
Abstract
Background
Malaria control efforts are highly skewed towards Plasmodium falciparum while overlooking other Plasmodium species such as P. malariae. A better understanding of the role of Plasmodium species other than P. falciparum is needed to strengthen malaria elimination initiatives. The aim of the present study was to elucidate the contribution of P. malariae to malaria transmission in Cameroon.
Methods
The study was conducted in the Ngatti Health District, a forest–savannah transition area in the Adamawa Region, Cameroon. A total of 497 individuals aged from 1 to 85 years were diagnosed with malaria in November 2020 using a rapid diagnostic test (RDT) and microscopy. Adult mosquitoes were collected between September 2019 and March 2020 by indoor aspiration and identified morphologically and molecularly. The infection status of Plasmodium spp. was also determined by quantitative PCR, and dried blood spots were collected from 156 participants with the aim to detect different Plasmodium species by nested PCR.
Results
The overall Plasmodium prevalence was 50.3%, 51.8% and 64.7%, as detected by microscopy, the RDT and PCR, respectively. Based on the PCR results, P. falciparum was the most prevalent species (43%); followed by co-infections P. falciparum/P. malariae (17%), P. falciparum/P. ovale (1.3%), P. falciparum/P. ovale/P. malariae (1.3%); and then by P. malariae mono-infection (2.5%). The same trend was observed using microscopy, with 35% of participants infected with P. falciparum, 11% co-infected with P. falciparum/P. malariae and 4% infected with P. malariae. The prevalence and parasite density of malaria infection varied significantly with age group (P < 0.05), with the highest prevalence rate observed in children aged 6–10 years (P = 0.0001) while the density of Plasmodium infection increased significantly in children aged < 5 years compared to the other age groups (P = 10−3). Among the 757 Anopheles mosquitoes collected, 737 (97.35%) were An. funestus sensu stricto, 15 (1.9%) were An. gambiae and 5 (0.6%) were An. hancocki. The Plasmodium species recorded at the head/thorax level were P. falciparum and P. malariae, with a sporozoite infection rate of 8.4%; the highest sporozoite infection rate was recorded at Mibellon village (13.6%).
Conclusion
The results of this study reveal the significant contribution of P. malariae, in addition to P. falciparum, to the high malaria transmission rate in this region. These findings highlight the need to deploy initiatives to also tackle this Plasmodium species to eliminate malaria in the region.
Graphical Abstract

Similar content being viewed by others
Background
The past decades have been witness to a substantial reduction in the number of malaria cases and deaths due to malaria worldwide by 37% and 60%, respectively, between 2000 and 2015 [1]. This reduction was mainly due to the use of long-lasting insecticide-treated bed nets (LLINs), indoor residual spraying (IRS), improvements in malaria diagnosis with rapid diagnostic tests (RDTs), affordable prices and easy access to the artemisinin-based combination therapy (ACT) [1, 2].
Despite past and ongoing efforts to eliminate malaria, the disease remains endemic in some parts of the world, with > 90% of cases recorded in the sub-Saharan region of Africa. This is largely due to the increasing resistance of mosquitoes to insecticides, which has diluted the effectiveness of malaria control measures [1]. Hence, there is an urgent need to find new strategies to eliminate or even eradicate this disease. In the case of malaria, the interaction between humans and vectors contributes to maintaining the circulation of malaria in endemic areas where hosts, even if they are highly immunised, constitute the infectious reservoir for plasmodial species [3, 4]. It has been reported that 16 Anopheles species can be considered to vectors of malaria disease to humans in Cameroon [5,6,7], with the most competent vectors being Anopheles gambiae sensu stricto (An. gambiae s.s.), An. coluzzii, An. arabiensis, An. funestus, An. nili and An. moucheti [6, 8]. To date, the malaria control programme in Cameroon has focused on the elimination of Plasmodium falciparum and, to a certain degree, Plasmodium vivax, making them the most studied Plasmodium species responsible for severe cases of malaria disease. However, Plasmodium malariae and Plasmodium ovale, which have been reported to be widely distributed in most malaria-endemic regions of the world, are less studied and difficult to identify by light microscopy because the low level of parasitaemia is often below detection thresholds of microscope readers. This failure is the result of sub-microscopic malaria, which accounts for 20–50% of all human-to-mosquito transmissions in sub-Saharan Africa [9]. A P. malariae infection is usually benign, but could cause morbidity such as severe anaemia, acute renal failure and splenomegaly [10, 11]. It is also responsible for quartan fever and can remain in the host for decades, complicating the fight against this disease [12]. In the case of co-infection with P. malariae and P. falciparum, it has been reported that P. malariae infection can reduce the burden of parasitaemia of P. falciparum infection by up to 30% and contribute to the reduction of the pathology due to P. falciparum as the people become even more asymptomatic [13,14,15]. A study carried out by Yman et al. [16] in Tanzania showed a reduction of P. malariae prevalence when the prevalence of P. falciparum decreased. These results demonstrate that there is a need for more information regarding the epidemiology of non-falciparum species in the context of the distribution of malaria vaccines targeting only the P. falciparum antigen. Four Plasmodium species have been recorded to circulate in Cameroon: P. falciparum, P. ovale, P. malariae and P. vivax [5, 17,18,19]. Plasmodium falciparum remains the predominant Plasmodium species followed by P. ovale and P. malariae, with average prevalence rates of infection of 95%, 3% and 1%, respectively [5]. These prevalence rates vary according to the localities sampled and methods used to determine the species. Malaria elimination also needs to consider the identification and contribution of non-falciparum parasites to the malaria burden. Most studies to date focus on P. falciparum malaria and especially on symptomatic malaria cases in patients aged < 15 years [20], even though the malaria reservoir is expected to be mainly concentrated in the asymptomatic human population when the transmission is low (asymptomatic malaria was defined as the presence of Plasmodium in a patient with an axillary temperature of < 37.5 °C). In the study reported here, we first conducted an entomological survey to identify the vector competent to transmit P. malariae. Then we used molecular, microscopic and RDT techniques to evaluate the dynamics of P. falciparum and P. malariae infection transmission in all age groups of the study population.
