
Abstract
Background
After several years without bluetongue disease, a ruminant illness caused by Culicoides-borne bluetongue virus (BTV), two new autochthonous cases were reported in 2018 in Germany. By contrast, Schmallenberg virus (SBV), another Culicoides-borne virus pathogenic to ruminants, has continuously circulated in Germany since its first emergence in 2011. The disease outbreaks have triggered numerous studies on the biology of the Culicoides vectors, but many ecological details are still obscure.
Methods
Culicoides biting midge species were collected with UV-light traps on 10 farms in Germany, with one trap inside and one trap outside barns on each of the farms. Traps were run once a week for 24 h from January to December 2019. Collected biting midges were morphologically identified, counted and statistically evaluated, with a focus on the Obsoletus Group and the Pulicaris Complex of the ceratopogonid genus Culicoides, which are believed to contain the major virus vectors. Temperature and relative humidity recorded at each trap were linked to the quantity of caught Culicoides. Correlations between relative Culicoides abundance and presence of livestock or type of husbandry were also investigated.
Results
A total of 38,886 Culicoides biting midges were trapped, with most of them belonging to the Obsoletus Group (51.0%) and the Pulicaris Complex (38.8%). The majority of captured specimens were collected in traps inside the barns. Obsoletus Group individuals were caught from late January to the last week of December while Pulicaris Complex individuals were captured from the end of March to early December. The lowest average temperatures at which members of the two groups were collected were 10.7 °C and 12.8 °C, respectively. While temperature had a statistically significant effect on the activity of both the Obsoletus Group and the Pulicaris Complex, relative humidity only significantly affected the activity of the latter. The presence of livestock significantly influenced the number of captured Obsoletus Group, but not of Pulicaris Complex specimens. Inside the barns, no statistical difference was found between numbers of caught Obsoletus Group and Pulicaris Complex specimens in livestock holdings with deep litter and manure scraper or slatted floor husbandry systems.
Conclusions
The almost year-round presence of Obsoletus Group biting midges and the demonstrated high relative abundance of other potential Culicoides vector species inside barns suggest a high risk of indoor virus transmission to ruminants should BTV or SBV circulate locally. Appropriate structural, organisational and vector control measures to reduce biting midge exposure should be implemented.
Graphical Abstract

Similar content being viewed by others


Background
Biting midges (family Ceratopogonidae) are distributed almost worldwide. Four genera of this family (Culicoides, Leptoconops, Lasiohelea, Austroconops) are known to contain species feeding on the blood of vertebrates [1]. With about 1300 described species, the genus Culicoides is the most important of these four genera from an infectiology point-of-view, comprising about 50 species that are potential vectors of disease agents. Major pathogens transmitted by members of the genus Culicoides include bluetongue virus (BTV; family Reoviridae, genus Orbivirus) and Schmallenberg virus (SBV; family Peribunyaviridae, genus Orthobunyavirus), both of which can cause severe infections in domestic and wild ruminants [1,2,3,4]. Bluetongue disease symptoms may include fever, lameness, muscle necrosis and facial oedema, and result in the death of the infected animals [5, 6], while Schmallenberg disease usually presents with milder symptoms such as fever, diarrhoea and a reduction in milk yield [7, 8]. In pregnant female ruminants, however, SBV infections often result in malformation of the offspring, stillbirths or abortions when occurring during a sensitive period of gestation [8, 9]. In addition to the negative effects on animal health, outbreaks of BTV will lead to restrictions on the trade of ruminants and their products. Vaccines are available for both BTV and SBV although they are administered restrictively unless in outbreak situations [10, 11]. Morbidity and mortality, trade restrictions and vaccination entail considerable costs and have a high economic impact on farmers and the livestock industry [8, 12, 13].
Until the outbreak in 2006 in central and northern Europe, BTV had circulated mainly in Africa, Asia and southern Europe where it had primarily been transmitted by Culicoides imicola Kieffer, 1913, an Afro-Asian species also established in the European Mediterranean area [14, 15]. The outbreak of bluetongue in central and northern Europe, where C. imicola was absent, suggested that native Culicoides species were competent to transmit BTV as well. Based on field and experimental findings, four members of the Obsoletus Group of the genus Culicoides (subgenus Avaritia: Culicoides obsoletus sensu stricto Meigen, 1818; Culicoides scoticus Downes & Kettle, 1952; Culicoides dewulfi Goethebuer, 1936; Culicoides chiopterus Meigen, 1830) and three members of the Pulicaris Complex of the same genus (subgenus Culicoides: Culicoides pulicaris sensu stricto Linnaeus, 1758; Culicoides punctatus Meigen, 1804; Culicoides impunctatus Goetghebuer 1920) were subsequently suggested as potential vectors [14, 16,17,18,19].
While no cases of BTV were recorded in Germany from 2010 to autumn 2018, two new cases were detected in cattle in southwestern Germany during routine BTV surveillance activities in December 2018 [13]. As a result, restrictions came into effect, and animals could only be moved from the affected region to unrestricted areas if they had been vaccinated against BTV or had a negative diagnostic test result (e.g. PCR, enzyme-linked immunosorbent assay [ELISA]) [13].
By contrast, SBV emerged in Germany and the Netherlands in 2011 as a completely novel virus spreading rapidly across central Europe [7, 20, 21]. It showed a phylogenetic similarity to viruses of the Simbu serogroup, which includes Akabane virus, a Culicoides-borne virus widely distributed in Asia and Africa [7, 22]. Due to the phylogenetic relatedness of the viruses, researchers concluded that Culicoides species were likely to transmit SBV as well [4, 7, 22,23,24]. Following the repeated detection of SBV in field-collected Obsoletus Group specimens in temperate European countries and laboratory evidence of virus susceptibility in both C. obsoletus and C. scoticus [25], some species of this biting midge group are now considered the main SBV vectors [22]. Unlike BTV, SBV has never disappeared from Germany and Europe but has been circulating cyclically since its first detection in 2011 [4, 9].
