
Abstract
Background
Anopheles anthropophagus is one of the major vectors of malaria in Asia. MicroRNAs (miRNAs) play important roles in cell development and differentiation as well as in the cellular response to stress and infection. In a former study, we have investigated the global miRNA profiles in relation to sex in An. anthropophagus. However, the miRNAs contributing to the blood-feeding and infection with Plasmodium are still unknown.
Methods
High-throughput sequencing was performed to identify miRNA profiles of An. anthropophagus midguts after blood-feeding and Plasmodium infection. The expression patterns of miRNA in different midgut libraries were compared based on transcripts per million reads (TPM), and further confirmed by Northern blots. Target prediction and pathway analysis were carried out to investigate the role of regulated miRNAs in blood-feeding and Plasmodium infection.
Results
We identified 67 known and 21 novel miRNAs in all three libraries (sugar-feeding, blood-feeding and Plasmodium infection) in An. anthropophagus midguts. Comparing with the sugar-feeding, the experssion of nine (6 known and 3 novel) and ten (9 known and 1 novel) miRNAs were significantly upregulated and downregulated respectively after blood-feeding (P < 0.05, fold change ≥ 2 and TPM ≥ 10). Plasmodium infection induced the expression of thirteen (9 known and 4 novel) and eleven (9 known and 2 novel) miRNAs significantly upregulated and downregulated, respectively, compared with blood-feeding. The representative upregulated miR-92a in blood-feeding and downregulated miR-275 in Plasmodium infection were further confirmed by Northern Blot. Putative targets of these regulated miRNAs were further investigated and classified into their pathways.
Conclusions
This study suggests that miRNAs are involved in the blood-feeding and Plasmodium infection in An. anthropophagus midgut. Further studies of the function of these differential expressed miRNAs will facilitate in better understanding of mosquito biology and anti-parasite immunity.
Similar content being viewed by others


Background
Malaria, caused by Plasmodium parasites, is a major threat to human health worldwide. According to the reports from WHO, there were 214 million new infection cases of malaria, and about 438,000 malaria deaths worldwide in 2015 [1]. Plasmodium spp. are transmitted to humans by the blood-feeding of Anopheles mosquitoes. To accomplish the life-cycle, the Plasmodium must overcome numerous attacks from the innate immunity system in mosquitoes [2, 3]. Mosquitoes also have developed various mechanisms to confront Plasmodium infection [3, 4]. The midgut represents the first barrier for the survival and development of Plasmodium [4–6]. A key concept that has emerged from recent studies is the molecular mechanisms of mosquito midgut that negatively and positively modulate the invasion of the parasite [5, 6].
MicroRNAs (miRNAs) are 18–24 nucleotides non-coding RNAs that regulate gene expression at the post-transcriptional level [7]. They are now considered as a key mechanism of gene regulation in many cellular processes including development, differentiation, apoptosis and innate immunity [8–10]. miRNAs are also involved in the physiological functions of mosquitoes, such as sexual differences and blood-feeding, even in the control of viral and parasitic infections [11–16]. Until now, the miRNA profiles of midguts from Anopheles gambiae and Anopheles stephensi have been reported, and several miRNAs expression levels were shown to be altered during the response to blood-feeding and Plasmodium infection [13, 15–18]. For example, blood-feeding and Plasmodium infection in An. stephensi revealed regulation of 13 and 16 miRNAs respectively [18]. Anopheles anthropophagus is a species of mosquito that sucks human blood and transmits malaria (Plasmodium vivax and Plasmodium falciparum) as well as Brugia malayi filariasis in Southeast Asia [12, 19, 20]. The role of miRNAs in An. anthropophagus during blood-feeding and Plasmodium infection are still unknown.
In this study, we employed small RNA sequencing to identify miRNA expression profiles from three samples of An. anthropophagus midguts: sugar-feeding, blood-feeding and Plasmodium infection. The differential expression of miRNAs were further analyzed by target prediction and pathway analysis to reveal their roles in blood-feeding and Plasmodium infection. Our results provide novel regulated miRNAs information of An. anthropophagus during blood-feeding and parasite infection. Understanding the functions of these regulated miRNAs will help investigate mosquito biology and control mosquito-borne infectious diseases.