Methods
Study areas
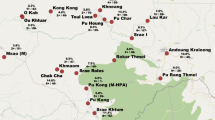
The entomological survey was performed by collecting mosquitoes in six locations between September 2019 (Nyakon, Mbougam, Mibellon, Ngatti) and March 2020 (Banyo, Panyere and Meng) (Fig. 1). The epidemiological survey was carried out in November 2020 in four villages: Moinkoing, Mibellon and Mbougam chefferie, all in the Ngatti health district located in the Adamawa Region of Cameroon, Bankim sub-division. The study carried out by Tchouakui et al. [21] showed a P. malariae/P. ovale or P. vivax infection rate of 4% in the mosquito population of Mibellon village, one of the villages in our study area.

Map of collection sites
A sampling of participants
The sample size of the human population was calculated following the formula: s = z2 × P × (1 − P)/m2 [22], where s = sample size for infinite population and z = Z-score (a Z-score of 1.96 was used with a 95% confidence level), P = the percentage of population probability (assumed to be 50% = 0.5) and m = the margin of error (taken to be 5% = 0.05). All people from year 1 and thereafter were considered for blood collection.
Blood collection and staining
During the survey, malaria diagnosis was carried out using thick blood smears, RDTs (CareStart™ Malaria Pf/PAN (HRP2/pLDH) Combo, Access Bio Inc. 65 Clyde Road. Somerset, NJ, 08873). and PCR analyses. Body weight, temperature, gender and age of subjects sampled in the study were taken into account. The blood was collected on slides in all the localities and on Whatman FTA cards (Sigma-Aldrich, St. Louis, MO, USA) only in Mibellon and Moinkoing villages.
Thick blood smears collected from the finger of each subject sampled in the study were air-dried and stained with 10% Giemsa solution for 20 min. The slides were screened by direct microscopy (100×) to visualise the different species of Plasmodium parasites. Parasite density was defined as the number of Plasmodium parasites per microlitre of blood, counted against 200 leukocytes and expressed according to the WHO guidelines as the number of asexual parasites per microlitre of blood, assuming a leukocyte count of 8000 cells/μl. Samples of blood drops were deposited on Whatman FTA cards and dried for 24 h at room temperature, following which they were stored in clean zippered bags until DNA extraction.
DNA extraction
DNA was extracted from blood on Whatman paper using the Chelex protocol. Briefly, 400 µl of 5% chelex100 was added to an Eppendorf tube containing a blood sample on filter paper. The tubes were placed in a shaking dry heater and incubated at 56 °C for 1 h, following which it was incubated for 30 min at 95 °C in a shaking heater. The tube was then removed and centrifuged at a maximum of 13,500 rpm for 10 min. The supernatant was then transferred into a labelled tube and stored at − 20 °C [23].
Real-time PCR for Plasmodium species identification
Molecular analysis
Real-time PCRs (RT-PCRs) for the identification of Plasmodium species infections were performed with all samples using the nested protocol with some minor modifications to differentiate the various species of Plasmodium [24]. A species-specific region of the 18S ribosomal DNA (rDNA) of Plasmodium was amplified. In the first reaction, PCR amplifications were performed in a 15-μl volume containing 1.5 μl PCR buffer A, 0.75 μl 25 mM MgCl2, 0.12 μl 25 mM dNTPs, 0.51 μl forward and reverse primers (10 mM), 0.12 μl Kapa Taq, 10.49 μl ddH2O and 1 μl DNA. A 5-μl sample of the first reaction’s product was used as a template in a second nested reaction to specifically differentiate P. falciparum, P. vivax, P. ovale and P. malariae products. The temperature profile for the second PCR was: 5 min at 95 °C, followed by 25 cycles of 1 min at 94 °C, 2 min at 58 °C, 2 min at 72 °C; with a final cycle at 72 °C for 10 min. The cycles were repeated 30 times for the second reaction at the same cycling parameters. After electrophoresis in 1.5% agarose gel and staining with Midori green, the amplicon products were visualized under UV light. Plasmodium identification was made based on amplicon sizes.