Despite the economic importance of Culicoides species as potential vectors of harmful pathogens, a research gap exists regarding distribution, biodiversity and vector competence of the species of this genus. Only a few studies have been conducted so far on the occurrence, phenology and composition of native Culicoides species in Germany and other European countries [e.g. 26–32]. The species mostly found in those studies were members of the Obsoletus Group and the Pulicaris Complex. The globally most important BTV vector, C. imicola, has so far only been detected in Europe in Mediterranean countries [17, 33] and once in southern Switzerland [34], but not further north.
Ultraviolet (UV)-light traps targeting adult biting midges are commonly used in studies on species distribution, activity and abundance of Culicoides [35, 36]. Biting midge monitoring usually focuses on livestock farms, where Culicoides species are most likely to occur, as they can find dung as a breeding substrate and vertebrates as hosts for a blood meal [37, 38]. In most of the studies examining Culicoides activity on farms, a higher number of Culicoides individuals were caught inside barns than outside these structures [26, 28, 33], whereas a greater species diversity was found outdoors [26, 30, 33, 39]. These observations could be attributed to temperature or biting behaviour (endo-/exophagy) of the species [40].
Knowledge about the occurrence, composition and ecology of biting midge vector species is still fragmentary. Efficient management of biting midges and biting midge-borne diseases, however, depends on detailed up-to-date data.
To contribute up-to-date data for Germany, the present study investigated the Culicoides species richness and relative abundance of the Obsoletus Group and Pulicaris Complex in relation to temperature, relative humidity and the presence of livestock inside and outside barns over the course of a calendar year on various German farms. Further, the influence of husbandry systems applied in barns on the occurrence of Culicoides was compared.
Methods
Collection sites
Eight farms with ruminant livestock (cattle, sheep or goat) and two horse farms were selected as sampling sites for the collection of biting midges (Table 1). Of these 10 farms, eight were located in the German federal state of Brandenburg, and one farm each was situated in the states of Thuringia and North Rhine-Westphalia (Fig. 1). The selection criteria for the farms were: (i) at least 10 ruminants or horses kept on the farms; (ii) livestock kept indoors predominantly or all-year round; and (iii) distance between sampled farms at least 5 km. In addition, the selected farms should either have deep litter, be cleaned with a manure scraper or have integrated a slatted floor. The selected barns were solid buildings with large openings that allowed flying insects to enter and leave anytime.

Map of Germany showing the geographical positions of the 10 study farms (A–J) in the federal states of Brandenburg, North Rhine-Westphalia and Thuringia. Letters correspond to the farm IDs listed in Table 1
Trapping
BG-Sentinel UV-light traps (Biogents, Regensburg, Germany) were used to collect Culicoides biting midges once a week for 24 h from January to December 2019. Sampling was conducted when weather conditions were suitable for biting midge activity, i.e. no or only slight rain and wind. On each study farm, one trap was operated inside and one trap outside the barn, approximately 1.5–2 m (top edge of the trap) above the ground. Traps inside the barns were placed at a maximum distance of 5 m from livestock. The outdoor traps were usually located next to the doors or other openings of the barns but at a maximum distance of 50 m from the barns. Traps on the same farm were operated simultaneously. Caught insects were collected in beakers filled with 75% (v/v) ethanol and, after removal from the trap, stored in the alcohol in a dark environment until morphological identification.
Culicoides identification
Collected biting midges were separated from other insects and identified under a stereomicroscope (Leica model M205 C; Leica Microsystems GmbH, Wetzlar, Germany) by wing patterns and other morphological characters, such as number of spermathecae, following the identification keys of Delécolle [41] and Mathieu et al. [42]. Because of the morphological similarity of some species (e.g. female C. obsoletus and C. scoticus), it was not possible to identify all individuals to the species level. If individual midges could not be identified to species, they were assigned to their species complex or group only.
All Culicoides were separated into females and males. Female Culicoides were further classified into engorged (hereafter referred to as ‘blood-fed’) and unengorged specimens.
Climate data collection
Data loggers (EBI 20-TH1; Xylem Analytics, Ingolstadt, Germany) were attached to each trap to record temperature and relative humidity hourly throughout the entire study period. The recorded temperatures and relative humidities were averaged according to the calendar week. Threshold temperatures and relative humidity for the start and the end of seasonal biting midge activity were calculated from the weekly temperature of the first and last annual catches of the Culicoides species group/complex per trap.
Influence of livestock and husbandry systems on the occurrence of Culicoides
To test whether the presence of livestock or the type of husbandry system in barns influenced the occurrence of Culicoides, the farmers provided information on livestock and farm management.
This study included seven deep litter barns, two with manure scrapers and one barn with a slatted floor (Table 1). In barns with deep litter, dung and urine fall onto a bed of straw. The straw is piled up repeatedly in the barn and accumulates over a long period (e.g. 1 year) [43]. In barns with manure scrapers, the area where the cattle defecates and urinates is cleaned several times a day by scraping the dung and urine into a pit [43]. In barns with slatted floors, the droppings disappear through slats into a pit below the slats [43]. In the latter two husbandry systems, only a bit of straw is present in the collection pits which are cleaned several times a year. Due to their similarities, these two husbandry systems were grouped together to analyse an impact on Culicoides activity, and compared with deep litter barns. Only barns mucked out once or twice a year were included in the analysis (Table 1). One of the seven deep litter barns was mucked out weekly, and excluded from this analysis.
Data analyses
For data analysis, identified Culicoides individuals were grouped into Obsoletus Group (including C. chiopterus, C. dewulfi, C. obsoletus s.s., C. scoticus), Pulicaris Complex (including C. pulicaris s.s., C. punctatus, C. impunctatus) and ‘other Culicoides’ (including all Culicoides species not belonging to the Obsoletus Group or the Pulicaris Complex).