Methods
Mosquito rearing and Plasmodium infection
Anopheles anthropophagus (China wild type strain) was reared and maintained in humidified incubators at 26 ± 1 °C on a 12 h light:dark cycle as described previously [12]. Mice (18–20 g male ICR mice) were used for mosquito blood-feeding and Plasmodium infection experiments. Stock solution of Plasmodium berghei ANKA strain (0.2 ml) were thawed and intraperitoneally (i.p.) injected into ICR mice by using a 1.0 ml tuberculin syringe. When the gametocytes were confirmed, naive 4–5 day-old female mosquitoes were fed on P. berghei-infected or uninfected ICR mice. Mosquitoes were kept at 26 ± 1 °C until dissection.
Mosquito dissections and total RNA isolation
Dissection of female adults midgut was performed 48 h post-blood-feeding on Plasmodium-infected or uninfected ICR mice. Adult female mosquitoes with 10% sugar-feeding were collected as control sample at 48 h. Dissections were performed on ice in RNAlater® Stabilization Solution (Ambion, Austin, U.S.A) and kept on ice. The midguts were dissected from the abdomen as described before [11]. Total RNAs were extracted from dissected tissue using mirVanaTMmiRNA Isolation Kit (Ambion). Quality and quantity of RNA was checked by using denaturalization agar gels and Du530 Spectrophotometer (Beckman, Krefeld, Gemany).
Small RNA sequencing
The small RNA samples were collected and subsequently sequenced by illumina Hiseq2000 as described before [12]. Briefly, the small RNAs were ligated with RNA adapter followed by reverse transcription using RT primers. Following PCR amplification of the adaptor enriched fragments, the PCR-amplified cDNAs were size-selected using electroelution to obtain the small RNA population with length 119–134 bp. These small RNA libraries were then sequenced using illumina Hiseq2000 (BGI, China). There were 3 biological replicates for each library sequencing.
Computational analysis of small RNA sequencing data
Raw reads generated by high-throughput sequencing was processed as previously described by us [11, 12] with slight modifications. First, low quality reads and reads smaller than 15 nucleotides (nt) were removed from the three small RNA read datasets of midguts, respectively. Clean reads derived from deep sequencing were trimmed and filtered with BOWTIE software to fetch sequences having an appropriate length (15–32 nt). Mature and pre-miRNA sequences of available mosquito species, i.e. Anopheles gambiae and Aedes aegypti, were used as a reference miRNAs database from miRBase v.21. Identification of the novel miRNA was performed by using RNA fold and miRDeep2 as described before [12]. The small RNA datas have been submitted to the NCBI/GEO database with the accession number GSE93545. Transcripts per million reads (TPM) for each miRNA in all three libraries was preformed to generate a comparative analysis of different midgut samples [17, 18]. A heatmap was generated by MeV software.
Northern blot
Northern hybridizations were conducted using digoxigenin-labeled miRCURY LNA probes (Exiqon, Vedbak, Denmark) as described before [12]. Briefly, the midguts of female mosquitoes were collected at 48 h after blood-feeding. The total RNA sample was extracted using mirVana™ miRNA Isolation Kit (Ambion). Total RNA (20 μg) was loaded in 15% denaturing polyacrylamide gels. The RNA gels were transferred to a nylon membrane (Ambion), crosslinked using a UV crosslinker, and prehybridized, then hybridized overnight in the ULTRAhyb-Oligo Hybridization Buffer (Ambion) with the appropriate DIG-labeled probe at 42 °C. After washed three times, the membranes were then incubated for 5 min in development buffer. Substrate (1:100 diluted in development buffer) was applied on to the membranes and incubated in dark for 10 min. Chemiluminescence signal was then measured to detect miRNA on the membrane. Antisense 5′ digoxigenin-labeled miRCURY LNA probes sequences as follow, aan-miR-92a: 5′-TCA GCC GCT GTC ACA CGC ACA G-3′; aan-miR-275: 5′-GAC CAA TCG CCG TCC CCG CCG-3′.
miRNA target prediction and pathway analysis
mRNA targets of regulated miRNAs during blood-feeding and Plasmodium infection were predicted according to the protocol reported before [18, 19]. Briefly, 3′UTR sequence of An. gambiae and Ae. aegypti were downloaded from VectorBase and subjected to target prediction using RNA hybrid tool. Target predictions were carried out based on the following three parameters: (i) the perfect complementarity of the miRNA with the 3′UTR sequence of mRNA targets; (ii) the energy of the miRNA:mRNA target duplex ≤ -20 Kcal/mol; (iii) P-value < 0.05. The selected miRNA:mRNA interaction networks were generated and visualized by Cytoscape.
