
Abstract
Background
Most of the studies on fish diseases focus on single infections, although in nature co-infections occur more often. The two freshwater myxozoan parasites of salmonids, having high economic and ecologic relevance are Tetracapsuloides bryosalmonae (Malacosporea), the etiological agent of proliferative kidney disease, and Myxobolus cerebralis (Myxosporea), the etiological agent of whirling disease. The present study aims to investigate immune modulation in rainbow trouts (Oncorhynchus mykiss) during single and co-infections by these parasites.
Methods
Fish were initially infected with T. bryosalmonae (one group) and M. cerebralis (another group) separately. At 30 days post-exposure (dpe), both the single species infected groups were co-infected, respectively, with the other parasite. Posterior kidney and cartilage cranium samples were collected at 30, 60, 90 and 120 dpe and RT-qPCR was performed on them to assess the transcription of suppressors of cytokine signaling (SOCS) -1 and -3, Janus kinase-1 (JAK-1) and signal transducer and activator of transcription-3 (STAT-3) genes.
Results
Kidney samples from the T. bryosalmonae-infected group showed upregulation of all immune genes tested between 60–120 dpe. Crania from the single M. cerebralis-infected group and the M. cerebralis and T. bryosalmonae co-infected group exhibited upregulation of SOCS-1 and JAK-1 between 60–120 dpe and SOCS-3 at 120 dpe. However, only in the single M. cerebralis-infected group, was a statistically significant expression of STAT-3 observed at 30 and 60 dpe.
Conclusions
The results of this study indicate that both T. bryosalmonae and M. cerebralis induce overexpression of SOCS-1 and SOCS-3 genes and modulate the host immune response during the development of parasite to cause immunosuppression.
Similar content being viewed by others


Background
Proliferative kidney disease (PKD) is caused by the myxozoan parasite, Tetracapsuloides bryosalmonae that belongs to the class Malacosprea and phylum Myxozoa [1, 2]. PKD causes high economic losses of farmed and wild autochthonous salmonids and is distributed in the regions of Europe and North America [3,4,5]. The life-cycle of T. bryosalmonae alternates between an invertebrate freshwater bryozoan host (Fredericella sultana) and a vertebrate salmonid host [4, 6]. PKD targets the kidney and induces chronic immunopathology, granulomatous-like lesions and lymphocytic hyperplasia of the interstitial kidney tissue, along with hyperimmunoglobulinemia [7, 8]. Since this disease is temperature-dependent, climate change plays a crucial role in its pathogenesis [5]. The infective T. bryosalmonae malacospores spread either through infected tolerant hosts, such as brown trout (Salmo trutta) [9], or infected bryozoan dispersal, via migrating zooids [10] and infected statoblasts [11]. During PKD the mortality rate can range from less than 20%, to 95–100% in serious outbreaks that are complicated by secondary infections and unfavorable farming or environmental conditions [5, 12, 13].
Fish that recover from PKD acquire a strong immunity and become resistant to re-infections [14]. This results from the massive activation of B cells, which induces hyperimmunoglobulinemia in response to the parasite’s extra-sporogonic histozoic proliferation [8, 15, 16]. Rainbow trout (Oncorhynchus mykiss) show upregulation of tumor necrosis factor (TNF-α2), cyclooxygenase (COX-2) and, to some degree, of transforming growth factor (TGF)-β, upon natural infection with T. bryosalmonae [8, 17]. The quality of the immune reaction to T. bryosalmonae is temperature-dependent, resulting in either a predominant Th2-like immune response with abundant B cell response at 15 °C or predominant Th1-like immune response with upregulation of the natural killer cell enhancement factor (NKEF) at 12 °C [18].
Whirling disease (WD) is a highly debilitating disease of salmonids that is caused by the myxozoan parasite Myxobolus cerebralis [19, 20]. M. cerebralis alternates between two hosts, an invertebrate oligochaetae host (Tubifex tubifex) and a vertebrate salmonid host, to complete a complex life-cycle [21, 22]. Mature myxospores that are formed within the fish cartilage can infect T. tubifex and once inside, T. tubifex triactinomyxons (TAMs) are formed and released into the water, thus infecting salmonids [23, 24]. WD is implicated in the decline of wild trout populations in North America [22]. The severity of WD depends largely on the age and size of the affected fish, with higher mortality rates in fingerlings, up to 90% of the infected populations [22].
The binding of a wide array of cytokines and growth factors to their cell receptors activates the associated Janus kinase (JAK) proteins and subsequently phosphorylates the cytokine receptor complex. Thereafter, the binding of signal transducer and activator of transcription (STAT) proteins to the site of the activated receptor and their phosphorylation occurs. Activated STAT proteins translocate to cell nucleus for signal transduction and initiate the gene transcription [25,26,27]. The suppressors of cytokine signaling (SOCS) are a group of intracellular molecules that act as strong negative regulators of the cytokine signaling through inhibition of the JAK-STAT pathway [28]. Even in Teleost fish, SOCS molecules are known to play an important role during the development of innate and acquired immunity [29], with SOCS-1 and SOCS-3 being selectively induced during PKD [8, 30].