Poisson generalised linear models (GLM) were used to check whether the numbers of caught Obsoletus Group and Pulicaris Complex specimens depended on temperature, relative humidity or the presence of livestock. The response variable was ‘biting midges counted’, explained by the predictors ‘temperature’, ‘relative humidity’ or ‘livestock presence’. The latter was a binary variable, set to ‘1’ when livestock was present and to ‘0’ when the livestock did not reside in the barns (c.f. Table 1).
To analyse the influence of the husbandry systems, the different barn sizes and the number of animals on the study farms had to be made comparable. To this end, the reference value of ‘livestock unit’ (LU), which facilitates the aggregation of animals of different species and ages and is based on the average annual number of animals on a farm [43], was employed to adjust livestock numbers. Also, the average number of Culicoides species group/complex per catch and farm was calculated for the indoor catches. These results were divided by the LU of each farm, thus guaranteeing meaningful and comparable statistics across farms. Two sample t-tests were used to compare the adjusted animal numbers between barns with deep litter and barns with manure scraper or slatted floor.
The data were managed in Microsoft Excel 2013 (Microsoft Corp., Redmond, WA, USA) and analysed and visualised in R version 4.1.3 [44] using the packages readxl [45], tidyverse [46] and ggplot2 [47], plm [48] and pglm [49]. In the calculation of whether the presence of livestock influenced the number of Culicoides caught, location-fixed effects were taken into account. For all calculations, P-values < 0.05 were considered significant.
Results
A total of 38,886 biting midge specimens belonging to the genus Culicoides were captured on the 10 farms, of which 63.1% (n = 24,553) were trapped inside and 36.9% (n = 14,333) outside the barns. These data show that Culicoides were caught 1.7-fold more frequently inside the barns than outside.
Overall, 24 different Culicoides species/species groups were determined morphologically, with 13 species/species groups trapped inside the barns and 23 species/species groups trapped outside the barns (Table 2). Of the captured Culicoides biting midges, 51.0% (n = 19,847) belonged to the Obsoletus Group, 38.8% (n = 15,072) belonged to the Pulicaris Complex and 10.2% (n = 3967) belonged to the classification ‘other Culicoides’. The relative proportion of biting midges per group/complex collected inside and outside the barns differed. More Obsoletus Group and Pulicaris Complex specimens were collected indoors than outdoors, while a larger proportion of ‘other Culicoides’ was trapped outdoors than indoors (Fig. 2).

Relative percentages of indoor and outdoor catches of the Obsoletus Group, the Pulicaris Complex and ‘other Culicoides’
By far more females (94.2%, n = 18,689) of the Obsoletus Group were caught than males (5.8%, n = 1158); this also applied to the Pulicaris Complex (females: 99.1%, n = 14,932; males: 0.9%, n = 140) (Additional file 1: Table S1).
Obsoletus Group biting midges were caught from calendar week 4 in January until calendar week 52 in December, both inside and outside the barns (Fig. 3). By contrast, Pulicaris Complex specimens were captured inside the barns from the end of March (calendar week 13) to the beginning of December (calendar week 49). Catches outside the barns containing Pulicaris Complex specimens occurred from calendar week 15 in the spring to calendar week 47 in the autumn. A heatmap (Fig. 3) shows an overview of the square root-transformed collection numbers of the Obsoletus Group and the Pulicaris Complex according to the location of the traps and the calendar week. A square root transformation presentation was applied due to the uneven distribution of the collection numbers and to show the differences between the catches from the different farms and traps more clearly [50].

Heatmap showing the square root-transformed collection numbers of Obsoletus Group and Pulicaris Complex specimens for each farm per calendar week (columns). Position of the traps inside and outside of barns are displayed in the rows
The highest proportion (39.8%) of the total number of Obsoletus Group individuals was caught at the location Lychen (26.8% relative proportion) (Table 3). Of the Pulicaris Complex, 63.9% (69.1% relative proportion) of the individuals were collected in Platkow and 25.4% (20.3% relative proportion) in Heidesee (Table 3). At these two farms, the numbers of captured Pulicaris Complex individuals were higher than those of the Obsoletus Group. Particularly in calendar weeks 30 and 31 in Platkow and in calendar weeks 26 and 32 in Heidesee, the numbers of the Pulicaris Complex exceeded by far those of the Obsoletus Group (Fig. 3).
A total of 794 blood-fed individuals from the Obsoletus Group and 958 blood-fed individuals from the Pulicaris Complex were captured, including at least one blood-fed female per trapping site (Table 4). Most blood-fed females (92.4%) were caught indoors, with the highest number recorded in Platkow, followed by the locations Eichelborn and Lychen (Table 4).
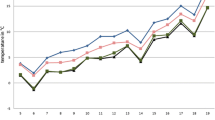
The numbers of total females, blood-fed females and males of the Obsoletus Group and Pulicaris Complex varied throughout the year, both inside and outside the barns, with most individuals caught between May and October (Table 5). With decreasing temperatures, the numbers of collected individuals decreased as well, with the highest collection numbers recorded between calendar week 20 in May and calendar week 35 in August (Fig. 4). The first and last seasonal Obsoletus Group catches were made at 10.7 °C (minimum 3.9 °C, maximum 23.2 °C) and approximately 75.6% relative humidity (minimum 61.0%, maximum 96.7%). For the Pulicaris Complex collection, the average threshold temperature was determined to be 12.8 °C (minimum 4.4 °C, maximum 24.9 °C) and the average relative humidity to be approximately 78.3% (minimum 53.0%, maximum 99.8%). Weekly temperature and relative humidity had no statistically significant effects on the total number of Culicoides collected (t = 1.80367 and t = 1.65711, respectively; both P > 0.05). However, the relative abundance of the Obsoletus Group was significantly correlated with temperature (t = 3.40309, P < 0.001), but not with relative humidity (t = 1.93182, P = 0.5338227). By contrast, the relative abundance of the Pulicaris Complex was significantly correlated with both temperature and relative humidity (t = 4.10764 and 3.47027, respectively; both P < 0.0001) (Table 6).

