
Abstract
Background
Protozoan pathogens from the genus Cryptosporidium cause the diarrhoeal disease cryptosporidiosis in humans and animals globally. Freshwater biota could act as potential reservoirs or zoonotic sources of Cryptosporidium infections for livestock and people, but Cryptosporidium occurrence in aquatic biota is largely unexplored. The aim of this study was to investigate the occurrence of Cryptosporidium in a range of freshwater organisms in upland rivers across England and Wales.
Methods
Fish were sampled by electrofishing, invertebrate larvae by kick sampling and the otter Lutra lutra and mink Mustela vison through faecal samples collected opportunistically as part of a nation-wide study. PCR targeting the small subunit ribosomal RNA gene was used to detect Cryptosporidium species.
Results
Cryptosporidium occurred in just 0.8% of all the samples and in none of 73 samples from nine invertebrate genera. Cryptosporidium was detected in two of 2/74 fish samples (2.7%), both salmonids, and in 2/92 otter faecal samples (2.17%), but there were no positive samples in mink (0/24) or the bullhead Cottus gobio (0/16).
Conclusions
Low detection rate of human-infective Cryptosporidium species in aquatic fauna indicates they may present a low risk of contamination of some upland freshwaters.
Graphical Abstract

Similar content being viewed by others
Background
Cryptosporidium, a waterborne protozoan parasite, is an important cause of gastroenteritis globally and the second leading cause of moderate to severe diarrhoea in young children in Southeast Asia and Africa [1]. There is currently no vaccine available to protect against cryptosporidiosis, and therapeutic drugs for the treatment of infection are extremely limited [2]. More than 40 Cryptosporidium species have been described, infective to a wide range of hosts, including humans, domestic and wild mammals, birds, reptiles, amphibians and fishes [3]. These parasites transmit between hosts via the faecal-oral route, either directly through contact with an infected individual or indirectly through transmission in faecally contaminated water or, less commonly, food [4, 5]. Water acts as an important medium for the dissemination of Cryptosporidium oocysts throughout the environment, and different water resources (including recreational, drinking, groundwater and wastewater) are frequently contaminated with this parasite [6, 7].
Cryptosporidium in freshwaters can arise from multiple sources, including runoff from contaminated soil or hard-standings, discharge from wastewater treatment facilities, sewage systems and infected livestock grazing in riparian zones [7, 8]. Young cattle (< 6 weeks) especially contribute to the environmental loading of Cryptosporidium parvum while older animals may shed other non-pathogenic Cryptosporidium species to humans [9]. During the first days of an active infection, symptomatic calves can excrete 1 × 1010 oocysts each day that are infective to other susceptible hosts [10]. This high excretion rate of oocysts favours parasite spread and results in Cryptosporidium being ubiquitous in aquatic ecosystems [11]. In upland rural areas, runoff from fields spread with cattle manure as a fertiliser or from fields where cattle are grazing can cause contamination of water courses, but farm management practices, including adequate composting, can reduce the risk [12, 13].
In the event of high rainfall, Cryptosporidium oocysts can be mobilised from agricultural land into surface water, including streams and rivers [12]. Cryptosporidium oocysts can then remain infective within cool and moist environments for 6 months at temperatures between 0 °C and 20 °C [14, 15]. Once within freshwater ecosystems, the oocysts may stay suspended or settle slowly into river sediments where they have the potential to be resuspended at a later stage [16]. Eukaryotic organisms inhabiting these aquatic environments may act as reservoirs or vectors of Cryptosporidium species [6]. Whereas some putative host or vector organisms have been identified, knowledge of the occurrence of Cryptosporidium in others is still patchy. For example, C. parvum and C. hominis oocysts have been isolated from Acanthamoeba species and freshwater sponges, respectively [17, 18], while the predation of Cryptosporidium oocysts has been demonstrated in free-living ciliated protozoa, amoebae and rotifers [19]. In contrast, little is known about the potential for the aquatic larvae of insects to act as Cryptosporidium vectors despite their abundance, diversity and involvement in a wide range of processes in running waters that could bring them into contact with Cryptosporidium oocysts.
