
Abstract
Background
In the inner city of Yogyakarta, Indonesia, insecticide resistance is expected in the main dengue vector, Aedes aegypti, because of the intensive local application of pyrethroid insecticides. However, detailed information about the nature of resistance in this species is required to assist the release of Wolbachia mosquitoes in a dengue control program, so that we can ensure that insecticide resistance in the strain of Ae. aegypti being released matches that of the background population.
Methods
High-resolution melt genotyping was used to screen for kdr mutations associated with pyrethroid resistance in the voltage-sensitive sodium channel (VSSC) gene in Ae. aegypti of some areas in the inner city of Yogyakarta.
Results
The results show that the V1016G mutation predominated, with individuals homozygous for the 1016G allele at a frequency of 82.1% and the mutant allele G at a frequency of 92%. Two patterns of co-occurrence of mutations were detected in this study, homozygous individuals V1016G/S989P; and heterozygous individuals V1016G/F1534C/S989P. We found the simultaneous occurrence of kdr mutations V1016G and F1534C at all collection sites, but not within individual mosquitoes. Homozygous mutants at locus 1016 were homozygous wild-type at locus 1534 and vice versa, and heterozygous V1016G were also heterozygous for F1534C. The most common tri-locus genotype co-occurrences were homozygous mutant 1016GG and homozygous wild-type FF1534, combined with homozygous mutant 989PP (GG/FF/PP) at a frequency of 38.28%.
Conclusions
Given the relatively small differences in frequency of resistance alleles across the city area, locality variations in resistance should have minor implications for the success of Wolbachia mosquito trials being undertaken in the Yogyakarta area.
Similar content being viewed by others

Background
Dengue is the most important mosquito-borne viral disease in the world [1]. Dengue is also a significant public health problem in Indonesia since the first reported dengue outbreak in 1968. All four dengue virus (DENV, Flavivirus) serotypes are detected, suggesting hyperendemicity in all of 34 provinces in Indonesia [2, 3]. Dengue virus is transmitted by Aedes aegypti, a mosquito that has also been linked with the transmission of other major arboviral diseases including chikungunya (CHIKV) and Zika (ZIKV). The virologically confirmed chikungunya was initially reported during an outbreak in Jambi in 1982 [4]. After nearly 20 years of absence, twenty-four distinct outbreaks of probable chikungunya re-emerged in South Sumatera, Aceh and West Java in the early 2000s [4]. To date, there has been no recorded fatality related to CHIKV infection in Indonesia [5]. Zika virus (ZIKV, family Flaviviridae) has become one of the global public health threats because of its association with Guillain-Barré syndrome and microcephaly. Until recently, most evidence for Zika virus infection reported in Asia, including in Indonesia, has been serological [6].
Indonesia has made dengue a notifiable disease. On average, 136,670 DENV infections and 1112 deaths were reported annually from 2013 to 2016, an incidence of about 54 dengue fever or dengue hemorrhagic fever cases per 100,000 population and a case fatality rate of approximately 1% [7]. Dengue cases in Indonesia are expected to be under-reported due to poor disease surveillance and a low level of reporting [8]. The surveillance database likely covers dengue probable cases with supportive dengue serology or with epidemiologic linkage, and dengue confirmed cases with confirmatory laboratory criteria [8, 9]. Dengue infections deprived of diagnostic tests may not end up in the surveillance database [8]. Despite the level of uncertainty on total case number, dengue visibly results in a considerable cost to the health sector, and a heavy economic and social impact is likely.
Indonesia has made progress in many areas of dengue prevention and control. In August 2016, Indonesia became the sixth country to approve the first licensed dengue vaccine, Dengvaxia® [10, 11]. To date, the vaccine so far has been approved in 20 countries and launched in 11, including Indonesia [11]. However, in December 2017, the National Agency of Drug and Food (Badan POM) suspended the use of dengue vaccine in Indonesia over safety concerns [12] and later in February 2018, the BPOM revised recommendation that the use of vaccine be limited for people between 9–16 years of age who had a dengue infection prior to vaccination and that the vaccine should not be administered to people who have never had a dengue infection before [13]. There is no rapid, reliable test for previous dengue infection, so the Dengvaxia® vaccine cannot be widely used [14]. Nevertheless, effective vector control methods are still essential, targeting Ae. aegypti in its immature and adult stages. It has been argued that even if the current vaccine is highly targeted and low-cost, sustained mosquito control will remain cost-effective [15].
The mainstay of the current vector control programme in Indonesia is environmental management, which in recent years, has emphasized community participation to reduce container breeding sites [2, 16]. The country is renowned for its 3M plus campaigns, aimed at covering and cleaning water containers and burying discarded water containers, complemented with biological approaches using natural predators or pathogens as alternatives for vector control [2, 16]. The efficacy of community-based approaches is measured by a larvae-free index (percentage of houses free from Ae. aegypti larvae and pupae infestation); unfortunately, data from the last five years showed a larvae-free index ranging between 24–80%, less than the target of 95% [7].
Chemical insecticides still play a central role in dengue vector control in Indonesia. Vector control during outbreaks depends primarily on chemical insecticides to effect a rapid reduction in the number of infected mosquitoes and to break the dengue transmission cycle [2, 16,17,18]. Since the 1970s, the organophosphates malathion and temephos have been widely used to control dengue, and starting in the 1980s, dengue vector control has been highly reliant on pyrethroids. Pyrethroids are also widely used in public health for prevention and control of other mosquito-borne diseases such as malaria and filariasis, and as agricultural insecticides [19]. Pyrethroids are also approved for household protection against dengue [20, 21]. The abundant and prolonged use of pyrethroids has led to the development of resistance in Ae. aegypti populations in many countries including Indonesia. Resistance to pyrethroids, based on bioassay data, has been reported in some areas of Indonesia [22,23,24,25,26].
