
Abstract
Background
Lignin is a highly abundant but strongly underutilized natural resource that could serve as a sustainable feedstock for producing chemicals by microbial cell factories. Because of the heterogeneous nature of the lignin feedstocks, the biological upgrading of lignin relying on the metabolic routes of aerobic bacteria is currently considered as the most promising approach. However, the limited substrate range and the inefficient catabolism of the production hosts hinder the upgrading of lignin-related aromatics. Particularly, the aerobic O-demethylation of the methoxyl groups in aromatic substrates is energy-limited, inhibits growth, and results in carbon loss in the form of CO2.
Results
In this study, we present a novel approach for carbon-wise utilization of lignin-related aromatics by the integration of anaerobic and aerobic metabolisms. In practice, we employed an acetogenic bacterium Acetobacterium woodii for anaerobic O-demethylation of aromatic compounds, which distinctively differs from the aerobic O-demethylation; in the process, the carbon from the methoxyl groups is fixed together with CO2 to form acetate, while the aromatic ring remains unchanged. These accessible end-metabolites were then utilized by an aerobic bacterium Acinetobacter baylyi ADP1. By utilizing this cocultivation approach, we demonstrated an upgrading of guaiacol, an abundant but inaccessible substrate to most microbes, into a plastic precursor muconate, with a nearly equimolar yields (0.9 mol/mol in a small-scale cultivation and 1.0 mol/mol in a one-pot bioreactor cultivation). The process required only a minor genetic engineering, namely a single gene knock-out. Noticeably, by employing a metabolic integration of the two bacteria, it was possible to produce biomass and muconate by utilizing only CO2 and guaiacol as carbon sources.
Conclusions
By the novel approach, we were able to overcome the issues related to aerobic O-demethylation of methoxylated aromatic substrates and demonstrated carbon-wise conversion of lignin-related aromatics to products with yields unattainable by aerobic processes. This study highlights the power of synergistic integration of distinctive metabolic features of bacteria, thus unlocking new opportunities for harnessing microbial cocultures in upgrading challenging feedstocks.
Similar content being viewed by others
Introduction
Lignin is the second most abundant polymer in nature, mainly consisting of three aromatic units comprising H- (p-coumaryl alcohol), G- (coniferyl alcohol), and S- (sinapyl alcohol) lignin types [1]. Large quantities of lignin, often considered as a waste material, are generated through pulping and biorefinery industries, agriculture, and wood processing [2]. Due to lignin’s complexity and heterogeneity, it is currently a strongly underutilized resource. However, it could serve as a renewable replacement for fossil-based resources in the production of chemicals and materials, for which biological lignin valorization is considered as the most promising approach. To access the monomers of the complex lignin polymer, the covalent C–C and C–O bonds connecting the units must be broken down via depolymerization or pyrolysis reactions [3]. The chemical structures of the obtained monomers are derivatives of the lignin units, varying with the number of their methoxyl and hydroxyl groups and the structure of the possible acrylate chain linked to the aromatic ring [4].
Biological lignin valorization often relies on the specific catabolic pathways of microbes, such as β-ketoadipate pathway [5], to funnel heterogeneous mixtures of aromatic compounds into central carbon metabolites [6, 7]. Previously several bacteria, including species of Pseudomonas and Rhodococcus, have been engineered to produce industrially relevant compounds, such as plastic precursor cis,cis-muconate (ccMA) [8,9,10,11], β-ketoadipate and muconolactone [12], as well as native products including polyhydroxyalkanoates (PHAs) and triacylglycerides (TAGs) [6], from lignin-derived aromatic compounds. We have previously engineered Acinetobacter baylyi ADP1 (hereafter referred to as ADP1) to produce 1-alkenes or alkanes [13,14,15] and wax esters [14] from lignin-related substrates or technical lignin. In addition, ADP1 has also been utilized to produce mevalonate from lignin-related aromatics [16].
However, the biological lignin valorization by microbes has its own hardships, including limited aromatic substrate range and the toxicity of the aromatic compounds causing growth inhibition at industrially relevant concentrations [6, 17]. For example, the valorization of guaiacol (2-methoxyphenol), one of the main monomeric products obtained from alkaline-pretreated softwood lignin, has been set as an important goal [6], but only few bacteria, like Rhodococcus rhodochrous [18] and Amycolatopsis sp. ATCC 39116 [19], can naturally catabolize the compound. The ability to utilize guaiacol has previously been engineered in non-native hosts including Pseudomonas putida EM42 [20] and ADP1 [21]. However, to achieve sufficient utilization of guaiacol, co-expression of redox partners [20], or evolution-based approaches [21] are often required.
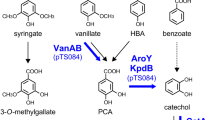
In addition to the restricted substrate range, also the natural pathways exhibit inherent challenges: the G- and S-lignin-derived compounds that contain one and two methoxyl groups, respectively, need to be demethylated prior to further catabolic steps. Aerobic O-demethylation is recognized as one of the major hurdles in the utilization of lignin-related aromatic compounds [22,23,24]. In aerobic conditions, O-demethylase reactions are catalyzed by enzymes belonging into either Rieske oxygenases (ROs), cytochrome P450 or tetrahydrofolate (THF)-dependent demethylases classes [25]. For example, VanAB enzyme pair in P. putida is RO-type, which demethylates G-lignin-derivate vanillate [26] and S-lignin-based syringate [27] into protocatechuate (PCA) and gallate, respectively. Also ADP1 harbors similar VanAB system and it is known to convert vanillate into PCA [28]. However, the O-demethylation reactions catalyzed by ROs or P450s produce highly cytotoxic formaldehyde [29] as their by-product [22, 26], hence further contributing to the toxicity of the aromatic compounds. To eliminate formaldehyde, cells harbor detoxification routes [29] which eventually degrade the compound into CO2 with a price of losing the carbon [23]. In addition, the aerobic O-demethylation causes intracellular cofactor imbalance and reduced primary metabolic activity [24]. Thus, to ensure efficient O-demethylation, a supporting carbon source maintaining the primary metabolism is often provided [24, 30]. Because of these issues, novel approaches to circumvent the problems related to aerobic O-demethylation are required.
Acetogens are anaerobic bacteria that can grow autotrophically on H2-CO2 mixtures using Wood–Ljungdahl pathway for synthesizing acetyl-CoA from CO2 [31, 32]. Because of their ability to fix gaseous CO2, acetogens could play a vital role in biotechnology for the sustainable production of biochemicals. The usage of acetogens to provide organic carbon compounds from CO2 in two-stage processes was recently reviewed by Ricci et al. [33]: in this approach, the fermentation products of acetogens, such as acetate or ethanol, are subsequently utilized and upgraded by aerobic bacteria. Both acetate and ethanol enter the central metabolism as acetyl-CoA, making the number of possible end products nearly limitless [33, 34]. We have previously utilized two-stage processes where we first cultivated Acetobacterium woodii on CO2 to produce acetate, which was then used as a substrate by ADP1 for wax ester [35] or alkane [36] syntheses.
In addition to H2-CO2 mixtures, acetogens utilize a variety of organic C1-compounds [37] as additional carbon sources and electron donors for the CO2 fixation. Interestingly, some acetogens can also utilize the methoxyl groups of aromatic compounds, including those derived from G-lignin, such as guaiacol, vanillate, and ferulate, by removing the methyl group by anaerobic O-demethylation [38]. Demethylating reactions have been observed in several types of acetogens including A. woodii, Moorella thermoacetica, and Sporomusa ovata [39]. In A. woodii, the O-demethylation involves a methyltransferase system that binds the cleaved methyl group to THF [40]. The methyl-THF can then be utilized to fix CO2 by Wood–Ljungdahl pathway, and eventually the carbons from methyl group and CO2 are used to form two-carbon compound acetate [40, 41]. Thus, by binding the methyl group to THF, the formation of cytotoxic formaldehyde can be avoided, while the process fixes CO2 and produces acetate [40]. The biomass production and the molar yield of the produced acetate are positively correlated with the number of the methoxyl groups in the aromatic compound [42]. In addition to the methoxyl group demethylation, A. woodii is also able to reduce any double bonds located in the acrylate side chain of the aromatic compounds, and to use the bonds as electron acceptor instead of CO2 [43], which results in increased biomass but lower acetate production yields [42]. During these processes, the aromatic ring stays unchanged [42].