Cryptosporidium parvum has also been detected in the microcrustacean, Artemia franciscana, prey of both wild and cultured fish and therefore of potential disease risk to them [20]. Our knowledge of fish-borne cryptosporidiosis is increasing steadily as parasites are being genetically characterised from wild, farmed and ornamental fishes in both marine and freshwater environments ([21], but see [22] for a more detailed review). While is not yet clear whether the presence of these parasite species in fish represents a true infection or mechanical transport, fish could still contribute to the transmission of Cryptosporidium through the aquatic food chain [23]. Notably, Cryptosporidium oocysts have been reported in the faeces of four species of otter including Eurasian otter (Lutra lutra) (in northwest Spain), giant otter (Pteronura brasiliensis), neotropical otter (Lontra longicaudis) (both in northern Brazil) and the North American river otter (Lontra canadensis) (in the Pacific Northwest of the USA). All four otter species are piscivorous predators; whether these parasites can infect and multiply in these host species is unknown [24,25,26].
This study aimed to evaluate Cryptosporidium occurrence and distribution in freshwater hosts from English and Welsh upland river systems. Detecting and characterising human-infecting Cryptosporidium species in fish, semi-aquatic mammals and or invertebrate larvae would alert us of the potential pathogen risk in aquatic biota.
Methods
UK study area
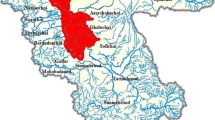
The study sites used were drawn partly from the interdisciplinary Natural Environment Research Council (NERC) ‘DURESS’ project ‘Diversity of Upland Rivers for Ecosystem Service Sustainability’ (see Fig. 1 and [27]) and partially from a UK wide surveillance project on otter corpses sampled opportunistically between 2011 and 2015 (Cardiff University Otter Project, https://www.cardiff.ac.uk/otter-project). In combination, the sites encompassed a range of geologies, soil formations, altitudes and land uses, and in many cases are supported by long-term biological and environmental data from stakeholder collaboration with the UK Environmental Agency, Natural Resources Wales, the Welsh Government and UK Forest Research.

The upland river study sites in England and Wales where fish, semi-aquatic mammals and aquatic invertebrates were collected between 2012 and 2015
Sample collection
Wild salmon (Salmo salar, n = 17), trout (Salmo trutta n = 41) and bullhead (Cottus gobio, n = 16) were collected under licence from 11 upland river sites across Wales in 2012 by electrofishing. Otter faeces (n = 92) were collected from post-mortem examinations (see section UK study area), and mink faecal samples (n = 24) were collected during bankside searches. The otter samples comprised faeces from sub-adults and adults. In this study, adult otters are defined as females with signs of reproductive activity or as males with a baculum > 60 mm; sub-adult females are defined as having a body weight > 2.1 kg with no signs of reproductive activity; and sub-adult males are defined as having a body weight > 2.1 kg with a baculum < 60 mm. Aquatic invertebrate larvae (n = 390, 109 used for method development and controls, and 281 for testing; Table 1) were collected in 2012 through 2-min-long, semi-quantitative kick sampling of six sites in central Wales, identified to the genus level (9 different genera) and preserved in 100% molecular grade ethanol.