Two main mechanisms for pyrethroid resistance have been identified in Ae. aegypti, metabolic resistance and target site resistance. Metabolic resistance occurs when increased levels or modified activities of one or more detoxifying enzymes result in a more rapid detoxification of the insecticide, preventing the insecticide from reaching its target in the nervous system. Metabolic resistance involves three groups of enzymes, i.e. esterases, multi-function oxidases P450 and glutathione s-transferases (GST) [27,28,29]. Limited studies about metabolic resistance based on biochemical assays are available for Indonesian dengue vectors [26, 30,31,32].
Pyrethroids act on the insect nervous system, targeting the voltage-sensitive sodium channel (VSSC). They modify the gating kinetics of the channel by slowing both the activation and the inactivation, stimulating the nerve cells to produce repetitive discharge that lead to paralysis and death of insects, an effect known as knockdown. Target site resistance is caused by point mutations in the VSSC gene, resulting in amino acid substitutions that affect pyrethroid binding sites, being known as kdr (knockdown resistance) mutations [33, 34]. A total of 12 point mutations in the VSSC of Ae. aegypti have been identified to be associated with kdr resistance to pyrethroids. Only five of these mutations have been functionally confirmed to reduce the sensitivity of mosquito sodium channels to pyrethroids, i.e. S989P, I1011M, V1016G/I, F1534C, and recently V410L [35]. The mutations L982W, G923V, I1011M and V1016G were the first reported sodium channel mutations in permethrin/DDT-resistant populations of Ae. aegypti from various countries [36], located in the domain IIS5 (for L982W and G923 mutations) and IIS6 (for I1011M and V1016G mutations) of the VSSC [36]. Further studies have reported novel mutations, including I1011V and V1016I in Latin American populations [37]. In Asian countries, the F1534C mutation (in domain IIIS6) was detected in Ae. aegypti mosquitoes from Thailand [38] and Vietnam [39]. The S989P mutation, located in linker between domains IIS5–S6, was first reported in Ae. aegypti populations in Thailand [40], the D1763Y mutation (in linker IVS5-S6) was reported in a Taiwan population [41], and mutation T1520I (in domain IIS6) was detected in an Indian population [42]. In 2017, the mutation V410L was first identified in Brazilian strains of Ae. aegypti [43], and in 2018, a novel mutation V419L was found in populations from Colombia [44] (both mutations are located in domain IS6).
Three of these mutations, V1016G, F1534C and S989P, are widely distributed and detected in pyrethroid-resistant populations in Southeast Asian countries including Thailand, Indonesia, Malaysia, Singapore, Vietnam, Cambodia and Laos [45]. Co-occurrence of kdr mutations has been a common phenomenon observed and, for some combinations, has been shown to confer a higher level of resistance than singly occurring mutations [46]. At least three patterns of mutational associations have been identified in Ae. aegypti from Southeast Asia: V1016G/F1534C, V1016G/S989P and V1016G/F1534C/S989P. Co-occurrence of V1016G and F1534C was reported in populations from Thailand [47], Myanmar [46], Malaysia [48] and Indonesia [49,50,51,52] and co-occurrence of V1016G and S989P point mutations was detected in Thailand [47], Myanmar [46], Indonesia, [49, 50, 52] and Papua New Guinea [53]. In addition, co-occurrence of triple mutations V1016G/F1534C/S989P in heterozygous form has been identified commonly in Ae. aegypti from Thailand [54], Myanmar [46] and Indonesia [49, 50, 52]. However, co-occurrence of triple homozygous point mutations (homozygous mutation for each of V1016G, F1534C and S989P) is very rare having only been reported from Myanmar at a frequency of 0.98% [46] and in Indonesia in one individual (at a frequency of 0.34%) [50]
Despite all of the strategies implemented in Indonesia, the existing methods of controlling dengue have limited success. The development of a novel strategy of vector control that can be incorporated into the existing vector control strategy is essential. One of the innovative approaches to preventing transmission of dengue virus involves introduction of strains of the bacterium Wolbachia into Ae. aegypti, which has both life-shortening effects on the mosquito and direct transmission-blocking effects on dengue virus. This new strategy, together with more traditional approaches for vector control including insecticide application, may provide promising results to reduce dengue transmission. With the support of communities and approval from regulators, Indonesia’s first field trial of Wolbachia-infected mosquitoes began in Yogyakarta in 2014 [55]. The first field trial in two areas in the outer city of Yogyakarta has yielded results that point to local invasion [56] and release of Wolbachia-infected mosquitoes on a broader scale in the inner city areas of Yogyakarta has since been initiated in August 2016 [57].
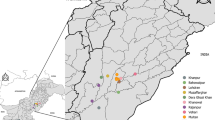
To assist the spread of Wolbachia, insecticide resistance in the strain of Ae. aegypti being released, needs to match that of the background population to ensure that released individuals persist and reproduce, allowing a Wolbachia invasion to take place [58]. In the inner city of Yogyakarta, resistance is expected because of the local heavy application of chemical insecticides. It is possible that insecticide usage is higher in these areas than in the outer rim of Yogyakarta, as inner city areas have more dengue cases [59] and are more densely populated [60]. In a previous study, we screened samples from Yogyakarta outer areas for kdr mutations and obtained a high frequency of kdr alleles in samples collected from the outer area of Yogyakarta [49]. The most common kdr co-occurrence was V1016G homozygous mutant/F1534 wild type, combined either with S989P heterozygous, or S989P wild type, or S989P homozygous mutant, respectively. This co-occurrence was more common in the surviving than in the dead mosquitoes [49] and a strong association between the V1016G mutation with pyrethroid resistance (type I and type II) was confirmed. In contrast, F1534C mutant homozygotes were rare and there was only a weak association between heterozygote individuals for the F1534C mutation and resistance to a type I pyrethroid. The S989P mutation, in addition to the V101G mutation, was found to have an additive effect in resistance to Type II pyrethroids [49]. In this study, we have screened Yogyakarta inner city areas for kdr mutations, based on a total of 1314 individuals collected from 27 localities (Fig. 1, Table 1) and aim to compare frequency and occurrence of kdr mutations between the inner and outer city.