DNA extraction from mosquitoes, species identification and detection of Plasmodium parasite
Female mosquitoes were captured inside houses using an electric aspirator during the early morning hours of between 05.30 a.m. and 08.00 a.m. Female Anopheles mosquitoes collected were morphologically identified using morphological keys [25]. These mosquitoes were kept in a tube containing the desiccant (silica gel) for further molecular analysis. Molecular identification was performed using a cocktail PCR to determine the species [26].
All female Anopheles mosquitoes were dissected individually into between heads, thoraxes and abdomens. Genomic DNA (gDNA) was extracted from each body part (head + thorax and abdomen) using the LIVAK method [27], and the Plasmodium sporozoite infection rate was assessed using the TaqMan assay [28]. As a last step, nested PCR was conducted on all positive samples confirm the results of the Taqman assay and discriminate the P. ovale, P. vivax and P. malariae species [24].
Epidemiological and statistical analyses
Data analysis was conducted using the Microsoft Excel sheet (Microsoft Corp., Redmond, WA, USA), GraphPad Prism V8.02R software (GraphPad Software Inc., San Diego, CA, USA). Data were summarised into means and standard deviations (SD), and percentages were used in the evaluation of the descriptive statistics. Proportions were compared using the Chi-square test (χ2). Parasitaemia was log transformed before analysis. Means were compared using an independent sample t-test and analysis of variance where appropriate; Kruskal–Wallis tests were used to make associations between P. falciparum, P. malariae and mixed infection and other variables of interest. P-values < 0.05 were considered to indicate significance, and a 95% confidence interval (CI) of geometric means that did not overlap was considered to indicate a significant difference.
Results
Mosquito species composition
A total of 757 Anopheles were collected during the study period. Three species were identified by evaluation of morphological characteristics: Anopheles funestus sensu stricto (An. funestus s.s.) (n = 737; 97.35%), Anopheles gambiae sensu lato (An. gambiae s.l.) (n = 15; 1.9%) and Anopheles hancocki (n = 5; 0.6%).
Plasmodium sporozoite and oocyst infection rate
The overall infection rate for Plasmodium sporozoites was 8.4%, recorded in An. funestus s.s. and An. gambiae s.l. (Table 1). The highest infections rate with P. falciparum (10.8%), P. OVM (P. malariae/P. ovale or P. vivax) (2.5%) and mixed infections (P. falciparum and P. OVM) (0.4%) were recorded in Mibellong village. Mosquitoes infected with P. OVM were found only in the Mibellong and Tibati localities, and two mosquitoes were found to be infected with a mixed infection (P. falciparum and P. OVM) in Mibellong and Meng (Table 1).
The Plasmodium oocyst infection rate was considered only in samples collected in 2019, and the results showed that mosquitoes from Mibellong, Mbougam and Niakon were infected by P. falciparum with a prevalence of 22.2%, 13.9% and 8.6%, respectively, followed by P.OVM with a prevalence of 7.5%, 5.4% and 4.3%, respectively (Table 1). The mixed infection rate (P. falciparum and P. OVM) was observed only in Mibellong (1.8%) and Mbougam (1.1%).
The nested-PCR results confirmed that the samples infected by P.OVM were P. malariae for those with successful amplification (Table 1).
Taken together, these observations indicate that P. falciparum and P. malariae are circulating in these localities and, therefore, that these localities are a good place to conduct an epidemiological survey of P. malariae.
Prevalence of multispecies malaria infection
For the microscopy analyses, samples were collected from 497 people (213 [42.8%] males, 284 [57.2] females) enrolled in this study. Due to some amplification issues, only 156 samples were analysed by PCR, at two locations (Mibellon and Moinkoing), from 67 (43%) males and 89 (57%) females, respectively. The prevalence of malaria infection assessed by microscopy, RDTs and PCR on the same set of samples (156) was 47.4%, 43.7% and 64.7%, respectively. The general prevalence of infection taking into account the entire study population was 51% (252/494) for microscopy and 51.8% (252/484) for microscopy (Table 2). There was a significant difference between the prevalence of Plasmodium spp. detected by PCR, microscopy and the RDT (P < 0.01).
Prevalence of asymptomatic malaria by microscopy, RDT and PCR.
Using the same sample set used as for PCR, 93.3% (147/157) of the individuals sampled were asymptomatic. Microscopy, RDTs and PCR showed that 44.8% (95% CI: 37.0–59.1), 43.7% (95% CI: 33.9–55.6) and 62.5% (95% CI: 37.0–59.1) of asymptomatic people, respectively, were infected with Plasmodium spp.