In this study, we present a novel concept for upgrading lignin-related aromatic compounds into industrially interesting products while simultaneously overcoming the issues of aerobic O-demethylation and carbon loss. To achieve this, we integrated the best features of acetogens and aerobic bacteria. First, we investigated the suitability of the O-demethylation end products of A. woodii for A. baylyi ADP1 growth. Following this, we established a two-stage culture showing the compatibility of A. woodii and ADP1 for synergistic utilization of lignin-related aromatic compounds. Finally, as a proof-of-concept, we demonstrated an equimolar conversion of guaiacol, a model compound of G-lignin and an inaccessible substrate for ADP1, into muconate (MA) by a one-pot coculture approach. In the process, A. woodii demethylates guaiacol into catechol, which is subsequently converted into MA by ADP1. Importantly, employing the anaerobic O-demethylation step ensured that the methoxyl group of guaiacol was not lost to CO2, but instead fixed into acetate, thus providing a carbon source for the growth of ADP1. Simultaneously, we were able to bypass the key bottleneck, aerobic O-demethylation, in the utilization of lignin-related aromatic compounds. This study highlights the potential of harnessing and integrating distinctive metabolic features of bacteria for improved carbon recovery and overcoming metabolic bottlenecks in the valorization of recalcitrant feedstock.
Results and discussion
Set-up for the utilization of aromatic compounds by integrating the metabolisms of anaerobic and aerobic bacteria
To investigate the utilization of lignin-related aromatic compounds by the collaboration of anaerobic and aerobic bacteria, we employed the acetogen A. woodii and the strictly aerobic ADP1 for this study. ADP1 was chosen as the aerobic production chassis because of its versatile metabolism and the previously demonstrated good performance in coculture and two-stage cultivations [44]. In the designed co-utilization scheme, A. woodii would utilize the methoxylated aromatic compounds via O-demethylation, yielding acetate and the hydroxylated counterparts of the original aromatics [42] (Fig. 1A). In addition, A. woodii would also reduce any double bonds located in the possible acrylate side chains of the aromatic substrates, if suitable electron donors are present [42, 45].

The valorization of lignin-related compounds to products by A. woodii and/or ADP1. Ferulate and guaiacol are used as example model compounds. A A. woodii utilizes the methyl groups of the compounds as carbon and electron donors and double bond of the ferulate as electron acceptor in a process that fixes CO2 and the cleaved methyl group into acetate. However, the modified aromatic compounds cannot be further utilized by A. woodii. B ADP1 cannot use guaiacol for its growth, so it is left unutilized. Ferulate is utilized as a substrate via β-ketoadipate pathway and can be directed to biomass or products of interest, but the O-demethylation produces toxic formaldehyde which is degraded into CO2 (presented with dashed circle). C By modifying ferulate and guaiacol first anaerobically with A. woodii, ADP1 can utilize all the obtained products for its substrate. Theoretically, higher yields can be obtained compared to the process including only ADP1. In addition, by executing the O-demethylation of the aromatic compounds anaerobically, the formation of the toxic by-product formaldehyde can be avoided
We hypothesized that the metabolites produced by A. woodii when cultivated on aromatic compounds could be used as substrates by ADP1 for biomass and product formation (Fig. 1C). By utilizing anaerobic O-demethylation, ADP1 would get an access to previously unavailable carbon: if ADP1 performed the O-demethylation reaction by itself, the carbon from the methoxyl group would form cytotoxic formaldehyde, eventually converted into CO2 (Fig. 1B).
The utilization of compounds modeling the anaerobic O-demethylation metabolites of A. woodii by ADP1
We first studied if ADP1 could grow on the substrates modeling the O-demethylation metabolites of A. woodii. In addition, we were interested to see if the growth metrics obtained on the metabolites would differ from the situation where ADP1 is cultivated solely on the unmodified methoxylated aromatics. According to the literature, when A. woodii is cultivated on 10 mM vanillate, 10 mM PCA and 7.5 mM acetate are produced [42] (Table S1, Fig. 2A). In our experiments, molar yields of similar range were obtained (Table S2). The effects of the different carbon sources on biomass yields were first examined by performing a flux balance analysis (FBA). According to the ADP1 model, 10 mM vanillate provides 0.77 g of cell dry weight (gCDW), whereas the biomass production from the mixture of 10 mM PCA and 7.5 mM acetate is 0.96 gCDW in a steady state environment. The results of the FBA were supported by the experimental data: when we cultivated ADP1 on 10 mM vanillate or on 10 mM PCA and 7.5 mM acetate mixture (modeling the metabolites produced by A. woodii during during growth on vanillate and CO2), faster cell growth and higher optical density measured at 600nm (hereafter OD600) were achieved in the latter culture (Fig. 2B). The cells grown on vanillate reached maximum OD600 of 0.85, whereas the cells grown on mixture reached OD600 of 0.99, corresponding to 0.32 gCDW and 0.38 gCDW, respectively (Supplementary Fig. S1). Thus, the ratio of the CDWs calculated from obtained OD600 values was similar, 0.84, to the ratio of the CDW values calculated by the FBA model, 0.80. Despite acetate (especially in combination with succinate) is known to repress the multiple operons related to the utilization of aromatic compounds, such as vanillate and benzoate, in ADP1 [46, 47], we observed no negative effects or diauxic growth pattern on the culture with a mixture of PCA and acetate.

A Anaerobic vanillate O-demethylation in A. woodii in the presence of CO2. 10 mM vanillate is converted into 10 mM PCA and 7.5 mM acetate. B The difference of ADP1 growth on 10 mM vanillate and on mixture containing 10 mM PCA and 7.5 mM acetate in aerobic conditions. The mixture represents the end products of vanillate O-demethylation and subsequent processing of the methyl group in the Wood–Ljungdahl pathway by A. woodii. The error bars indicate the standard deviations from the average values of the biologically independent triplicates. In some cases, the error bars are smaller than the size of the marker
Thus, the anaerobic conversion of the methoxylated aromatic substrates into a more accessible form, accompanied by CO2 fixation into acetate, has clear advantages, seen as both higher biomass and improved growth rate of ADP1.
The utilization of ferulate and coumarate by A. woodii and ADP1
Ferulate and coumarate are model compounds of G- and H-lignin, respectively, that both contain acrylate side chains. According to the literature, A. woodii produces 1.0 mol dihydrocaffeate ((3-(3,4-dihydroxyphenyl)propanoate) and 0.5 mol acetate from 1.0 mol ferulate [42]. Coumarate does not contain any methoxyl groups, so the only possible modification is the reduction of the double bond in the acrylate tail, theoretically resulting in the formation of stoichiometric amounts of phloretate (3-(4-hydroxyphenyl)propionate) in the presence of electron donors [48]. We studied the growth of A. woodii on 13 mM ferulate or 14 mM coumarate under N2-CO2. When ferulate was used as a substrate, we detected notable amounts of caffeate (3-(3,4-dihydroxyphenyl)-2-propenoate) in addition to dihydrocaffeate and acetate (Fig. S2A), indicating that the O-demethylation of the methoxyl group of ferulate along with CO2 fixation occurs preferentially over double bond reduction in the studied conditions. The molar yields of dihydrocaffeate and acetate from ferulate were 0.70 ± 0.24 mol/mol and 0.66 ± 0.14 mol/mol, respectively (Table S3). Unlike methoxyl groups that act as electron donors, double bonds in acrylate side chains are electron acceptors, similar to CO2 [48]. When A. woodii was cultivated on coumarate, minor formation of phloretate (3-(4-hydroxyphenyl)propionate) and acetate was detected, despite the absence of added electron donor, indicating that energy for the double bond reduction and CO2 fixation was potentially provided by carry-over fructose from the preculture (Fig. S2B) (Table S4).