Even though co-infections occur frequently in the aquatic environment, the research on such a subject is still in the infancy, with most of the studies often being conducted on single infections [31,32,33]. During co-infections, the host immune response induced by one pathogen can alter the pathogenesis of the secondary infections through the suppression or stimulation of the immune system [31,32,33]. The interaction between different parasites could be either synergistic or antagonistic in the infected host [32, 33]. Mixed infection of five myxozoan parasite species: T. bryosalmonae, Sphaerospora truttae, Chloromyxum schurovi, C. truttae, and Myxobolus sp. has been examined in farmed brown trout (Salmo trutta) [34]. A big knowledge gap still exists in the understanding of immune response mechanism in salmonids, during co-infections. Consequently, a substantial need for investigating the interactions between heterogeneous micro-organisms during co-infections, is there [35].
Recently, the impact of T. bryosalmonae and M. cerebralis co-infection on pathology of the target organs of rainbow trout has been examined [36]. The present study was designed to examine the expression of immune genes in rainbow trout when co-infected with two myxozoan parasites, by measuring the transcription levels of JAK/STAT signaling induced genes and SOCS genes in the posterior kidney and cranial cartilages (the target tissues of PKD or WD pathogenesis), respectively.
Methods
Fish and the experiment design
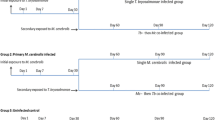
The experimental challenges, consisting of single infection and co-infection with parasites, were performed as described previously in the first part of this study and the focus was on the pathological assessment (Fig. 1) [36]. Briefly, pathogen-free rainbow trout (mean length 4.02 ± 0.26 cm, mean weight 0.6 ± 0.15 g) were divided into three groups of 96 each: the first group was infected with T. bryosalmonae spores, according to Kumar et al. [37], while the second group was infected with M. cerebralis TAMs, according to Hedrick et al. [38], and the third group was kept as uninfected control. Thirty days later, half of the fish from the first two infected groups were reciprocally co-infected with these parasites. At 30, 60, 90 and 120 days post-exposure (dpe), the fish were euthanized using an overdose of tricaine methanesulfonate (500 mg/l, MS-222, Sigma-Aldrich, Steinheim, Germany) and posterior kidneys and crania were dissected out and preserved in RNAlater (Sigma-Aldrich).

The experimental design. Three primary groups of rainbow trout: T. bryosalmonae-infected, M. cerebralis-infected, and uninfected control. Primary infected fish were exposed to M. cerebralis and T. bryosalmonae at 30 dpe, thereafter fish were sampled at the specified time points (adapted from Kotob et al. [36])
RNA extraction and cDNA synthesis
Total RNA was extracted from the posterior kidneys and cranial samples of each group (n = 4) at each time point, starting from 30 dpe (time of co-infection), using RNeasy Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. An on-column DNase digestion step was also included in order to remove any residual DNA contamination. RNA concentration was determined using a Nanodrop 2000c spectrophotometer (Thermo Fisher Scientific, Wilmington, USA) and one microgram of total RNA was used to synthesize cDNA, with iScript cDNA Synthesis Kit (Bio-Rad, Hercules, USA).
Reverse transcription quantitative PCR (RT-qPCR)
Parasite load determination (respectively for T. bryosalmonae and M. cerebralis) and quantification of the expression of SOCS-1, SOCS-3, JAK-1 and STAT-3 genes, were performed using samples from posterior kidneys and the crania of infected fish, following the previously described methodology [8, 39,40,41]. Primers used for the gene transcription assessment are summarized in Table 1. The quantity of gene expression level was measured with CFX96 Touch Real-Time PCR detection system (Bio-Rad). The PCR reaction of 20 μl final volume contained 4 μl of 1:10-fold diluted cDNA, 1× SsoAdvanced Universal SYBR Green Supermix (Bio-Rad), 0.4 μM of each primer, and DEPC-treated sterile distilled water (Bio-Rad). The PCR reaction consisted of an initial 5 min of cDNA denaturation at 95 °C, followed by 35 cycles of 95 °C for 30 s, 57–62 °C for 30 s and 72 °C for 30 s. A melting-point curve was measured, starting from 57 °C with an increase of 0.5 °C at every 10 s up to 95 °C, for detecting non-specific binding. Elongation factor 1 alpha [42] was used as a reference gene for normalizing the expression of targeted genes and calculation of relative gene expression was done using CFX manager software version 3.1 (Bio-Rad).
Statistical analysis
The differences in the expression of genes that were tested at different time points for each group were analyzed by a general linear model with repeated measurements and Sidak’s procedure was used for multiple comparison. One-way ANOVA with Tukey’s α-correction was used to examine the differences of expressions between all groups at each time point. Pearson’s product-moment correlation coefficient (r) was measured to determine the correlation between relative expressions of all tested immune genes. The statistical differences were considered significant at a P-value < 0.05 and all the data were analyzed in IBM SPSS software version 24.
