
Abstract
Background
Onchocerciasis control using ivermectin alone has been achieved in some endemic savannah zones of Africa. In the forest regions, the co-endemicity with Loa loa has led to severe adverse events (SAEs) resulting in poor adherence of community members to ivermectin mass drug administration (MDA). This may jeopardize achieving the interruption of transmission of onchocerciasis. Therefore, to accelerate the elimination of onchocerciasis in L. loa co-endemic zones, alternative treatment strategies (ATS) including ground larviciding may be necessary. This study aimed at identifying Simulium breeding sites, cytospecies, transmission profile, susceptibility of Simulium larvae to insecticide (temephos) and identification of some non-target aquatic fauna prior to the implementation of the COUNTDOWN consortium ground larviciding alternative strategy in the Meme River Basin in South West Cameroon.
Methods
A topographic map and entomological survey were used to determine breeding sites. Larvae and adults were identified using standard identification keys. Susceptibility tests were carried out on collected larvae by exposing them to decreasing concentrations of temephos and assessing survival rates while the cytospecies were identified using cytotaxonomy. Various entomological indicators were assessed from dissected flies. Fishing was used as proxy to traps to assess some aquatic fauna at different sites.
Results
Twenty-two breeding sites were prospected in the Meme River Basin with eight productive for larvae. A concentration of 0.5–0.1 mg/l temephos induced 100% larval mortality. As the concentration of temephos decreased from 0.05 to 0.0025 mg/l, mortality of larvae also decreased from 98.7 to 12%. Nine cytospecies were observed in the Meme River Basin; 13,633 flies were collected and 4033 dissected. A total of 1455 flies were parous (36.1%), 224 flies were infected (5.5%), and 64 were infective (1.6%). Aquatic fauna observed included Cyprinus spp., Clarias spp., crabs, tadpoles, beetles and larvae of damsel fly.
Conclusions
Onchocerciasis is being actively transmitted within the Meme River Basin. Simulium larvae are susceptible to temephos, and nine cytospecies are present. Non-target fauna observed included fishes, frogs, crabs and insects. Besides treatment with ivermectin, vector control through ground larviciding may be a complementary strategy to accelerate onchocerciasis elimination in the study area.
Graphical Abstract

Similar content being viewed by others


Background
Onchocerciasis, a parasitic disease caused by the filarial nematode Onchocerca volvulus [1], is a public health problem found in some of the world’s most disadvantaged communities in remote, rural sub-Saharan Africa. Adult worms live in subcutaneous nodules and produce millions of microfilariae during their reproductive life span (10–15 years). The microfilariae (mf) can live between 1 and 2 years in the skin, from where they are taken up by black fly vectors during a blood meal. Mf develops into infective larvae within black fly, which infect another person during a subsequent bite, thus maintaining the transmission cycle. Host inflammatory reactions to dead mf in the skin cause onchocercal dermatological disease and in the eyes ocular disease that may end in blindness. Because of the public health problem and socioeconomic effects of onchocerciasis, large-scale control programmes were created in the past with strategies for its control while in recent years strategies have been directed at its elimination [2]. In 11 West African countries, the Onchocerciasis Control Programme (OCP) aerial larviciding led to the control of onchocerciasis as a public health problem [3].
The advent of ivermectin and its donation by Merck & Co. (NJ, USA) [4] provided the opportunity for its large-scale use for the treatment of onchocerciasis by the African Programme for Onchocerciasis Control (APOC) in 20 endemic African countries. Ivermectin mass drug administration (MDA) has led to the interruption of transmission in some foci in Mali, Senegal, Sudan and Uganda [5,6,7,8]. Unfortunately, the same level of achievement has not been made in Cameroon, especially in the South West Region where onchocerciasis is co-endemic with loiasis.
In the South West Region of Cameroon, the persistence of onchocerciasis transmission is due to the existence of abundant breeding sites in fast-flowing rivers, which results in very high numbers of biting black flies transmitting the parasite and poor adherence of the community members to ivermectin mass drug administration [9, 10]. The poor adherence to ivermectin MDA is due to the fear of SAEs that follow ivermectin treatment of highly infected Loa loa individuals in co-endemic areas of onchocerciasis and L. loa [10, 11]. Faced with these challenges and coupled with the change in paradigm from onchocerciasis control to elimination, it was important for APOC to offer new approaches to improve or complement ivermectin MDA [12]. This led to adoption of alternative treatment strategies (ATS). These ATS include complementary vector control, enhanced community-directed treatment with ivermectin (CDTI), community-directed treatment (CDT) with drug combinations or new drugs and test-and-treat (TNT) strategies [13]. The ATS are essentially directed towards highly endemic onchocerciasis foci where the disease cannot be eliminated by annual CDTI alone.