Species-specific infection prevalence
Among the 156 samples analysed by PCR, 66 (42.3%) tested positive for P. falciparum, four (2.5%) for P. malariae, 27 (17.1%) for P. falciparum/P. malariae, two (1.3%) for P. falciparum/P. ovale and two (1.3%) for P. falciparum/P. ovale/P. malariae (Fig. 2). Also, 497 samples were analysed by microscopy, of which 174 (35%) were positive for P. falciparum, 21 (4%) for P. malariae, 57 (11%) for P. falciparum/P. malariae and two (0.4%) for P. falciparum/P. ovale (Table 3). No samples were infected with P. vivax. The general prevalence of infection detected by microscopy in Mibellon (64.8%), Moinkoing (54.6%) and Ngatti (51%) villages was high, while the lowest prevalence of 12% was recorded in Mbougam village (Table 3). Among these villages, the highest proportion of mixed infections (P. falciparum/P. malariae) was recorded in Mibellon (25.7%) (Table 3). Microscopy showed that P. malariae prevalence was 6%, 5.4%, 3.3% and 0% for Moinkoing, Mibellon, Ngatti and Mbougam chefferie villages, respectively. The prevalence of infection as assessed by PCR and per village was determined in Mibellon and Moinkoing only. The results showed an overall prevalence of 64% and 53.9%, with 3.2% and 1.5% infection with P. malariae, in Mibellon and Moinkoing, respectively (Table 3).

Schematic representation of the number of Plasmodium parasites per species (n = 157 samples). The circles indicate the number of samples that tested positive for each Plasmodium species (blue: Plasmodium falciparum; yellow: Plasmodium malariae; green: Plasmodium ovale). The sections where circles overlap represent the number of co-infections of each combination of > 1 Plasmodium species
According to age groups, the highest P. falciparum prevalence of 47.7% (95% CI: 34–64) was observed among older children aged 6–10 years, followed (in order of decreasing prevalence) by teenagers aged 11–15 years (36.7%, 95% CI: 44–57%), infants aged 1–5 years (34.7%, 95% CI: 26–49%), young adults aged 16–20 years (34%, 95% CI: 19–56%) and adults, with the latter age group showing the lowest prevalence of 22% (95% CI 19–26%) (Fig. 3). The prevalence of mixed infection (P. falciparum/P. malariae) followed the same trend, with the highest prevalence of 27% observed in children aged 5–10 years, followed (in order of decreasing prevalence) by subjects in the age groups 1–5, 10–14, 16–20 and > 21 years, with 22%, 14.2%, 4.5% and 1% infection rates, respectively (Fig. 3). For P. malariae mono-infection, the highest prevalence (9%) was recorded in subjects aged 16–20 years, followed a prevalence of 7.1% in subjects aged 11–15 years. The prevalence of P. malariae obtained in subjects aged 6–10, 1–5 and 21+ years was 3.4%, 3.3% and 2%, respectively (Fig. 3). In general, the probability of having a high prevalence of infection with P. falciparum and mixed infections (P. falciparum/P. malariae) was highest in children aged 6–10 years (odds ratio [OR] = 4.2, 95% CI 2.3–7.6%; P = 0.0001), while the younger adolescents (aged 10–14 year) were more infected with P. malariae with a prevalence of 9%.

Prevalence of Plasmodium infection by age group.
The overall geometric mean P. falciparum and P. malariae parasitaemia was 4354.7 ± 7991.6 and 606.1 ± 572.9 parasites/μl, respectively. The highest parasitaemia was recorded in children aged 1–5 years, at 5873.9 ± 8910.2 and 854.4 ± 608.4 parasites/μl for P. falciparum and P. malariae,, respectively. The lowest parasitaemia was observed in adults (age group: 21–85 years), at 1901.8 ± 2389 and 215 ± 119.8 parasites/μl for P. falciparum and P. malariae, respectively (Fig. 4). The mean of asexual parasite densities did not follow the that of prevalence of infection; hence, the density of Plasmodium infection increased significantly in children aged < 5 years compared to the other age groups (Fig. 4), decreasing from the younger age group to the older ones, leading to a significant reduction in the trophozoite density in subjects in the older age groups (Kruskal Wallis test, H = 20.03, P = 10−3).

Parasite density by age groups
Discussion
In this study we assessed the transmission dynamics of P. falciparum and P. malariae infections in mosquito and human populations. The study produced strong evidence that malaria remains highly prevalent in the study locations in Cameroon.
An. funestus is the predominant malaria vector in Central Cameroon
Three anopheline species were identified in the study area: An. funestus s.s., An. gambiae s.l. and An. hancocki, of which An. funestus s.s.(97.3%) was the most prevalent species. Our results are similar to those obtained by Tchouakui et al. in Mibellon [29], where there was 100% An. funestus prevalence. Our findings vary slightly from those of Tchouakui et al. [21] who recorded 95% An. funestus s.s. in the same village and from those of Menze et al. [30] who observed 80.1% prevalence, possibly due to the season in which the mosquitoes were collected.