Total number of Culicoides specimens caught on the 10 farms in relation to the mean temperature measured at each trap
The presence of livestock had a significant impact on the relative abundance of the Obsoletus Group (t = 44.368, P < 0.0001) and ‘other Culicoides’ (t = − 36.004, P = < 0.0001), but did not significantly affect the relative abundance of the Pulicaris Complex (t = 1.229, P = 0.219) (Table 7).
No significant differences of the relative abundance of the Obsoletus Group, the Pulicaris Complex and ‘other Culicoides’ were found between barns with deep litter and barns with manure scraper or slatted floor (Additional file 2: Table S2).
Discussion
By analysing systematic collection data, this study examines the seasonal activity of culicoid Obsoletus Group and Pulicaris Complex biting midges on 10 farms, both inside and outside barns in Germany.
Although the number of Culicoides individuals caught inside and outside barns varies from study to study, most authors report higher collection numbers inside than outside [26, 28, 33]. The same result was registered in the present study (63.1% of all individuals caught inside), with an inside:outside ratio of 1.7:1, a ratio that approximately corresponds to that obtained by Kameke et al. [31] (1.6:1).
In the present study, 23 different Culicoides species/species groups were caught outside the barns—almost twice as many as inside (13 species/species groups). This result is in line with findings reported by Baldet et al. [26], Romón et al. [33] and Sarvašová et al. [39], who also recorded higher species richness outside than inside barns in northern France, northern Spain and eastern Slovakia, respectively. The number of species mentioned in these studies varied, with the numbers identified in this study being somewhere in between. However, in the present study, the number of Culicoides species might actually well be higher than determined, as not all ‘other Culicoides’ could be morphologically identified to the species level. In fact, in addition to the Obsoletus Group and the Pulicaris Complex, ‘other Culicoides’ might also include species contributing to virus transmission. For example, the collections contained Culicoides nubeculosus (captured indoors and outdoors), which has previously been shown to be a competent BTV-vector [51, 52]. Other studies have discussed ‘other Culicoides’ species as potential vectors [53,54,55], but evidence is still missing.
The authors of various studies have raised the question of whether light trap catches underrepresent the abundance of some species of ‘other Culicoides’ [55,56,57]. If this were the case, vector species of this group might play a more important role in the occurrence and spread of Culicoides-borne diseases than commonly thought. Therefore, more efforts should be invested in the elucidation of the ecology of such ‘other Culicoides’.
Despite the high species diversity recorded in the present study, species of the Obsoletus Group and the Pulicaris Complex, particularly C. obsoletus/C. scoticus, C. pulicaris and C. punctatus, were the predominant species indoors and outdoors in the present study. Thus, more than half of the collected individuals could be assigned to the Obsoletus Group, while almost 40% were Pulicaris Complex specimens. Studies from central and northern Europe confirm the general high relative abundance of Obsoletus Group species [14, 28, 31, 32, 58], accounting for up to > 90% of the total catches [29]. The share in captured Obsoletus Group individuals in the present study, however, is lower than that recorded that in other investigations [14, 28, 29, 31, 32], probably due to an unexpectedly large number of Pulicaris Complex individuals being caught during several weeks at two of the 10 study farms (Heidesee and Platkow).
Previous studies have shown that the Pulicaris Complex was predominant at specific locations [59] or at certain times [28, 29, 39]. Mehlhorn et al. [29] suggest that the large number of Pulicaris Complex individuals during specific periods coincides with the simultaneous hatching of many individuals of this complex. This hypothesis might be supported by a study in Slovakia, during which extraordinarily high numbers of C. punctatus, a species of the Pulicaris Complex, were caught on a single trapping day [39], both indoors and outdoors. Based on the results of this study, a possible explanation for the locally and temporally high occurrence of Pulicaris Complex specimens appears to be the strong dependence on both temperature and relative humidity, while the activity of the Obsoletus Group was only correlated with temperature, as also reported in other studies [32, 39, 60]. To determine the true reasons for the restricted local and temporal peaks in the emergence of individuals of the Pulicaris Complex, further data are needed. Factors not investigated in the present study, such as land cover, soil type, soil moisture or vegetation, may be responsible.
It has been described that species of the Obsoletus Group tend to be endophilic while the Pulicaris Complex species C. pulicaris and C. punctatus rather appear to be exophilic [26, 40, 61]. Our results only partially support this notion, as considerable numbers of Obsoletus Group individuals were caught outdoors and many Pulicaris Complex individuals were caught indoors. However, these contradictory findings can possibly be attributed to the open construction of the barns which facilitated the easy entry and exit of biting midges. Meiswinkel et al. [40] also stresses that the building used in their study did not prevent Culicoides from getting inside. In addition, it has been suggested that a certain proportion of the C. punctatus population may adopt endophilic behaviour during their abundance peak [39].
In the present study, most of the indoor-collected Pulicaris Complex individuals were observed on two specific farms (Platkow and Heidesee). On the other study farms, relatively few individuals of the Pulicaris Complex were caught without a tendency for being predominantly collected outside or inside. Based on the literature [31, 56, 62], the types of livestock and animal husbandry practice might have affected the collection numbers of individuals of this complex, although these two parameters also differed between the locations Heidesee and Platkow (Heidesee: horse farm, 10 animals; Platkow: cattle farm, 820 animals). Moreover, the observation that such a high number of Pulicaris Complex specimens were trapped on individual days was not made on any other study farm that kept cattle or horses. Therefore, on the farms included in this study, factors other than the held animal species are likely to influence the behaviour of the Pulicaris Complex species. Different types of barn construction or landscape around the farms may affect the abundance and activity of Culicoides in general [31, 63]. However, these factors are not adequate to explain the high relative abundance or the behaviour of certain species in certain periods of the season.