We then investigated the growth metrics of ADP1 on ferulate and coumarate in comparison to the growth on the metabolites provided by A. woodii cultivated on ferulate and coumarate, namely, caffeate, dihydrocaffeate, and phloretate (Fig. 3A). In this experiment, we concentrated solely on the aromatic metabolites and thus neglected the positive effect of acetate on growth (Fig. 2B). A clear difference in the growth of ADP1 was observed between the different substrates: in all cases, the obtained biomass and growth rate on demethylated and/or reduced aromatics were higher compared to those on ferulate and coumarate (Fig. 3B, C), and the effect was even more pronounced with higher concentrations (Fig. S3, S4); when ADP1 was cultivated on 15 mM ferulate, the lag-phase lasted approximately 1 day and the obtained biomass was modest, whereas with caffeate, the exponential growth phase was reached earlier, and the biomass growth resulted in OD600 1.2. The final OD600 of the cells grown on dihydrocaffeate was the same than with caffeate-grown cells, but the growth occurred more slowly. The difference in the cell growth on coumarate and phloretate was less drastic, although phloretate-grown cells had shorter lag-phase with final OD600 of 1.2, whereas with coumarate-grown cell, the final OD600 remained under 1.

A A simplified scheme of the modification carried out by A. woodii for ferulate and coumarate. B. The growth of ADP1 on 15 mM of ferulate, caffeate, and dihydrocaffeate. C The growth of ADP1 on 15 mM coumarate and phloretate. The error bars in graph B and C indicate the standard deviations from the average values of the biologically independent triplicates. In some cases, the error bars are smaller than the size of the marker
These results affirm that the modifications carried out by A. woodii for the aromatic compounds are beneficial for ADP1 growth: the growth metrics were enhanced, indicating that caffeate, dihydrocaffeate, and phloretate are more readily consumed and potentially less toxic carbon sources compared to ferulate and coumarate, respectively. However, interestingly, the ADP1 growth was faster on double bond-containing caffeate than on dihydrocaffeate lacking the double bond. We hypothesize the phenomenon to be related to the catabolic route of dihydrocaffeate in ADP1; unlike with caffeate, the catabolism of dihydrocaffeate requires an additional enzymatic step, where 3,4-dehydroxyphenolpropionyl-CoA is converted to caffeoyl-CoA, which is a direct degradation intermediate of caffeate catabolism [49]. This finding creates an interesting opportunity: when A. woodii was cultivated on ferulate, complete conversion of ferulate into dihydrocaffeate was not observed, but notable amounts of caffeate were still present in the medium after 164 h (Fig. S2A). Thus, much shorter cultivation time of A. woodii would be sufficient, making the overall cultivation process faster and simultaneously providing the most suitable carbon source and maximal amount of acetate for ADP1 growth.
The negative effect of the methoxyl group on ADP1 growth is evident when ferulate and caffeate are compared, because the only difference between the two compounds is the lack of the methoxyl group in caffeate [49]. Most likely the lack of methoxyl group on both coumarate and phloretate molecules explain why the growth metrics of ADP1 on these compounds did not differ that dramatically. To the best of our knowledge, there are no previous studies reporting the growth of ADP1 on phloretate. Therefore, the exact metabolic pathway for phloretate catabolism is not yet known and the reason why ADP1 grew better on phloretate than coumarate remains unclear.
Complete utilization of guaiacol by A. woodii and ADP1 coculture
After confirming the suitability of A. woodii metabolites on ADP1 growth, we next established a two-stage culture for the complete utilization of a lignin-related model compound, guaiacol. Unlike vanillate, ferulate, or coumarate, ADP1 cannot naturally utilize guaiacol. We further confirmed this by cultivating ADP1 in MSM with guaiacol as a sole carbon source (data not shown). It has been previously reported that A. woodii can grow on guaiacol, producing catechol and acetate as end-products [50, 51]. To obtain quantitative information about the process, we cultivated A. woodii on 6, 11, and 18 mM guaiacol as a sole organic carbon source (Table S5). The obtained OD600 values and the molar yields of catechol and acetate from guaiacol were in line with previous literature, being approximately 0.85 mol/mol for catechol and 0.7 mol/mol for acetate.
While ADP1 cannot grow on guaiacol, both catechol and acetate serve as suitable carbon sources for its growth. However, we were concerned about the toxicity of catechol to ADP1. For example, Kohlstedt et al. [52] have reported that catechol concentrations above 2 mM caused reduced growth rates, while 8 mM catechol completely inhibited the cell growth of P. putida. Thus, the tolerance of ADP1 toward catechol was first tested by cultivations in MSM supplemented with 0.5, 1, 2, 5, 7.5, and 10 mM of catechol as a sole carbon source (Supplementary Fig. S5). ADP1 showed unexpectedly high tolerance toward catechol, as the highest OD600s were achieved with 5 mM and 7.5 mM catechol concentrations, which were 0.7 and 0.8, respectively. However, 7.5 mM catechol caused a notable lag-phase, and the exponential cell growth started only after 30 h of cultivation.
As no drastic differences in the obtained biomass between 5 mM and 7.5 mM catechol concentrations were detected, we decided to test the two-stage coculture of A. woodii and ADP1 using 6 mM guaiacol as a sole organic carbon source (Fig. 4). A. woodii consumed all guaiacol within 48 h of cultivation and converted it to catechol and acetate. After the anaerobic phase, the cultivation tubes were opened and ADP1 cells were added to the culture. At the end of the aerobic cultivation with ADP1, no acetate or catechol were detected in the medium.

Two-stage cocultivation of A. woodii and ADP1 on acetobacterium medium supplemented with 6 mM guaiacol as the sole organic carbon source. The increase of acetate concentration at the beginning of the cultivation is a result of the carry-over acetate from A. woodii inoculation. ADP1 was inoculated later, at 141 h, marked in the graph with dashed vertical line. The error bars indicate the standard deviations from the average values of the biologically independent duplicates. In some cases, the error bars are smaller than the size of the marker
It was confirmed that ADP1 was able to grow on the authentic metabolites produced by A. woodii in the acetobacterium medium (ABM). By utilizing the coculture approach, a complete degradation of guaiacol was achieved using wild-type cells, without the need for genetic engineering, although ADP1 is not able to utilize guaiacol naturally. Thus, this cocultivation approach can be utilized to widen the aromatic substrate range of aerobic bacteria by modifying the compounds in more accessible form by employing bacteria capable of anaerobic O-demethylation.
Production of muconate from guaiacol with A. woodii and ADP1 coculture
To demonstrate that lignin-related aromatic compounds could be converted into products by utilizing the integrated metabolisms of A. woodii and ADP1, we established the production of MA from guaiacol. Muconate is a natural intermediate compound of the β-ketoadipate pathway, located in the catechol branch. To allow the accumulation of MA, we constructed a knock-out mutant strain with catBC deletion, thus preventing further catabolism of MA. The resulting knock-out strain was designated as ADP1ΔcatBC.
We tested the production of MA in a two-stage process, where we first cultivated A. woodii on 11 mM guaiacol as a sole organic carbon source. After the conversion of guaiacol to 10.9 ± 0.3 mM of catechol and 8.4 ± 1.8 mM of acetate, the anaerobic cultivation was stopped and A. woodii cells were removed from the cultivation by centrifugation. The cultivation medium was diluted to lower the catechol concentration to 5.4 ± 0.2 mM and ADP1ΔcatBC cells were added to the culture.
ADP1ΔcatBC was able to grow and produce MA solely from A. woodii metabolites. Because of the knock-out mutation, only acetate was used to produce biomass reaching the OD600 1.0 ± 0.0. The strain converted all catechol into ccMA, but because of the spontaneous isomerization of the compound, also small amounts of cis–trans-muconate (ctMA) were detected from the medium. We further confirmed the product to be MA with liquid chromatography mass spectrometry (LC–MS) analysis. In the diluted cultivation, the total concentration of produced MA was 4.9 ± 0.1 mM. Therefore, the total molar yield of MA from catechol was 0.91 ± 0.05 mol/mol, while the total molar yield of MA from guaiacol was 0.87 ± 0.04 mol/mol.