The focus on the collection of indoor blood-fed mosquitoes may have prevented us from collecting the outdoor resting species of subfamily Anophelinae. Otherwise, our results showed that An. funestus is the dominant species in this locality and that the prevalence could vary according on the season since the different collections did not take place during the same season. To the contrary, in Tibati, we recorded 94.4% An. funestus prevalence compared to 55.2 %and 16% prevalence in collections from 2015 and 2017, respectively, by Feufack-Donfack et al. [31]. This difference in mosquito distribution in Tibati could be due to the month of mosquito collection, as our work was conducted during the dry season when the temporary breeding sites necessary for An. gambiae development become scarce.
High Plasmodium sporozoite and oocyst infection rates
Both An. funestus and An. gambiae were infected with Plasmodium spp. A slight increase in the overall infection rate was observed in An. funestus in the present study compared to samples collected in 2017 in Mibellon by Menze et al. [30] (5% vs 13.6%; X2 = 3.43, P = 0.06) and also to samples collected in Tibati in 2017 by Feufack-Donfack et al. [31] (5.8% vs 2.04%; X2 = 3.14, P = 0.07). Although not significant, this increase in sporozoite infection rate could be due to the reduction in efficacy of LLINs (approx. 3 years) that were massively distributed in this part of the country in 2015. The prevalence of infection was also low in another location.
The significantly higher overall infection rate found in the abdomen of the mosquitoes compared to the head + thorax (34.2% vs 12.3%; X2 = 48.98, P = 0.0001) and, in particular, the infection rate by P. malariae (8.2% vs 1.9%; X2 = 15.08, P = 0.0001) of mosquitoes collected in 2019 strongly support the role of physical barriers played by the mosquito midgut and immune genes in the process of oocyst migration to the salivary gland [32]. In addition, the large difference in sporozoite infection between P. falciparum and P. malariae could also be due to the lifespan of mosquitoes, which is generally no longer than 3 weeks in nature. In fact, P. malariae has a longer sporogonic cycle (21 days) [33] than P. falciparum (14 days), which may explain the reduced likelihood of finding P. malariae sporozoites in the mosquitoes. The study site of Mibellon had a high infection rate with P. falciparum and P. malariae; this can possibly be explained by the perennial presence of the malaria vector, mainly An. funestus, during the year in Mibellon. The high infection rate of oocysts and sporozoites was recorded among the mosquito population of Mibellon village despite the human factors, vector immunity, midgut microbiota [34] and midgut epithelium [35], all of which are the causes of the reduction in the number of malaria parasites in the mosquito host. These results point to Mibellon as the area of high malaria endemicity and suggest that this location and surrounding villages are suitable for parasitological investigation of P. malariae in the human population.
Multispecies malaria infection prevalence in human population
The high prevalence of infection and high parasitaemia indicate that the Ngatti health district is a high-risk zone for malaria transmission. This is supported by earlier reports of the prevalence of malaria infection in the Adamawa Region varying from 8.1% to 10.6% in symptomatic children aged between 2 and 9 years [5, 20, 31]. Moreover, the mode of malaria transmission is known to be influenced by environmental factors, such as temperature, rainfall and humidity. The high sporozoite infection rate is the main contributor to the maintenance of malaria endemicity in Mibellon. The 64.7% prevalence of malaria infection recorded in this study was lower than expected. Abdulraheem et al. [36] recorded a high prevalence (80%) of malaria in the asymptomatic population in Nigeria using PCR; this prevalence was almost twofold higher than that obtained by microscopy (47%). Similar results were obtained in the DRC by Nundu et al. [15], who recorded 62% and 33% infection rate of asymptomatic individuals using PCR and microscopy, respectively. The low Plasmodium spp. infection rate recorded in the present study could be due to several factors, such as the sampling season and the molecular method used for species identification. The PCR test used in this study targeted the 18S rRNA gene [28] whereas the studies cited above targeted the mt gene cox III [37] in the PCR assays. Unlike the standard 18S rRNA PCR method, the cox III method is able to detect an additional 10–50% more cases with high sensitivities for all four Plasmodium species due to the high number of parasite copies (20–150) found in cox III compared to 18S (4–8 copies) [37].
The results also revealed an association between the prevalence of malaria and age at infection. The highest prevalence of malaria (P. falciparum and Plasmodium mixed) infection was recorded in children in the age group 6–10 years. Regarding this result, it should be noted that malaria control campaigns target children aged < 5 years of age and pregnant women, with parents advised to ensure that children aged < 5 years, who are considered at high risk of contracting malaria, sleep under a LLIN. This possibly explains the reduction in the prevalence of infection among children in this young age group. However, the good use of ITNs for children aged < 5 years may impede or delay the development of malaria protective immunity in children aged 6–10 years [38, 39]. Also, the prevalence of infection with P. falciparum decreased significantly with age from 10 to 85 years, which could be accounted for by the acquisition of protective immunity by adolescents and adults following multiple exposures to malaria infection [40].