Results
Tetracapsuloides bryosalmonae and Myxobolus cerebralis parasite burden
The relative expression of 60S ribosomal protein L18 of T. bryosalmonae (RPL18) and 18S rRNA genes of M. cerebralis in the posterior kidneys and the crania, respectively, showed an increasing burden during the pathogenesis progression in single and co-infections (Fig. 2). Tb burden reached significantly higher levels in the Mc-then-Tb co-infected group, in the posterior kidneys during 60 and 90 days post secondary exposure to T. bryosalmonae (F(2, 6) = 5.969, P < 0.05; F(2, 6) = 8.750, P < 0.05) (Fig. 2a). Mc burden in the Tb-then-Mc co-infected group was significantly lower than the single Mc-infected group at 30 dpe to Mc (F(1, 4) = 8.240, P = 0.045), then Mc burden increased at 60 dpe to Mc in all Mc-infected groups and reached significantly higher levels in Mc-then-Tb co-infected group at 90 and 120 dpe to Mc (F(2, 6) = 64.105, P < 0.01; F(1, 4) = 18.489, P = 0.013, respectively) (Fig. 2b).

Parasite burden quantification in the posterior kidneys and cranial cartilages of infected rainbow trout. a T. bryosalmonae RPL18 expression in posterior kidneys of T. bryosalmonae-infected rainbow trout, b M. cerebralis 18S rRNA expression in cranial cartilages of M. cerebralis-infected rainbow trout. Bars indicate standard deviation (n = 4)
Immune gene expression in the posterior kidneys during single and co-infections
Kidneys of the single Mc-infected group displayed no significant differences in the expression of SOCS-1, JAK-1 and STAT-3 genes at all the time points, when compared to the uninfected control (F(1, 4) = 2.670, P > 0.05). However, only SOCS-3 gene expression increased from 90 dpe and showed a significant increase at 120 dpe (F(1, 4) = 158.623, P = 0.001), indicating the stimulation of systemic immune response during the late stages of WD (Figs. 3, 4).

Relative gene expression of SOCS-1 (a) and SOCS-3 (b) in posterior kidneys during single and co-infections. RT-qPCR data from different time points were normalized to EF-1α expression and relative gene expression data were statistically analyzed. Bars indicate standard deviation (n = 4). Time points were calculated from the primary exposures to T. bryosalmonae and M. cerebralis

Relative gene expression of JAK-1 (a) and STAT-3 (b) in posterior kidneys during single and co-infections. Details are the same as in Fig. 3
The expression of SOCS-1, SOCS-3 and JAK-1 genes started to increase in the single Tb-infected group compared to the single Mc-infected and uninfected control groups at 30 dpe (F(2, 6) = 46.157, P < 0.001 for SOCS-1; F(2, 6) = 108.095, P < 0.001 for SOCS-3 and F(2, 6) = 8.116, P < 0.05 for JAK-1). No significant difference in the transcription of STAT-3 gene was observed between all the groups at this stage (F(2,6) = 2.471, P = 0.165) (Figs. 3, 4).
At 60 dpe, SOCS-1 gene expression was upregulated in all Tb-infected groups which was significant in the single Tb-infected group compared to single Mc-infected and uninfected control groups (F(2, 6) = 25.766, P = 0.001), and other co-infected groups (F(2, 6) = 5.486, P = 0.044). When all the groups were compared, the upregulation of SOCS-3 was significant in the single Tb and Tb-then-Mc co-infected groups (F(4, 10) = 27.941, P < 0.01) and upregulation of JAK-1 and STAT-3 gene expression was significant in all Tb-infected groups (F(4, 10) = 56.291, P < 0.01 for JAK-1 and F(4, 10) = 58.690, P < 0.01 for STAT-3) (Figs. 3, 4).
At 90 dpe, increased gene expression of SOCS-1, SOCS-3, JAK-1 and STAT-3 transcriptions was seen in all the Tb-infected groups, especially in the Mc-then-Tb co-infected group, compared to the uninfected control and single Mc-infected groups (F(4, 10) = 21.612, P < 0.001 for SOCS-1; F(4, 10) = 38.323, P < 0.001 for SOCS-3; F(4, 10) = 22.507, P < 0.001 for JAK-1 and F(4, 10) = 20.744, P < 0.001 for STAT-3) (Figs. 3, 4).
At 120 dpe, all the Tb-infected groups showed significant upregulation of SOCS-1 and SOCS-3 gene expression when compared with the uninfected control group (F(4, 10) = 24.981, P < 0.05 for SOCS-1 and F(4, 10) = 27.407, P < 0.01 for SOCS-3). Moreover, gene expression of JAK-1 and STAT-3 genes were upregulated in the single Tb-infected and Tb-then-Mc co-infected groups (F(4, 10) = 15.852, P < 0.01 for JAK-1 and F(4, 10) = 4.417 P < 0.05 for STAT-3) compared to other groups (Figs. 3, 4).