Sample collection sites for Aedes aegypti from inner city of Yogyakarta, Indonesia. The map was sourced from ‘Map of case control area borders, city of Yogyakarta, Eliminate Dengue Project Yogyakarta’
Methods
Mosquito samples
Aedes aegypti samples (male and female adults) were collected in June-July 2015 using BioGent-Sentinel (BG-S) traps (Biogents AG, Regensburg, Germany), each locality was covered by several BG-traps. Samples were collected every week and identified to species level and A. aegypti were preserved in absolute ethanol.
High-resolution melt (HRM) genotyping of kdr mutations in Ae. aegypti samples from Yogyakarta
A total of 1293 samples from 27 collection sites were screened for kdr mutations at 1016, 1534 and 989 following the HRM assay protocol of our previous study [49]. The PCR primer pairs, the region amplified in VSSC and the product size are shown in Table 2.
We used the High Pure PCR Template Preparation Kit (Roche, Mannheim, Germany) to extract mosquito DNA in a final volume of 200 μl of elution buffer and prepared a 10-fold dilution of the template DNA. PCR reactions (10 μl) contained 2 μl of 1 in 10 diluted template DNA, 0.4 μl each of the primers at 10 μM, 1 μl of the ThermoPol reaction buffer (NEB Inc., Ipswich, MA, USA), 0.064 μl of dNTPs at 25 mM (Bioline, Alexandria, NSW, Australia), 0.4 μl of MgCl2 (50 mM) (Bioline, Alexandria, NSW, Australia), 0.25 μl of the LightCycler® 480 High Resolution Melting Master (Roche, Mannheim, Germany), 0.01 μl of IMMOLASETM DNA polymerase (10 u/μl) (Bioline, Alexandria, NSW, Australia) and 5.476 μl of ddH2O (Honeywell, Burdick and Jackson; Muskegon, MI, USA).
Amplification was performed on a Roche LightCycler® 480 system (384-well format) using the following temperature cycling conditions: 95 °C for 10 min, 20 cycles of 95 °C for 5 s, 65 °C (reduce 0.5 °C each cycle) for 15 s, 72 °C for 15 s, followed by an additional 20 cycles of 95 °C for 5 s, 55 °C for 15 s and 72°C for 15 s. Fluorescence information was captured at the end of each 72 °C step. Following amplification, PCR products were subjected to HRM analysis. The HRM step involved heating the PCR products to 95 °C for 1 min, cooling to 40 °C for 20 s and then increasing the temperature to 65 °C and then melting at 0.05 °C/s with continuous acquisition of fluorescence as the temperature increased from 65 to 95 °C. Melt curves were generated in the Gene Scanning module of the Roche LightCycler® 480 software package. Melting data were normalized, temperature shifted, and displayed as derivative curves compared to the known genotyped samples. We used the same PCR protocol and thermocycling conditions, and HRM step for all kdr mutations except the parameter settings for melt curve normalization.
Statistical analysis
Statistical comparisons of locations for genotypic frequencies for the V1016G, F1534C and S989P loci were undertaken using a Chi-square contingency test (IBM SPSS version 22) with degrees of freedom dictated by the number of localities being compared. We also tested for spatial signature in the data using the average GPS points for the BG traps in a locality and then computing Moran’s I using the software Spatial Analysis in Macroecology [61].
Hardy-Weinberg equilibrium for observed genotyped frequencies for each kdr mutation was calculated using GenAlEx 6.5 (http://biology.anu.edu.au/GenAlEx/) [62, 63]. Linkage disequilibrium between the two-kdr mutations, V1016G and F1534C, was also tested. Data were pooled across adjacent sites to increase sample size for detecting linkage disequilibrium.
We identified three patterns of mutational associations: (i) nearly all homozygous mutant individuals 1016GG occurred in conjunction with homozygous wild-type 1534FF; (ii) homozygous mutant individuals 1534CC occurred with the wild-type 1016VV; and (iii) heterozygous individuals for V1016G were commonly heterozygous for F1534C. We used Pearson’s correlation (MS Excel v.16.23) to measure the strength of relationship of these mutational associations.
Results
HRM genotyping of kdr mutations in Ae. aegypti samples from Yogyakarta
A total of 1314 of Ae. aegypti were collected from 27 different sites in the inner city of Yogyakarta (Fig. 1, Table 1). Of these, we genotyped 1293 for kdr mutations at V1016G, F1534C and S989P sites using the high-resolution melt (HRM) protocol [49] and compared these according to their geographical location (Fig. 1). V1016G was the most widespread kdr point mutation and was detected in all collection sites: 82.06% of the individuals had the homozygous mutant genotype 1016GG, 16.86% were heterozygous 1016VG and the remaining 1.08% were homozygous wild type 1016VV (Table 3). The allelic frequencies of V1016 and 1016G were 0.105 and 0.895, respectively. All collection sites displayed a high frequency of the mutant allele 1016G, ranging between 0.760–1.000. The observed frequency of the mutant allele 1016G was statistically different between collection sites as indicated by a Chi-square contingency test (\(\chi_{(26)}^{2}\) = 44.145, P = 0.015). For the 1016 allele frequencies, Moran’s I was not significant for any distance class, indicating locality differences unrelated to distance.
The second most common kdr mutation detected in our samples was S989P. The frequencies of genotypes detected overall were as follows: homozygous wild-type 989SS, 14.54%; heterozygous 989SP, 47.18%; and homozygous mutant 989PP, 38.28%. Allelic frequencies overall were 0.599 for S989 and 0.401 for 989P. The mutant allelic frequency 989P ranged between 0.360–0.700 across localities and overall there was a statistically significant difference between localities (Chi-square contingency test, \(\chi_{(26)}^{2}\)= 43.473, P = 0.017). For the 989 allele frequencies, Moran’s I did not show a significant departure from randomness at the smallest distance class (0.85 km), but there was a positive association at 1.7 km and negative association at the next class up (2.2 km), suggesting a complex pattern not related directly to distance.
For the 1534 site, the homozygous wild-type genotype 1534FF was the most common at 83.06%, while the homozygous mutant genotype 1534CC was rare at 1.00% and heterozygous genotype 1534F/C was at 15.94%. A contingency test indicates no significant difference between the localities in allele frequencies (\(\chi_{(26)}^{2}\)= 32.151, P = 0.188). Moran’s I was not significant for any distance classes for the 1534 alleles.
