47.
Schaffner F, Medlock JM, Van Bortel W. Public health significance of invasive mosquitoes in Europe. Clin Microbiol Infect. 2013;19:685–92.
Gjenero-Margan I, Aleraj B, Krajcar D, Lesnikar V, Klobučar A, Pem-Novosel I, et al. Autochthonous dengue fever in Croatia, August-September 2010. Euro Surveill [Internet]. European Centre for Disease Control and Prevention (ECDC); 2011;16. Available from: https://doi.org/10.2807/ese.16.09.19805-en.
La Ruche G, Souarès Y, Armengaud A, Peloux-Petiot F, Delaunay P, Desprès P, et al. First two autochthonous dengue virus infections in metropolitan France, September 2010. Euro Surveill. European Centre for Disease Control and Prevention (ECDC); 2010;15:19676.
Marchand E, Prat C, Jeannin C, Lafont E, Bergmann T, Flusin O, et al. Autochthonous case of dengue in France, October 2013. Euro Surveill. European Centre for Disease Control and Prevention (ECDC); 2013;18:20661.
Schmidt-Chanasit J, Haditsch M, Schoneberg I, Gunther S, Stark K, Frank C. Dengue virus infection in a traveller returning from Croatia to Germany. Euro Surveill [Internet]. European Centre for Disease Control and Prevention (ECDC); 2010;15. Available from: https://doi.org/10.2807/ese.15.40.19677-en.
Succo T, Leparc-Goffart I, Ferré J-B, Roiz D, Broche B, Maquart M, et al. Autochthonous dengue outbreak in Nîmes, South of France, July to September 2015. Euro Surveill [Internet]. 2016;21. Available from: https://doi.org/10.2807/1560-7917.ES.2016.21.21.30240.
Annual Epidemiological Reports (AERs) [Internet]. European Centre for Disease Prevention and Control. [cited 2023 Mar 8]. Available from: https://www.ecdc.europa.eu/en/publications-data/monitoring/all-annual-epidemiological-reports.
Giron S, Franke F, Decoppet A, Cadiou B, Travaglini T, Thirion L, et al. Vector-borne transmission of Zika virus in Europe, southern France, August 2019. Euro Surveill [Internet]. 2019;24. Available from: https://doi.org/10.2807/1560-7917.ES.2019.24.45.1900655.
Rezza G, Nicoletti L, Angelini R, Romi R, Finarelli AC, Panning M, et al. Infection with chikungunya virus in Italy: an outbreak in a temperate region. Lancet. 2007;370:1840–6.
Delisle E, Rousseau C, Broche B, Leparc-Goffart I, L’Ambert G, Cochet A, et al. Chikungunya outbreak in Montpellier, France, September to October 2014. Eurosurveillance. European Centre for Disease Prevention and Control; 2015;20:21108.
Grandadam M, Caro V, Plumet S, Thiberge JM, Souarès Y, Failloux A-B, et al. Chikungunya virus, southeastern France. Emerg Infect Dis. Centers for Disease Control and Prevention (CDC); 2011;17:910–3.
Venturi G, Di Luca M, Fortuna C, Remoli ME, Riccardo F, Severini F, et al. Detection of a chikungunya outbreak in Central Italy, August to September 2017. Euro Surveill [Internet]. 2017;22. Available from: https://doi.org/10.2807/1560-7917.ES.2017.22.39.17-00646.
Versteirt V, Schaffner F, Garros C, Dekoninck W, Coosemans M. Van Bortel W. Introduction and establishment of the exotic mosquito species Aedes japonicus japonicus (Diptera: Culicidae) in Belgium. J Med Entomol. 2009;46:1464–7.
Schaffner F, Kaufmann C, Hegglin D, Mathis A. The invasive mosquito Aedes japonicus in Central Europe. Med Vet Entomol. 2009;23:448–51.