The correlation of tested immune genes was positively strong between SOCS-1 and JAK-1 in the single Tb-infected group (r = 0.619, P = 0.032) (see Additional file 1: Table S1). The correlation between the expression of SOCS-1 and STAT-3 genes was also highly positive in the single Tb-infected group (r = 0.577 and P = 0.049). Interestingly, a strong and highly significant positive correlation was detected in expression of SOCS-1 and SOCS-3 genes in the single Tb-infected and Mc-then-Tb co-infected groups (r = 0.959 and P = 0.0001; r = 0.895 and P = 0.001, respectively). SOCS-3 correlated positively with JAK-1 and this correlation was highly significant in the single Mc-infected group (r = 0.785 and P = 0.002). A weak positive correlation was found between SOCS-3 and STAT-3 in all groups except in the single Mc-infected group, where the correlation was strong (r = 0.722 and P = 0.008) and the expression of both genes was found to be high at 90 and 120 dpe. Strong positive correlation was also found between JAK-1 and STAT-3 in single Tb and Mc-infected groups (r = 0.779 and P = 0.003; r = 0.775 and P = 0.003, respectively) (see Additional file 1).
Immune gene expression in the crania during single and co-infections
The transcription of SOCS-1 and SOCS-3 genes was significantly higher between 60–120 dpe (F(3, 8) = 92.682, P < 0.001 at 60 dpe; F(3, 8) = 9.596, P < 0.01 at 90 dpe and F(3, 8) = 36.842, P < 0.001 at 120 dpe) (Fig. 5a) and at 120 dpe in the single Mc and Mc-then-Tb co-infected groups (F(3, 8) = 39.441, P < 0.01) (Fig. 5b), respectively.

Relative gene expression of SOCS-1 (a) and SOCS-3 (b) in cranial cartilages during single and co-infections. Details are the same as in Fig. 3
In the case of JAK-1 and STAT-3 gene expression, a remarkable increase was observed in the single Mc-infected group at 30 dpe (F(1, 4) = 713.629, P < 0.001 for JAK-1 and F(1, 4) = 1489.910, P < 0.0001). The transcription of JAK-1 gene was significantly increased between 60–120 dpe in the single Mc and Mc-then-Tb co-infected groups compared to other groups (F(3, 8) = 36.765, P < 0.001 at 60 dpe; F(3, 8) = 54.622, P < 0.001 at 90 dpe and F(3, 8) = 16.834, P < 0.01 at 120 dpe) (Fig. 6a). However, statistically significant expression of STAT-3 was only observed in the single Mc-infected group which continued at 60 dpe (F(3, 8) = 29.919, P < 0.001) (Fig. 6b).

Relative gene expression of JAK-1 (a) and STAT-3 (b) in cranial cartilages during single and co-infections. Details are the same as in Fig. 3
Since there was a low level of expression of all immune genes in the cranial cartilages, we did not observe any correlation between the immune genes expression.
Discussion
Myxozoan parasites can modulate the pro-inflammatory cellular responses including phagocytosis, oxidative phagocytic activity and complement activity [43]. Additionally, lysozyme, peroxidases, and acute-phase proteins, as the players of humoral immune mechanisms, are also involved in the immune response against those parasites [8, 43]. In this study, we have investigated, for the first time, the immune response of rainbow trout during single and co-infections between T. bryosalmonae and M. cerebralis, focusing on selective genes that are involved in the SOCS/JAK/STAT signaling pathway. We also determined the correlations of gene expression between these immune-related genes. The data obtained from this study demonstrate that the posterior kidneys of all T. bryosalmonae-infected fish had elevated levels of tested immune genes and that the pathway was highly induced during the pathogenesis of PKD, which was present either as a single disease or in concomitance with WD. However, the relative gene expressions of SOCS-1 and SOCS-3 were much higher than JAK-1 and STAT-3. Additionally, the highest load of both parasites was detected in fish co-infected initially with M. cerebralis and then with T. bryosalmonae at 90 dpe. The kidneys of fish from this group showed increased expressions of tested immune genes at 90 dpe and their crania also showed the highest gene expression levels of SOCS-1 and SOCS-3 at 120 dpe.