Statistical analysis
Statistical tests for identifying significant differentially expressed miRNAs were performed using t-test. The P-value cut-off was carried out on the data with the significance selected as 0.05.
Results
Small RNA sequencing analysis
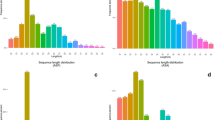
The small RNA libraries were enriched, and separately yielded 17.51 million, 15.87 million and 6.5 million raw reads from the midgut samples of sugar-feeding (SF), blood-feeding (BF) and Plasmodium infection (PI). After filtering for linker sequences, and removing ambiguous reads, high quality clean reads with sizes ranged from 15 to 32 nt were collected, of which 14.45 million (82.56%) reads for SF, 11.75 million (74.02%) reads for BF, and about 3.2 million (50.38%) reads for PI (Table 1). The main populations of small RNA in the length distribution is 20–23 nt (Fig. 1), of which 10.1 million (68.89%) reads for SF, 5.76 million for BF (49.01%) and 1.45 million for PI (45.51%). The 20–23 nt peaks of small RNA are consisted with the expected size of microRNAs. After aligned to the known miRNA and pre-miRNA in miRBase (version 21.0), the miRNA reads from SF, BF and PI midguts which can match to the miRBase are 4.03 million, 1.88 million and 0.21million separately (Table 1).

Length distribution of small RNA reads in the An. anthropophagus midguts. Female mosquitoes midguts at 48 h sugar feeding (SF), blood-feeding (BF) and Plasmodium infected-blood-feeding (PI)
Identification of miRNAs in midguts of An. anthropophagus
A total of 67 known and 21 novel miRNAs were identified in the SF, BF and PI midgut libraries (Tables 2 and 3). Both known and novel miRNAs were present in the miRNA databases of An. anthropophagus adult as described before [12]. The distribution and relative abundance of these miRNAs were analyzed according to the set of “abundant” (TPM ≥ 1000) or “rare” (TPM < 10). Most of known and novel miRNAs were found in the SF library (n = 87) with 29 abundant and 20 rare miRNAs. In case of BF library, 26 miRNAs were found to be abundant and 7 miRNAs (miR-193, miR-2944b, miR-307, miR-309, miR-79-5p, miR-965-1 and miR-N6) were not detected. Meanwhile, 27 abundant and 20 rare miRNAs were found in the PI library, of which 7 miRNAs (miR-193, miR-210, miR-252, miR-263b-5p, miR-2944b, miR-307 and miR-87) have no read counts. A novel miRNA miR-N6 is unique in the PI midguts.
Differential expression of miRNAs after blood-feeding and Plasmodium infection
To investigate the miRNAs regulated in blood-feeding and Plasmodium infection, the TPM value of individual miRNA between BF and SF, PI and BF were compared in the form of the fold change (Fig. 2). The set of significantly regulated miRNAs were selected on the basis of their P-value < 0.05, fold change ≥ 2 and TPM ≥10 (Additional file 1: Table S1 and Additional file 2: Table S2). With respect to BF library, six known miRNAs (miR-252, miR-315-5p, miR-92a, miR-92b, miR-932-3p and miR-989) and three novel miRNAs (miR-N10, miR-N16 and miR-N21) showed significantly upregulated expression compared with SF. Especially, the expression levels for miR-92a, miR-932-3p, miR-N16 and miR-N21 were enhanced more than 5-fold after blood-feeding. Meanwhile, the expression of 10 miRNAs (miR-1, miR-1175-3p, miR-12, miR-13, miR-276, miR-279, miR-283, miR-2b, miR-996 and miR-N3) was found to be significantly downregulated after blood-feeding (Tables 2 and 3).