An estimation of malaria parasite density is necessary for patient management. Our study revealed that asymptomatic people coming from different villages in the Ngatti health district tolerated the high burden of asexual parasitaemia of up to 56,600 parasites/μl blood for P. falciparum without manifesting any sign of the disease; this burden could be more if we combine the parasitaemia in the case of co-infection. Plasmodium falciparum and P. malariae parasitaemia were significantly associated with age (P < 0.001) and, not surprisingly, the highest parasitaemia was recorded in children aged < 5 years. This result is explained by the low levels of immunity in young children and increasing naturally acquired immunity in older children through repeated exposure to the parasite [41, 42].
The prevalence of P. malariae obtained by microscopy was low, although higher than the general prevalence observed in Cameroon (3%) [5]. The highest prevalence was also observed in individuals between 11 and 20 years old. According to the “niche theory”, the elimination of P. falciparum in the lower age groups can be assumed to allow non-falciparum species, such as P. malariae, to become more important as P. falciparum is eliminated. In general, high transmission areas constitute a significant infectious reservoir of malaria parasites due to the potentially high percentage of asymptomatic malaria parasite carriers in this location [13, 14], which is what was observed in the Ngatti district health.
Conclusion
The aim of this study was to examine the prevalence of malaria infection, targeting both P. falciparum as well as P. malariae, in the asymptomatic population. The findings revealed that malaria endemicity in the Adamawa Region of Cameroon is driven by multiple Plasmodium species, although P. falciparum remains the most prevalent. In addition, P. malariae infection and P. falciparum/P. malariae co-infections are both transmitted to humans by An. funestus in the studied localities and play an important role in malaria transmission. We found that older children were the age group most infected with malaria. Thus, beyond the control of malaria targeting only the under-fives, pregnant women and P. falciparum, a comprehensive policy targeting all age groups of the population should be put in place to eliminate malaria and its reservoir species more effectively.
Availability of data and materials
All data are fully available without restriction.
Abbreviations
- ACT:
-
Artemisinin-based combination therapy
- IRS:
-
Indoor residual spraying
- ITNs:
-
Insecticide treated nets
- LLINs:
-
Long-lasting insecticide-treated bed nets
- P. OVM :
-
Plasmodium malariae, Plasmodium ovale OR Plasmodium vivax
- RDTs:
-
Rapid diagnostic tests
References
WHO. World malaria report 2015. https://apps.who.int/iris/handle/10665/200018. Accessed 15 Feb 2022.
Hemingway J, Shretta R, Wells TN, Bell D, Djimdé AA, Achee N, et al. Tools and strategies for malaria control and elimination: what do we need to achieve a grand convergence in malaria? PLoS Biol. 2016;14:e1002380.
Drakeley C, Akim N, Sauerwein R, Greenwood B, Targett G. Estimates of the infectious reservoir of Plasmodium falciparum malaria in The Gambia and in Tanzania. Trans R Soc Trop Med Hyg. 2000;94:472–6.
Ross A, Killeen G, Smith T. Relationships between host infectivity to mosquitoes and asexual parasite density in Plasmodium falciparum. Am J Trop Med Hyg. 2006;75(2 Suppl):32-7. https://doi.org/10.4269/ajtmh.2006.75.32.
Antonio-Nkondjio C, Ndo C, Njiokou F, Bigoga JD, Awono-Ambene P, Etang J, et al. Review of malaria situation in Cameroon: technical viewpoint on challenges and prospects for disease elimination. Parasit Vectors. 2019;12:1–23.
Antonio-Nkondjio C, Awono-Ambene H, Etang J, Fontenille D, Ndo C, Simard F, et al. Epidemiological importance of the Anopheles Nili group of malaria vectors in equatorial Villages of Cameroon; Central Africa. Bull liaison doc-OCEAC. 2009;1:13-20.
Wanji S, Tanke T, Atanga SN, Ajonina C, Nicholas T, Fontenille D. Anopheles species of the Mount Cameroon region: biting habits, feeding behaviour and entomological inoculation rates. Trop Med Int Health. 2003;8:643–9.
Fontenille D, Cohuet A, Awono-Ambene P, Antonio-Nkondjio C, Wondji C, Kengne P, et al. Systématique et biologie des anophèles vecteurs de Plasmodium en Afrique, données récentes. Med Trop. 2003;63:247–53.
Okell LC, Ghani AC, Lyons E, Drakeley CJ. Submicroscopic infection in Plasmodium falciparum-endemic populations: a systematic review and meta-analysis. J Infect Dis. 2009;200:1509–17.
Badiane AS, Diongue K, Diallo S, Ndongo AA, Diedhiou CK, Deme AB, et al. Acute kidney injury associated with Plasmodium malariae infection. Malar J. 2014;13:1–5.