Becker N, Huber K, Pluskota B, Kaiser A. Ochlerotatus japonicus japonicas—a newly established neozoan in Germany and a revised list of the German mosquito fauna. Eur Mosq Bull. 2011;29:88–102.
Koban MB, Kampen H, Scheuch DE, Frueh L, Kuhlisch C, Janssen N, et al. The Asian bush mosquito Aedes japonicus japonicus (Diptera: Culicidae) in Europe, 17 years after its first detection, with a focus on monitoring methods. Parasit Vectors. 2019;12:109.
Seidel B, Montarsi F, Huemer HP, Indra A, Capelli G, Allerberger F, et al. First record of the Asian bush mosquito, Aedes japonicus japonicus, in Italy: invasion from an established Austrian population. Parasit Vectors. 2016;9:284.
Seidel B, Duh D, Nowotny N, Allerberger F. First record of the mosquitoes Aedes (Ochlerotatus) japonicus japonicus (Theobald, 1901) in Austria and Slovenia 2011 and for Aedes (Stegomyia) albopictus (Skuse, 1895) in Austria 2012. Entomol Zeitschrift. 2012;122:223–6.
Ibáñez-Justicia A, Kampen H, Braks M, Schaffner F, Steeghs M, Werner D, et al. First report of established population of Aedes japonicus japonicus (Theobald, 1901) (Diptera, Culicidae) in the Netherlands. J Eu Mosq Control Assoc. 2014;32:9–13.
Seidel B, Nowotny N, Bakonyi T, Allerberger F, Schaffner F. Spread of Aedes japonicus japonicus (Theobald, 1901) in Austria, 2011–2015, and first records of the subspecies for Hungary, 2012, and the principality of Liechtenstein, 2015. Parasit Vectors. 2016;9:356.
Krebs T, Bindler P, L’Ambert G, Toty C, Perrin Y, Jourdain F. First establishment of Aedes japonicus japonicus (Theobald, 1901) (Diptera: Culicidae) in France in 2013 and its impact on public health. J Vector Ecol. 2014;39:437–40.
Klobučar A, Lipovac I, Žagar N, Mitrović-Hamzić S, Tešić V, Vilibić-Čavlek T, et al. First record and spreading of the invasive mosquito Aedes japonicus japonicus (Theobald, 1901) in Croatia. Med Vet Entomol. 2019;33:171–6.
Janssen N, Graovac N, Vignjević G, Bogojević MS, Turić N, Klobučar A, et al. Rapid spread and population genetics of Aedes japonicus japonicus (Diptera: Culicidae) in southeastern Europe (Croatia, Bosnia and Herzegovina, Serbia). PLoS ONE. 2020;15:e0241235.
Eritja R, Ruiz-Arrondo I, Delacour-Estrella S, Schaffner F, Álvarez-Chachero J, Bengoa M, et al. First detection of Aedes japonicus in Spain: an unexpected finding triggered by citizen science. Parasit Vectors. 2019;12:53.
Versteirt V, De Clercq EM, Fonseca DM, Pecor J, Schaffner F, Coosemans M, et al. Bionomics of the established exotic mosquito species Aedes koreicus in Belgium. Europe J Med Entomol. 2012;49:1226–32.
Deblauwe I, De Wolf K, De Witte J, Schneider A, Verlé I, Vanslembrouck A, et al. From a long-distance threat to the invasion front: a review of the invasive Aedes mosquito species in Belgium between 2007 and 2020. Parasit Vectors. 2022;15:206.
Capelli G, Drago A, Martini S, Montarsi F, Soppelsa M, Delai N, et al. First report in Italy of the exotic mosquito species Aedes (Finlaya) koreicus, a potential vector of arboviruses and filariae. Parasit Vectors. 2011;4:188.
Montarsi F, Drago A, Martini S, Calzolari M, De Filippo F, Bianchi A, et al. Current distribution of the invasive mosquito species, Aedes koreicus [Hulecoeteomyia koreica] in northern Italy. Parasit Vectors. 2015;8:614.