The transcription level of the T. bryosalmonae RPL18 gene in cDNAs was measured to assess the relative burden of the parasite T. bryosalmonae in posterior kidneys [8]. The single Tb-infected and Tb-then-Mc co-infected groups showed a decrease in the gene expression of T. bryosalmonae RPL18 at 90 and 120 dpe, suggesting the activation of immune system and parasite clearance during this time. However, this was not in agreement with the immunohistochemistry (IHC) results obtained by counting the number of parasites [36]. The disparity between both the results could be because of the fact that the IHC data reflected both dead and viable parasites while the data obtained by T. bryosalmonae RPL18 gene expression reflected only viable parasites in the samples. Nevertheless, the relative gene expression of T. bryosalmonae RPL18 in Mc-then-Tb co-infected group persisted to increase, indicating active parasite proliferation and the suppression of immune response. Furthermore, the primary infection of rainbow trout with M. cerebralis and then with T. bryosalmonae fostered the pathogenesis of both infections and this co-infected group exhibited high parasitic burden of both parasites (Fig. 2a, b). These data were in accordance with the counting of T. bryosalomae stages in the kidney of infected fish [36]. The synergistic interaction that occurred in this case of co-infection might be due to the immunosuppressive effect of secondary exposure to T. bryosalmonae, which was 30 days after primary M. cerebralis exposure [8, 17, 44]. PKD-mediated immunosuppression occurs due to the downregulation of some key regulatory genes and significant decrease in phagocytic and respiratory burst activity of kidney macrophages [44]. During the course of primary infection with M. cerebralis, the spores successfully parasitize and multiply in the cranium and evade the immune system before the secondary exposure to T. bryosalomonae, because of which the infected fish exhibits exacerbated form of clinical signs and lesions of both diseases along with the highest levels of transcription of the specific genes of parasites load (T. byrosalmonae RPL18 and M. cerebralis 18S rRNA) [36]. On the contrary, the secondary infection with M. cerebralis counteracts the development and the load of M. cerebralis in the cranium of Tb-infected rainbow trout, evident from the lower parasitic load in this group (Fig. 2b). The decreased parasitic burden could be due to the cross-reactivity between the sporogonic stages of both parasites that might induce cross-immunity and thereby interfering with the pathogenesis and parasitic burden of secondary M. cerebralis [45].
The pathogenesis of PKD is characterized by an anti-inflammatory reaction, T helper cell-like activity and an intense B cell/antibody response with a marked upregulation of interleukin (IL)-6, IL-10, IL-11 and antimicrobial peptides [8]. In our study, we found that the expression of SOCS-1 and SOCS-3 genes was higher during the progression of PKD pathogenesis, further reaffirming the fact that the parasite causes an immunosuppression reaction in host in order to evade its immune system. This agrees with previous studies that have demonstrated specifically that the expression of SOCS-1 and SOCS-3 genes strongly correlate with the parasitic burden and pathology progression [8, 30]. In addition, one of the most striking findings was the presence of increased SOCS-1 and SOCS-3 gene transcription in the Mc-then-Tb co-infected group at 90 dpe that correlated positively with the kidney swelling index and pathological lesions of kidneys from this group [36]. This could be attributed to the presence of synergistic interaction during the period of co-infection [36]. It has been demonstrated earlier that SOCS-1 has a negative regulatory effect on interferon (IFN) mediated JAK-STAT signaling in fish, and that there exists a direct negative interaction between SOCS-1 and STAT-1 and between SOCS-1 and Tyrosine kinase 2 (Tyk2) [28]. SOCS-1 blocks the differentiation of Th1 subset through inhibition of IFN-γ-STAT-1 and IL-12-STAT-4 pathways and hence its deficiency can lead to constitutive expression of IFN-γ and STAT-1 inducing Th1 differentiation, preventing Th17 differentiation [46]. SOCS-1 and SOCS-3 have a negative regulatory role in IFN-γ signaling within human keratinocytes and their overexpression inhibits IFN-γ-induced phosphorylation of IFN-γRα and activation of STAT-1 and STAT-3, leading to impaired IFN-γ-dependent inflammatory responses [47]. The pathogenesis of PKD in rainbow trout is strongly inducing the transcription of Type II IFN-γ [8]. Despite the upregulation of all immune tested genes observed in this study, we found that the relative gene expressions of SOCS-1 and SOCS-3 in the posterior kidney were much higher than JAK-1 and STAT-3. Kumar et al. [42] found that the gene expression of transforming protein RhoA, which regulates the signal transduction pathway of a wide range of cellular processes, was upregulated in the kidney of brown trout infected with T. bryosalmonae and suggested that differential modulation of genes may support the parasite development in fish hosts. In our experiment, we studied the induced expressions of the tested JAK-STAT genes in the trout kidneys, during T. bryosalmonae infection and found that the gene expressions of SOCS-1 and SOCS-3 were most prominent, which indicated their support for parasite development in the fish host.