Evaluation of Cryptosporidium detection in invertebrate larvae
To evaluate the sample preparation methods used in the detection of Cryptosporidium oocysts from invertebrate larvae, we used an artificial contamination procedure with C. hominis oocysts which we predicated would be unlikely to be present naturally in these samples. Cryptosporidium hominis oocysts were first purified from anonymised stools using the saturated salt flotation method and enumerated using an improved Neubauer haemocytometer. A sample of field-collected invertebrate larvae was then seeded with oocysts via a Nanoject II injector (Drummond Scientific Broomall, PA, USA) with pulled glass capillary needles into the abdomen (targeted soft tissue between the exoskeleton). In this evaluation, three replicates of the following genera were seeded with 200 oocysts each: the ephemeropterans, Baetis and Rhithrogena; the trichopteran, Diplectrona; the plecopterans, Amphinemura and Leuctra; and the dipteran, Simulium. Three replicates of Baetis and Amphinemura were also injected with suspensions containing 5, 25, 50 or 100 oocysts. Controls included positive (200 C. hominis oocysts with and without the addition of Baetis larvae) as well as negative (reverse osmosis water) samples. Prior to DNA extraction, seeded invertebrate larvae were ground up in liquid nitrogen and freeze-thawed in lysis buffer. DNA extractions were performed using the Gentra Puregene Kit according to the manufacturer’s instructions (Qiagen, Hilden, Germany). To detect only the seeded oocysts, we used a real-time C. hominis-specific PCR that amplified a 169-bp fragment of the LIB13 locus [34], duplexed with a commercial non-competitive (primer-limited) internal control PCR (PrimerDesign, Southampton, UK). Thermocycling conditions were 95 °C for 10 min, followed by 55 cycles of 95 °C for 15 s and 60 °C for 60 s, using the forward primer 5′-TCCTTGAAATGAATATTTGTGACTCG-3′, the reverse primer 5′-AAATGTGGTAGTTGCGGTTGAAA-3′ and the minor groove binding probe VIC-5′-CTTACTTCGTGGCGGCGT-3′ MGB-NFQ [28].
Sample preparation, DNA extraction and PCR
Non-seeded invertebrate larvae (281 individual larvae from 9 different genera) were pooled in groups of one to five larvae of the same genus per pool depending on the number collected (Table 1). Each pool was ground in liquid nitrogen and DNA extracted as described above. Paired fish gill samples (n = 74) alongside scrapings of the entire intestinal tracts (n = 73) were recovered and stored at − 20 °C until processed. DNA was extracted using the QIAamp DNA Mini Kit Tissue protocol as described in the manufacturer’s instructions (Qiagen). DNA was extracted directly from otter and mink faeces using the QIAamp Fast DNA Stool Mini Kit according to the manufacturer’s instructions (Qiagen).
Cryptosporidium DNA was identified by nested PCR of the small subunit (SSU) ribosomal RNA (rRNA) gene that detects all Cryptosporidium species [29]. A 1325-bp PCR product was amplified first in a primary PCR using the primers 5′-TTCTAGAGCTAATACATGCG-3′ and 5′- CCCATTTCCTTCGAAACAGGA-3′ [36]. PCR conditions involved HotStar Taq (Qiagen) activation at 95 °C for 15 min, followed by 40 cycles of 94 °C for 45 s, 60 °C for 45 s and 72 °C for 1 min, with a final extension at 72 °C for 7 min. In the secondary PCR, a 2-µl aliquot of the primary PCR product (approx. 830 bp) was used as a template for amplification with forward primer 5′-GGAAGGGTTGTATTTATTAGATAAAG-3′ and reverse primer 5′-CTCATAAGGTGCaGAAGGAGTA-3′ [30, 31] under the same cycling conditions as used for the primary PCR except for annealing at 62 °C. The C. hominis-specific real-time PCR with a duplexed internal amplification control PCR provided useful information about the efficiency of sample preparation and DNA extraction as well as the potential effect of PCR inhibition from invertebrate larvae. This PCR was not used for screening field samples because it was unknown which Cryptosporidium species would be present, so for field samples we used the nested PCR of the SSU rRNA gene, known to provide sensitive and specific amplification of all Cryptosporidium species [29]; at the time of the study, a real-time PCR targeting the SSU rRNA gene was not available in our laboratory. Crptosporidium hominis-positive samples were also tested by a nested PCR targeting the GP60 locus in an attempt to identify the subtype. The primary PCR was performed using primers 5′-ATAGTCTCCGCTGTATTC-3′ (from [32]) and 5′-GGAAGGAACGATGTATCT-3′ (from [33]). PCR conditions consisted of an initial denaturation at 95 °C for 3 min, 35 cycles of denaturation at 94 °C for 45 s, annealing at 50 °C for 45 s and extension at 72 °C for 60 s, with a final extension of 72 °C for 10 min [34]. A secondary PCR product (800–850 bp) was then amplified from a 2-µl aliquot of the primary PCR product using the primers 5′-TCCGCTGTATTCTCAGCC-3′ and 5′-GCAGAGGAACCAGCATC-3′ (both from [32]), with the same cycling conditions as used for the primary PCR.