Previously, production of MA from guaiacol has been achieved by engineered Amycolatopsis sp. ATCC 39116 [53] with near equimolar and P. putida [30] with equimolar yields. However, in addition to guaiacol, the cultivations with growing cells were supplemented with glucose to support primary metabolism and cell growth; Almqvist et al. [30] reported that guaiacol O-demethylation caused a severe redox equivalent imbalance to the cells, and by adding glucose to the cultivations, 10-fold higher MA production rates were achieved. They tested whether vanillin, present in their depolymerized lignin feedstock, could replace glucose as an additional carbon source, but too low ratio of vanillin to guaiacol in the feedstock as well as inhibitory effect caused by higher vanillin concentrations was found to be problematic. In contrast, by utilizing our coculture approach, guaiacol can serve as the sole organic carbon source, as the anaerobic O-demethylation of guaiacol along with CO2 fixation provides acetate for ADP1 growth and circumvents the redox imbalance issue.
A one-pot cocultivation of A. woodii and ADP1 for muconate production
While A. woodii and ADP1 represent strictly anaerobic and aerobic bacteria, respectively, our previous work indicates that it might be possible to carry out a one-pot cocultivation of A. woodii and ADP1 with alternating anaerobic and aerobic phases [54, 55]. The one-pot approach allows for more straight-forward culture operation and potentially reduces the conversion time of substrates to products. To that end, we established a three-step batch process in a bioreactor for the production of MA from guaiacol.
We started the cultivation by adding ADP1ΔcatBC in ABM medium prepared without using any anaerobic techniques. In addition to 5 mM guaiacol, the medium was supplemented with 10 mM glucose to ensure the oxidation activity of ADP1 during the first phase of the cultivation, referred to as ‘deoxygenation phase.’ However, the addition of glucose turned out to be unnecessary as ADP1 did not utilize glucose during the first phase (Fig. S6). After the cell inoculation, the reactor was closed, and ADP1ΔcatBC was able to rapidly deoxygenize the medium. Thereafter, A. woodii cells were anaerobically inoculated into the reactor, which started the anaerobic phase. After 3 days, A. woodii had completely demethylated guaiacol into catechol. Minor amounts of glucose (1.3 mM) were consumed during the anaerobic phase (Fig. S6). The final aerobic phase was initiated by starting the airflow, enabling the growth of ADP1ΔcatBC cells. ADP1ΔcatBC was able to convert all the available catechol into ccMA. During the aerobic phase, all acetate produced by A. woodii was completely consumed. The rate of glucose consumption by ADP1 was notably slow when other carbon sources were present, and it was not consumed during the deoxygenation phase (Fig. S6), indicating that glucose supplementation is not needed in the process.
In the one-pot approach, the metabolic integration between the two bacteria is even more stringent compared to two-stage cultivations as both bacteria are dependent on the metabolic activities of the other: by the deoxygenation of the medium, ADP1 modified the growth environment to be suitable for A. woodii growth. The growth of ADP1 also produces CO2, required for the O-demethylation reactions of the acetogen, while ADP1 is dependent on the end-metabolites of A. woodii (Fig. 5A). Encouragingly, our results suggest that both bacteria were able to stay active in the bioreactor conditions: the consumption of acetate (Fig. S6) and catechol (Fig. 5C) that occurred already during the anaerobic phase indicates that ADP1ΔcatBC was metabolically active by consuming the minor amounts of oxygen that potentially leaked into the bioreactor, while keeping the medium anaerobic and suitable for A. woodii to grow. These exciting results suggest that indeed, it is possible to cultivate the strictly anaerobic acetogen A. woodii together with a strictly aerobic bacteria, opening novel and intriguing possibilities for the utilization of A. woodii. To the best of our knowledge, our study represents the first example of utilizing A. woodii in a one-pot cocultivation containing both aerobic and anaerobic cultivation steps.

Production of MA by ADP1ΔcatBC and A. woodii one-pot cocultivation. A Hypothetical schematics of the cultivation phases and the substrates and products in each phase. ED, Entner–Doudoroff pathway, TCA, Tricarboxylic acid cycle, WL, Wood–Ljungdahl pathway, DM, O-demethylation, BKA, β-ketoadipate pathway. B Experimental data of the three-phase one-pot coculture of A. woodii and ADP1ΔcatBC. The carry-over acetate resulted from the inoculation of A. woodii is subtracted from the acetate concentration, which therefore describes solely the acetate produced by A. woodii during the coculture.DO, deoxygenation phase. C. Close-up of the experimental data from 65.5 h to 85.5 h. Because of the spontaneous isomerization of ccMA to ctMA, the concentration of ccMA decreases during the aerobic phase in the graphs B and C
Equimolar conversion of guaiacol into MA (1.00 mol/mol) in the one-pot cultivation was achieved. Notably, ADP1ΔcatBC converted catechol very fast, within few hours, into MA. The conversion of catechol to MA occurred before all the acetate was consumed (Fig. 5), indicating that the conversion is not under carbon catabolite repression (CCR) caused by acetate, unlike we would have anticipated based on a previous study on other aromatic compounds [47]. CCR effect was not encountered either when ADP1 WT was cultivated in acetate–catechol mixture: in contrast, both substrates were co-utilized, and catechol was consumed even faster than acetate (Supplementary Fig. S7). In addition, ADP1 exhibited good tolerance and metabolic activity toward catechol, as no inhibition of growth or MA production was detected. These observations well justify the selection of ADP1 as the optimal strain for the coculture set-up, as catechol has been identified as the most toxic lignin-related compound to for example P. putida, which has been previously broadly employed for MA production [11].
Nowadays, many of the state-of-the-art approaches to MA production rely on the heterogeneous expression of PCA decarboxylases converting PCA to catechol [56,57,58]. Using PCA decarboxylases such as AroY [59] can broaden the range of suitable substrates for MA accumulation, including vanillate, ferulate, and coumarate. However, producing PCA from lignin-related methoxylated aromatics still requires the problematic step of aerobic O-demethylation. Thus, expressing AroY in ADP1 could be a potential approach, allowing for the production of MA also from methoxyl-free and readily accessible caffeate, dihydrocaffeate, and phloretate produced by A. woodii.
Conclusions
Aerobic O-demethylation is a crucial step in the catabolism of the methoxyl group-rich G- and S-lignin-derived compounds, but the process produces cytotoxic formaldehyde causing cofactor imbalance and results in wasting the carbons from the methoxyl groups into CO2. Here, we presented a novel approach based on the optimal integration of anaerobic and aerobic metabolisms to circumvent these problems while achieving more efficient utilization of lignin-related compounds: by employing anaerobic O-demethylation of the acetogen A. woodii, the carbons from the methoxyl groups and CO2 are reduced to acetate, which together with the demethylated aromatic compounds can be utilized and upgraded to products by the aerobic bacterium A. baylyi ADP1. Thus, our approach converts the methoxyl group of the aromatic compound from a noxious vice into a valuable asset and makes the previously unavailable carbon accessible for much wider range of microbes while substantially enhancing the carbon recovery. Importantly, not only does the acetate contribute to the growth of aerobic ADP1, but the aromatic compounds modified by A. woodii are also more accessible and better tolerated by ADP1.
By employing the coculture approach, we were able to achieve utilization of guaiacol for production of industrially interesting chemicals, which has traditionally been considered as a difficult task. We demonstrated a nearly equimolar production of MA from guaiacol by metabolic integration of A. woodii and APD1ΔcatBC. By the stringent integration of the metabolisms, the consortium was able to accomplish tasks neither bacterium could perform alone: guaiacol cannot be utilized by ADP1, and A. woodii cannot perform ring fission reaction required for MA production. The process required only minimal genetic engineering of ADP1ΔcatBC, namely, a single knock-out to accumulate MA, thus allowing for robust and stable production system. ADP1 also showed good tolerance and high catabolic activity toward catechol, a key intermediate in the system. In addition, the acetogenic O-demethylation of guaiacol coupled with CO2 fixation did provide not only precursor for the product, but also the growth substrate for ADP1, eliminating the need for using additional carbon sources. Such comprehensive utilization of carbon would not be possible using only aerobic production system. We further demonstrated that the MA production using this consortium would be possible in more easily operated one-pot bioreactor conditions. In these conditions, we reported a simultaneous activity of both bacteria, which opens intriguing possibilities for other production systems comprising both anaerobic and aerobic micro-organisms.