Suter T, Flacio E, Fariña BF, Engeler L, Tonolla M, Müller P. First report of the invasive mosquito species Aedes koreicus in the Swiss-Italian border region. Parasit Vectors. 2015;8:402.
Pfitzner WP, Lehner A, Hoffmann D, Czajka C, Becker N. First record and morphological characterization of an established population of Aedes (Hulecoeteomyia) koreicus (Diptera: Culicidae) in Germany. Parasit Vectors. 2018;11:662.
Steinbrink A, Zotzmann S, Cunze S, Klimpel S. Aedes koreicus-a new member of the genus Aedes establishing in Germany? Parasitol Res. Springer Sci Bus Med LLC. 2019;118:1073–6.
Kurucz K, Manica M, Delucchi L, Kemenesi G, Marini G. Dynamics and distribution of the invasive mosquito Aedes koreicus in a temperate European city. Int J Environ Res Public Health. 2020;17:2728.
European Centre for Disease Prevention and Control and European Food Safety Authority. Mosquito maps [internet]. Stockholm: ECDC; 2023. Available from: https://ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/mosquito-maps.
Montarsi F, Rosso F, Arnoldi D, Ravagnan S, Marini G, Delucchi L, et al. First report of the blood-feeding pattern in Aedes koreicus, a new invasive species in Europe. Sci Rep. 2022;12:15751.
Schönenberger AC, Wagner S, Tuten HC, Schaffner F, Torgerson P, Furrer S, et al. Host preferences in host-seeking and blood-fed mosquitoes in Switzerland. Med Vet Entomol. 2016;30:39–52.
Turell MJ, O’Guinn ML, Dohm DJ, Jones JW. Vector competence of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J Med Entomol. 2001;38:130–4.
Sardelis MR, Turell MJ, Andre RG. Experimental transmission of St. Louis encephalitis virus by Ochlerotatus j. japonicus. J Am Mosq Control Assoc. 2003;19:159–62.
Huber K, Jansen S, Leggewie M, Badusche M, Schmidt-Chanasit J, Becker N, et al. Aedes japonicus japonicus (Diptera: Culicidae) from Germany have vector competence for Japan encephalitis virus but are refractory to infection with West Nile virus. Parasitol Res. 2014;113:3195–9.
Jansen S, Heitmann A, Lühken R, Jöst H, Helms M, Vapalahti O, et al. Experimental transmission of Zika virus by Aedes japonicus japonicus from southwestern Germany. Emerg Microbes Infect. 2018;7:192.
Sardelis MR, Turell MJ, Andre RG. Laboratory transmission of La Crosse virus by Ochlerotatus j. japonicus (Diptera: Culicidae). J Med Entomol. 2002;39:635–9.
Schaffner F, Vazeille M, Kaufmann C, Failloux AB, Mathis A. Vector competence of Aedes japonicus for chikungunya and dengue viruses. Eur Mosq Bull. University of East London; 2011;141–2.
Jansen S, Cadar D, Lühken R, Pfitzner WP, Jöst H, Oerther S, et al. Vector Competence of the Invasive Mosquito Species Aedes koreicus for Arboviruses and Interference with a Novel Insect Specific Virus. Viruses. 2021;13. Available from: https://doi.org/10.3390/v13122507.
Montarsi F, Ciocchetta S, Devine G, Ravagnan S, Mutinelli F, Frangipane di Regalbono A, et al. Development of Dirofilaria immitis within the mosquito Aedes (Finlaya) koreicus, a new invasive species for Europe. Parasit Vectors. 2015;8:177.
Ciocchetta S, Prow NA, Darbro JM, Frentiu FD, Savino S, Montarsi F, et al. The new European invader Aedes (Finlaya) koreicus: a potential vector of chikungunya virus. Pathog Glob Health. 2018;112:107–14.