DNA sequencing of all SSU rRNA amplicons was performed by Source Bioscience (Cambridge, UK).
Ethics statement
Fish sample collection was conducted under licence through National Resources Wales (NRW) under Schedule 5 of the Wildlife and Countryside Act (1981, as amended).
Results
Evaluation of the detection of Cryptosporidium in invertebrate larvae
All three replicate samples of Diplectrona, Simulium and Rhithrogena tested positive for C. hominis after being injected with 200 oocysts, while for the other genera examined two of the three replicate samples were positive (Table 2). All Baetis and Amphinemura larvae seeded with 100 oocysts were positive; for the lower oocyst dosages of 5, 25 and 50 oocysts, at least one replicate sample was negative (Table 3). These results suggested that it was possible to detect oocysts from larvae but that the limit of detection was between 50 and 100 oocysts per extraction, although no oocysts were detected in our unseeded field samples. The cycle threshold (Ct) values between the 200 oocyst C. hominis control and the spiked invertebrate larvae matrix were variable, but always higher than the control value (Table 2).
Occurrence of Cryptosporidium species in riverine hosts
Cryptosporidium species were detected in four out of 471 (0.8%) samples from freshwater biota collected from the full array of English and Welsh sites. The parasites were found in 2/74 fish and 2/116 mammals sampled, but not in the nine genera of aquatic invertebrates (Table 1). Screening of fish gut samples identified C. hominis in one salmon and one trout individual, by SSU rRNA gene sequencing. Unfortunately, these C. hominis samples failed to amplify when tested by the LIB13 and GP60 assays. Of the 92 otter faecal specimens tested, two samples were positive, one for C. muris and the other for Cryptosporidium mink genotype. Cryptosporidium was not detected in the 24 mink samples.
Discussion
This study investigated the occurrence of Cryptosporidium spp. in some of the fauna inhabiting rivers within England and Wales. We detected Cryptosporidium species, including one pathogenic to humans, in Atlantic salmon (C. hominis), brown trout (C. hominis) and otters (C. muris and Cryptosporidium species mink genotype). Although Cryptosporidium was not detected in the nine genera of aquatic invertebrates sampled, we demonstrate that molecular screening for Cryptosporidium in aquatic invertebrate larvae is achievable, although the limit of detection does need to be improved. Such invertebrates may still serve as mechanical vectors for disseminating oocysts, as has been shown for wild filth flies [35], but whether Cryptosporidium naturally occurs in invertebrates remains unknown. In a single study in northwestern Spain, samples containing a community of aquatic invertebrate nymphs and larvae were ground, filtered and screened by immunofluorescent microscopy for Cryptosporidium [36]. The authors of the study detected oocysts in four of these samples, but the number of oocysts detected was not reported and so could be below the limit of detection of our assay. Furthermore, the invertebrate host could not be identified as the whole community was tested together [36].
In the present study, C. hominis oocysts were found in both trout and salmon, confirming the presence of this protozoan parasite in English and Welsh rivers. The lack of amplification at the LIB13 and GP60 loci may well be due to the sensitivity of the assay if low numbers of parasites were present, but we cannot exclude the presence of a C. hominis variant that does not amplify with those more specific primer sets. This human pathogenic Cryptosporidium species is considered to be anthroponotic, and previous reports in animals are rare, suggesting water contamination arose from wastewater [37]. This is not unheard of in upland waters; an outbreak of C. hominis in northwestern Wales was directly linked to inadequate wastewater treatment [38]. Given the rural nature of the study sites, it is likely that C. parvum was present in the river catchments, but the levels would be influenced by the number, type and age of livestock and their husbandry, the time of year, weather events and geographical features of the catchment as well as wastewater inputs.