This study highlights and repurposes unique anaerobic characteristics and metabolic features that have been previously overlooked in the context of lignin valorization. By combining these features with aerobic metabolism in rationally designed synthetic cocultures, many challenges related to upgrading the recalcitrant feedstock can be tackled. Moreover, the concept of synergistically integrating metabolisms could be potentially applied across various microbial production platforms and to substrates beyond those derived from lignin.
Materials and methods
Strains and cultivations
Acinetobacter baylyi ADP1 (DSM 24193), A. baylyi ADP1ΔcatBC::tdk/kan, and Acetobacterium woodii (DSM 1030) were used in this study. ADP1 DSM24193 was used as a background strain for the knock-out strain with CatB (ACIAD1446) and CatC (ACIAD1447) deletions, referred here after as ADP1ΔcatBC. The molecular work for constructing the knock-out strain was done by using established methods. All the reagents and the primers were purchased from ThermoFisher Scientific (USA).
The three gene components for the knock-out cassette were first amplified separately by PCR as described previously [60] with following exceptions: the upstream flanking of the cassette was constructed by amplifying the region between CatB_P3 and CatB_P4 primers from the ADP1 genome. The downstream flanking was amplified from ADP1 genome by using the primers CatC_P5 and CatC_P6. The tdk/kanr [61] segment of the cassette, containing kanamycin resistance gene, was amplified from the genome of Acinetobacter baylyi ADP1Δacr1::tdk/kanr [62] (a kind gift of Veronique de Berardinis) by using the primers Tdk_kanF and Tdk_kanR. The knock-out cassette was constructed by using overlap extension PCR, and the complete cassette was amplified by using primers CatB_P3 and CatC_P6. The sequence of the obtained cassette and the used primers are presented in the supplementary material. The knock-out cassette was transformed to ADP1 as described previously [63]. The kanamycin concentration was 25 µg/mL.
Unless stated otherwise, ADP1 was precultivated in 5 mL of minimal salt medium (MSM) [64] containing K2HPO4 (3.88 g/L), NaH2PO4 (1.63 g/L), (NH4)2SO4 (2.00 g/L), MgCl2 ∙ 6 H2O (0.1 g/L), EDTA (10 mg/L), ZnSO4 ∙ 7 H2O (2 mg/L), CaCl2 ∙ 2 H2O (1 mg/L), FeSO4 ∙ 7 H2O (5 mg/L), Na2MoO4 ∙ 2 H2O (0.2 mg/L), CuSO4 ∙ 5 H2O (0.2 mg/L), CoCl2 ∙ 6 H2O (0.4 mg/L), and MnCl2 ∙ 2 H2O (1 mg/L), supplemented with 0.2% casamino acids and 0.4% d-glucose. For ADP1ΔcatBC precultivations, also 30 µg/mL kanamycin was added. ADP1 and ADP1ΔcatBC cultivations were done at + 30 ℃ and 300 rpm, unless stated otherwise. 96-well plate cultivations were done using Tecan Spark multimode microplate reader (Tecan, Switzerland) with 96-well µClear®plates (Greiner bio-one, Germany). The cultivation volume was 200 µL and the temperature was kept at + 30 ℃. Shaking amplitude was 2.5 mm with the frequency of 108 rpm, and the shaking duration was 180 s. Shaking was performed before and after the measurements of optical density at 600 nm, which occurred once in an hour.
Unless stated otherwise, A. woodii was cultivated in 12 mL of acetobacterium medium (DSMZ 135, referred as ABM) which was prepared according to DSMZ’s instructions. The medium was prepared under N2-CO2 80:20 headspace and the final composition of the medium was the following (g/L): NaHCO3 (11.04), D-Fructose (9.79), yeast extract (1.96), NH4Cl (0.98 g/L), L-Cysteine HCl ∙ H2O (0.49), Na2S ∙ 9 H2O (0.49), K2HPO4 (0.44), KH2PO4 (0.32), MgSO4 ∙ 7 H2O (0.06), nitrilotriacetic acid (0.03), NaCl (0.02), MnSO4 ∙ H2O (0.01), CoSO4 ∙ 7 H2O (0.004), ZnSO4 ∙ 7 H2O (0.004), CaCl2 ∙ 2 H2O (0.002), FeSO4 ∙ 7 H2O (0.002), NiCl2 ∙ 6 H2O (5.9e-4), Na-resazurin, (4.9e-4), AlK(SO4)2 ∙ 12 H2O (3.9e-4), H3BO3 (2.0e-4), Na2MoO4 ∙ 2 H2O (2.0e-4), CuSO4 ∙ 5 H2O (2.0e-4), pyridoxine hydrochloride (9.8e-5), nicotinic acid (4.9e-5), p-aminobenzoic acid (4.9e-5), (DL)-alpha-lipoic acid (4.9e-5), calcium D-( +)-pantothenate (4.9e-5), thiamine HCl (4.9e-5), riboflavin (4.9e-5), biotin (2.0e-5), folic acid (2.0e-5), Na2WO4 ∙ 2 H2O (7.8e-6), Na2SeO3 ∙ 5 H2O (5.9e-6), and vitamin B12 (9.8e-7). Unless stated otherwise, A. woodii cultivations were performed at + 30 ℃ and 150 rpm. The inoculations for every cultivation containing A. woodii cells were done from fructose-grown cultivations, which were less than 1 week old. The inoculation volume was 0.2 mL (resulting in a starting OD600 of 0.01–0.04), unless stated otherwise.
Flux balance analysis
FBA was done by using COBRApy Python package (version 0.28.0) [65]. The metabolic network model for ADP1 was obtained from Durot et al. [66] and it was used to examine the effects of using the products from anaerobic O-demethylation process as substrates. To perform that, new exchange reactions related to PCA intake were added to the model.
Cultivations
The growth test of ADP1 on 10 mM vanillate and on a mixture containing 10 mM PCA and 7.5 mM acetate as well as the test where ADP1 was cultivated on 0, 5, 7.5, 10, and 15 mM of ferulate, caffeate, dihydrocaffeate, coumarate, or phloretate were done on MSM in 96-well plate setting. The starting OD600 was set to be 0.1.
The growth of A. woodii on vanillate, ferulate, coumarate, and guaiacol was studied on ABM without fructose supplementation. For vanillate and guaiacol, the tested concentrations were 6, 12, and 18 mM. For ferulate and coumarate, the concentrations were 13 mM and 14 mM, respectively.
The utilization of guaiacol by ADP1 was studied with 96-well plate setting by cultivating ADP1 on 0, 2.5, and 5 mM of guaiacol in MSM. Also, the growth of ADP1 on 0, 0.5, 1, 2, 5, 7.5, and 10 mM catechol was studied in similar cultivation. In both studies, the starting OD600 was set to be 0.1.
The ability of ADP1 to grow on the catechol and acetate produced by A. woodii from guaiacol was determined in a cultivation study, where A. woodii was cultivated first on 12 mM guaiacol in a 12.5 mL volume without fructose supplementation. After all the guaiacol was demethylated into catechol, the cultivations were transformed into sterile 50 mL Falcon tubes with vented lids. After the pH was adjusted to 7 by using 5% HCl, ADP1 cells were added to the cultivation at OD600 0.1. The pH was adjusted again to 7 on the 2nd day of ADP1 cultivation phase.
The production of ccMA with ADP1ΔcatBC and A. woodii cocultivation was tested by first cultivating A. woodii on 11 mM guaiacol in ABM without fructose supplementation. After the anaerobic cultivation phase, the A. woodii cells were removed from the medium by centrifugation. To ensure that the catechol concentration would not be too high for ADP1ΔcatBC to grow and to prevent pH increase in aerobic phase caused by the presence of NaHCO3, we diluted the medium 1:1 with MQ and added MOPS buffer to a final concentration of 120 mM. In addition, 0.2% casamino acids were added to the cultivations to ensure that ADP1 would not suffer from the presence of l-cysteine [35]. Kanamycin concentration of 15 µg/mL was used. The final volume of the new medium was 15 mL. ADP1ΔcatBC cells were collected from liquid preculture by centrifugation, resuspended into MSM and added to the medium to obtain a starting OD600 0.3.