Heatmap of differential expressed miRNAs in the An. anthropophagus midgut. Expression profile of known miRNAs (a) and novel miRNAs (b) in sugar-feeding midgut (SF), blood-feeding midgut (BF) and Plasmodium-infected midguts (PI). Colour scale-bar from light green to dark red indicates relative increase in miRNA expression
In the case of the PI library, 9 known miRNAs (miR-1, miR100, miR-12, miR-13, miR-263a, miR-276, miR-308-5p, miR-31, miR-iab-4) and 4 novel miRNAs (miR-N1, miR-N6, miR-N14 and miR-N20) exhibited significantly upregulated expression. Notably, the TPM value of aan-miR-N16 was almost 150-fold enhanced in PI midgut compared with BF midgut. Meanwhile, the expression of miR-31 was enhanced more than 10-fold after Plasmodium infection. On the other hand, the expression levels of 9 known (miR-133, miR-14, miR-210, miR-252, miR-2a, miR-275, miR-92b, miR-957 and miR-980) and 2 novel miRNAs (miR-N16 and miR-N18) were significantly downregulated in the Plasmodium infection midgut. In particular, the TPM value for miR-252 was 191 in BF midgut but zero in the PI midgut (Tables 2 and 3).
Furthermore, several miRNA exhibited different expression patterns in BF and PI midguts. For example, four miRNAs including miR-1, miR-12, miR-13 and miR-276 were significantly downregulated in the BF midguts but upregulated in the PI midguts. Conversely, the expression of miR-252 and miR-N16 showed significant upregulation in the BF midguts but were downregulated in the PI midguts (Tables 2 and 3). These data suggest that the same miRNAs may play different roles in the blood-feeding and Plasmodium infection midguts.
Confirmation of mosquito miRNAs
Having identified the microRNAs expression patterns of An. anthropophagus midguts after blood-feeding and Plasmodium infection, Northern hybridization was performed to validate some of these miRNAs. The representative abundant miRNA (miR-275 and miR-92a) were selected in the Northern blot analysis. Total RNA from SF, BF and PI 48 h were probed using locked nucleic acid (LNA) probe. The expression patterns of miR-275 and miR-92a in SF, BF and PI are shown in Fig. 3. The northern signals at ~20 nt indicated that the expression level of miR-92a is upregulated to over 5-fold after blood-feeding, and miR-275 is downregulated to more than 3-fold after Plasmodium infection, which is consistent with the sequencing results.

Northern blot of differentially expressed miRNAs in the An. anthropophagus midguts. Abbreviations: SF, sugar-feeding midgut; BF, blood-feeding midgut; PI, Plasmodium-infected midgut
Target prediction and network analysis
Target prediction were carried out to understand the putative function of regulated miRNAs by using RNA hybrid (P-value < 0.05). All the mRNA targets are derived from the orthologous genes of An. gambiae in the Vector Base. Maximum numbers of mRNA targets were predicted for miR-92a (n = 744). The targets were further analyzed by KOBAS, and a total of 38 different pathways were regulated by miRNAs after blood-feeding and Plasmodium infection (Fig. 4). In the upregulated miRNA after blood-feeding, miR-252 and miR-92a were identified to target oxidative phosphorylation and peroxisome pathway. miR-92a was also found to target glycolysis, proteasome, ribosome and TGF-beta signaling pathway. Meanwhile, ribosome was commonly targeted by the downregulated miRNAs including miR-13, miR-279 and miR-2b in BF midguts. After Plasmodium infection, endocytosis and fructose mannose metabolism pathway is specifically targeted by miR-31. Spliceosome was common between miR100 and miR-14. In the downregulated miRNA, miR-210 and miR-275 targeted RNA transport and purine metabolism separately (Fig. 4).

KOBAS analysis of miRNA targets. miRNA targets involved in blood-feeding and Plasmodium infection were predicted by RNA hybrid. Pie chart represents the pathway targeted by the miRNAs which are listed in the pie area
The interaction networks of miRNA:mRNA targets were generated. In BF midguts, AGAP000348-RA and AGAP000120-RA were targeted by downregulated miR-1. A total of 493 mRNA targets were regulated by upregulated miR-989, of which AGAP007839-RA, AGAP0004451-RA and AGAP008345-RA were targeted by two or more upregulated and downregulated miRNAs (Additional file 3: Figure S1). Furthermore, 573 mRNAs were targeted by upregulated miR-31 in the PI midguts. Three mRNA (AGAP000204-RA, AGAP009077-RA and AGAP010306-RA) were targeted by downregulated miR-957 (Additional file 4: Figure S2). Further investigations are important to understand the role of these mRNAs during blood-feeding and Plasmodium infection.