It has been reported previously that innate immune response genes namely, IFN-γ, interferon regulatory factor 1 (IRF-1) and inducible nitric oxide synthase (iNOS) in addition to Ubiquitin-like protein 1 were significantly upregulated both in susceptible as well as resistant rainbow trout strains after exposure to M. cerebralis [41]. However, STAT-3 and metallothionein B were consistently upregulated in the resistant Hofer strain and remained unchanged in the susceptible TL trout strain following M. cerebralis infection [41, 48]. In the present study, we found the expressions of the different immune genes to be significantly lower in the cranial cartilage than in the kidney, a main hematopoietic organ of fish. Therefore, the kidney is characterized as an organ that elicits strong immune response against the invading pathogens. The transcription of SOCS-1 gene was found to be upregulated in the cranial cartilages from 60 dpe to 120 dpe. The highest level of SOCS-1 and SOCS-3 gene expression in the cranium and the highest load of M. cerebralis were detected in Mc-then-Tb co-infected group at 120 dpe, indicating an association between expression of SOCS genes and disease progression and severity. This is in accordance with observations from the first part of this study wherein synergistic effects were elicited during Mc-then-Tb co-infection along with exacerbated pathological lesions of PKD and WD [36]. The elevated expression of JAK-1 gene in the cranium of single Mc-infected and Mc-then-Tb co-infected group at 60 dpe (Fig. 6), lowered with the course of time. This expression reciprocally correlated with increased expression of SOCS genes in the Mc-then-Tb co-infected group with the course of time (Fig. 5). Therefore, we suggest that M. cerebralis modulates the immune gene expression to overcome the host cellular response. The results obtained from this study will help in understanding the host-pathogen interaction during single and co-infections with PKD and WD.
Conclusions
In this study, we highlight the SOCS/JAK/STAT signaling pathway and its role in co-infections with two myxozoan parasites. Rainbow trout infected with M. cerebralis and then subsequently with T. bryosalmonae showed the highest loads of both parasites in the posterior kidneys and cranial cartilages, thereby indicating a synergistic interaction. This study showed differential immunomodulation of SOCS genes and post-receptor JAK/STAT induced genes during myxozoans co-infection when compared to single infection. Our results suggest that T. bryosalmonae and M. cerebralis alter the JAK/STAT signaling pathway via a strong overexpression of SOCS-1 and SOCS-3 genes. However, further studies are required to fully understand the early innate immune response of fish during myxozoans co-infections.
Abbreviations
- dpe:
-
Days post-exposure
- Mc :
-
Myxobolus cerebralis
- PKD:
-
Proliferative kidney disease
- RT-qPCR:
-
Reverse transcription quantitative PCR
- TAMs:
-
Triactinomyxons
- Tb :
-
Tetracapsuloides bryosalmonae
- WD:
-
Whirling disease
References
Canning EU, Curry A, Feist SW, Longshaw M, Okamura B. Tetracapsula bryosalmonae n. sp. for PKX organism, the cause of PKD in salmonid fish. Bull Eur Ass Fish Pathol. 1999;19:203–6.
Canning EU, Curry A, Feist SW, Longshaw M, Okamura B. A new class and order of myxozoans to accommodate parasites of bryozoans with ultrastructural observations on Tetracapsula bryosalmonae (PKX organism). J Eukaryot Microbiol. 2000;47:456–68.
El-Matbouli M, Hoffmann RW. Influence of water quality on the outbreak of proliferative kidney disease-field studies and exposure experiments. J Fish Dis. 2002;25:459–67.
Grabner DS, El-Matbouli M. Transmission of Tetracapsuloides bryosalmonae (Myxozoa: Malacosporea) to Fredericella sultana (Bryozoa: Phylactolaemata) by various fish species. Dis Aquat Organ. 2008;79:133–9.
Okamura B, Hartikainen H, Schmidt-Posthaus H, Wahli T. Life cycle complexity, environmental change and the emerging status of salmonid proliferative kidney disease. Freshwater Biol. 2011;56:735–53.
Morris DJ, Adams A. Transmission of Tetracapsuloides bryosalmonae (Myxozoa: Malacosporea), the causative organism of salmonid proliferative kidney disease, to the freshwater bryozoan Fredericella sultana. Parasitology. 2006;133:701–9.
Hedrick RP, MacConnell E, De Kinkelin P. Proliferative kidney disease of salmonid fish. Ann Rev Fish Dis. 1993;3:277–90.
Gorgoglione B, Wang T, Secombes CJ, Holland JW. Immune gene expression profiling of proliferative kidney disease in rainbow trout Oncorhynchus mykiss reveals a dominance of anti-inflammatory, antibody and T helper cell-like activities. Vet Res. 2013;44:55.
Soliman H, Kumar G, El-Matbouli M. Tetracapsuloides bryosalmonae persists in brown trout Salmo trutta for five years post exposure. Dis Aquat Organ. 2018;127:151–6.
Gorgoglione B, Kotob MH, El-Matbouli M. Migrating zooids allow the dispersal of Fredericella sultana (Bryozoa) to escape from unfavourable conditions and further spreading of Tetracapsuloides bryosalmonae. J Invertebr Pathol. 2016;140:97–102.
Abd-Elfattah A, El-Matbouli M, Kumar G. Structural integrity and viability of Fredericella sultana statoblasts infected with Tetracapsuloides bryosalmonae (Myxozoa) under diverse treatment conditions. Vet Res. 2017;48:19.