The three-step batch cocultivation was done by using Biostat B Plus bioreactor (Sartorius, Germany) equipped with pH-meter (Hamilton, Switzerland) and pO2-meter (Hamilton). Aerobically prepared low-NaHCO3 ABM (NaHCO3 concentration only 1.25 g/L) without fructose supplementation was used as a medium. The only carbon sources and electron donors added to the medium in the beginning were 10 mM glucose and 5 mM guaiacol. The initial cultivation volume was 555 mL. Before the addition of ADP1ΔcatBC cells, the medium was aired with pressured air for couple of hours. To obtain a required volume of ADP1ΔcatBC to reach the desired starting OD600 of 0.3, ADP1ΔcatBC was cultivated on similarly to the normal preculture, but without kanamycin and the cultivation volume was increased to 50 mL in flask. The cells were centrifugated and resuspended into 10 mL MSM. The pO2-value was calibrated to 100% just before the resuspended ADP1ΔcatBC cells were added to the reactor. The flow of the pressured air was stopped, and the reactor was sealed with clamps. The reactor was covered to protect the light-sensitive guaiacol. When pO2-value was 0.7%, A. woodii cells were added to the reactor, which initiated the anaerobic phase. HPLC results were used to determine the optimal starting time of the last aerobic phase. Throughout the cultivation, OD600 was measured in every 12 min by Arc view device (Hamilton) and manually from each sample taken. To minimize the effect of the resazurin indicator to the OD600 results, manually measured OD600 values were used as a starting point and a gradual step-by-step transformation from one value to the next one was done based on the tendencies of machine measured values. The pH was set to 7.0 for the whole cultivation and the required adjustments were done automatically by using concentrated NaOH and H3PO4 stocks.
Analytical techniques
Quantitative analysis of compounds was done using high-performance liquid chromatography (HPLC) instruments (Shimadzu, Japan) equipped with PDA and RID detectors.
Aromatic compounds were analyzed on Luna C18 column (150 × 4.6 mm, 5 µm) (Phenomenex, USA) at 40 ℃ using the mixture of MQ/methanol/formic acid (80/20/0.16 v/v/v) as the eluent at the flow rate 1 mL/min. The injection volume was 5 µL. Aromatic substances were detected at the wavelength 254 nm or 272 nm.
Sugar and organic acid were analyzed on Rezex RHM-Monosaccharide H + (8%) column (300 × 7.8 mm) (Phenomenex) at 40 ℃ using 5 mM H2SO4 as the eluent with the flow rate 0.5 mL/min. The injection volume was 10 µL. Compounds were detected by refractive index detector.
The obtainment of ccMA and ctMA in the two-stage MA production test with A. woodii and ADP1ΔcatBC was confirmed by LC–MS using Agilent 1260 Infinity II (USA) which was equipped with ultraviolet (UV) detector and connected with Jeol AccuTOF LC Plus (JMS-T100LP) (Japan). Compounds were analyzed on Poroshell 120 EC-C18 column (100 × 4.6 mm, 2.7 µm) (USA). The temperature of the column was + 30 ℃ and the injection volume was 2 µL. The mixture of 0.1% formic acid and methanol (80/20 v/v) was used as eluent with the flow rate 0.2 mL/min. Calculation of ccMA and ctMA was performed from extracted ion chromatograms and UV absorption chromatograms at 272 nm using external standard method. In both cases molar weight of MA was calculated as approximately 141 g/mol.
Availability of data and materials
The data supporting the findings of this study are available within the paper and its supplementary information files. The raw data and the additional data sets regarding this manuscript are available from the corresponding authors upon reasonable request.
Abbreviations
- ABM:
-
Acetobacterium medium
- ADP1:
-
Acinetobacter baylyi ADP1
- ccMA:
-
cis,cis-Muconate
- CCR:
-
Carbon catabolite repression
- ctMA:
-
cis,trans-Muconate
- FBA:
-
Flux balance analysis
- CDW:
-
Cell dry weight
- HPLC:
-
High-performance liquid chromatography
- LC–MS:
-
Liquid chromatography mass spectrometry MA Muconate
- OD600:
-
Optical density measured in 600 nm
- PCA:
-
Protocatechuate
- PHA:
-
Polyhydroxyalkanoate
- RO:
-
Rieske oxygenase
- TAG:
-
Triacylglyceride
- THF:
-
Tetrahydrofolate
- UV:
-
Ultraviolet
References
Guadix-Montero S, Sankar M. Review on catalytic cleavage of C-C inter-unit linkages in lignin model compounds: towards lignin depolymerisation. Top Catal. 2018;61(3):183–98.
Laurichesse S, Avérous L. Chemical modification of lignins: towards biobased polymers. Prog Polym Sci. 2014;39(7):1266–90.
Chio C, Sain M, Qin W. Lignin utilization: a review of lignin depolymerization from various aspects. Renew Sustain Energy Rev. 2019;1(107):232–49.
Ralph J, Lapierre C, Boerjan W. Lignin structure and its engineering. Curr Opin Biotechnol. 2019;1(56):240–9.
Wells T, Ragauskas AJ. Biotechnological opportunities with the β-ketoadipate pathway. Trends Biotechnol. 2012;30(12):627–37.
Beckham GT, Johnson CW, Karp EM, Salvachúa D, Vardon DR. Opportunities and challenges in biological lignin valorization. Curr Opin Biotechnol. 2016;1(42):40–53.
Xu Z, Lei P, Zhai R, Wen Z, Jin M. Recent advances in lignin valorization with bacterial cultures: microorganisms, metabolic pathways, and bio-products. Biotechnol Biofuels. 2019;12(1):32.
van Duuren JBJH, Wijte D, Karge B, Martins dos Santos VAP, Yang Y, Mars AE, et al. pH-stat fed-batch process to enhance the production of cis, cis-muconate from benzoate by Pseudomonas putida KT2440-JD1. Biotechnol Prog. 2012;28(1):85–92.
Becker J, Kuhl M, Kohlstedt M, Starck S, Wittmann C. Metabolic engineering of Corynebacterium glutamicum for the production of cis, cis-muconic acid from lignin. Microb Cell Fact. 2018;17(1):115.
Cai C, Xu Z, Xu M, Cai M, Jin M. Development of a Rhodococcus opacus cell factory for valorizing lignin to muconate. ACS Sustain Chem Eng. 2020;8(4):2016–31.
Kohlstedt M, Weimer A, Weiland F, Stolzenberger J, Selzer M, Sanz M, et al. Biobased PET from lignin using an engineered cis, cis-muconate-producing Pseudomonas putida strain with superior robustness, energy and redox properties. Metab Eng. 2022;1(72):337–52.
Okamura-Abe Y, Abe T, Nishimura K, Kawata Y, Sato-Izawa K, Otsuka Y, et al. Beta-ketoadipic acid and muconolactone production from a lignin-related aromatic compound through the protocatechuate 3,4-metabolic pathway. J Biosci Bioeng. 2016;121(6):652–8.
Luo J, Lehtinen T, Efimova E, Santala V, Santala S. Synthetic metabolic pathway for the production of 1-alkenes from lignin-derived molecules. Microb Cell Fact. 2019;18(1):48.
Salmela M, Lehtinen T, Efimova E, Santala S, Santala V. Alkane and wax ester production from lignin-related aromatic compounds. Biotechnol Bioeng. 2019;116(8):1934–45.
Salmela M, Lehtinen T, Efimova E, Santala S, Santala V. Towards bioproduction of poly-α-olefins from lignocellulose. Green Chem. 2020;22(15):5067–76.
Arvay E, Biggs BW, Guerrero L, Jiang V, Tyo K. Engineering Acinetobacter baylyi ADP1 for mevalonate production from lignin-derived aromatic compounds. Metab Eng Commun. 2021;24(13): e00173.