All genotypes at the three loci in all field collections were in Hardy-Weinberg equilibrium except for Co1 (Rejowinangun) (Table 3). When combined across sites, genotype frequencies were also in Hardy-Weinberg equilibrium (Table 4).
Mutation combinations
We observed that there were nine out of 27 possible genotype combinations (three genotypes at three loci) in our 1293 individual mosquitoes (Fig. 2). Figure 2 shows the frequency of each of the nine tri-locus combinations. The most common tri-locus genotypes were the homozygous mutant 1016GG and homozygous wild-type FF1534, combined with either the homozygous mutant 989PP (GG/FF/PP) (38.28%), the heterozygous S989P (GG/FF/SP) (35.43%), or the homozygous wild-type SS989 (GG/FF/SS) (8.35%). These genotypes totalled 495, 458 and 108 mosquitoes, respectively. Triple mutations were found only in the heterozygous genotype state (VG/FC/SP) at a frequency of 11.14% (144 individuals).

BG trap deployment (mark as triangles) at sample collection sites for Aedes aegypti from inner city of Yogyakarta, Indonesia. The map was sourced from ‘Map of case control area borders, city of Yogyakarta, Eliminate Dengue Project Yogyakarta’
The V1016G homozygote mutant genotype was found alone (GG/FF/SS) in 108 (8.35%) individuals, and there were 8 (1.00%) individuals exhibiting the homozygous mutant genotype for F1534C (VVCCSS). S989P was only found in conjunction either with V1016G (as a homozygote or heterozygote) or F1534C (only as a heterozygote). We only obtained one wild- type individual for the three mutations (VV/FF/SS), likewise there was no individual that expressed triple mutations (homozygous mutants for each site V1016G, F1534C and S989P) among the 1293 mosquito individuals analysed.
Interestingly, we found that nearly all homozygous mutant 1016GG individuals occurred in conjunction with homozygous wild-type 1534FF, and homozygous mutant individuals 1534CC occurred with the wild-type 1016VV. Heterozygous individuals for V1016G were commonly heterozygous for F1534C, although there was a small number of combined homozygous/heterozygous genotypes (Table 3, Fig. 2). Pearson’s correlation was used to assess these relationship patterns and there was a significant positive correlation between co-occurrence of 1016G homozygous mutants and F1534 wild-type at all collection sites (r = 0.999, n = 27, P < 0.0005), with 1016G homozygous mutants explaining 99.8% of the variation in V1016G mutations (Fig. 3). A strong positive correlation was also found between the co-occurrence of individuals with heterozygous V1016G and heterozygous F1534C (r = 0.988, n = 27, P < 0.0005) (Fig. 4). The most common genotype for V1016C and F1534C was the homozygous mutant genotype 1016GG combined with the homozygous wild-type 1534FF with a frequency of 82.1%, followed by the double heterozygous 1016VG with 1534FC which had a frequency of 9.74% and finally the homozygous wild-type 1016VV combined with the homozygous mutant 1534CC which had a frequency of 1.01%. However, there were 12 individuals with heterozygous 1016VG that were combined with homozygous wild-type 1534FF. Table 4 shows the linkage disequilibrium coefficients (Rij), chi-square and associated probabilities obtained between pairwise loci. To calculate linkage disequilibrium, samples from adjacent collection sites were pooled. In all pooled collection sites, there was very strong linkage disequilibrium between V1016G and F1534C, with coefficients (Rij) ranged from 0.863 to 1.

The combinations of kdr mutations V1016G (T/G) F1534C (T/G) and S989P (T/C) in Aedes aegypti samples collected from the inner city of Yogyakarta

Correlation between the number of Aedes aegypti individuals with homozygous mutant genotype V1016G and those with homozygous wild-type genotype F1534C across sample areas
We also compared V1016G, F1534C and S989P from those sites where data were available from our previous study in 2012 [49] with this study in 2015. Most allele frequencies were similar between the two sampling periods and did not differ significantly by contingency tests, but there was a significant difference at Cokrodiningratan for S989 (\(\chi_{(1)}^{2}\) = 9.88, P = 0.002) where frequency of the mutant 989P increased (Table 5).
Discussion
The diagnostic identification of the kdr mutations in Ae. aegypti natural populations is nowadays an important tool to predict resistance to pyrethroids in the field. In this study, we genotyped kdr mutations (F1534C, V1016G and S989P) that have been confirmed as being associated with pyrethroid resistance in the VSSC of Ae. aegypti [49, 50, 64, 65]. The homozygous V1016G mutation predominated, with individuals homozygous for the 1016G allele being at a frequency of 82.1% and the mutant allele (G) being at a frequency of 92%. The high frequency of the mutant allele is similar to previous findings for Ae. aegypti samples from the outer region of Yogyakarta [49], and some other provinces in Indonesia, including Denpasar (70% in a resistant group and 50% in susceptible groups [52], Jakarta (65% in a resistant group and 35% in a susceptible group) [51], and Central Java [50]. In Asia, a high frequency of the mutant allele V1016G is also common, including in Myanmar (84.4% [46]), and in southern China (95.5% [66]). The V1016G mutation was detected at a lower frequency in Thailand [47], Vietnam [46] and Malaysia [48].
For locus 1534, the homozygous wild-type genotype was frequent at all collection sites. A high frequency of the wild-type genotype was also found in Ae. aegypti mosquitoes from Jakarta (70.70%), Central Java (91.16%) [50] and Denpasar (63.91%) [52]. The 1534C allele frequency in this study (8%) was lower compared to the neighbouring countries of Southeast Asia, including Malaysia (40-80%) [48], Vietnam (22%) [46], Myanmar (21%) [46] and Thailand (51%) [67].