The Meme River Basin of South West Cameroon has been under CDTI for > 18 years. A situational analysis of onchocerciasis conducted in this region by Wanji et al. [9] demonstrated persistent transmission despite ivermectin MDA with prevalence values as high as 52% [9]. This persistence in onchocerciasis transmission motivated the COUNTDOWN Consortium to evaluate implementation of one of the recommended ATS, ground larviciding, as a potential tool to accelerate the elimination of onchocerciasis in this endemic focus. Ground larviciding is known to significantly reduce the number of biting black flies, thus reducing disease transmission and infection rates. Pre-requisites to ground larviciding are: identification of breeding sites, susceptibility test of Simulium larvae to a larvicide of interest, identification of cytospecies in the river basin and identification of non-target aquatic fauna. In this study, we chose to use temephos (Abate® 500 EC-BASF, Douala, Cameroon) as the larvicide. Temephos is an organophosphate larvicide which has been shown to be biodegradable and non-toxic to non-target fauna with a good carry during the rainy season. This article presents the breeding sites, identified Simulium cytospecies, current onchocerciasis transmission profile, susceptibility of Simulium damnosum larvae to temephos and a baseline survey of some non-target aquatic fauna present in the Meme River Basin prior to the implementation of the COUNTDOWN Consortium ground larviciding alternative strategy implementation trial.
Methods
Study area
Meme River Basin is situated in the rain forest of the South West Region of Cameroon approximately 60 km inland from the Atlantic Ocean. The topography is very diverse; the main feature is a mountain range (Rumpi Hills, characterized by a volcanic ridge culminating at 1764 m with a northeast orientation [14]. The volcanic ridge is broken by several valleys and constitutes a watershed that is the source for several rivers, the Manyu, Meme, Mungo and Ndian. These rivers go down steep slopes generating fast currents and appropriate flow rates that favour the establishment of permanent Simulium breeding sites conducive to high intensity of onchocerciasis transmission. The climate is characterized by 8 months of rainfall and a short dry season from December to March. The annual rainfall varies between 2500 and 4000 mm with annual temperatures ranging from 25 to 32 °C.
River Meme and its tributaries originate from the Rumpi Hills and flow down precipitous slopes some kilometres from the selected communities. The main tributaries of the Meme River are the Mbile River, which flows pass Marumba I and II, and the Uve River in Bakumba and Big Massaka. These tributaries create important Simulium breeding sites as they enter the Meme River.
Identification of breeding sites
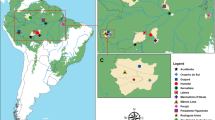
Potential Simulium breeding sites in the Meme River were selected using a topographical map of 1:200,000 scale [28] followed by an entomological survey to check for the presence of rapids, trailing vegetation and larvae to confirm that it is a breeding site. The Melange River in Ndian Division was set as the control area, which will be used to monitor the stability of entomological indicators after ground larviciding. The larvae and adults were identified using standard identification keys [15, 16]. All breeding sites were geo-referenced using a GPS. The co-ordinates were used to generate a topographical map of the breeding sites (Fig. 1a, b).

Map showing breeding sites geo-referenced using a GPS. a Map of the study and control area with prospected breeding sites. b Map showing productive and non-productive breeding sites of the study area
Study sites
The study was conducted from March to July 2017. River Meme and its tributaries were surveyed, 20 breeding sites from 12 communities (Betenge, Small Massaka Lokando Big Butu, Small Butu, Big Massaka, Kumukumu, Bombele, Bobanda Kwakwa Nake and Marumba 1) were visited, but due to absence of larvae in some sites at the time of the study, only 8 sites from six communities (Betenge, Small Massaka, Big Massaka, Bombele, Kwakwa and Marumba 1) were chosen for fly and larvae collection. Due to accessibility, two sites near one community (Berenge) were surveyed in the Melange River Basin (control area). Flies were collected and dissected from one of the sites that had larvae to monitor the stability of entomological indicators in an untreated area after ground larviciding. Some sites in the Meme River Basin did not have enough larvae, so larvae for cytotaxonomy were collected only from three sites (Big Massaka, Bombele and Kwakwa). In the study arm, the breeding sites chosen were at:
-
Marumba I (River Mbile, a tributary of Meme). Here, S. damnosum larvae were collected and a fly dissection point was set up at the river bank.