Clifton-Hadley RS, Bucke D, Richards RH. Economic importance of proliferative kidney disease in salmonid fish in England and Wales. Vet Rec. 1986;119:305–6.
Gorgoglione B, Kotob MH, Unfer G, El-Matbouli M. First Proliferative Kidney Disease outbreak in Austria, linking to the aetiology of Black Trout Syndrome threatening autochthonous trout populations. Dis Aquat Organ. 2016;119:117–28.
Bailey C, Segner H, Wahli T. What goes around comes around: an investigation of resistance to proliferative kidney disease in rainbow trout Oncorhynchus mykiss (Walbaum) following experimental re-exposure. J Fish Dis. 2017;11:1599–612.
Olesen NJ, Vestergard-Jørgensen PE. Quantification of serum immunoglobulin in rainbow trout Salmo gairdnieri under various environmental conditions. Dis Aquat Org. 1986;1:183–9.
Kumar G, Abd-Elfattah A, El-Matbouli M. Identification of differentially expressed genes of brown trout (Salmo trutta) and rainbow trout (Oncorhynchus mykiss) in response to Tetracapsuloides bryosalmonae (Myxozoa). Parasitol Res. 2015;114:929–39.
Holland JW, Gould CRW, Jones CS, Noble LR, Secombes CJ. The expression of immune-regulatory genes in rainbow trout, Oncorhynchus mykiss, during a natural outbreak of proliferative kidney disease (PKD). Parasitology. 2003;126:95–102.
Bailey C, Segner H, Casanova-Nakayama A, Wahli T. Who needs the hotspot? The effect of temperature on the fish host immune response to Tetracapsuloides bryosalmonae the causative agent of proliferative kidney disease. Fish Shellfish Immunol. 2017;63:424–37.
Hofer B. Über die Drehkrankheit der Regenbogenforelle. Allg Fisch Ztg. 1903;28:7–8.
Hoffman GL. Myxobolus cerebralis, a worldwide cause of salmonid whirling disease. J Aquat Anim Health. 1990;2:30–7.
Wolf K, Markiw ME. Biology contravenes taxonomy in the Myxozoa: new discoveries show alternation of invertebrate and vertebrate hosts. Science. 1984;225:1449–52.
Hedrick RP, El-Matbouli M, Adkinso MA, MacConnell E. Whirling disease: re-emergence among wild trout. Immunol Rev. 1998;166:365–76.
El–Matbouli M, Hoffmann RW, Mandok C. Light and electron microscopic observations on the route of the triactinomyxon-sporoplasm of Myxobolus cerebralis from epidermis into rainbow trout cartilage. J Fish Biol. 1995;46:919–35.
El-Matbouli M, Hoffmann RW. Light and electron microscopic studies on the chronological development of Myxobolus cerebralis to the actinosporean stage in Tubifex tubifex. Int J Parasitol. 1998;28:195–217.
Yoshimura A, Naka T, Kubo M. SOCS proteins, cytokine signalling and immune regulation. Nat Rev Immunol. 2007;7:454–65.
O’Shea JJ, Gadina M, Schreiber RD. Cytokine signaling in 2002: new surprises in the Jak/Stat pathway. Cell. 2002;109:S121–31.
Ilangumaran S, Ramanathan S, Rottapel R. Regulation of the immune system by SOCS family adaptor proteins. Semin Immunol. 2004;16:351–65.
Skjesol A, Liebe T, Iliev DB, Thomassen EI, Tollersrud LG, Sobhkhez M, et al. Functional conservation of suppressors of cytokine signaling proteins between teleosts and mammals: Atlantic salmon SOCS1 binds to JAK/STAT family members and suppresses type I and II IFN signaling. Dev Comp Immunol. 2014;45:177–89.
Wang T, Secombes CJ. Rainbow trout suppressor of cytokine signalling (SOCS)-1, 2 and 3: Molecular identification, expression and modulation. Mol Immunol. 2008;45:1449–57.
Wang T, Gorgoglione B, Maehr T, Holland JW, Vecino JLG, Wadsworth S, et al. Fish suppressors of cytokine signaling (SOCS): gene discovery, modulation of expression and function. J Signal Transduct. 2011;2011:905813.
Lello J, Boag B, Fenton A, Stevenson IR, Hudson PJ. Competition and mutualism among the gut helminths of a mammalian host. Nature. 2004;428:840–4.
Telfer S, Birtles R, Bennett M, Lambin X, Paterson S, Begon M. Parasite interactions in natural populations: insights from longitudinal data. Parasitology. 2008;135:767–81.
Kotob MH, Menanteau-Ledouble S, Kumar G, Abdelzaher M, El-Matbouli M. The impact of co-infections on fish: a review. Vet Res. 2016;47:98.
Holzer AS, Sommerville C, Wootten R. Molecular studies on the seasonal occurrence and development of five myxozoans in farmed Salmo trutta L. Parasitology. 2006;132:193–205.