In the present study, there were three patterns of co-occurrence of point mutations, namely, V1016G/F1534C, V1016G/S989P and V1016G/F1534C/S989P. We found the simultaneous occurrence of kdr mutations V1016G and F1534C in all collection sites. Homozygous mutants at locus 1016 had homozygous wild-type alleles at locus 1534 and vice versa, and heterozygous V1016G were also heterozygous for F1534C, apart from a small number of combined homozygous/heterozygous genotypes. The main co-occurrence was homozygous mutants at 1016 and homozygous wild-type alleles at 1534 (1016G/F1534 haplotype) at a frequency of 82.1%. Pearson’s correlation analysis strongly supports statistical associations between mutations in all collection sites. Correspondingly, a similar linkage association is also found in the Ae. aegypti population from our previous study, where these genotypes occurred at 69.2% [49], and from Myanmar where the equivalent frequency was 59.3% [46]. In contrast, the more common association in Ae. aegypti from Thailand was a homozygous F1534C mutation with V1016G wild-type at frequency 43.5% [47]. Double mutants of 1016G and 1534C were never found. This is consistent with other studies, which have noted that in natural populations of Ae. aegypti, the double homozygotes for V1016G and F1534C mutations are rare [46,47,48,49] or absent [47], suggesting a possible fitness cost related to this haplotype or absence of recombination to date to bring homozygotes for V1016G and F1534C mutations onto the same haplotype [54]. However, a small number of mosquitoes homozygous for 1016G and 1534C occur in Malaysia [48] and Myanmar [46]. Diverse haplotypes associated with V1016G and F1534C co-occurrences across the countries suggest that selection has acted differently across sites perhaps related to pyrethroid usage, or else that different genetic backgrounds have had an impact on evolutionary trajectories [68]. Given the linkage disequilibrium between V1016G and F1534C, there is a non-random association of alleles at the two kdr loci which may indicate a selection process acting simultaneously at these two loci.
The co-occurrence of the S989P mutation with the V1016G mutation has been noted in previous studies from some other Asian countries [46, 66, 68,69,70]. However, it also appears that the S989P mutation can be found in the absence of the V1016G mutation. A small number of homozygous and heterozygous mutants at S989P were found to coexist with the F1534C mutation instead of the V1016G mutation in Ae. aegypti samples from Yogyakarta outer city areas [49] and Central Java [50]. Additionally, the S989P mutation has been reported as occurring alone in samples from Central Java [50]. The most common tri-locus genotype co-occurrences were homozygous mutant 1016GG and homozygous wild-type FF1534, combined with homozygous mutant 989PP (GG/FF/PP), followed by heterozygous S989P (GG/FF/SP). These results are somewhat different from our previous findings, which detected the most common combination being the homozygous mutant 1016G and homozygous wild-type F1534, combined with heterozygous S989P which was recorded at 39.75% in the resistant group, 26.83% in the susceptible group, and 31% in the field collection [49]. These differences may reflect local selection processes for resistance that differ between the city and outer areas of Yogyakarta.
The spatial variation in resistance allele frequencies found in this study may have minor implications for Wolbachia mosquito trials being undertaken in the Yogyakarta area [56]. At present, some areas of the city have been invaded by Wolbachia whereas other areas are acting as uninvaded controls. The expectation is that dengue incidence will be lower in invaded areas because there is no local transmission or at least reduced transmission where Wolbachia is common, based on the fact that the wMel strain released in Ae. aegypti mosquitoes partially blocks dengue transmission [71]. However, if there are differences in pesticide resistance across areas, then this may confound interpretations because increased resistance may be associated with a higher local density of mosquitoes which could influence transmission as well. This problem will be alleviated to some extent by backcrossing the Wolbachia mosquito release strain to the local wild mosquitoes, so that the local insecticide resistance genetic profile is introgressed to the release strain [56]. Given the relatively small differences in frequency of resistance alleles, we do not anticipate large confounding effects of locality variation in resistance, but it is nevertheless worthwhile controlling for such an effect when comparing impacts of Wolbachia (Fig. 5).

Correlation between the number of Aedes aegypti individuals with heterozygous mutant genotype V1016G and heterozygous mutant genotype F1534C across sampling areas
Finally, the data suggest that there is no spatial structure to variation in resistance mutations across the city area. This seems to point to local selection events driving resistance frequencies rather than migration at this scale. Still, any attempts to manage pesticide resistance such as through rotating active ingredients will need to be mindful of selection pressures that can locally increase resistance and aim to alter behaviour across the entire inner-city area.
Conclusions
This study identified kdr mutations in Ae. aegypti from some areas in the inner city of Yogyakarta managed for Wolbachia releases. We detected the highest frequency of the V1016G mutation, and the lowest frequency of the F1534C mutation. The low frequency of the F1565C mutation in the samples may indicate that Type I pyrethroids have not been used as extensively as Type II pyrethroids. We also observed the pattern of the simultaneous occurrence of kdr mutations V1016G and F1534C: the V1016/C1534 double homozygote (VV/CC); the G1016/C1534 double heterozygote (VG/FC) and the G1016/F1534 homozygote (GG/FF); and a small amount of combined heterozygous/ homozygous (VG/FF) genotypes. Given the linkage disequilibrium between V1016G and F1534C, there is a non-random association of alleles at the two kdr loci which may indicate a selection process acting simultaneously at these two loci. The most common tri-locus genotype co-occurrences were a homozygous mutant 1016GG and a homozygous wild-type FF1534, combined with a homozygous mutant 989PP (GG/FF/PP). This is a bit different from the co-occurrence from Yogyakarta outer city samples which was a homozygous mutant 1016G and a homozygous wild-type F1534, combined with a heterozygous S989P. These differences are perhaps due to their having different local selection processes for resistance between the city and outer areas of Yogyakarta. The relatively low spatial variation in resistance allele frequency across the city may have minor implication for Wolbachia mosquito trials.
Availability of data and materials
Data are presented in the main manuscript.
References
WHO. Global strategy for dengue prevention and control 2012–2020. France: World Health Organization; 2014.
Indonesia KKR. Demam berdarah dengue di Indonesia tahun 1968–2009. Buletin Jendela Epidemiologi. 2010;2:1–14.
Indonesia KKR. Situasi DBD di Indonesia. Edited by Indonesia KKR. Jakarta: Pusdatin Kementrian Kesehatan Republik Indonesia; 2016.