-
Bombele, where the Meme River runs over several rock features developing rapids, which constitute breeding sites along its way to join the Atlantic Ocean.
-
Kwakwa (along River Mbile, a tributary of Meme). Here, many larvae were found and a fly collection point was set up at the bank of the river.
-
Big Massaka (River Uve, tributary of Meme). A survey of this tributary revealed many breeding sites which contained S. damnosum larvae. A catching point was therefore located in this community.
-
Betenge, where the Meme runs. A fly collection and dissection point were located in this community
-
Small Massaka (River Ka, tributary of Meme). A fly collection and dissection point were located in this community
Susceptibility of S. damnosum larvae to temephos (Abate 500EC)
Larvae of S. damnosum were collected from several breeding sites, namely Marumba 1, Big Massaka, Bombele and KwaKwa in the Meme River Basin located in the South West Region of Cameroon. These larvae were exposed using a bowl containing local bottled water (Supermont) to ten concentrations of temephos (0.0005 mg/l to 0.5 mg/l) (Additional file 1: Text S1) for a period of 3 h [17]. After this, mortality was determined by observation of larvae motility response to stimulation [18].
Dosage-mortality regression line
Adjusted mortality for each concentration was calculated utilising the control mortality frequency that ran in parallel with the test. If the control mortality was between 5 and 20%, then Abbot’s formula [19] was used to correct the mortality and ensure that other factors were not contributing to the mortality of the larvae. No adjustment was done if control mortality was < 5% and results were discarded for any testing with control mortality > 20%.
Abbot’s formula
Temephos dosage-dependent mortality curve was constructed using prism 7.0 (Graphpad, San Diego, CA, USA).
Identification of cytospecies of S. damnosum in the Meme River Basin
Cytotaxonomy was used to identify the different cytospecies in the Meme River Basin. Cytotaxonomy requires the extraction and analysis of polythene chromosomes (giant chromosomes caused by cells that have undergone repeated rounds of replication) present in the silk gland of black fly larvae. Larvae for cytotaxonomy were collected from three breeding sites (Big Massaka, Bombele and Kwakwa) because the other breeding site did not have enough larvae for cytotaxonomy. The sixth and seventh stage larvae were collected and put in Carnoy’s fixative (3 parts ethanol: 1part acetic acid) that was freshly prepared at the riverside. A minimum of 30 larvae were put in 24 ml of Carnoy. The Carnoy’s solution was changed after 3 h and 24 h. The silk glands were dissected followed by hydrolysation to enable all the silk in the glands to be removed. Thereafter, chromosomes were stained for 5 min with Feulgen stain, which stains the DNA pink. Continual addition of orcien to the silk gland occurred to prevent dehydration. The gonads of the larvae were inspected to identify the sex. If male, the gonads are round, but if female, the gonads are elongated in shape. Squashing was done to separate the chromosomes while a light microscope was used to visualize and identify them. Photographs were taken at × 400. Identification of each larval chromosome was done using information from pictorial guide and identification key by Post et al. [20].
Collection and identification of Simulium flies
Simulium flies were collected from seven different sites: Bombele and Betenge (River Meme), Big Massaka (River Uve), Kwakwa and Marumba I (River Mbile), Small Massaka (River Ka) and Berenge (River Melange). Residents of endemic communities for onchocerciasis were trained to carry out collections; Simulium flies landing on exposed legs for a blood meal were captured using suction tubes or mouth aspirators (locally adapted) before they bit [21].
Adult flies were either dissected to determine the parity and infectivity rate or stored in 80% alcohol for future use. Onchocerca volvulus transmission indicators (parous rate, infection rate, infectivity rate and monthly transmission potential) were computed after fly collection and dissection as described by Walsh et al. [22].