The activity of Culicoides species varies during the year. For example, Clausen et al. [28] found different activity levels at different times of the year on German farms and, not surprisingly, observed reduced activity of the Obsoletus Group and the Pulicaris Complex during the winter. Mehlhorn et al. [29] also noted that, despite reduced activity, C. obsoletus specimens were caught during the winter. In this context, a possible vector-free period was brought into question [28]. In the present study, the Obsoletus Group was active both indoors and outdoors during all months of the year. Individuals of the Pulicaris Complex were active from March onwards but were also caught during the cold months until early December. Thus, our data support the existence of a vector-low, rather than a vector-free, period.
Biting midges are commonly collected by UV-light traps [29, 32, 35]. Using this approach, females are overrepresented over males [26, 32, 64], but at the same time are more important from a vector ecological point-of-view. As in other studies [26, 39, 40], in the present study the blood-fed females of the Obsoletus Group were mainly trapped indoors. While most of these females were caught during the summer months, Sarvašová et al. [39] reported a decrease in the number of blood-fed females in the barn to a negligible level at that time of the year. These authors assumed this decrease to be caused by the livestock being kept outdoors and the endophagic behaviour of the Obsoletus Complex individuals. The observation that the number of blood-fed Pulicaris Complex individuals caught indoors exceeded that of the Obsoletus Group in Platkow again contradicts the described exophagic behaviour of this complex [26]. On almost all farms included in our study, livestock was kept indoors even during summer nights, probably resulting in more blood-fed biting midge females caught indoors. In general, barns offer sheltered habitats for Culicoides species where they are not directly exposed to wind, precipitation and adverse temperatures [39]. These environmental parameters may decrease Culicoides activity outdoors [63], while individuals staying indoors remain active and are prone to being trapped [39].
The first catches of the Obsoletus Group and of the Pulicaris Complex during the study period were made at the end of January and at the end of March, respectively. The delayed activity of Pulicaris Complex individuals might be attributed to a higher threshold temperature, which was calculated to be 12.8 °C in the present study. By contrast, Kameke et al. [31] measured a threshold temperature of 10.9 °C for the Pulicaris Complex species C. punctatus and C. pulicaris [31]. Due to a lack of data, those authors could not calculate a threshold temperature for the Obsoletus Group. However, based on the observation that the Obsoletus Group was active sooner in the season, they concluded that the threshold temperature had to be higher for the Pulicaris Complex than for the Obsoletus Group [31]; it would appear that our results confirm this assumption.
Silbermayr et al. [65] correlated the occurrence of the genus Culicoides with weather data and identified a significant influence of temperature and relative humidity in Austria, which we could confirm only for the Pulicaris Complex, but neither for the genus Culicoides in general nor for the Obsoletus Group in particular. However, in many other studies, all Culicoides species were considered as a whole [32, 39, 62]. Further studies that have demonstrated a correlation between the abundance of Culicoides species and temperature and relative humidity focus on species that do not occur in Germany, such as C. imicola [66, 67], or which were conducted in other climatic zones [65, 67] or under constant laboratory conditions [66, 68].
The present study was also able to prove statistically that the ‘presence of livestock’ influenced the number of caught individuals of the Obsoletus Group. As opposed to that, ‘presence of livestock’ does not appear to affect the activity of the Pulicaris Complex. This result is in agreement with the stenoecious behaviour of the Pulicaris Complex [69].
Finally, the impact of deep litter, which was removed once or twice a year, and a manure scraper or slatted floor cleaning system on the presence of Culicoides biting midges was examined. Between these two husbandry approaches, there were no significant differences in the catch numbers of either the Obsoletus Group or the Pulicaris Complex. At least for the Obsoletus Group, this result was surprising as some members of this group have been described as preferring dung and manure for larval development [37, 70, 71]. However, on farms examined in the present study that were equipped with manure scrapers or slatted floors, the manure was not completely removed from the barns or their vicinity. According to the farmers, the scraped-up manure was stored in pits under or near the barn or fell into an open pit under the slatted barn floor. Thus, the manure was still available as a breeding habitat until the pits were cleaned out. However, as the results from only nine traps and farms were used for this analysis, further investigations are necessary to make a well-founded statement on whether the husbandry system influences the occurrence of Culicoides.
The considerable duration of seasonal activity of putative Culicoides vector species inside and outside barns demonstrates the importance of spatially and temporally more differentiated analyses of the risk of virus transmission by Culicoides biting midges. It also highlights details on which further research is needed. In addition, the presence of host-seeking Culicoides females on all farms indicates that extensive virus transmission and disease risk can be inferred, provided the virus circulates in the study regions.
Conclusion
Most specimens collected in the present study belonged to the Obsoletus Group and Pulicaris Complex of the ceratopogonid genus Culicoides, which are believed to contain the major vectors of SBV and BTV. These two species groups/complexes were caught on all livestock farms investigated in the present study, highlighting the risk of virus transmission if the virus becomes introduced into Germany. Compared to the Pulicaris Complex, individuals of the Obsoletus Group have a lower threshold temperature for activity and were caught from January to December. Individuals of both the Obsoletus Group and the Pulicaris Complex were caught inside and outside barns, suggesting that virus transmission in and around the barns is possible even in winter. Due to highest collection numbers of Culicoides from late May to August, the transmission risk appears to be the highest during this time period. Winter activity, however, was also observed and should be studied more closely. In addition, further investigations should be considered to explain the sudden high occurrence of the Pulicaris Complex at certain times and locations and to examine the ecology and epidemiological role of Culicoides species not belonging to the Obsoletus Group and the Pulicaris Complex.
Availability of data and materials
All data supporting the conclusions of this article are included within the article and in its additional files.