Luo J, McIntyre EA, Bedore SR, Santala V, Neidle EL, Santala S. Characterization of highly ferulate-tolerant Acinetobacter baylyi ADP1 isolates by a rapid reverse engineering method. Appl Environ Microbiol. 2022;88(2):e01780-e1821.
Eltis LD, Karlson U, Timmis KN. Purification and characterization of cytochrome P450RR1 from Rhodococcus rhodochrous. Eur J Biochem. 1993;213(1):211–6.
Sutherland JB. Demethylation of veratrole by cytochrome P-450 in Streptomyces setonii. Appl Environ Microbiol. 1986;52(1):98–100.
García-Hidalgo J, Ravi K, Kuré LL, Lidén G, Gorwa-Grauslund M. Identification of the two-component guaiacol demethylase system from Rhodococcus rhodochrous and expression in Pseudomonas putida EM42 for guaiacol assimilation. AMB Express. 2019;11(9):34.
Tumen-Velasquez M, Johnson CW, Ahmed A, Dominick G, Fulk EM, Khanna P, et al. Accelerating pathway evolution by increasing the gene dosage of chromosomal segments. Proc Natl Acad Sci U S A. 2018;115(27):7105–10.
Mallinson SJB, Machovina MM, Silveira RL, Garcia-Borràs M, Gallup N, Johnson CW, et al. A promiscuous cytochrome P450 aromatic O-demethylase for lignin bioconversion. Nat Commun. 2018;9(1):2487.
Zhang Z, Wang Y, Zheng P, Sun J. Promoting lignin valorization by coping with toxic C1 byproducts. Trends Biotechnol. 2021;39(4):331–5.
Otsuka Y, Araki T, Suzuki Y, Nakamura M, Kamimura N, Masai E. High-level production of 2-pyrone-4,6-dicarboxylic acid from vanillic acid as a lignin-related aromatic compound by metabolically engineered fermentation to realize industrial valorization processes of lignin. Bioresour Technol. 2023;1(377):128956.
Erickson E, Bleem A, Kuatsjah E, Werner AZ, DuBois JL, McGeehan JE, et al. Critical enzyme reactions in aromatic catabolism for microbial lignin conversion. Nat Catal. 2022;5(2):86–98.
Brunel F, Davison J. Cloning and sequencing of Pseudomonas genes encoding vanillate demethylase. J Bacteriol. 1988;170(10):4924–30.
Notonier S, Werner AZ, Kuatsjah E, Dumalo L, Abraham PE, Hatmaker EA, et al. Metabolism of syringyl lignin-derived compounds in Pseudomonas putida enables convergent production of 2-pyrone-4,6-dicarboxylic acid. Metab Eng. 2021;1(65):111–22.
Segura A, Bünz PV, D’Argenio DA, Ornston LN. Genetic analysis of a chromosomal region containing vanA and vanB, genes required for conversion of either ferulate or vanillate to protocatechuate in Acinetobacter. J Bacteriol. 1999;181(11):3494–504.
Chen NH, Djoko KY, Veyrier FJ, McEwan AG. Formaldehyde stress responses in bacterial pathogens. Front Microbiol. 2016;3(7):257.
Almqvist H, Veras H, Li K, Garcia Hidalgo J, Hulteberg C, Gorwa-Grauslund M, et al. Muconic acid production using engineered Pseudomonas putida KT2440 and a guaiacol-rich fraction derived from kraft lignin. ACS Sustain Chem Eng. 2021;9(24):8097–106.
Drake HL, Gößner AS, Daniel SL. Old Acetogens, New Light. Ann N Y Acad Sci. 2008;1125(1):100–28.
Schuchmann K, Müller V. Energetics and application of heterotrophy in Acetogenic Bacteria. Appl Environ Microbiol. 2016;82(14):4056–69.
Ricci L, Seifert A, Bernacchi S, Fino D, Pirri CF, Re A. Leveraging substrate flexibility and product selectivity of acetogens in two-stage systems for chemical production. Microb Biotechnol. 2023;16(2):218–37.
Ma X, Liang H, Panda S, Fung VKY, Zhou JFJ, Zhou K. C2 feedstock-based biomanufacturing of value-added chemicals. Curr Opin Biotechnol. 2022;1(73):240–5.
Lehtinen T, Efimova E, Tremblay PL, Santala S, Zhang T, Santala V. Production of long chain alkyl esters from carbon dioxide and electricity by a two-stage bacterial process. Bioresour Technol. 2017;1(243):30–6.
Lehtinen T, Virtanen H, Santala S, Santala V. Production of alkanes from CO2 by engineered bacteria. Biotechnol Biofuels. 2018;11(1):228.
Katsyv A, Müller V. Overcoming energetic barriers in acetogenic C1 conversion. Front Bioeng Biotechnol. 2020;23:8.
Sharak Genthner BR, Bryant MP. Additional characteristics of one-carbon-compound utilization by Eubacterium limosum and Acetobacterium woodii. Appl Environ Microbiol. 1987;53(3):471–6.
Khomyakova MA, Slobodkin AI. Transformation of methoxylated aromatic compounds by anaerobic microorganisms. Microbiol. 2023;92(2):97–118.
Kremp F, Müller V. Methanol and methyl group conversion in Acetogenic bacteria: biochemistry, physiology and application. FEMS Microbiol Rev. 2021;45(2):fuaa040.
Frazer AC, Young LY. Anaerobic C1 metabolism of the O-Methyl-14C-Labeled substituent of vanillate. Appl Environ Microbiol. 1986;51(1):84–7.
Bache R, Pfennig N. Selective isolation of Acetobacterium woodii on methoxylated aromatic acids and determination of growth yields. Arch Microbiol. 1981;130(3):255–61.
Müller V. Energy conservation in Acetogenic Bacteria. Appl Environ Microbiol. 2003;69(11):6345–53.
Santala S, Santala V. Acinetobacter baylyi ADP1-naturally competent for synthetic biology. Essays Biochem. 2021;65(2):309–18.
Schmidt S, Biegel E, Müller V. The ins and outs of Na+ bioenergetics in Acetobacterium woodii. Biochim Biophys Acta. 2009;1787(6):691–6.
Fischer R, Bleichrodt FS, Gerischer UC. Aromatic degradative pathways in Acinetobacter baylyi underlie carbon catabolite repression. Microbiol. 2008;154(10):3095–103.
Bleichrodt FS, Fischer R, Gerischer UC. The β-ketoadipate pathway of Acinetobacter baylyi undergoes carbon catabolite repression, cross-regulation and vertical regulation, and is affected by Crc. Microbiol. 2010;156(5):1313–22.
Dilling S, Imkamp F, Schmidt S, Müller V. Regulation of caffeate respiration in the Acetogenic Bacterium Acetobacterium woodii. Appl Environ Microbiol. 2007;73(11):3630–6.
Stuani L, Lechaplais C, Salminen AV, Ségurens B, Durot M, Castelli V, et al. Novel metabolic features in Acinetobacter baylyi ADP1 revealed by a multiomics approach. Metabolomics. 2014;10(6):1223–38.
Kalil MS, Stephens GM. Catechol production by O-demethylation of 2-methoxyphenol using the obligate anaerobe Acetobacterium woodii. Biotechnol Lett. 1997;19(12):1165–8.
Kalil MS, Asyiqin Z, Zaki M. Catechol synthesis via demethylation of guaiacol by Anaerobic Bacterium Acetobacterium woodii DSM 1030. Pak J Biol Sci. 2002;5(11):1186–8.
Kohlstedt M, Starck S, Barton N, Stolzenberger J, Selzer M, Mehlmann K, et al. From lignin to nylon: cascaded chemical and biochemical conversion using metabolically engineered Pseudomonas putida. Metab Eng. 2018;1(47):279–93.
Barton N, Horbal L, Starck S, Kohlstedt M, Luzhetskyy A, Wittmann C. Enabling the valorization of guaiacol-based lignin: Integrated chemical and biochemical production of cis, cis-muconic acid using metabolically engineered Amycolatopsis sp ATCC 39116. Metab Eng. 2018;1(45):200–10.