Laras K, Sukri NC, Larasati RP, Bangs MJ, Kosim R, Djauzi, et al. Tracking the re-emergence of epidemic chikungunya virus in Indonesia. Trans R Soc Trop Med Hyg. 2005;99:128–41.
Harapan H, Michie A, Mudatsir M, Nusa R, Yohan B, Wagner AL, et al. Chikungunya virus infection in Indonesia: a systematic review and evolutionary analysis. BMC Infect Dis. 2019;19:243.
Yudhaputri FA, Trimarsanto H, Perkasa A, Yohan B, Haryanto S, Wiyatno A, et al. Genomic characterization of Zika virus isolated from Indonesia. Virology. 2017;510:248–51.
Indonesia KKR. Profil kesehatan Indonesia tahun 2016. Jakarta: Kementerian Kesehatan Republik Indonesia; 2017.
Karyanti MR, Uiterwaal CSPM, Kusriastuti R, Hadinegoro SR, Rovers MM, Heesterbeek H, et al. The changing incidence of dengue haemorrhagic fever in Indonesia: a 45-year registry-based analysis. BMC Infec Dis. 2014;14:412.
WHO. Comprehensive guidelines for prevention and control of dengue and dengue haemorrhagic fever. Revised and expanded edition. India: World Health Organization Regional Office for South-East Asia; 2011.
Badan POM Republik Indonesia. The Drug and Food Monitoring Agency of Indonesia has approved dengue vaccine distribution permit in Indonesia. 2017. http://pionas.pom.go.id/artikel/badan-pom-menyetujui-izin-edar-vaksin-dengue-di-indonesia. Accessed 20 May 2018.
Pasteur S. First dengue vaccine approved in more than 10 countries. Lyon: Sanofi Pasteur; 2016.
Badan POM Republik Indonesia. The Drug and Food Monitoring Agency of Indonesia clarifies concerns about dengue vaccine safety. 2017. https://www.pom.go.id/new/view/more/klarifikasi/72/PENJELASAN-BADAN-POM-RI–TERKAIT-ISU-KEAMANAN-VAKSIN-DENGUE–DEMAM-BERDARAH-.html. Accessed 20 May 2018.
Badan POM Republik Indonesia. The Drug and Food Monitoring Agency of Indonesia clarifies concerns about dengue vaccine safety. 2018. http://www.pom.go.id/new/view/more/klarifikasi/79/PENJELASAN-BADAN-POM-RI—-TERKAIT–ISU-KEAMANAN-VAKSIN-DENGUE–DEMAM-BERDARAH-.html. Accessed 20 May 2018.
The dengue vaccine dilemma. Lancet Infect Dis. 2018;18:123.
Fitzpatrick C, Haines A, Bangert M, Farlow A, Hemingway J, Velayudhan R. An economic evaluation of vector control in the age of a dengue vaccine. PLoS Negl Trop Dis. 2017;11:e0005785.
Kementerian Kesehatan Republik Indonesia. Technical guideline for implementing mosquito larval source reduction (PSN 3M-Plus), and 1 house 1 mosquito larval inspector. Jakarta: Kementerian Kesehatan RI; 2016. http://www.jumantik.org/images/book/Juknis_1_Rumah_1_Jumantik.pdf.
Kementerian Kesehatan Republik Indonesia General information about dengue haemorrhagic fever. 2011. https://www.scribd.com/doc/108669362/Copy-of-Informasi-Umum-Dbd-2011.
Kementerian Kesehatan Republik Indonesia Module for dengue haemorrhagic fever prevention. Jakarta: Direktorat Jenderal Pengendalian Penyakit Dan Penyehatan Lingkungan; 2011. https://www.scribd.com/document/364139674/Modul-Pengendalian-Demam-Berdarah-Dengue.
Ministry of Health Republic of Indonesia. Review of national vector control policy in Indonesia. 2010. http://www.actmalaria.net/files/vector_control/i_resistance/Indonesia.pdf. Accessed 27 Mar 2012.
Pratamawati DA, Irawan AS, Widiarti W. Relationship between knowledge of vector with household insecticide usage behavior in dengue hemorrhagic fever endemic areas in Bali Province. Vektora. 2012;4:99–116.
Setyawati DR. Knowledge, behaviour and practice on household insecticide usage in Jakarta and Surabaya. 2010. http://repository.ipb.ac.id/handle/123456789/27154. Accessed 25 Jan 2016.
Ikawati B, Sunaryo S, Widiastuti D. Map of the susceptibility status of Aedes aegypti (Linn) to insecticide cypermethrin and malathion in Central Java. Aspirator. 2015;7:23–8.
Widiarti W, Heriyanto B, Boewono DT, Widyastuti U, Mujiono M, Lasmiati L, et al. Map of insecticide resistance status of dengue haemorrhagic fever vector Aedes aegypti to organophosphates, carbamates and pyrethroids in Central Java and D.I. Yogyakarta Yogyakarta. Bul Pen Kes. 2011;39:176–89.
Ahmad I, Astari S, Rahardjo B, Tan M, Munif A. Resistance of Aedes aegypti from three provinces in Indonesia, to pyrethroid and organophosphate insecticides. In: International Conference on Mathematics and Natural Sciences (ICMNS). Bandung, Indonesia; 2006. https://sith.itb.ac.id/wp-content/uploads/sites/56/2016/06/Resistance-aegyptyi-to-pyrethroids-intan-ahmad.pdf.
Mantolu Y, Kustiati K, Ambarningrum TB, Yusmalinar S, Ahmad I. Status and development of resistance in Aedes aegypti (Linnaeus) (Diptera: Culicidae) strains of Bandung, Bogor, Makassar, Palu, and VCRU against permethrin insecticide over five generations of selection. J Entomol Indonesia. 2016;13:1–8.
Putra RE, Ahmad I, Prasetyo DB, Susanti S, Rahayu R, Hariani N. Detection of insecticide resistance in the larvae of some Aedes aegypti (Diptera: Culicidae) strains from Java, Indonesia to temephos, malathion and permethrin. Int J Mosq Res. 2016;3:23–8.