Dissection of Simulium flies
Captured flies were killed using chloroform, counted and dissected in physiological saline under a dissecting microscope (Humanscope, HUMAN Diagnostics Worldwide, USA). Flies caught were dissected on an hourly basis to determine parity and infection status. This consisted of holding the fly with a needle in the thorax, piercing the abdomen with a dissecting needle at the posterior end and then pulling out the different internal organs to examine the fat bodies, state of the Malpighian tubules and ovaries to distinguish parous from nulliparous flies (nulliparous ovary is generally clear with follicles that are bigger and have voluminous fatty substance while parous flies generally have yellowish, spotted ovaries because of follicular relicts. The follicles are much smaller with little or no fatty substance). The head, thorax and abdomen of parous flies were further dissected separately and examined for O. volvulus developing larvae (L1, L2 and L3). Any larvae found were counted and recorded on a dissection sheet [23] and the entomological indices computed [22]
Survey of some non-target aquatic fauna
Survey of some non-target aquatic fauna was conducted to obtain a picture mainly of the common fish species and also other common vertebrate as well as invertebrate non-target fauna to serve as indicators to guide measurement of any detrimental effect of temephos during ground larviciding. For this to be done, fishermen and -women were contacted to determine the species of aquatic vertebrates common in the Meme River and its tributaries. Thereafter, they were requested to carry out fishing exercises to determine the vertebrate aquatic fauna found at the different study sites. Locally made fishing nets were placed in water overnight to trap any available vertebrate aquatic fauna as modified from the protocol adopted by OCP in 1991 [24]. During the collection of Simulium larvae, invertebrate aquatic fauna present on the same substrate as the Simulium larvae were observed. Any aquatic fauna observed were grouped as either vertebrate or invertebrate. The vertebrate aquatic fauna were identified with reference to Temegne and Momo [25] while the invertebrate aquatic fauna were identified using a key by Danladi et al. [26].
Data analysis
Fly collection data were entered in a template created in Epi info version 3.5.3 (https://www.cdc.gov/epiinfo/index.html), exported to Microsoft Excel and then later to SPSS version 24 for analysis. The different entomological indicators generated were biting densities, daily biting rate, monthly biting rate, parous rates infection rates, infective rates and monthly transmission potential. The chi-square test for trend was used to establish whether there was any association among parous rate, infection rate and infective rate.
Entomological indices were calculated using the following formulae:
-
Parous rates were computed as the number of parous flies divided by the total number of flies dissected and multiplied by 100.
-
Infection rates were computed as number of flies carrying any developmental form of O. volvulus larvae divided by the total number of parous flies dissected and multiplied by 100.
-
Infective rates were computed as the total number of flies carrying infective larvae in the head divided by the total number of parous flies dissected and multiplied by 100.
-
Biting density (number of flies collected at different hours plotted against the hours collected).
-
Monthly transmission potential: (number of days in month × number of infective larvae)/number of fly collection days × (number of flies collected/number of flies dissected).
Results
Identification of breeding sites
Twenty-two breeding sites from 13 communities (Berenge, Betenge, Small massaka, Lokando, Big butu, Small butu, Big massaka, Kumukumu, Bombele, Bobanda, Kwakwa, Nake and Marumba 1) in both the control and study arms were prospected based on accessibility (Fig. 1a, b). Nine of these sites from seven communities (Berenge, Betenge, Small Massaka, Big Massaka, Bombele, Kwakwa and Marumba 1) had larvae (productive) at the time of prospection.
Breeding sites in Bombele and Betenge (River Meme), Big Massaka (River Uve), Kwakwa and Marumba I (River Mbile) and Small Massaka (River Ka) were identified as the main breeding sites of the study area; as such, fly and larvae collection points were set up at these sites. In the case of larval density, substrates were not calibrated but on average about ten larvae were found per substrate. Simulium damnosum was found to be the only anthropophagic species in the study area.
Susceptibility test results
At a concentration between 0.5 and 0.1 mg/l, a mortality of 100% was recorded. It was observed that as the concentration of temephos decreased (from 0.05 to 0.00025 mg/l), mortality of larvae also decreased uniformly (from 12 to 98.7%) as seen in Table 1. Thus, S. damnosum larvae in these areas demonstrated good susceptibility to temephos (Abate 500EC).
Dosage-mortality regression curve was plotted for susceptibility testing (Fig. 2). The control mortality was set to a concentration of 0.0001 mg/l as taking a logarithm of 0 is undefinable. This shows that the concentration of temephos required to kill 50% of larvae (EC50) is 0.005 mg/l and the concentration required to kill 95% of larvae (EC95) is 0.041 mg/l. The Hill slope (steepness of curve) was not constrained and included the control mortality.

Dosage-mortality regression curve of concentrations. The control mortality was set to a concentration of 0.0001 mg/l as taking a logarithm of 0 is undefinable. The Hill slope (steepness of curve) was not constrained and included the control mortality
Identification of cytospecies in the Meme River Basin
A total of 328 larvae were collected for cytotaxonomy with 40 from Bombele, 241 from Big Massaka and 47 from Kwakwa. Sixty larvae chosen for chromosome preparation due to their large size and white histoblast. This included 13 from Bombele, 29 from Big Massaka and 18 from Kwakwa. Chromosome preparations of 60 larvae were examined to identify high-quality images where the banding patterns on the chromosome were distinctive and clear. Additionally, some preparations were removed if the chromosomes were tightly coiled or broken. This led to the cytotaxonomic analysis of 15 larvae; 9 S. squamosum subcomplex larvae were found, which were three times the larvae found for any other subcomplex (Table 2). Four larvae from the S. squamosum subcomplex could not be identified as either male or female. The cytospecies S. soubrense (Beffa form) was observed but has not been previously described in Cameroon (Fig. 3, in blue).