Limited data are available on the taxonomy, epidemiology and distribution of Cryptosporidium species and genotypes in fish [39]. Highly variable parasite prevalences have been reported, ranging between 0.8% and 100%, with the top end of this range mainly found in farmed juveniles, while larger fish generally show fewer infections over time [40]. Human Cryptosporidium species have been detected at consistently low prevalences (< 1%) in fish hosts [6]. However, to fully assess the prevalence of Cryptosporidium species in fish, further sampling from areas with high levels of human or agricultural faecal contamination is required. There have been inconsistent reports as to whether fish act as natural hosts for human infecting Cryptosporidium species [40,41,42]. Although there are no known reports of fishborne cryptosporidiosis in humans at present, C. parvum oocysts were detected from commercial Atlantic blue crabs (Callinectes sapidus) during animal handling and preparation [43]. Foodborne cryptosporidiosis following the ingestion of these crabs will most likely be prevented during cooking processes, but nevertheless parasite exposure can still result in the contamination of storage areas potentially leading to future disease. Furthermore, 56% of hand swabs taken from urban anglers in Baltimore were positive for Cryptosporidium species [44]. Thus, handling of Cryptosporidium-contaminated animals, including fish, may pose a significant infection risk to humans.
Within freshwater and marine foraging otter populations, reported Cryptosporidium prevalence ranges from 3.9% to 41.7% [24,25,26]; our findings are at the lower end of this range, at 2.17%, but the spatial distribution of the Cryptosporidium-positive otter samples suggests that Cryptosporidium could be widespread. Although Cryptosporidium oocysts have been detected in these aquatic mustelids, the parasite species are not routinely identified. This study is the first report of the Cryptosporidium sp. mink genotype and C. muris in otters. The habitat and diets of the introduced American mink and Eurasian otter overlap to some extent [45], and both species prey on small rodents known to harbour C. muris (although consumption of rodents by otters is relatively rare; see [46]). Interestingly, the C. muris-positive otter did have a rat’s tail in its stomach upon post-mortem examination. Parasite transmission may therefore occur throughout the food chain. Alternatively, otters may have been exposed to both parasite species following the ingestion of faecally contaminated water. As with fish, the presence of these parasites in Eurasian otters does not necessarily indicate an active infection, and it remains unknown whether otters are true hosts of these Cryptosporidium species or merely another reservoir of infection.
The current study has several limitations. First, as detection of Cryptosporidium species was based solely upon PCR assays and, therefore, the detection of Cryptosporidium DNA, there is no direct evidence that the positive fish and otter samples contained the parasite transmission stage, the oocyst. Secondly, as there was no quantification of the positive samples, we do not know whether these hosts had a low or high burden of Cryptosporidium, or, quite possibly in the case of the C. hominis-positive fish, whether the parasite was passing through the gut rather than arising from an active infection. Thirdly, as sampling relied on collection from two separate projects covering different study sites, the spatial distribution of the positive samples could not be analysed. Fourthly, the oocyst limit of detection in invertebrate larvae was relatively high, quite feasibly more than would realistically be expected to be ingested by the larvae. The limit of detection might be influenced by the invertebrate sample matrix through PCR inhibition or losses during DNA extraction, which could explain the variability between Ct values of the different insect genera and the positive control.
Conclusions
We detected Cryptosporidium species in 0.8% of samples from freshwater biota tested in this study. More specifically, the human-infective species, C. hominis, was detected in fish as well as C. muris and Cryptosporidium species mink genotype in otters. The low detection rate of Cryptosporidium species known to be pathogenic to humans may indicate that, in the areas sampled, there is a low risk of human infection from freshwater biota. Further investigation is needed to determine whether the occurrence of Cryptosporidium species and genotypes found in this study represent true infections or whether these animals act as transport vectors, and to further refine detection of oocysts in the plethora of potential invertebrate hosts.
Availability of data and materials
Data is publicly available at the UK CEH Environmental Information Data Centre and can be accessed via the following: https://doi.org/10.5285/84242834-dc78-49a6-83cb-951edac65d18. Sequence data presented in this study, deposited in the NCBI GenBank database, is openly available under Accession numbers: OP999667, OP999668, OP999670 and OP999669.