Hemingway J. The molecular basis of two contrasting metabolic mechanisms of insecticide resistance. Insect Biochem Mol Biol. 2000;30:1009–15.
Hemingway J, Ranson H. Insecticide resistance in insect vectors of human disease. Ann Rev Entomol. 2000;45:371–91.
Hemingway J, Hawkes NJ, McCarroll L, Ranson H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem Mol Biol. 2004;34:653–5.
Ahmad I, Astari S, Tan M. Resistance of Aedes aegypti (Diptera: Culicidae) in 2006 to pyrethroid insecticides in Indonesia and its association with oxidase and esterase levels. Pak J Biol Sci. 2007;10:3688–92.
Astari S, Ahmad I. Insecticide resistance and effect of piperonyl butoxide as a synergist in three strains of Aedes aegypti (Linn.) (Diptera: Culicidae) against insecticides permethrin, cypermethrin, and d-allethrin. Bul Pen Kes. 2005. 33:73–9.
Widiastuti D, Sunaryo S, Pramestuti N, Martini M. Monooxygenase activities in Aedes aegypti mosquito population of Tembalang district. Semarang. Aspirator J Vector-borne Dis Stud. 2015;7:1–6.
Soderlund DM. Pyrethroids, knockdown resistance and sodium channels. Pest Manag Sci. 2008;64:610–6.
Dong K, Du Y, Rinkevich F, Nomura Y, Xu P, Wang L, et al. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem Mol Biol. 2014;50:1–17.
Saavedra-Rodriguez K, Maloof FV, Campbell CL, Garcia-Rejon J, Lenhart A, Penilla P, et al. Parallel evolution of VGSC mutations at domains IS6, IIS6 and IIIS6 in pyrethroid resistant Aedes aegypti from Mexico. Sci Rep. 2018;8:6747.
Brengues C, Hawkes NJ, Chandre F, McCarroll L, Duchon S, Guillet P, et al. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med Vet Entomol. 2003;17:87–94.
Saavedra-Rodriguez K, Urdaneta-Marquez L, Rajatileka S, Moulton M, Flores AE, Fernandez-Salas I, et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol Biol. 2007;16:785–98.
Yanola J, Somboon P, Walton C, Nachaiwieng W, Prapanthadara L. A novel F1552/C1552 point mutation in the Aedes aegypti voltage-gated sodium channel gene associated with permethrin resistance. Pestic Biochem Physiol. 2010;96:127–31.
Kawada H, Higa Y, Komagata O, Kasai S, Tomita T, Thi Yen N, et al. Widespread distribution of a newly found point mutation in voltage-gated sodium channel in pyrethroid-resistant Aedes aegypti populations in Vietnam. PLoS Negl Trop Dis. 2009;3:e527.
Srisawat R, Komalamisra N, Eshita Y, Zheng M, Ono K, Itoh TQ, et al. Point mutations in domain II of the voltage-gated sodium channel gene in deltamethrin-resistant Aedes aegypti (Diptera: Culicidae). Appl Entomol Zool. 2010;45:275–82.
Chang C, Shen WK, Wang TT, Lin YH, Hsu EL, Dai SM. A novel amino acid substitution in a voltage-gated sodium channel is associated with knockdown resistance to permethrin in Aedes aegypti. Insect Biochem Mol Biol. 2009;39:272–8.
Kushwah RBS, Dykes CL, Kapoor N, Adak T, Singh OP. Pyrethroid-resistance and presence of two knockdown resistance (kdr) mutations, F1534C and a novel mutation T1520I, in Indian Aedes aegypti. PLoS Negl Trop Dis. 2015;9:e3332.
Haddi K, Tomé HVV, Du Y, Valbon WR, Nomura Y, Martins GF, et al. Detection of a new pyrethroid resistance mutation (V410L) in the sodium channel of Aedes aegypti: a potential challenge for mosquito control. Sci Rep. 2017;7:46549.
Granada Y, Mejía-Jaramillo AM, Strode C, Triana-Chavez O. A point mutation V419L in the sodium channel gene from natural populations of Aedes aegypti is involved in resistance to lambda-cyhalothrin in Colombia. Insects. 2018;9:23.
Amelia-Yap ZH, Chen CD, Sofian-Azirun M, Low VL. Pyrethroid resistance in the dengue vector Aedes aegypti in Southeast Asia: present situation and prospects for management. Parasit Vectors. 2018;11:332.
Kawada H, Oo SZM, Thaung S, Kawashima E, Maung YNM, Thu HM, et al. Co-occurrence of point mutations in the voltage-gated sodium channel of pyrethroid-resistant Aedes aegypti populations in Myanmar. PLoS Negl Trop Dis. 2014;8:e3032.
Stenhouse SA, Plernsub S, Yanola J, Lumjuan N, Dantrakool A, Choochote W, et al. Detection of the V1016G mutation in the voltage-gated sodium channel gene of Aedes aegypti (Diptera: Culicidae) by allele-specific PCR assay, and its distribution and effect on deltamethrin resistance in Thailand. Parasit Vectors. 2013;6:253.
Ishak IH, Jaal Z, Ranson H, Wondji CS. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in the dengue vectors Aedes aegypti and Aedes albopictus from Malaysia. Parasit Vectors. 2015;8:181.
Wuliandari JR, Lee SF, White VL, Tantowijoyo W, Hoffmann AA, Endersby-Harshman NM. Association between three mutations, F1565C, V1023G and S996P, in the voltage-sensitive sodium channel gene and knockdown resistance in Aedes aegypti from Yogyakarta, Indonesia. Insects. 2015;6:658–85.
Sayono S, Hidayati APN, Fahri S, Sumanto D, Dharmana E, Hadisaputro S, et al. Distribution of voltage-gated sodium channel (Nav) alleles among the Aedes aegypti populations in Central Java Province and its association with resistance to pyrethroid insecticides. PLoS ONE. 2016;11:e0150577.
Hamid PH, Prastowo J, Ghiffari A, Taubert A, Hermosilla C. Aedes aegypti resistance development to commonly used insecticides in Jakarta. Indonesia. PLoS ONE. 2017;12:e0189680.