Discussion
Anopheles mosquitoes are the main vector for the transmission of malaria, and the invasion of midguts is one of the most critical steps for the survival and development of Plasmodium [21]. Because of the lack of an adaptive immune system, RNA interference (RNAi) is the most important and primary defense employed by mosquitoes to protect themselves from pathogens [22, 23]. miRNAs are the most important part of RNA interference, and some of them have been proven to be involved in fighting against virus and parasite infections [21–24]. In a previous study, we have investigated the miRNA profiles of An. anthropophagus, and discovered that several miRNAs exhibit sexual differences and stage-specific functions [12]. However, their role in Plasmodium infection is poor understood. This study was conducted to identify and elucidate role of miRNAs after blood-feeding and Plasmodium infection in An. anthropophagus midguts.
In the present study, the expression profiles of miRNA at 48 h after blood-feeding and Plasmodium infection were investigated using small RNA sequencing; sugar-feeding midguts were taken as the control. Compared with sugar feeding, there is a significant downregulation for the percent of miRNA library (20–23 nt) after blood-feeding and Plasmodium infection. The similar downregulation expression profiling of miRNAs was also reported in the parasitized blood-feeding midguts of Anopheles stephensi [18] suggested that pathogens including Plasmodium and viruses invade mosquitoes midguts by inhibiting or downregulating the miRNA expression [22–24].
Blood-feeding is a critical physiological activity for the mosquito and its ability to transmit disease [25]. Female mosquitoes take blood meals to carry out egg production, and acquire pathogens such as malaria parasites and dengue viruses from an infected host [26–28]. Furthermore, mosquito biological characteristics are affected by triggering a cascade of gene regulatory events in the midgut after blood-feeding [28, 29]. In this study, the correlation between blood-feeding and miRNA expression profiles in An. anthropophagus midguts was investigated. We indentified nine significantly upregulated and ten downregulated expression miRNAs in the blood-feeding midguts compared with sugar feeding. For example, miR-92a exhibited a significant enhanced expression level after blood-feeding according to the sequencing and Northern blot results. Li et al. [11] reported that blood-feeding can induce the expression of miR-92a in the midgut of Ae. aegypti females. While Wolbachia infection can downregulate the expression of miR-92a in mosquito cell [30]. In our study, the expression of miR-92a is significantly upregulated in blood-feeding. By target prediction and networks analysis, miR92a was found to target 744 mRNAs and several pathways including the oxidative phosphorylation, proteasome, ribosome and TGF-beta singaling pathway. Our study may shed light on the possible roles of miR92a in An. anthropophagus mosquito physiology related to blood-feeding.
Midgut represents the first barrier for the pathogens to establish infection in mosquitoes [2, 4]. Pathogens such as parasites and endosymbiotic bacteria are known to alter host miRNA profiles for their invasion and development [15, 31]. For example, Wolbachia can induce the expression of miR-12 in An. aegypti mosquito cells to maintan the persistence of infection [31]. In our study, we indentified 13 upregulated miRNAs including miR-12. This finding suggests that miR-12 is also involved in Plasmodium infection, and the role of miR-12 in Plasmodium infection of An. anthropophagus need to be further elucidated. Meanwhile, we also found downregulated expression for 11 miRNA in Plasmodium infection midguts of An. anthropophagus, six of which including miR-14, miR-2a, miR-92b, miR-957, miR-980 and miR-275, were also shown to be downregulated in Plasmodium-infected Anopheles stephensi [18]. miR-275 is indispensable for blood digestion and egg development in the mosquito Ae. aegypti [32]. In this study, we confirmed that miR-275 is also involved in Plasmodium infection of An. anthropophagus by sequencing and further Northern blot analysis.
Conclusions
In conclusion, our study provides the significant experimental data on the expression profile of microRNAs in An. anthropophagus midgut after blood-feeding and Plasmodium infection. Differentially expressed miRNA in SF, BF and PI were identified by small RNA sequencing, and further validated by Northern blot. By comparative analyzing differentially expression levels of the miRNA in sugar-feeding, blood-feeding and Plasmodium infection, we found several significant miRNAs involved in the interaction of mosquito host and parasite Plasmodium. Our study provides novel regulated miRNAs information of An. anthropophagus during blood-feeding and parasite infection. Elucidation of regulated miRNA functions will provide strong foundation for better understanding of the biology of the mosquitoes and mosquito-parasite interactions.