Betson M, Clifford S, Stanton M, Kabatereine NB, Stothard JR. Emergence of nonfalciparum Plasmodium infection despite regular artemisinin combination therapy in an 18-month longitudinal study of Ugandan children and their mothers. J Infect Dis. 2018;217:1099–109.
Sutherland CJ. Persistent parasitism: the adaptive biology of malariae and ovale malaria. Trends Parasitol. 2016;32:808–19.
Black J, Hommel M, Snounou G, Pinder M. Mixed infections with Plasmodium falciparum and P malariae and fever in malaria. Lancet. 1994;343:1095.
Mason DP, McKenzie FE, Bossert WH. The blood-stage dynamics of mixed Plasmodium malariae–Plasmodium falciparum infections. J Theor Biol. 1999;198:549–66.
Nundu SS, Culleton R, Simpson SV, Arima H, Muyembe J-J, Mita T, et al. Malaria parasite species composition of Plasmodium infections among asymptomatic and symptomatic school-age children in rural and urban areas of Kinshasa. Malaria J. 2021;20:1–13.
Yman V, Wandell G, Mutemi DD, Miglar A, Asghar M, Hammar U, et al. Persistent transmission of Plasmodium malariae and Plasmodium ovale species in an area of declining Plasmodium falciparum transmission in eastern Tanzania. PLoS Negl Trop Dis. 2019;13:e0007414.
Mbenda Ngassa, Gaelle Huguette, Aparup D. Molecular evidence of Plasmodium vivax mono and mixed malaria parasite infections in Duffy-negative native Cameroonians. PLoS One. 2014;9:e103262. https://doi.org/10.1371/journal.pone.0103262.
Fru-Cho J, Bumah VV, Safeukui I, Nkuo-Akenji T, Titanji VP, Haldar K. Molecular typing reveals substantial Plasmodium vivax infection in asymptomatic adults in a rural area of Cameroon. Malar J. 2014;13:1–11.
Russo G, Faggioni G, Paganotti GM, Dongho GBD, Pomponi A, De Santis R, et al. Molecular evidence of Plasmodium vivax infection in Duffy negative symptomatic individuals from Dschang. Malaria J. 2017;16:1–9.
Kwenti TE, Nkume FA, Tanjeko AT, Kwenti TD. The effect of intestinal parasitic infection on the clinical outcome of malaria in coinfected children in Cameroon. PLoS Negl Trop Dis. 2016;10:e0004673.
Tchouakui M, Chiang M-C, Ndo C, Kuicheu CK, Amvongo-Adjia N, Wondji MJ, et al. A marker of glutathione S-transferase-mediated resistance to insecticides is associated with higher Plasmodium infection in the African malaria vector Anopheles funestus. Sci Rep. 2019;9:1–12.
Manly BF. The design and analysis of research studies. Cambridge: Cambridge University Press; 1992.
Bereczky S, Martensson A, Gil JP, Farnert A. Rapid DNA extraction from archive blood spots on filter paper for genotyping of Plasmodium falciparum. Am J Trop Med Hyg. 2005;72:249–51.
Snounou G, Viriyakosol S, Jarra W, Thaithong S, Brown KN. Identification of the four human malaria parasite species in field samples by the polymerase chain reaction and detection of a high prevalence of mixed infections. Mol Biochem Parasitol. 1993;58:283–92.
Gillies M, Coetzee M. A supplement to the Anophelinae of Africa South of the Sahara. Publ S Afr Inst Med Res. 1987;55:1–143.
Koekemoer L, Kamau L, Hunt R, Coetzee M. A cocktail polymerase chain reaction assay to identify members of the Anopheles funestus (Diptera: Culicidae) group. The American journal of tropical medicine and hygiene. 2002;66 6:804–11.
Livak KJ. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics. 1984;107 4:611–34.
Bass C, Nikou D, Blagborough AM, Vontas J, Sinden RE, Williamson MS, et al. PCR-based detection of Plasmodium in Anopheles mosquitoes: a comparison of a new high-throughput assay with existing methods. Malaria journal. 2008;7 1:1–9.
Tchouakui M, Riveron JM, Djonabaye D, Tchapga W, Irving H, Soh Takam P, et al. Fitness costs of the glutathione S-transferase epsilon 2 (L119F-GSTe2) mediated metabolic resistance to insecticides in the major African malaria vector Anopheles funestus. Genes. 2018;9:645.
Menze BD, Wondji MJ, Tchapga W, Tchoupo M, Riveron JM, Wondji CS. Bionomics and insecticides resistance profiling of malaria vectors at a selected site for experimental hut trials in central Cameroon. Malar J. 2018;17:1–10.
Feufack-Donfack LB, Sarah-Matio EM, Abate LM, Tuedom AGB, Bayibéki AN, Ngou CM, et al. Epidemiological and entomological studies of malaria transmission in Tibati, Adamawa region of Cameroon 6 years following the introduction of long-lasting insecticide nets. Parasit Vectors. 2021;14:1–14.