Observed cytospecies of S. damnosum in the Meme River Basin. The cytospecies S. soubrense (Beffa form) was observed but has not been previously described in Cameroon (in blue)
Entomological indicators of O. volvulus transmission
Entomological indices of O. volvulus transmission in the study site are found in Table 3. A total of 13,633 flies were collected and 4033 were dissected. Of the flies dissected, 1455 were parous, giving a parous rate of 36.1%; 224 were infected (infection rate 5.5%) and 64 were infective giving an infective rate of 1.6%. Kwakwa had the highest parous rate (43.7%) while Marumba I recorded the lowest parous rate (17.8%). On the other hand, Bombele had the highest infection rate (10%) while Betenge had the lowest infection rate (1.4%).
Biting density
More flies were caught during the afternoon hours with the peak biting hours for most of the communities between 1 and 3 p.m. This coincides with the peak farming hours. The most flies per hour were caught at Bombele with 1177 flies caught between 2 and 3 p.m. Biting density decrease towards the evening period (Fig. 4).

Biting density at different study sites. Peak biting hours for most of the communities were between 1–3 p.m. with as many as 1177 flies caught per hour at Bombele
Daily biting rate (DBR) and monthly biting rate (MBR)
The DBR and MBR for the various collection sites are shown in Table 4. The highest daily and monthly biting rates were recorded at Kwakwa (1311 and 40618 respectively) while the lowest daily and monthly biting rate was recorded at Betenge (142 and 4402 respectively).
Trend in entomological indicators within the study sites.
Figure 5 shows that there was some trend in parasitological indicators. Bombele with the highest parous rate also recorded the highest infection rates and eventually one of the highest infective rates. On the other hand, Betenge had a very high parous rate but recorded the lowest infection rate. As such, a chi-square test of trend indicated that there was no statistically significant association (Chi-square test, x2 = 2.332, df = 1, P-value = 0.127) among parous rate, infection rate and infective rate.

Variation in entomological indicators across study sites. A chi-square test of trend indicates that there was no statistically significant association (Chi-square test, x2 = 2.332, df = 1, P = 0.127) among parous rate, infection rate and infective rate
Survey of some non-target aquatic fauna
The aquatic fauna found in the Meme River Basin were the same in all the study sites at the time of survey. This included two species of fish: Cyprinus (Mbanga fish) and Clarias sp. (African catfish). Other aquatic fauna observed were crabs and tadpoles. The presence of tadpoles indicated that frog species were also present though no frogs were captured. During the collection of S. damnosum larvae, aquatic Coleoptera (beetle) and the larvae of damsel flies (Odonata: Zygoptera) were also observed on the same substrate.
Discussion
This study was designed to assess entomological indicators of onchocerciasis transmission and the susceptibility of Simulium larvae to temephos prior to the implementation of ground larviciding as an alternative treatment strategy for the acceleration of onchocerciasis elimination in the Meme River Basin. To realise this, we mapped the Meme River system, conducted a survey of the aquatic fauna, tested the susceptibility of S. damnosum larvae to temephos (Abate 500EC) and dissected a sample of adult black fly to determine entomological parameters.
In this study, exposure of black fly larvae to decreasing concentrations of larvicide (from 0.5 to 0.0005 mg/l) resulted in decreasing mortality of S. damnosum larvae. One hundred percent mortality was achieved at concentration exposures of between 0.5 and 0.1 mg/l and we determined EC50 of 0.005 mg/l. We therefore conclude that Simulium larvae are susceptible to ABATE 500 EC and this insecticide is effective for use in ground larviciding to accelerate the elimination of onchocerciasis in the study area. This is in line with the study carried out by Kalinga et al. [27] where there was 100% mortality of Simulium larvae to temephos between 0.129 and 0.34 mg/l pre- and 0.144–0.211 mg/l (95% CI, P < 0.05) post-treatment. The concentrations that were lethal to S. damnosum larvae in this study fall within the standard diagnostic dose of ≤ 0.4 mg/l for susceptible S. damnosum (s.l.) populations. This also shows the rapid activity of temephos, which has been shown to be highly efficacious against S. damnosum (s.l.) larvae at concentrations giving 0.05 mg/l at high river discharges (> 25 cubic m/s) and at 0.1 mg/l at low discharges (when insecticide carry is reduced). This is an indication that larvae from the Meme River Basin are highly susceptible to temephos.
