
Abstract
Background
Chronic Granulomatous Disease (CGD) is a primary immunodeficiency that causes susceptibility to recurrent fungal and bacterial infections. The CYBB gene encodes gp91phox component of the Phagocytic Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase and specifically, X-linked CGD is caused by mutations in the CYBB gene, located on the X chromosome. The aim of the study was to characterize functional and genetic mutations in X-linked CGD.
Methods
Functional analysis was conducted on the whole blood of seventeen male individuals who were suspected to have X-linked chronic granulomatous disease (CGD). Flow cytometry was employed to assess the capacity of NADPH oxidase, measuring both H2O2 production and gp91phox protein expression in neutrophils. Additionally, DNA Sanger sequencing was performed for genetic analysis. The pathogenicity of novel mutations was assessed by pathogenicity prediction tools.
Result
Among the seventeen patients evaluated, five patients (P1, P2, P3, P4, and P5) displayed impaired H2O2 production by their neutrophils upon stimulation with Phorbol myristate acetate (PMA), accompanied by abnormal gp91phox expression. DNA sequencing of the CYBB gene identified specific mutations in each patient. In P1 and P2 (previously reported cases), a hemizygous missense mutation, c.925G > A/p.E309K was identified. In P3 and P4 (novel cases), hemizygous nonsense mutations, c.216T > A/p.C72X were found. Lastly, in P5 (also a novel case), a hemizygous missense mutation, c.732T > G/p.C244W was detected. These mutations reside in exons 9,3 and 7 of the CYBB gene, respectively.
Conclusions
The current study contributes to the understanding of the clinical and genetic spectrum associated with X-linked chronic granulomatous disease (CGD). It highlights the significance of early diagnosis in CGD and emphasizes the importance of lifelong prophylaxis to prevent severe infections.
Similar content being viewed by others