Abbreviations
- BF:
-
Blood-feeding
- FPKM:
-
Fragments Per Kilobase of transcript per Million mapped reads
- miRNAs:
-
microRNAs
- PI:
-
Plasmodium infection
- RNAi:
-
RNA interference
- SF:
-
Sugar feeding
References
World Malaria Report 2015. http://www.who.int/malaria/publications/world-malaria-report-2015/en/. Accessed on 3 Jan. 2016
Sinden RE, Alavi Y, Raine JD. Mosquito-malaria interactions: a reappraisal of the concepts of susceptibility and refractoriness. Insect Biochem Mol Biol. 2004;34:625–9.
Meister S, Koutsos AC, Christophides GK. The Plasmodium parasite - a ‘new’ challenge for insect innate immunity. Int J Parasitol. 2004;34:1473–82.
Whitten MM, Shiao SH, Levashina EA. Mosquito midguts and malaria: cell biology, compartmentalization and immunology. Parasite Immunol. 2006;28:121–30.
Zhang G, Niu G, Franca CM, Dong Y, Wang X, Butler NS, et al. Anopheles Midgut FREP1 mediates Plasmodium invasion. J Biol Chem. 2015;290:16490–501.
Nikolaeva D, Draper SJ, Biswas S. Toward the development of effective transmission-blocking vaccines for malaria. Expert Rev Vaccines. 2015;14:653–80.
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–97.
Miska EA. How microRNAs control cell division, differentiation and death. Curr Opin Genet Dev. 2005;15:563–8.
Zhang R, Wang YQ, Su B. Molecular evolution of a primate-specific microRNA family. Mol Biol Evol. 2008;25:1493–502.
Bushati N. MicroRNA Functions. Ann Rev Cell Dev Biol. 2007;23:175–205.
Li S, Mead AE, Liang SH, Tu ZJ. Direct sequencing and expression analysis of a large number of miRNAs in Aedes aegypti and a multi-species survey of novel mosquito miRNAs. BMC Genomics. 2009;10:581.
Liu W, Huang H, Xing C, Li C, Tan F, Liang S. Identification and characterization of the expression profile of microRNAs in Anopheles anthropophagus. Parasit Vectors. 2014;7:159.
Winter F, Edaye S, Hüttenhofer A, Brunel C. Anopheles gambiae miRNAs as actors of defence reaction against Plasmodium invasion. Nucleic Acids Res. 2007;35(20):6953–62.
Skalsky LR, Vanlandingham LD, Scholle F, Higgs S, Cullen RB. Identification of microRNAs expressed in two mosquito vectors, Aedes albopictus and Culex quinquefasciatus. BMC Genomics. 2010;11:119.
Biryukova I, Ye T, Levashina E. Transcriptome-wide analysis of microRNA expression in the malaria mosquito Anopheles gambiae. BMC Genomics. 2014;15:557.
Castellano L, Rizzi E, Krell J, Di Cristina M, Galizi R, Mori A, et al. The germline of the malaria mosquito produces abundant miRNAs, endo-siRNAs, piRNAs and 29-nt small RNAs. BMC Genomics. 2015;16:100.
Jain S, Rana V, Tridibes A, Sunil S, Bhatnagar RK. Dynamic expression of miRNAs across immature and adult stages of the malaria mosquito Anopheles stephensi. Parasit Vectors. 2015;8:179.
Jain S, Rana V, Shrinet J, Sharma A, Tridibes A, Sunil S, Bhatnagar RK. Blood feeding and Plasmodium infection alters the miRNome of Anopheles stephensi. PLoS One. 2014;9:e98402.
Shrinet J, Jain S, Jain J, Bhatnagar RK, Sunil S. Next generation sequencing reveals regulation of distinct Aedes microRNAs during chikungunya virus development. PLoS Negl Trop Dis. 2014;8(1):e2616.
Geng YJ, Gao ST, Huang DN, Zhao YR, Liu JP, Li XH, Zhang RL. Differentially expressed genes between female and male adult Anopheles anthropophagus. Parasitol Res. 2009;105:843–51.
Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J, et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasit Vectors. 2010;3:117.
Carol DB. Mosquito RNAi is the major innate immune pathway controlling arbovirus infection and transmission. Future Microbiol. 2011;6:265–77.
Cirimotich CM, Dong Y, Garver LS, Sim S, Dimopoulos G. Mosquito immune defenses against Plasmodium infection. Dev Comp Immunol. 2010;34:387–95.
Campbell LC, Harrison T, Hess MA, Ebel DG. MicroRNA levels are modulated in Aedes aegypti following exposure to Dengue-2. Insect Mol Biol. 2014;23(1):132–9.
Dennison NJ, BenMarzouk-Hidalgo OJ, Dimopoulos G. MicroRNA-regulation of Anopheles gambiae immunity to Plasmodium falciparum infection and midgut microbiota. Dev Comp Immunol. 2015;49:170–8.
Cator LJ, Pietri JE, Murdock CC, Ohm JR, Lewis EE, Read AF, et al. Immune response and insulin signalling alter mosquito feeding behaviour to enhance malaria transmission potential. Sci Rep. 2015;5:11947.
Harrington LC, Edman JD, Scott TW. Why do female Aedes aegypti (Diptera: Culicidae) feed preferentially and frequently on human blood? J Med Entomol. 2001;38:411–22.
Ferguson HM, Read AF. Mosquito appetite for blood is stimulated by Plasmodium chabaudi infections in themselves and their vertebrate hosts. Malar J. 2004;3:12.
Putnam JL, Scott TW. Blood-feeding behavior of dengue-2 virus-infected Aedes aegypti. Am J Trop Med Hyg. 1995;52:225–7.
Mayoral JG, Etebari K, Hussain M, Khromykh AA, Asgari S. Wolbachia infection modifies the profile, shuttling and structure of microRNAs in a mosquito cell line. PLoS One. 2014;9:e96107.
Osei-Amo S, Hussain M, O’Neill SL, Asgari S. Wolbachia-induced aae-miR-12 miRNA negatively regulates the expression of MCT1 and MCM6 genes in Wolbachia-infected mosquito cell line. PLoS One. 2012;7(11):e50049.
Bryant B, Macdonald W, Raikhel AS. microRNA miR-275 is indispensable for blood digestion and egg development in the mosquito Aedes aegypti.Proc Natl Acad Sci USA. 2010;107(52):22391–8.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (81071389), the Commonweal Technology Application Project of Zhejiang Province (2015C33101) and the Technology Project of Wenzhou City (Y20140481).
Funding
The National Natural Science Foundation of China (81071389), the Commonweal Technology Application Project of Zhejiang Province (2015C33101) and the Technology Project of Wenzhou City (Y20140481).
Availability of data and materials
The datasets supporting the conclusions of this article are available in the Gene Expression Omnibus repository, [GSE93545, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE93545].
Authors’ contributions
SHL and WQL conceived and designed the study. WQL, ZHH, LYH, LZC, QMW and LYC performed the experiments, analyzed the data and drafted the manuscript. SHL critically revised the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval
Animal experiments were performed in accordance with National animal ethics guidelines of the Government of P. R. China after approval by the Animal Ethics Committees of Wenzhou Medical University (Permit number: SYXKZHE20150009).
Author information
Authors and Affiliations
Corresponding author
Additional files
Additional file 1: Table S1.
Known miRNAs in Anopheles anthropophagus midgut. (XLSX 14 kb)
Additional file 2: Table S2.
Novel miRNAs in Anopheles anthropophagus midgut. (XLS 25 kb)
Additional file 3: Figure S1.
Interaction network of regulated miRNA and mRNA targets after blood-feeding. (JPG 1651 kb)
Additional file 4: Figure S2.
Interaction network of regulated miRNA and mRNA trargets after Plasmodium infection. (JPG 1638 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

{kind=link}
{kind=link}
Cite this article
Liu, W., Hao, Z., Huang, L. et al. Comparative expression profile of microRNAs in Anopheles anthropophagus midgut after blood-feeding and Plasmodium infection. Parasites Vectors 10, 86 (2017). https://doi.org/10.1186/s13071-017-2027-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-017-2027-6