Johnson PT, Hoverman JT. Parasite diversity and coinfection determine pathogen infection success and host fitness. Proc Natl Acad Sci USA. 2012;109:9006–11.
Kotob MH, Gorgoglione B, Kumar G, Abdelzaher M, Saleh M, El-Matbouli M. The impact of Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections on pathology in rainbow trout. Parasit Vectors. 2017;10:442.
Kumar G, Abd-Elfattah A, Saleh M, El-Matbouli M. Fate of Tetracapsuloides bryosalmonae (Myxozoa) after infection of brown trout (Salmo trutta) and rainbow trout (Oncorhynchus mykiss). Dis Aquat Org. 2013;107:9–18.
Hedrick RP, McDowell TS, Gay M, Marty GD, Georgiadis MP, MacConnell E. Comparative susceptibility of rainbow trout Oncorhynchus mykiss and brown trout Salmo trutta to Myxobolus cerebralis, the cause of salmonid whirling disease. Dis Aquat Org. 1999;37:173–83.
Kelley GO, Zagmutt-Vergara FJ, Leutenegger CM, Adkison MA, Baxa DV, Hedrick RP. Identification of a serine protease gene expressed by Myxobolus cerebralis during development in rainbow trout Oncorhynchus mykiss. Dis Aquat Organ. 2004;59:235–48.
Djordjevic B, Škugor S, Jørgensen SM, Øverland M, Mydland LT, Krasnov A. Modulation of splenic immune responses to bacterial lipopolysaccharide in rainbow trout (Oncorhynchus mykiss) fed lentinan, a beta-glucan from mushroom Lentinula edodes. Fish Shellfish Immunol. 2009;26:201–9.
Baerwald MR. Temporal expression patterns of rainbow trout immune-related genes in response to Myxobolus cerebralis exposure. Fish Shellfish Immunol. 2013;35:965–71.
Kumar G, Abd-Elfattah A, El-Matbouli M. Differential modulation of host genes in the kidney of brown trout Salmo trutta during sporogenesis of Tetracapsuloides bryosalmonae (Myxozoa). Vet Res. 2014;45:101.
Alvarez-Pellitero P. Fish immunity and parasite infections: from innate immunity to immunoprophylactic prospects. Vet Immunol Immunopathol. 2008;126:171–98.
Chilmonczyk S, Monge D, De Kinkelin P. Proliferative kidney disease: cellular aspects of the rainbow trout, Oncorhynchus mykiss (Walbaum), response to parasitic infection. J Fish Dis. 2002;25:217–26.
Morris DJ, Molnar K, Longshaw M, Adams A. Immunostaining of spores and plasmodia of disparate myxozoan genera with comments on the properties of the sporular mucus envelope. Parasitology. 2006;132:781–90.
Tanaka K, Ichiyama K, Hashimoto M, Yoshida H, Takimoto T, Takaesu G, et al. Loss of suppressor of cytokine signaling 1 in helper T cells leads to defective Th17 differentiation by enhancing antagonistic effects of IFN-γ on STAT3 and Smads. J Immunol. 2008;180:3746–56.
Federici M, Giustizieri ML, Scarponi C, Girolomoni G, Albanesi C. Impaired IFN-γ-dependent inflammatory responses in human keratinocytes overexpressing the suppressor of cytokine signaling 1. J Immunol. 2002;169:434–42.
Baerwald MR, Welsh AB, Hedrick RP, May B. Discovery of genes implicated in whirling disease infection and resistance in rainbow trout using genome-wide expression profiling. BMC Genomics. 2008;9:37.
Acknowledgments
We are thankful to the ministry of higher education in Egypt for offering a PhD scholarship to MHK. We are grateful to the whole team of the Clinical Division of Fish Medicine, University of Veterinary Medicine Vienna for their help during this study.
Funding
This study was funded in part by the University of Veterinary Medicine and the Austrian Science Fund (FWF) project no. P29294-B25.
Availability of data and materials
All data supporting the findings of this study are presented in the main text and its Additional file.
Author information
Authors and Affiliations
Contributions
MEM and BG designed the research plan. MEM and GK supervised this study. MHK performed the gene expression screening. MHK and GK analyzed the data. MHK, GK and MS wrote the manuscript. MEM, BG and MA revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was approved by the institutional ethics committee of the University of Veterinary Medicine, Vienna and the national authority, and conducted according to §26 of the Austrian Law for Animal Experiments, Tierversuchsgesetz 2012–TVG 2012 under permit no. GZ 68.205/0141-WF/V/3b/2015.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Table S1 The correlation of coefficient among different kidney immune genes expression in single and co-infected groups. (DOCX 14 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Kotob, M.H., Kumar, G., Saleh, M. et al. Differential modulation of host immune genes in the kidney and cranium of the rainbow trout (Oncorhynchus mykiss) in response to Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections. Parasites Vectors 11, 326 (2018). https://doi.org/10.1186/s13071-018-2912-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-018-2912-7