Change history
09 October 2023
This article has been corrected since original publication; please see the linked erratum for further details.
19 October 2023
A Correction to this paper has been published: https://doi.org/10.1186/s13071-023-05987-8
References
Kotloff KL, Nasrin D, Blackwelder WC, Wu Y, Farag T, Panchalingham S, et al. The incidence, aetiology, and adverse clinical consequences of less severe diarrhoeal episodes among infants and children residing in low-income and middle-income countries: a 12-month case-control study as a follow-on to the Global Enteric Multicenter Study (GEMS). Lancet Glob Health. 2019;7:5.
Korpe PS. The silent reservoir of cryptosporidiosis. Clin Infect Dis. 2021;72:1367–8.
Zahedi A, Paparini A, Jian F, Robertson I, Ryan U. Public health significance of zoonotic Cryptosporidium species in wildlife: critical insights into better drinking water management. Int J Parasitol Parasites Wildl. 2016;5:88–109.
Cope JR, Prosser A, Nowicki S, Roberts MW, Roberts JM, Scheer D, et al. Preventing community-wide transmission of Cryptosporidium: a proactive public health response to a swimming pool-associated outbreak—Auglaize County, Ohio, USA. Epidemiol Infect. 2015;143:3459–67.
Wells B, Paton C, Bacchetti R, Shaw H, Stewart W, Plowman J, et al. Cryptosporidium prevalence in calves and geese co-grazing on four livestock farms surrounding two reservoirs supplying public water to mainland Orkney, Scotland. Microorganisms. 2019;7:11.
Couso-Pérez S, Ares-Mazás E, Gómez-Couso H. A review of the current status of Cryptosporidium in fish. Parasitology. 2022;149:444–56.
Kong Y, Lu P, Yuan T, Niu J, Li Z, Yang B. Cryptosporidium contamination and attributed risks in Yunlong lake in Xuzhou, China. Can J Infect Dis Med Microbiol. 2017;2017:4819594.
Armon R, Gold D, Zukerman U, Kurzbaum E. Environmental aspects of Cryptosporidium. J Vet Med. 2016;3:1048.
Thomson S, Hamilton CA, Hope JC, Katzer F, Mabbott NA, Morrison LJ, et al. Bovine cryptosporidiosis: impact, host-parasite interaction and control strategies. BMC Vet Res. 2017;48:42.
Nydam DV, Wade SE, Schaaf SL, Mohammed HO. Number of Cryptosporidium parvum oocysts or Giardia spp. cysts shed by dairy calves after natural infection. Am J Vet Res. 2001;62:1612–5.
Ahmed SA, Karanis P. Cryptosporidium and cryptosporidiosis: the perspective from the gulf countries. Int J Environ Res Public Health. 2020;17:1–34.
Brankston G, Boughen C, Ng V, Fisman DN, Sargeant JM, Greer AL. Assessing the impact of environmental exposures and Cryptosporidium infection in cattle on human incidence of cryptosporidiosis in southwestern Ontario, Canada. PLoS ONE. 2018;13:4.
Wells B. Controlling Cryptosporidium in the environment. Vet Rec. 2015;13:615–6.
Fayer R. Cryptosporidium: a water-borne zoonotic parasite. Vet Parasitol. 2004;126:37–56.
Shirley DAT, Moonah SN, Kotloff KL. Burden of disease from cryptosporidiosis. Curr Opin Infect Dis. 2012;25:555–63.
Lake IR, Bentham G, Kovats RS, Nichols GL. Effects of weather and river flow on cryptosporidiosis. J Water Health. 2005;3:469–74.
Gómez-Couso H, Paniagua-Crespo E, Ares-Mazás E. Acanthamoeba as a temporal vehicle of Cryptosporidium. Parasitol Res. 2007;100:1151–4.
Masangkay FR, Milanez GD, Tsiami A, Somsak V, Kotepui M, Tangpong J, et al. First report of Cryptosporidium hominis in a freshwater sponge. Sci Total Environ. 2020;700:134447.