Salmela M, Lehtinen T, Efimova E, Santala S, Mangayil R. Metabolic pairing of aerobic and anaerobic production in a one-pot batch cultivation. Biotechnol Biofuels. 2018;3(11):187.
Mangayil R, Efimova E, Konttinen J, Santala V. Co-production of 1,3 propanediol and long-chain alkyl esters from crude glycerol. New Biotechnol. 2019;25(53):81–9.
Salvachúa D, Johnson CW, Singer CA, Rohrer H, Peterson DJ, Black BA, et al. Bioprocess development for muconic acid production from aromatic compounds and lignin. Green Chem. 2018;20(21):5007–19.
Sonoki T, Takahashi K, Sugita H, Hatamura M, Azuma Y, Sato T, et al. Glucose-free cis, cis-muconic acid production via new metabolic designs corresponding to the heterogeneity of lignin. ACS Sustain Chem Eng. 2018;6(1):1256–64.
Xie NZ, Liang H, Huang RB, Xu P. Biotechnological production of muconic acid: current status and future prospects. Biotechnol Adv. 2014;32(3):615–22.
Sonoki T, Morooka M, Sakamoto K, Otsuka Y, Nakamura M, Jellison J, et al. Enhancement of protocatechuate decarboxylase activity for the effective production of muconate from lignin-related aromatic compounds. J Biotechnol. 2014;20(192):71–7.
Santala S, Efimova E, Kivinen V, Larjo A, Aho T, Karp M, et al. Improved triacylglycerol production in Acinetobacter baylyi ADP1 by metabolic engineering. Microb Cell Fact. 2011;10(1):36.
Metzgar D, Bacher JM, Pezo V, Reader J, Döring V, Schimmel P, et al. Acinetobacter sp. ADP1: an ideal model organism for genetic analysis and genome engineering. Nucleic Acids Res. 2004;32(19):5780.
de Berardinis V, Vallenet D, Castelli V, Besnard M, Pinet A, Cruaud C, et al. A complete collection of single-gene deletion mutants of Acinetobacter baylyi ADP1. Mol Syst Biol. 2008;4:174.
Bedore SR, Neidle EL, Pardo I, Luo J, Baugh AC, Duscent-Maitland CV, et al. Chapter 8—Natural transformation as a tool in Acinetobacter baylyi streamlined engineering and mutational analysis. Methods Microbiol Genome Eng. 2023;52:207–34.
Hartmans S, Smits JP, van der Werf MJ, Volkering F, de Bont JAM. Metabolism of styrene oxide and 2-phenylethanol in the styrene-degrading Xanthobacter Strain 124X. Appl Environ Microbiol. 1989;55(11):2850–5.
Ebrahim A, Lerman JA, Palsson BO, Hyduke DR. COBRApy: COnstraints-based reconstruction and analysis for python. BMC Syst Biol. 2013;7:74.
Durot M, Le Fèvre F, de Berardinis V, Kreimeyer A, Vallenet D, Combe C, et al. Iterative reconstruction of a global metabolic model of Acinetobacter baylyi ADP1 using high-throughput growth phenotype and gene essentiality data. BMC Syst Biol. 2008;7(2):85.
Acknowledgements
The authors are grateful to Dr. Pauli Losoi for his help with FBA of ADP1, Dr. Milla Salmela for constructing the knock-out strain A. baylyi ADP1ΔcatBC, and Henna Ovaska BSc (Tech) for her contributions in constructing the correlation curve between OD600 and CDW. Figures 1A and 5 were created with BioRender.com.
Funding
Open access funding provided by Tampere University (including Tampere University Hospital). This work was financially supported by the Research Council of Finland (grant numbers 347204, 353587). SS and VS would also like to thank the Novo Nordisk Foundation (grants NNF21OC0067758, NNF22OC0079579).
Author information
Authors and Affiliations
Contributions
S.S., V.S., and E.M. designed the experimental set-up. E.M. performed the experimental work, interpreted the data, and wrote the original draft of the manuscript. E.E contributed to metabolite analyses and performed LC–MS analysis. S.S. supervised the study. E.M., V.S., and S.S. participated in reviewing and editing the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
13068_2024_2526_MOESM1_ESM.docx
Supplementary Material 1. Cultivating A. woodii on vanillate as a sole organic carbon source. Cultivating A. woodii on ferulate and coumarate as a sole organic carbon source. Cultivating A. woodii on guaiacol. Sequence of the knock-out cassette. Table S1: Stoichiometric equations of vanillate, syringate, and ferulate utilization by A. woodii according to Bache and Pfennig. Table S2: The production of PCA and acetate by A. woodii when cultivated on vanillate as a sole organic carbon source. Standard deviations describe the differences between the duplicates. Table S3: The production of caffeate, dihydrocaffeate, and acetate by A. woodii when cultivated on ferulate as a sole organic carbon source. Standard deviations describe the differences between the duplicates. Table S4: The production of phloretate and acetate by A. woodii when cultivated on coumarate as a sole organic carbon source. Standard deviations describe the differences between the duplicates. Table S5: The production of catechol and acetate by A. woodii when cultivated on guaiacol as a sole organic carbon source. The standard deviations describe the differences between the duplicates. Table S6: List of the used primers and their names. Figure S1: A correlation curve between OD600 and CDW in ADP1. Figure S2: A A. woodii cultivated in acetobacterium medium with A ferulate or B coumarate as a sole organic carbon source. The fructose present at the beginning of the cultivation is a carry-over substrate coming from the inoculation. Error bars in both graphs are describing the differences between the duplicates. The error bars indicate the standard deviations from the average values of the biologically independent duplicates. In some cases, the error bars are smaller than the size of the marker. Figure S3: The growth of ADP1 on ferulate or caffeate and dihydrocaffeate modeling the aromatic metabolites obtained when A. woodii is cultivated on ferulate. Used concentrations are A 5 mM, B 7.5 mM, and C 10 mM. The error bars indicate the standard deviations from the average values of the biologically independent triplicates. In some cases, the error bars are smaller than the size of the marker. Figure S4: The growth of ADP1 on coumarate or phloretate modeling the aromatic metabolites obtained when A. woodii is cultivated on coumarate. Used concentrations are A 5 mM, B 7.5 mM, and C 10 mM. The error bars indicate the standard deviations from the average values of the biologically independent triplicates. In some cases, the error bars are smaller than the size of the marker. Figure S5: The growth of ADP1 in MSM with catechol as a sole carbon source. The cultivation volume was 200 µl. The linear increase of OD600 in 10 mM cultivations was caused by the color change of catechol in the medium, and therefore, it is not representing the accumulation of biomass. The error bars indicate the standard deviations from the average values of the biologically independent triplicates. In some cases, the error bars are smaller than the size of the marker. Figure S6: Experimental data of the three-phase one-pot coculture of A. woodii and ADP1ΔcatBC. In this graph, only the utilization and production of sugars and organic acids are presented. Glucose is supplemented in the medium at the beginning of the cultivation. Fructose and acetate peaks at the first dashed lined are caused by the inoculation of A. woodii. Please notice that compared to Fig. 5, the presented acetate concentration here describes the overall concentration of acetate, not only the acetate produced by A. woodii. In these growth conditions, ADP1 consumed acetate faster than glucose during the aerobic phase, which is potentially related to the complex medium composition and a long oxygen-depletion period; previously, ADP1 has been reported to consume acetate and glucose simultaneously (5). DO = deoxygenation phase. Figure S7: Consumption of catechol and acetate by ADP1 WT. The cultivation volume was 30 mL. The error bars indicate the standard deviations from the average values of the biologically independent duplicates. In some cases, the error bars are smaller than the size of the marker. One of the duplicate cultivations was lost after 7 h, and thus, the rest of the data points are describing values obtained from single cultivation.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Meriläinen, E., Efimova, E., Santala, V. et al. Carbon-wise utilization of lignin-related compounds by synergistically employing anaerobic and aerobic bacteria. Biotechnol Biofuels 17, 78 (2024). https://doi.org/10.1186/s13068-024-02526-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13068-024-02526-0