Miles JA. Some ecological aspects of the problem of arthropod-borne animal viruses in the Western Pacific and South-East Asia regions. Bull World Health Organ. 1964;30:197–210.
Flacio E, Engeler L, Tonolla M, Lüthy P, Patocchi N. Strategies of a thirteen year surveillance programme on Aedes albopictus (Stegomyia albopicta) in southern Switzerland. Parasit Vectors. 2015;8:208.
Scholte E, Den Hartog W, Dik M, Schoelitsz B, Brooks M, Schaffner F, et al. Introduction and control of three invasive mosquito species in the Netherlands, July–October 2010. Euro Surveill [Internet]. 2010;15. Available from: https://www.ncbi.nlm.nih.gov/pubmed/21087591.
Tanaka K, Mizusawa K, Saugstad ES. A revision of the adult and larval mosquitoes of Japan (including the Ryukyu Archipelago and the Ogasawara Islands) and Korea (Diptera: Culicidae). Contrib Am Entomol Inst. 1979;16:1–987.
Seok S, Kim Z, Nguyen V, Lee Y. The poential invasion into North America and Europe by non-native mosquito, Aedes koreicus (Diptera: Culicidae). J Med Entomol. 2023;2023:1.
Fonseca DM, Campbell S, Crans WJ, Mogi M, Miyagi I, Toma T, et al. Aedes (Finlaya) japonicus (Diptera: Culicidae), a newly recognized mosquito in the United States: analyses of genetic variation in the United States and putative source populations. J Med Entomol. 2001;38:135–46.
Sumruayphol S, Apiwathnasorn C, Ruangsittichai J, Sriwichai P, Attrapadung S, Samung Y, et al. DNA barcoding and wing morphometrics to distinguish three Aedes vectors in Thailand. Acta Trop. 2016;159:1–10.
Sauer FG, Jaworski L, Erdbeer L, Heitmann A, Schmidt-Chanasit J, Kiel E, et al. Geometric morphometric wing analysis represents a robust tool to identify female mosquitoes (Diptera: Culicidae) in Germany. Sci Rep. 2020;10:17613.
Börstler J, Lühken R, Rudolf M, Steinke S, Melaun C, Becker S, et al. The use of morphometric wing characters to discriminate female Culex pipiens and Culex torrentium. J Vector Ecol. Society for Vector Ecology; 2014;39:204–12.
Martinet J-P, Ferté H, Sientzoff P, Krupa E, Mathieu B, Depaquit J. Wing Morphometrics of Aedes mosquitoes from North-Eastern France. Insects. 2021;12. Available from: https://doi.org/10.3390/insects12040341.
Blattner L, Gerecke R, von Fumetti S. Hidden biodiversity revealed by integrated morphology and genetic species delimitation of spring dwelling water mite species (Acari, Parasitengona: Hydrachnidia). Parasit Vectors. 2019;12. Available from: https://doi.org/10.1186/s13071-019-3750-y.
Fang Y, Shi W-Q, Zhang Y. Molecular phylogeny of Anopheles hyrcanus group (Diptera: Culicidae) based on mtDNA COI. Infect Dis Poverty. 2017;6:61.
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012;9:676–82.
Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012;9:671–5.
Lorenz C, Patané JSL, Suesdek L. Morphogenetic characterisation, date of divergence, and evolutionary relationships of malaria vectors Anopheles cruzii and Anopheles homunculus. Infect Genet Evol. 2015;35:144–52.
Simões RF, Wilke ABB, Chagas CRF, de Menezes RMT, Suesdek L, Multini LC, et al. Wing geometric morphometrics as a tool for the identification of Culex subgenus mosquitoes of Culex (Diptera: Culicidae). Insects. 2020;11:567.
Wilke ABB, de Christe RO, Multini LC, Vidal PO, Wilk-da-Silva R, de Carvalho GC, et al. Morphometric wing characters as a tool for mosquito identification. PLoS ONE. 2016;11:e0161643.