Abbreviations
- BTV:
-
Bluetongue virus
- ELISA:
-
Enzyme-linked immunosorbent assay
- GLM:
-
Generalised linear model
- LU:
-
Livestock unit
- SBV:
-
Schmallenberg virus
- UV:
-
Ultraviolet
References
Mellor PS, Boorman J, Baylis M. Culicoides biting midges: their role as arbovirus vectors. Annu Rev Entomol. 2000;45:307–40.
Boorman J. Biting midges (Ceratopogonidae). In: Lane RP, Crosskey RW, editors. Medical insects and arachnids. London: Springer: Chapman & Hall; 1993. p. 288–309.
Borkent A, Dominiak P. Catalog of the biting midges of the world (Diptera: Ceratopogonidae). Zootaxa. 2020;4787:1–377.
Beer M, Wernike K. Akabane virus and Schmallenberg virus (Peribunyaviridae). In: Bamford DH, Zuckerman M, editors. Encyclopedia of virology. Oxford: Academic Press; 2021. p. 34–9.
MacLachlan NJ. Bluetongue: pathogenesis and duration of viraemia. Vet Ital. 2004;40:462–7.
Elbers ARW, Backx A, Meroc E, Gerbier G, Staubach C, Hendrickx G, et al. Field observations during the bluetongue serotype 8 epidemic in 2006 I. Detection of first outbreaks and clinical signs in sheep and cattle in Belgium, France and the Netherlands. Prev Vet Med. 2008;87:21–30.
Hoffmann B, Scheuch M, Höper D, Jungblut R, Holsteg M, Schirrmeier H, et al. Novel orthobunyavirus in cattle, Europe, 2011. Emerg Infect Dis. 2012;18:469–72.
Beer M, Conraths FJ, van der Poel WHM. ’Schmallenberg virus’—a novel orthobunyavirus emerging in Europe. Epidemiol Infect. 2013;141:1–8.
Wernike K, Beer M. Re-circulation of Schmallenberg virus, Germany, 2019. Transbound Emerg Dis. 2020;67:2290–5.
Baetza HJ. Eradication of bluetongue disease in Germany by vaccination. Vet Immunol Immunopathol. 2014;158:116–9.
Wernike K, Aebischer A, Audonnet JC, Beer M. Vaccine development against Schmallenberg virus: from classical inactivated to modified-live to scaffold particle vaccines. One Health Outlook. 2022;4:13.
Veldhuis AMB, Santman-Berends IMGA, Gethmann JM, Mars MH, van Wuyckhuise L, Vellema P, et al. Schmallenberg virus epidemic: impact on milk production, reproductive performance and mortality in dairy cattle in the Netherlands and Kleve district, Germany. Prev Vet Med. 2014;116:412–22.
Gethmann J, Probst C, Conraths FJ. Economic impact of a bluetongue serotype 8 epidemic in Germany. Front Vet Sci. 2020;7:65.
Meiswinkel R, van Rijn P, Leijs P, Goffredo M. Potential new Culicoides vector of bluetongue virus in northern Europe. Vet Rec. 2007;161:564–5.
Saegerman C, Berkvens D, Mellor PS. Bluetongue epidemiology in the European Union. Emerg Infect Dis. 2008;14:539–44.
Caracappa S, Torina A, Guercio A, Vitale F, Calabrò A, Purpari G, et al. Identification of a novel bluetongue virus vector species of Culicoides in Sicily. Vet Rec. 2003;153:71–4.
Torina A, Caracappa S, Mellor PS, Baylis M, Purse BV. Spatial distribution of bluetongue virus and its Culicoides vectors in Sicily. Med Vet Entomol. 2004;18:81–9.
Nolan DV, Carpenter S, Barber J, Mellor PS, Dallas JF, Mordue Luntz AJ, et al. Rapid diagnostic PCR assays for members of the Culicoides obsoletus and Culicoides pulicaris species complexes, implicated vectors of bluetongue virus in Europe. Vet Microbiol. 2007;124:82–94.
Savini G, Goffredo M, Monaco F, Di Gennaro A, Cafiero MA, Baldi L, et al. Bluetongue virus isolations from midges belonging to the Obsoletus Complex (Culicoides, Diptera: Ceratopogonidae) in Italy. Vet Rec. 2005;157:133–9.
Rasmussen LD, Kristensen B, Kirkeby C, Rasmussen TB, Belsham GJ, Bødker R, et al. Culicoids as vectors of Schmallenberg virus. Emerg Infect Dis. 2012;18:1204–6.
Wernike K, Hoffmann B, Conraths FJ, Beer M. Schmallenberg virus recurrence, Germany, 2014. Emerg Infect Dis. 2015;21:1202–4.
Sick F, Beer M, Kampen H, Wernike K. Culicoides biting midges—underestimated vectors for arboviruses of public health and veterinary importance. Viruses. 2019;11:376.
Elbers A, Meiswinkel R, van Weezep E, van Kooi E. Schmallenberg virus detected by RT-PCR in Culicoides biting midges captured during the 2011 epidemic in the Netherlands. Emerg Infect Dis. 2013;19:106–9.
Larska M, Lechowski L, Grochowska M, Żmudziński JF. Detection of the Schmallenberg virus in nulliparous Culicoides obsoletus/scoticus complex and C. punctatus—the possibility of transovarial virus transmission in the midge population and of a new vector. Vet Microbiol. 2013;166:467–73.
Barber J, Harrup LE, Silk R, Veronesi E, Gubbins S, Bachanek-Bakowska K, et al. Blood-feeding, susceptibility to infection with Schmallenberg virus and phylogenetics of Culicoides (Diptera: Ceratopogonidae) from the United Kingdom. Parasit Vectors. 2018;11:116.
Baldet T, Delécolle JC, Cêtre-Sossah C, Mathieu B, Meiswinkel R, Gerbier G. Indoor activity of Culicoides associated with livestock in the bluetongue virus (BTV) affected region of northern France during autumn 2006. Prev Vet Med. 2008;87:84–97.