Hamid PH, Prastowo J, Widyasari A, Taubert A, Hermosilla C. Knockdown resistance (kdr) of the voltage-gated sodium channel gene of Aedes aegypti population in Denpasar, Bali Indonesia. Parasit Vectors. 2017;10:283.
Demok S, Endersby-Harshman N, Vinit R, Timinao L, Robinson LJ, Susapu M, et al. Insecticide resistance status of Aedes aegypti and Aedes albopictus mosquitoes in Papua New Guinea. Parasit Vectors. 2019;12:333.
Plernsub S, Saingamsook J, Yanola J, Lumjuan N, Tippawangkosol P, Sukontason K, et al. Additive effect of knockdown resistance mutations, S989P, V1016G and F1534C, in a heterozygous genotype conferring pyrethroid resistance in Aedes aegypti in Thailand. Parasit Vectors. 2016;9:417.
World Mosquito Program. Communities show support for mosquito releases. 2014. http://www.eliminatedengue.com/id/progress/view/news/242/year/2014/month/1/pg/1. Accessed 7 Aug 2018.
Tantowijoyo W, Andari B, Arguni E, Budiwati N, Nurhayati I, Fitriana I, et al. Stable establishment of wMel Wolbachia in Aedes aegypti populations in Yogyakarta, Indonesia. PLoS Negl Trop Dis. 2020;14:e0008157.
Yogyakarta EDP. Yogya City residents are ready to take care of Aedes aegypti Wolbachia infected mosquitoes to control DHF. Yogyakarta: Eliminate Dengue Project. Yogyakarta; 2016.
Garcia GdA, Sylvestre G, Aguiar R, Borges da Costa G, Martins AJ, Lima JBP, et al. Matching the genetics of released and local Aedes aegypti populations is critical to assure Wolbachia invasion. PLoS Negl Trop Dis. 2019;13:e0007023.
Dinas Kesehatan D.I. Yogyakarta. Health profile of D.I. Yogyakarta 2016.Yogyakarta: Dinas Kesehatan D.I. Yogyakarta; 2016. https://www.kemkes.go.id/resources/download/profil/PROFIL_KES_PROVINSI_2016/14_DIY_2016.pdf. Accessed 2 July 2018.
Statistics of D.I. Yogyakarta. The population density by regency/city in D.I. Yogyakarta. 2016. https://yogyakarta.bps.go.id/dynamictable/2017/08/02/31/kepadatan-penduduk-menurut-kabupaten-kota-di-d-i-yogyakarta.html. Accessed 2 July 2018.
Rangel TF, Diniz-Filho JAF, Bini LM. SAM: a comprehensive application for Spatial Analysis in Macroecology. Ecography. 2010;33:46–50.
Peakall R, Smouse PE. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research—an update. Bioinformatics. 2012;28:2537–9.
Peakall R, Smouse PE. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes. 2006;6:2888–295.
Du Y, Nomura Y, Satar G, Hu Z, Nauen R, He SY, et al. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc Natl Acad Sci USA. 2013;110:11785–90.
Hirata K, Komagata O, Itokawa K, Yamamoto A, Tomita T, Kasai S. A single crossing-over event in voltage-sensitive Na+ channel genes may cause critical failure of dengue mosquito control by insecticides. PLoS Negl Trop Dis. 2014;8:e3085.
Li CX, Kaufman PE, Xue RD, Zhao MH, Wang G, Yan T, et al. Relationship between insecticide resistance and kdr mutations in the dengue vector Aedes aegypti in Southern China. Parasit Vectors. 2015;8:325.
Saingamsook J, Saeung A, Yanola J, Lumjuan N, Walton C, Somboon P. A multiplex PCR for detection of knockdown resistance mutations, V1016G and F1534C, in pyrethroid-resistant Aedes aegypti. Parasit Vectors. 2017;10:465.
Plernsub S, Saingamsook J, Yanola J, Lumjuan N, Tippawangkosol P, Walton C, et al. Temporal frequency of knockdown resistance mutations, F1534C and V1016G, in Aedes aegypti in Chiang Mai city, Thailand and the impact of the mutations on the efficiency of thermal fogging spray with pyrethroids. Acta Trop. 2016;162:125–32.
Srisawat R, Komalamisra N, Apiwathnasorn C, Paeporn P, Roytrakul S, Rongsriyam Y, et al. Field-collected permethrin-resistant Aedes aegypti from central Thailand contain point mutations in the domain IIS6 of the sodium channel gene (kdr) Southeast Asian. J Trop Med Public Health. 2012;43:1380–6.
Kasai S, Komagata O, Itokawa K, Shono T, Ng LC, Kobayashi M, et al. Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti: target site insensitivity, penetration, and metabolism. PLoS Negl Trop Dis. 2014;8:e2948.
Walker T, Johnson PH, Moreira LA, Iturbe-Ormaetxe I, Frentiu FD, McMeniman CJ, et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature. 2011;476:450–3.
Acknowledgements
The study was funded by the National Health and Medical Research Council with a Programme Grant/Award Numbers: 1037003, 1132412 and a Fellowship to AAH; NHMRC Fellowship Grant no. 1118640; FNIH Vector-based Control of Transmission: Discovery Research (VCTR) program Eliminate Dengue; JJT Foundation and the Tahija Foundation.
Funding
The funding body had no role in the design of the study and collection, analysis and interpretation of data or in writing the manuscript.
Author information
Authors and Affiliations
Contributions
JRW, NMEH and AAH designed the study. WT collected the samples. JRW collected the data. JRW and AAH analysed the data. JRW, NMEH and AAH wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
No ethics approval was required for this study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wuliandari, J.R., Hoffmann, A.A., Tantowijoyo, W. et al. Frequency of kdr mutations in the voltage-sensitive sodium channel (VSSC) gene in Aedes aegypti from Yogyakarta and implications for Wolbachia-infected mosquito trials. Parasites Vectors 13, 429 (2020). https://doi.org/10.1186/s13071-020-04304-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-04304-x