Baken EK, Collyer ML, Kaliontzopoulou A, Adams DC. geomorph v4.0 and gmShiny: enhanced analytics and a new graphical interface for a comprehensive morphometric experience. Methods Ecol Evol. 2021;12:2355–63.
Virginio F, Oliveira Vidal P, Suesdek L. Wing sexual dimorphism of pathogen-vector culicids. Parasit Vectors. 2015;8:159.
Venables WN, Ripley BD. Modern applied statistics with S. Springer Science & Business Media; 2003.
Adams DC, Collyer M, Kaliontzopoulou A, Sherratt E. geomorph: Software for geometric morphometric analyses. R package version 3.0.6. Comprehensive R Archive Network; 2018 [cited 2023 Jun 20]; Available from: https://rune.une.edu.au/web/handle/1959.11/21330.
R Core Team. R: A Language and Environment for Statistical Computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2022. Available from: https://www.R-project.org/.
Wickham H. ggplot2. Cham: Springer; 2016.
Gómez GF, Márquez EJ, Gutiérrez LA, Conn JE, Correa MM. Geometric morphometric analysis of Colombian Anopheles albimanus (Diptera: Culicidae) reveals significant effect of environmental factors on wing traits and presence of a metapopulation. Acta Trop. 2014;135:75–85.
Dujardin J-P. Morphometrics applied to medical entomology. Infect Genet Evol. 2008;8:875–90.
Sontigun N, Sukontason KL, Zajac BK, Zehner R, Sukontason K, Wannasan A, et al. Wing morphometrics as a tool in species identification of forensically important blow flies of Thailand. Parasit Vectors. 2017;10. Available from: https://doi.org/10.1186/s13071-017-2163-z.
Barreaux AM, Stone CM, Barreaux P, Koella JC. The relationship between size and longevity of the malaria vector Anopheles gambiae (ss) depends on the larval environment. Parasit Vectors. 2018;2018:1.
Lorenz C, Suesdek L. Evaluation of chemical preparation on insect wing shape for geometric morphometrics. Am J Trop Med Hyg. 2013;89:928–31.
Lorenz C, Marques TC, Sallum MAM, Suesdek L. Morphometrical diagnosis of the malaria vectors Anopheles cruzii, An. homunculus and An. bellator. Parasit Vectors. 2012;5:257.
Rodrigues PJ, Gomes W, Pinto MA. DeepWings©: Automatic wing geometric morphometrics classification of honey bee (Apis mellifera) subspecies using deep learning for detecting landmarks. Big Data Cogn Comput. 2022;6:70.
Acknowledgements
We thank Nicole Sittig for assistance in collecting the samples in the field.
Funding
Open Access funding enabled and organized by Projekt DEAL. FGS, LR, KK and RL are funded by the Federal Ministry of Education and Research of Germany (BMBF) under the project NEED (01Kl2022) and UL by the German Federal Ministry of Food and Agriculture (BMEL) through the Federal Office for Agriculture and Food (BLE) with the Grant No. 2819107A22.
Author information
Authors and Affiliations
Contributions
FGS and RL wrote the main manuscript text. WPP and HJ conducted the field sampling. KK, UL, AH, SJ conducted laboratory work. KK, LR and FGS collected the landmark data. FGS and RL analysed the data. FGS and RL designed the study. All authors revised the manuscript text. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare there are no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1. Figure S1
: Variation of the superimposed shape coordinates for each landmark of the female specimens. Figure S2: Variation of the superimposed shape coordinates for each landmark of the male specimens.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sauer, F.G., Pfitzner, W.P., Jöst, H. et al. Using geometric wing morphometrics to distinguish Aedes japonicus japonicus and Aedes koreicus. Parasites Vectors 16, 418 (2023). https://doi.org/10.1186/s13071-023-06038-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-06038-y