Stott R, May E, Ramirez E, Warren A. Predation of Cryptosporidium oocysts by protozoa and rotifers: Implications for water quality and public health. Water Sci Technol. 2003;47:77–83.
Méndez-Hermida F, Gómez-Couso H, Ares-Mazás E. Possible involvement of Artemia as live diet in the transmission of cryptosporidiosis in cultured fish. Parasitol Res. 2007;101:823–7.
Golomazou E, Karanis P. Cryptosporidium species in fish: an update. Environ Sci Proc. 2020;2:13.
Robertson LJ, Clark CG, Debenham JJ, Dubey JP, Kváč M, Li J, et al. Are molecular tools clarifying or confusing our understanding of the public health threat from zoonotic enteric protozoa in wildlife? Int J Parasitol Parasites Wildl. 2019;13:323–41.
Certad G, Follet J, Gantois N, Hammouma-Ghelboun O, Guyot K, Benamrouz-Vanneste S, et al. Prevalence, molecular identification, and risk factors for Cryptosporidium infection in edible marine fish: a survey across sea areas surrounding France. Front Microbiol. 2019;10:1307.
Borges JCG, Lima DDS, Calera BM, Marmontel M, da Silva EM, de Oliveira Moreira AL, et al. Cryptosporidium spp. and Giardia sp. in Neotropical river otters (Lontra longicaudis) and giant otters (Pteronura brasiliensis) in northern Brazil. J Mar Biol Assoc. 2018;98:2153–7.
Gaydos JK, Miller WA, Gilardi KVK, Melli A, Schwantje H, Engelstoft C, et al. Cryptosporidium and Giardia in marine-foraging river otters (Lontra canadensis) from the Puget Sound Georgia Basin ecosystem. J Parasitol. 2007;93:198–202.
Méndez-Hermida F, Gómez-Couso H, Romero-Suances R, Ares-Mazás E. Cryptosporidium and Giardia in wild otters (Lutra lutra). Vet Parasitol. 2007;144:153–6.
Durance I, Bruford MW, Chalmers R, Chappell NA, Christie M, Cosby BJ, et al. The challenges of linking ecosystem services to biodiversity: lessons from a large-scale freshwater study. Adv Ecol Res. 2016;54:87–134.
Hadfield SJ, Robinson G, Elwin K, Chalmers RM. Detection and differentiation of Cryptosporidium spp. in human clinical samples by use of real-time PCR. J Clin Microbiol. 2011;49:918–24.
Zhao W, Xu J, Xiao M, Cao J, Jiang Y, Huang H, et al. Prevalence and characterization of Cryptosporidium species and genotypes in four farmed deer species in the northeast of China. Front Vet Sci. 2020;10:7.
Xiao L, Alderisio K, Limor J, Royer M, Lal AA. Identification of species and sources of Cryptosporidium oocysts in storm waters with a small-subunit rRNA-based diagnostic and genotyping tool. Appl Environ Microbiol. 2000;66:5492–8.
Jiang J, Alderisio KA, Xiao L. Distribution of Cryptosporidium genotypes in storm event water samples from three watersheds in New York. Appl Environ Microbiol. 2005;71:4446–54.
Peng M, Matos O, Gatei W, Das P, Stantic-Pavlinic M, Bern C, et al. A comparison of Cryptosporidium subgenotypes from several geographic regions. J Eukaryot Microbiol. 2001;48:31S.
Alves M, Xiao L, Sulaiman I, Lal AA, Matos O, Antunes F. Subgenotype analysis of Cryptosporidium isolates from humans, cattle, and zoo ruminants in Portugal. J Clin Microbiol. 2003;41:2744–7.
Glaberman S, Moore JE, Lowery CJ, Chalmers RM, Sulaiman I, Elwin K, et al. Three drinking-water-associated Cryptosporidiosis outbreaks, Northern Ireland. Emerg Infect Dis. 2002;8:631–3.