Hillyer JF, Barreau C, Vernick KD. Efficiency of salivary gland invasion by malaria sporozoites is controlled by rapid sporozoite destruction in the mosquito haemocoel. Int J Parasitol. 2007;37:673–81.
Pinilla YT, Boussougou-Sambe ST, Gräßle S, Ngossanga B, Doumba-Ndalembouly AG, Weierich A, et al. Experimental Transmission of Plasmodium malariae to Anopheles gambiae. J Infect Dis. 2021;223:522–6.
Dillon RJ, Dillon V. The gut bacteria of insects: nonpathogenic interactions. Annu Rev Entomol. 2004;49:71–92.
Zieler H, Dvorak JA. Invasion in vitro of mosquito midgut cells by the malaria parasite proceeds by a conserved mechanism and results in death of the invaded midgut cells. Proc Natl Acad Sci USA. 2000;97:11516–21.
Abdulraheem MA, Ernest M, Ugwuanyi I, Abkallo HM, Nishikawa S, Adeleke M, et al. High prevalence of Plasmodium malariae and Plasmodium ovale in co-infections with Plasmodium falciparum in asymptomatic malaria parasite carriers in southwestern Nigeria. Int J Parasitol. 2022;52:23–33.
Isozumi R, Fukui M, Kaneko A, Chan CW, Kawamoto F, Kimura M. Improved detection of malaria cases in island settings of Vanuatu and Kenya by PCR that targets the Plasmodium mitochondrial cytochrome c oxidase III (cox3) gene. Parasitol Int. 2015;64:304–8.
Apinjoh TO, Anchang-Kimbi JK, Mugri RN, Tangoh DA, Nyingchu RV, Chi HF, et al. The effect of insecticide treated nets (ITNs) on Plasmodium falciparum infection in rural and semi-urban communities in the South West Region of Cameroon. PLoS One. 2015;10:e0116300.
Njua-Yafi C, Achidi EA, Anchang-Kimbi JK, Apinjoh TO, Mugri RN, Chi HF, et al. Malaria, helminths, co-infection and anaemia in a cohort of children from Mutengene, south western Cameroon. Malar J. 2016;15:1–11.
Ndamukong-Nyanga JL, Guria PC, Ndamukong NC, Nji DN, Yunga NB. The influence of Helicobacter pylori infection on malaria parasitaemia among symptomatic patients in Buea Cameroon. Int J Trop Dis Health. 2018;31:1–7. https://doi.org/10.9734/IJTDH/2018/42812.
Kimbi HK, Keka F, Nyabeyeu H, Ajeagah H, Tonga C, Lum E, et al. An update of asymptomatic falciparum malaria in school children in Muea, Southwest Cameroon. J Bacteriol Parasitol. 2012;3:2.
Sumbele IU, Ning TR, Bopda OS, Nkuo-Akenji T. Variation in malariometric and red cell indices in children in the Mount Cameroon area following enhanced malaria control measures: evidence from a repeated cross-sectional study. Malar J. 2014;13:1–10.
Acknowledgements
We thanks the local residents of Mibellon, Moinkoing, Ngatti and Mbougam villages for their participation in this study. We are also grateful to Yvan Fotso Toguem, who drew the map of study area, and Amelie Wamba and Jonas Kengne, who also read the manuscript.
COMAL study group
Romuald Agonhossou, Romaric Bidossessi Akoton, Barclaye Ngossanga1, Ange G Doumba-Ndalembouly1, Abel Lissom, Jacques Dollon Mbama Ntabi and Sara Buezo-Montero.
Funding
This work received financial support of the German Research funding [DFG BO 2494/3-1] and Wellcome Trust Senior Research Fellowship in Biomedical Sciences to Charles S. Wondji (217188/Z/19/Z).
Author information
Authors and Affiliations
Contributions
DNN performed the parasitological survey, laboratory work, slide reading, data analysis and writing of the first draft. NF performed the parasitological survey and laboratory work. CD performed the parasitological survey, commented and approved the manuscript. J-PA performed the parasitological survey and slide reading. STBS commented on and approved the manuscript. SB contributed to the design of the study, and commented on and approved the manuscript. AAA contributed to the design of the study, and commented on and approved the manuscript. FN contributed to the design of the study, and commented on and approved the manuscript. LD contributed to the design of the study, and commented on and approved the manuscript. CSW, contributed to the design of the study, coordinated the field and laboratory work and commented on and approved the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was approved by the national research ethics committee for human health No. 1234 CE/CNERSH/SP. Before enrolment, informed consent was obtained from participants or parents on behalf of their children and was documented.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Nguiffo-Nguete, D., Nongley Nkemngo, F., Ndo, C. et al. Plasmodium malariae contributes to high levels of malaria transmission in a forest–savannah transition area in Cameroon. Parasites Vectors 16, 31 (2023). https://doi.org/10.1186/s13071-022-05635-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05635-7