De Deken G, Madder M, Deblauwe I, De Clercq K, Fassotte C, Losson B, et al. Vector monitoring at Belgian outbreak sites during the bluetongue epidemic of 2006. Prev Vet Med. 2008;87:64–73.
Clausen PH, Stephan A, Bartsch S, Jandowsky A, Hoffmann-Köhler P, Schein E, et al. Seasonal dynamics of biting midges (Diptera: Ceratopogonidae, Culicoides spp.) on dairy farms of central Germany during the 2007/2008 epidemic of bluetongue. Parasitol Res. 2009;105:381–6.
Mehlhorn H, Walldorf V, Klimpel S, Schmahl G, Al-Quraishy S, Walldorf U, et al. Entomological survey on vectors of bluetongue virus in Northrhine-Westfalia (Germany) during 2007 and 2008. Parasitol Res. 2009;105:321–9.
Nielsen SA, Nielsen BO, Chirico J. Monitoring of biting midges (Diptera: Ceratopogonidae: Culicoides Latreille) on farms in Sweden during the emergence of the 2008 epidemic of bluetongue. Parasitol Res. 2010;106:1197–203.
Kameke D, Kampen H, Walther D. Activity of Culicoides spp. (Diptera: Ceratopogonidae) inside and outside of livestock stables in late winter and spring. Parasitol Res. 2017;116:881–9.
Collins AB, Mee JF, Doherty ML, Barrett DJ, England ME. Culicoides species composition and abundance on Irish cattle farms: implications for arboviral disease transmission. Parasit Vectors. 2018;11:472.
Romón P, Higuera M, Delécolle JC, Baldet T, Aduriz G, Goldarazena A. Phenology and attraction of potential Culicoides vectors of bluetongue virus in Basque Country (northern Spain). Vet Parasitol. 2012;186:415–24.
Cagienard A, Griot C, Mellor PS, Denison E, Stärk KDC. Bluetongue vector species of Culicoides in Switzerland. Med Vet Entomol. 2006;20:239–47.
Probst C, Gethmann JM, Kampen H, Werner D, Conraths FJ. A comparison of four light traps for collecting Culicoides biting midges. Parasitol Res. 2015;114:4717–24.
Viennet E, Garoos C, Lancelot R, Alléne X, Gardés L, Rakotoarivony I, et al. Assessment of vector/host contact: comparison of animal-baited traps and UV-light/suction trap for collecting Culicoides biting midges (Diptera: Ceratopogonidae), vectors of orbiviruses. Parasit Vectors. 2011;4:119.
Thompson G, Jess S, Murchie AK. Differential emergence of Culicoides (Diptera: Ceratopogonidae) from on-farm breeding substrates in northern Ireland. Parasitology. 2013;140:699–708.
Lassen S, Nielsen S, Kristensen M. Identity and diversity of blood meal hosts of biting midges (Diptera: Ceratopogonidae: Culicoides Latreille) in Denmark. Parasit Vectors. 2012;5:143.
Sarvašová A, Kočišová A, Liptáková E, Hlavatá H, Mathieu B. First insights into indoor and outdoor Culicoides activity related to the risk period for bluetongue virus transmission in eastern Slovakia. Acta Parasitol. 2016;61:743–55.
Meiswinkel R, Goffredo M, Dijkstra EG, van der Ven IJ, Baldet T, Elbers A. Endophily in Culicoides associated with BTV-infected cattle in the province of Limburg, south-eastern Netherlands, 2006. Prev Vet Med. 2008;87:182–95.
Delécolle JC. Nouvelle contribution à lʼétude systématique et iconographique des espèces du genre Culicoides (Diptera: Ceratopogonidae) du Nord‑Est de la France. PhD thesis. Strasbourg: University of Strasbourg; 1985.
Mathieu B, Cêtre-Sossah C, Garros C, Chavernac D, Balenghien T, Carpenter S, et al. Development and validation of IIKC: an interactive identification key for Culicoides (Diptera: Ceratopogonidae) females from the western Palaearctic region. Parasit Vectors. 2012;5:137.
Pain B, Menzi H, Benda I, Döhler H, Eckel H, Euric-Menden B, et al. Glossary of terms in livestock and manure management. 2nd ed. Darmstadt: Association for Technology and Structures in Agriculture (KTBL); 2011.
R Core Team, R: a language and environment for statistical computing; 2022. https://www.R-project.org/. Accessed 22 May 2023.
Wickham H, Bryan J. readxl: read Excel files; 2019. https://cran.r-project.org/package=readxl. Accessed 22 May 2023.
Wickham H, Averick M, Bryan J, Chang W, D’Agostino McGowan L, François R, et al. Welcome to the Tidyverse. J Open Source Softw. 2019;4:1686.
Wickham H. ggplot2: elegant graphics for data analysis. New York: Springer; 2016.
Croissant Y, Millo G. Panel data econometrics in R: the plm package. J Stat Softw. 2008;27:1–43.
Croissant Y. pglm: panel generalized linear models. https://cran.r-project.org/package=pglm. Accessed 22 May 2023.
Legendre P, Legendre L. Numerical ecology. 2nd ed. Amsterdam: Elsevier; 1998.
Foxi C, Meloni G, Puggioni G, Manunta D, Roccigiani A, Vento L, et al. Bluetongue virus detection in new Culicoides species in Sardinia. Italy Vet Rec. 2019;184:621.
Jennings DM, Mellor PS. The vector potential of British Culicoides species for bluetongue virus. Vet Microbiol. 1988;17:1–10.
Veronesi E, Henstock M, Gubbins S, Batten C, Manley R, Barber J, et al. Implicating Culicoides biting midges as vectors of Schmallenberg virus using semi-quantitative RT-PCR. PLoS ONE. 2013;8:e57747.