Previously within Cameroon, there were eight cytospecies of S. damnosum recorded in the literature [20]. This record gives the most cytospecies that have been recorded in one country in West Africa [28]. This could be because Cameroon is situated in the Gulf of Guinea, which is known for its high biodiversity [29] with many different cytospecies occurring in proximity to different subcomplexes. In this study, the cytospecies S. soubrese (Beffa form) was found in two breeding sites (Kwakwa and Big Massaka). Notably, there have been no published studies that have recorded this cytospecies in Cameroon. However, this cytospecies has been recorded in Nigeria, which is a neighbouring country to Cameroon precisely in the Jos Plateau, which is 500 km from the Meme River Basin, and the Guarara River, which is 600 km away from the Meme River Basin [20]. The addition of S. soubrense (Beffa form) gives a record of nine cytospecies of S. damnosum found in Cameroon (Fig. 3).
There has been a decrease in infected flies from 12.9% in 2015 by Wanji et al. [9] to 5.5%. There has also been a decrease in infectivity from 3.9% in 2015 [9] to 1.6% obtained by this study. Results from this study show that onchocerciasis is still being actively transmitted in the Meme River Basin despite > 18 rounds of mass drug administration.
The availability of fast-flowing rivers in this basin contributes to the large numbers of black flies necessary for high transmission. In this pre-intervention baseline study, biting density was up to 36 bites/person/day, highlighting the need for a focal vector control [30].
In areas of high pre-intervention baseline infection endemicity, it has been suggested that vector control could be used as a complementary intervention strategy [31]. Modelling results using EPIONCHO and ONCHOSIM [32] suggest that in such settings vector control could be used in conjunction with ivermectin MDA to enhance effectiveness or to consolidate the gains made towards the path of onchocerciasis elimination. The aquatic fauna found in the Meme River Basin were similar in all the study sites. Both vertebrate aquatic fauna and invertebrate aquatic fauna were observed. It is expected that this aquatic fauna will not be affected since temephos is relatively non-toxic to fish and other aquatic vertebrates [33].
This study is limited in that it failed to collect quantitative pre-control data for non-target fauna, which would have been compared with post-quantitative data for the sake of environmental impact assessment.
Conclusions
Onchocerciasis is being actively transmitted within the Meme River Basin. Simulium damnosum larvae in this river basin are susceptible to temephos and there are nine cytospecies of S. damnosum complex within the basin. Non-target fauna observed included fishes, frogs, crabs, and insects. Beside treatment with ivermectin, vector control through ground larviciding may complement and accelerate onchocerciasis elimination in the study area.
Availability of data and materials
All data analysed during this study are included within the paper and supplementary information file.
Abbreviations
- APOC:
-
African Programme for Onchocerciasis Control
- ATS:
-
Alternative treatment strategies
- CDT:
-
Community directed treatment
- CDTI:
-
Community directed treatment with ivermectin
- COUNTDOWN:
-
Calling time on neglected tropical diseases
- DBR:
-
Daily biting rate
- DNA:
-
Deoxyribonucleic acid
- EC:
-
Emulsifiable concentrate
- MBR:
-
Monthly biting rate
- MDA:
-
Mass drug administration
- Mg/l:
-
Milligram per litre
- OCP:
-
Onchocerciasis control programme
- ONCHOSIM:
-
Onchocerciasis transmission elimination simulation model
- SAEs:
-
Severe adverse effects
References
Duke BO, Lewis DJ, Moore PJ. Onchocerca-Simulium complexes. Transmission of forest and Sudan-savanna strains of Onchocerca volvulus, from Cameroon, by Simulium damnosum from various West African bioclimatic zones. Ann Trop Med Parasitol. 1966;60:318–26.
Remme J, Boatin B, Boussinesq M. Helminthic diseases: onchocerciasis and loiasis. In: Quah SR, Cockerham WC, editors. The international encyclopedia of public health. 2nd ed. Oxford: Elsevier; 2017. p. 576–87.
Boatin B. The Onchocerciasis control programme in West Africa (OCP). Ann Trop Med Parasitol. 2008;102:13–7.
Thylefors B. The Mectizan donation program (MDP). Ann Trop Med Parasitol. 2008;102:39–44.