Graczyk TK, Fayer R, Knight R, Mhangami-Ruwende B, Trout JM, da Silva AJ, et al. Mechanical transport and transmission of Cryptosporidium parvum oocysts by wild filth flies. Am J Trop Med Hyg. 2000;63:178–83.
Reboredo-Fernández A, Prado-Merini Ó, García-Bernadal T, Gómez-Couso H, Ares-Mazás E. Benthic macroinvertebrate communities as aquatic bioindicators of contamination by Giardia and Cryptosporidium. Parasitol Res. 2014;113:1625–8.
Vermeulen LC, van Hengel M, Kroeze C, Medema G, Spanier JE, van Vliet MTH, et al. Cryptosporidium concentrations in rivers worldwide. Water Res. 2019;149:202–14.
Chalmers RM, Robinson G, Elwin K, Hadfield SJ, Thomas E, Watkins J, et al. Detection of Cryptosporidium species and sources of contamination with Cryptosporidium hominis during a waterborne outbreak in north west Wales. J Water Health. 2010;8:311–25.
Shahbazi P, Ahmad N. Identification of zoonotic Cryptosporidium parvum in freshwater ornamental fish. J Zoonotic Dis. 2020;3:48–55.
Alvarez-Pellitero P, Quiroga MI, Sitjà-Bobadilla A, Redondo MJ, Palenzuela O, Padrós F, et al. Cryptosporidium scophthalmin sp. (Apicomplexa: Cryptosporidiidae) from cultured turbot Scophthalmus maximus. Light and electron microscope description and histopathological study. Dis Aquat Organ. 2004;62:133–45.
Moratal S, Dea-Ayuela MA, Cardells J, Marco-Hirs NM, Puigcercós S, Lizana V, et al. Potential risk of three zoonotic protozoa (Cryptosporidium spp., Giardia duodenalis, and Toxoplasma gondii) transmission from fish consumption. Foods. 2020;9:12.
Arcay L, Baez de Borges E, Bruzual E. Criptosporidiosis experimental en la escala de vertebrados: I infecciones experimentales, II estudio histopatológico. Parasitol Día. 1995;19:20–9.
Graczyk TK, McOliver C, Silbergeld EK, Tamang L, Roberts JD. Risk of handling as a route of exposure to infectious waterborne Cryptosporidium parvum oocysts via Atlantic blue crabs (Callinectes sapidus). Appl Environ Microbiol. 2007;73:4069–70.
Roberts JD, Silbergeld EK, Graczyk T. A probabilistic risk assessment of Cryptosporidium exposure among Baltimore urban anglers. J Toxicol Environ Health A. 2007;70:1568–76.
Bonesi L, Chanin P, Macdonald DW. Competition between Eurasian otter Lutra lutra and American mink Mustela vison probed by niche shift. Oikos. 2004;106:19–26.
Moorhouse-Gann RJ, Kean EF, Parry G, Valladares S, Chadwick EA. Dietary complexity and hidden costs of prey switching in a generalist top predator. Ecol Evol. 2020;10:13.
Acknowledgements
We thank the Cardiff University Otter Project for samples, and the UK Environmental Agency, Natural Resources Wales, the Welsh Government and the UK Forest Research for fieldwork support.
Funding
This work was funded by the Natural Environment Research Council via the DURESS project (NE/J014818/1) and a GW4 + FRESH studentship (NE/R0115241).
Author information
Authors and Affiliations
Contributions
GR, RMC, ID and SJO designed the study. GR, ID and SO contributed to sample collection. LH, GR and RMC analysed the dataset. LH prepared the original draft with JC. All authors reviewed and edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Licensing for the collection of fish samples was through National Resources Wales (NRW) under Schedule 5 of the Wildlife and Countryside Act (1981, as amended).
Consent for publication
Not applicable.
Competing interests
The authors declare they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hayes, L., Robinson, G., Chalmers, R.M. et al. The occurrence and zoonotic potential of Cryptosporidium species in freshwater biota. Parasites Vectors 16, 209 (2023). https://doi.org/10.1186/s13071-023-05827-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-05827-9