Balenghien T, Pagés N, Goffedo M, Carpenter S, Augot D, Jacquier E, et al. The emergence of Schmallenberg virus across Culicoides communities and ecosystems in Europe. Prev Vet Med. 2014;116:360–9.
Paslaru AI, Mathis A, Torgerson P, Veronesi E. Vector competence of pre-alpine Culicoides (Diptera: Ceratopogonidae) for bluetongue virus serotypes 1, 4 and 8. Parasit Vectors. 2018;11:466.
Viennet E, Garros C, Gardés L, Rakotoarivony I, Alléne X, Lancelot R, et al. Host preferences of palaearctic Culicoides biting midges: implications for transmission of orbiviruses. Med Vet Entomol. 2013;27:255–66.
Carpenter S, Szmaragd C, Barber J, Labuschagne K, Gubbins S, Mellor P. An assessment of Culicoides surveillance techniques in northern Europe: have we underestimated a potential bluetongue virus vector? J Appl Ecol. 2008;45:1237–45.
Cuéllar AC, Kjaer LJ, Kirkeby C, Skovgard H, Nielsen SA, Stockmarr A, et al. Spatial and temporal variation in the abundance of Culicoides biting midges (Diptera: Ceratopogonidae) in nine European countries. Parasit Vectors. 2018;11:112.
Bartsch S, Bauer B, Wiemann A, Clausen PH, Steuber S. Feeding patterns of biting midges of the Culicoides obsoletus and Culicoides pulicaris groups on selected farms in Brandenburg, Germany. Parasitol Res. 2009;105:373–80.
Magliano A, Scaramozzino P, Ravagnan S, Montarsi F, Da Rold G, Cincinelli G, et al. Indoor and outdoor winter activity of Culicoides biting midges, vectors of bluetongue virus, in Italy. Med Vet Entomol. 2018;32:70–7.
Viennet E, Garros C, Rakotoarivony I, Allène X, Gardès L, Lhoir J, et al. Host-seeking activity of bluetongue virus vectors: endo/exophagy and circadian rhythm of Culicoides in western Europe. PLoS ONE. 2012;7:e48120.
Elbers A, Meiswinkel R. Culicoides (Diptera: Ceratopogonidae) and livestock in the Netherlands: comparing host preference and attack rates on a Shetland pony, a dairy cow, and a sheep. J Vector Ecol. 2015;40:308–17.
Lühken R, Kiel E. Distance from stable affects trapping of biting midges (Diptera: Ceratopogonidae). J Vector Ecol. 2012;37:453–7.
Venter GJ, Labuschagne K, Hermanides KG, Boikanyo SNB, Majatladi DM, Morey L. Comparison of the efficiency of five suction light traps under field conditions in South Africa for the collection of Culicoides species. Vet Parasitol. 2009;166:299–307.
Silbermayr K, Hackländer K, Doscher C, Koefer J, Fuchs K. A spatial assessment of Culicoides spp. distribution and bluetongue disease risk areas in Austria. Berl Münch Tierärztl Wochenschr. 2011;124:228–35.
Venter GJ, Boikanyo SNB, de Beer CJ. The influence of temperature and humidity on the flight activity of Culicoides imicola both under laboratory and field conditions. Parasit Vectors. 2019;12:4.
Ortega MD, Holbrook FR, Llyod JE. Seasonal distribution and relationship to temperature and precipitation of the most abundant species of Culicoides in five provinces of Andalusia. Spain. J Am Mosq Control Assoc. 1999;15:391–9.
Barceló C, Miranda MA. Development and lifespan of Culicoides obsoletus s.s. (Meigen) and other livestock-associated species reared at different temperatures under laboratory conditions. Med Vet Entomol. 2021;35:187–201.
Werner D, Groschupp S, Bauer C, Kampen H. Breeding habitat preferences of major Culicoides species (Diptera: Ceratopogonidae) in Germany. Int J Environ Res Public Health. 2020;17:5000.
Ninio C, Augot D, Dufour B, Dapaquit J. Emergence of Culicoides obsoletus from indoor and outdoor breeding sites. Vet Parasitol. 2011;183:125–9.
Kameke D, Kampen H, Wacker A, Werner D. Field studies on breeding sites of Culicoides LATREILLE (Diptera: Ceratopogonidae) in agriculturally used and natural habitats. Sci Rep. 2021;11:10007.
Acknowledgements
The authors thank the farmers who allowed the traps to be placed on their premises. They are also grateful to former colleagues Nadja Pernat, Martina Pusch, Nathalie Richter, Antje Wehrhan and Maria Will at the Leibniz Centre for Agricultural Landscape Research, Müncheberg, Germany, for their support. The authors would also like to thank Bruno Mathieu (University Strasbourg, France) for his assistance in identifying some Culicoides specimens.
Funding
Open Access funding enabled and organized by Projekt DEAL. The research was part of the CeratoVir project, which was funded by the Federal Ministry of Food and Agriculture (BMEL) through the Federal Office for Agriculture and Food (BLE) under Grant numbers 281B101816 and 281B101916.
Author information
Authors and Affiliations
Contributions
SG, HK and DW designed the study. SG collected the biting midge samples and did the morphological identification, performed the data analyses and wrote the first manuscript draft. HK and DW supervised the study and data analyses and revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Numbers and percentages (in brackets) of Culicoides and other biting midges captured with UV-light traps inside and outside the barns on all study farms.
Additional file 2: Table S2.
Non-significant results of the two sample t-tests comparing the numbers of Culicoides caught in the barns with deep litter, manure scraper or slatted floor.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Groschupp, S., Kampen, H. & Werner, D. Occurrence of putative Culicoides biting midge vectors (Diptera: Ceratopogonidae) inside and outside barns in Germany and factors influencing their activity. Parasites Vectors 16, 307 (2023). https://doi.org/10.1186/s13071-023-05920-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-05920-z