Diawara L, Traore MO, Badji A, Bissan Y, Doumbia K, Goita SF, et al. Feasibility of onchocerciasis elimination with ivermectin treatment in endemic foci in Africa: first evidence from studies in Mali and Senegal. PLoS Negl Trop Dis. 2009;3:e497.
Higazi TB, Zarroug IM, Mohamed HA, Elmubark WA, Deran TC, Aziz N, et al. Interruption of Onchocerca volvulus transmission in the Abu Hamed focus. Sudan Am J Trop Med Hyg. 2013;89:51–7.
Katabarwa M, Richards F. Twice-yearly ivermectin for onchocerciasis: the time is now. Lancet Infec Dis. 2014;14:373–4.
Traore S, Wilson MD, Sima A, Barro T, Diallo A, Ake A, et al. The elimination of the onchocerciasis vector from the island of Bioko as a result of larviciding by the WHO African Programme for Onchocerciasis Control. Acta Trop. 2009;111:211–8.
Wanji S, Kengne-Ouafo JA, Esum ME, Chounna PW, Tendongfor N, Adzemye BF, et al. Situation analysis of parasitological and entomological indices of onchocerciasis transmission in three drainage basins of the rain forest of South West Cameroon after a decade of ivermectin treatment. Parasit Vectors. 2015;8:202.
Forrer A, Wanji S, Obie ED, Nji TM, Hamill L, Ozano K, et al. Why onchocerciasis transmission persists after 15 annual ivermectin mass drug administrations in South-West Cameroon. BMJ Glob Health. 2021;6:e003248.
Wanji S, Kengne-Ouafo JA, Esum ME, Chounna PW, Adzemye BF, Eyong JE, et al. Relationship between oral declaration on adherence to ivermectin treatment and parasitological indicators of onchocerciasis in an area of persistent transmission despite a decade of mass drug administration in Cameroon. Parasit Vectors. 2015;8:667.
WHO/APOC: African Programme for Onchocerciasis Control. Strategic options and alternative treatment strategies for accelerating onchocerciasis elimination in Africa. http://who.int/apoc/ATS Report_2015.12.pdf. Accessed on 26 Jan, 2021.
Boussinesq M, Fobi G, Kuesel AC. Alternative treatment strategies to accelerate the elimination of onchocerciasis. Int Health. 2018;10:i40–8.
Hawkins P, Brunt M, Hawkins P, Brunt M. The soils and ecology of West Cameroon report to the government of cameroon FAO expanded program of technical Assistance No 2083 Rome. Food Agric Organizat. 1965;76:1–3.
Crosskey RW. The identification of the larvae of African Simulium. Bull World Health Organ. 1962;27:483–9.
Dang PT, Peterson BV. Pictorial keys to the main species and species groups within the Simulium damnosum Theobald complex occurring in West Africa (Diptera: Simuliidae). Tropenmed Parasitol. 1980;31:117–20.
Mouchet J, Guillet P. Insecticide resistance in blackfly of the Simulium damnosum complex in West Africa. In: Research on insecticides for the control of onchocerciasis vectors. Research on Insecticides for the Control of Onchocerciasis Vectors. OCP/SAP Working Group, Geneva 1978.
Davies JB, Crosskey RW: World Health Organization. Division of Control of Tropical Diseases. Simulium - vectors of onchocerciasis. World Health Organization 1991; https://apps.who.int/iris/handle/10665/59007.
WHO. pesticide evaluation scheme, control of neglected tropical diseases. guidelines for efficacy testing of insecticides for indoor and outdoor ground-applied space spray applications 2009; who/htm/ntd/whopes.
Post RJ, Onyenwe E, Somiari SA, Mafuyai HB, Crainey JL, Ubachukwu PO. A guide to the Simulium damnosum complex (Diptera: Simuliidae) in Nigeria, with a cytotaxonomic key for the identification of the sibling species. Ann Trop Med Parasitol. 2011;105:277–97.
Wanji S, Tendongfor N, Esum M, Ndindeng S, Enyong P. Epidemiology of concomitant infections due to Loa loa, Mansonella perstans, and Onchocerca volvulus in rain forest villages of Cameroon. Med Microbiol Immunol. 2003;192:15–21.
Walsh JF, Davies JB, Le Berre R, Grams R. Standardization of criteria for assessing the effect of Simulium control in onchocerciasis control programmes. Trans R Soc Trop Med Hyg. 1978;72:675–6.
Duke BO. The differential dispersal of nulliparous and parous Simulium damnosum. Tropenmed Parasitol. 1975;26:88–97.
WHO. Report of the twelfth session of the ecological group Bouake, 1991. Expert advisory committee Twelfth session Ouagadougou.
Temegne N, Momo N. Review on aquaculture research in cameroon: fish farming. Int J Oceanogr aquac. 2019;3:000170.
Danladi MU, Jon SH, Michael JW. Freshwater Invertebrates of the Mambilla Plateau Nigeria. Photographic School of Biological Sciences University of Canterbury 2013.
Kalinga AK, Mweya CN, Barro T, Maegga BT. Susceptibility of Simulium damnosum complex larvae to temephos in the Tukuyu onchocerciasis focus, Southwest Tanzania. Tanzan Health Res Bull. 2007;9:19–24.
Post RJ, Mustapha M, Krueger A. Taxonomy and inventory of the cytospecies and cytotypes of the Simulium damnosum complex (Diptera: Simuliidae) in relation to onchocerciasis. Trop Med Int Health. 2007;12:1342–53.
Mustapha M, Post RJ, Enyong P, Lines J. A new cytotype of Simulium squamosum (Diptera: Simuliidae) from southwest Cameroon. Med Vet Entomol. 2004;98:18296–300.
Wanji S, Nji TM, Hamill L, Dean L, Ozano K, Njouendou AJ, et al. Implementation of test-and-treat with doxycycline and temephos ground larviciding as alternative strategies for accelerating onchocerciasis elimination in an area of loiasis co-endemicity: the COUNTDOWN consortium multi-disciplinary study protocol. Parasit Vectors. 2019;12:574.
WHO/APOC. Report of the consultative meetings on strategic options and alternative treatment strategies for accelerating onchocerciasis elimination in Africa. Ouagadougou African Programme for Onchocerciasis Control. 2015; http://www.who.int/apoc/ATS_Report_2015.12.pd. Accessed on 26 Jan. 2021.
Verver S, Walker M, Kim YE, Fobi G, Tekle AH, Zoure HGM, et al. How can onchocerciasis elimination in africa be accelerated? modeling the impact of increased ivermectin treatment frequency and complementary vector control. Clin Infect Dis. 2018;66:S267–74.
WHO. report of the meeting on ground larviciding of blackflies during the post-OCP period Expert Advisory Committee Twenty-second session Ouagadougou. 2001https://apps.who.int/iris/bitstream/handle/10665/311937/OCP-EAC22eng. Accessed 26 Jan 2021.
Acknowledgements
The authors are grateful to the expert group invited by the COUNTDOWN consortium for their advice in designing the study protocol. Thanks to all those who helped in the execution of the study, especially the fly collectors and Mr Oben Bruno, who was involved in collecting larvae from the river, to the Ministry of Public Health Cameroon (Division of Operational Research) and to the National Onchocerciasis Control Programme. Thanks also to the South West Regional Delegation of Public Health for their collaboration in the implementation of the study and to the Research and Evidence Division (RED) of the Department for International Development (DFID) for funding this study.
Funding
Funding for this study was obtained from the Research and Evidence Division (RED) of the Department for International Development (DFID) MG8-RBPS107080. The funds were used for data collection, analysis and interpretation of results.
Author information
Authors and Affiliations
Contributions
Conceptualization: SW. Data curation: RAA, RE, AJN, AA, EDO, JFC MEE, SW. Data analysis: RAA, RE AJN, SW. Field investigations: PE, LH, RE, ST, AA, BLN, RAA, DAN. Supervision of activities: SW, JT, MT. Methodology: SW, PE, MT, JT, RE, RAA, LH. Resources: SW, MT. Laboratory investigation: LH, ST, RE, PE. Wrote the draft manuscript: RE, PE, RAA, AJN, SW. Review and editing of manuscript: RE, EDO, PE, LH, ST, RAA, AJN, AA, MEE, JFC, DAN, BLN, MT, JT, SW. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was incorporated into the study research protocol 16–027 by COUNTDOWN consortium who were granted ethical approval by Liverpool School of Tropical Medicine (LSTM), the University of Buea and Cameroon National Ethics Committee. Before collectors were trained for fly collection, the nature and objectives of the study were explained to them and informed consent was obtained from all collectors. Fly collectors were given a dose of ivermectin as prophylactic treatment against filariasis before fly collection and 3 months after fly collection.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Text S1.
Preparation of Abate dilutions.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ekanya, R., Obie, E.D., Hamill, L. et al. The preparatory phase for ground larviciding implementation for onchocerciasis control in the Meme River Basin in South West Cameroon: the COUNTDOWN Consortium alternative strategy implementation trial. Parasites Vectors 15, 219 (2022). https://doi.org/10.1186/s13071-022-05300-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05300-z