Background
The Phagocytic Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase plays a vital role in eliminating intracellular pathogens by generating microbicidal reactive oxygen species (ROS) [1, 2]. Under resting conditions, NADPH oxidase is comprised of three cytosolic proteins and two membrane proteins, which are encoded by five structural genes (CYBB, CYBA, NCF1, NCF2, and NCF4). The membrane proteins, p22phox, and gp91phox, are encoded by CYBA and CYBB genes, respectively. On the other hand, the cytosolic proteins, p47phox, p67phox, and p40phox, are encoded by NCF1, NCF2, and NCF4 respectively [3, 4]. Upon pathogens stimulation, the cytosolic proteins (p47phox, p67phox, and p40phox) associated with RAC2 combine with p22phox and gp91phox to form an active NADPH oxidase complex that produces ROS [5].
Mutations in the five structural genes (NCF1, NCF2, NCF4, CYBA, CYBB) are associated with chronic granulomatous disease (CGD), a condition characterized by impaired production of reactive oxygen species (ROS) [2]. In CGD, the defective phagocytic NADPH oxidase-killing ability of pathogens results in chronic fungal and bacterial infections, leading to the formation of characteristic granulomas, a hallmark of the disease [3]. Notably, recurrent life-threatening infections in CGD are often caused by pathogens such as S. aureus, Aspergillus species, Salmonella species, Klebsiella species, mycobacteria spp., and Burkholderia cepacia [6].
CGD was first identified as an X-linked/autosomal recessive disease in early 1950s [4]. Among the various forms of CGD, the X-linked form of CGD is the most common (∼ 65%) and is caused by mutations in the CYBB gene positioned on the X-Chromosome. This gene encodes the gp91phox protein, also known as NOX2 [7]. Autosomal recessive CGD (ARCGD) results from mutations in CYBA (∼ 5%), NCF1(∼ 20% ), NCF2 (∼ 5%), or NCF4 (1%) genes [6, 8]. Three variants of X-linked CGD have been characterized so far: X910(complete absence of gp91phox expression and NADPH oxidase activity), X91−( reduced gp91phox expression and NADPH oxidase activity), and X91+(normal gp91phox expression with residual NADPH oxidase activity first). These variants resulting from heterogenous hemizygous mutations that differently affecting the function and expression of gp91phox protein [9].
Lack of medical knowledge, limited awareness about CGD, and the absence of diagnostic centers in many countries, including Pakistan, contribute to the delay in diagnosing CGD [10, 11]. Early and accurate diagnosis of CGD, particularly X-linked CGD, is crucial for the prognosis of affected individuals. Any delay in diagnosis hinders the timely initiation of specific treatments for CGD patients, leading to poor prognosis and an increased risk of mortality in these individuals.
In Pakistan, physicians rarely consider genetic diagnosis for CGD, and diagnosis primarily relies on clinical phenotypes. However, the clinical overlap among different conditions often makes it challenging to establish a definitive diagnosis. While there is extensive research on the molecular diagnosis of CGD patients globally, limited information is available regarding the molecular diagnosis of CGD patients in Pakistan. therefore, the purpose of our study was to investigate and characterize genetic alterations in CYBB gene in Pakistani patients using biochemical and genetic analyses.
Methods
Ethical statements and patients
This study included seventeen male patients with strong suspicion of X-linked chronic granulomatous disease (XCGD). All patients were confirmed to be HIV-negative. The collection of blood samples followed the approved protocol after taking consent from either the patients themselves or their parents. Blood samples were obtained from both the patients and control subjects. The collected blood samples were used for functional assessment as well as genetic analysis. This study has obtained approval from the ethical committee of Kohat University of Science and technology (KUST), Pakistan.
Dihydrorhodamine- 123 assay for quantification of H2O2
The oxidative burst, specifically the production of H2O2, in neutrophils was analyzed following a previously published protocol [12]. Briefly, a total of 5 ml of blood was collected from both healthy controls and patients in heparinized tubes. The whole blood samples were then stimulated with phorbol myristate acetic acid (PMA) at a concentration of 300 ng/ml for a duration of 30 min. Following this, the samples were incubated with the dihydroergotamine (DHR) 123 assay at 370 C for 20 min. The Whole blood was then lysed using RBC lysis solution at room temperature, and the cells were subjected to two washes with PBS. subsequently, the cells were fixed using a 2% paraformaldehyde solution. Flow cytometry analysis was performed using a BD FACSCanto II flow cytometer. The data obtained were analyzed using FlowJo software (Treestar, Inc, Ashland, Ore).
Flow cytometric analysis of NADPH oxidase complex
Flow cytometric analysis was performed to evaluate the expression of NADPH oxidase protein (gp91phox),as previously described [13, 14]. To specifically identify the membrane-associated NADPH oxidase (gp91phox) protein, neutrophils were incubated with anti-gp91phox antibody and subsequently washed twice with PBS. The fixation process was carried out in PBS comprising 1% paraformaldehyde, and the analysis was performed using a BD FACSCanto II flow cytometer. To identify the components of cytoplasmic proteins, the cells were permeabilized and fixed using the Cytoperm/Cytofix Plus kit, as per the manufacturer’s instructions. Subsequently, the cells were stained for 30 min at 4 °C with anti-p40phox, anti-p47phox, and anti-p67phox antibodies. After two washes with PBS, the cells were analyzed using a BD FACSCanto II flow cytometer. Data analysis was conducted using FlowJo software (Treestar, Inc, Ashland, Ore).
CYBB gene sequence analysis
Genomic DNA (gDNA) was extracted from ethylenediaminetetraacetic acid (EDTA)-chelated blood samples using the Wizard® Genomic DNA Purification kit from Promega. Polymerase chain reaction (PCR) was performed to amplify a total of 13 exons of CYBB, including exon-intron boundaries. The DYEnamic ET Terminator Cycle Sequencing Kit was utilized to sequence the PCR amplified products using the MegaBACE 1000 sequencer.
Assessment of pathogenic genetic variations in CYBB gene
Pathogenicity-predicating tools such as Polyphen-2, SIFT, and Mutation Taster were utilized to evaluate the mutation pathogenicity [15,16,17].
Evaluation of multiple sequence alignment and 3D structure
Sequences of gp91phox from various organisms were obtained from the protein database (www.uniprot.org) and was multiple-aligned with gp91phox mutants employing clustal X [18]. Comparative structure of full-length human gp91phox (with a missing sequence of 200–280 amino acids) were constructed through program MODELLER 9v19 [18] while taking gp91phox from Cylindrospermum stagnale and human as template (PDB id: 5O0T, 5O0X and 3A1F) [19]. The Output of MODELLER contained 3D model for the target sequence (wild type and mutants of gp9phox) were used with the entire non-hydrogen side as well as main chain atoms. We have developed various comparative models for every wild type and mutants gp91phox and the PROCHECK and ProSA programs were utilized to choose the best structure [20].
Evaluation of M. tuberculosis growth in monocytes derived macrophages
We further assessed the phagocytic capacity of monocyte-derived macrophages (MDMs) from both NOX2- deficient patients and control. We also examined their ability to control the growth of M. tuberculosis in response to in vitro treatment with recombinant human interferon gamma (rhIFN), following the previously described protocol [21].
Statistical analysis
Statistical analysis was conducted using GraphPad PRISM 5.00 software from GraphPad Software (San Diego, CA, USA). An unpaired t-test was employed for the analysis, and a significant level of P < 0.05 was used to determine statistical significance.
Results
Clinical presentations in X-linked CGD patients
Among the Pakistani XCGD patients (P1, P2, P3, P4, and P5), a higher frequency of BCG complications (23.53%) was observed. Other clinical presentations included Liver abscess and Otitis media (11.65%), Urinary tract infection and Pneumonia (11.76%), diarrhea, Tuberculosis (TB), Septicemia, Pulmonary aspergillosis, Skin infection, and Lymphadenitis 5.88% each (Fig. 1).

Map of study areas, 4 CGD patients belongs to district Mardan, 8 were from district Peshawar, 3 from district Kohat and 2 from district Karak. Percentage load of infections in clinically suspected patients
The X-linked CGD patients displayed impaired oxidative burst and NOX2 expression
The DHR-123 test was conducted to measure H2O2 level in 17 male CGD patients. Among them, five patients (P1, P2, P3, P4 and P5) exhibited reduced H2O2 production in their neutrophils (Fig. 2). Flow cytometric analysis was conducted to evaluate the expression of the gp91phox component of NADPH oxidase. The results indicated that only two patients (P3 and P4) had the X910 CGD variant, characterized by the absence of gp91phox expression and the complete loss of NADPH oxidase activity. On the other hand, three patients (P1, P2 and P5) exhibited the X91-CGD variant, characterized by low levels of gp91phox expression and a corresponding reduction in H2O2 production compared to healthy controls. It should be noted that the presented data in Fig. 2B only represents the NADPH expression (gp91phox) of the genetically characterized patients, while the other components of NADPH oxidase (p22phox, p47phox, p67phox, and p40phox) were found to be normal (Data not shown).

(A) show reduced H2O2 by NADPH oxidase of patients (P1, P2, P3, P4 and P5) in comparison to healthy controls, similarly (B) flow cytometric analysis of gp91phox in neutrophils of P1, P2, P3, P4, and P5 reveal aberrant expression respectively. The unfilled circles showing the control group of the same ages while the black filled circle representing the XCGD patients
Genetic analysis of XCGD patients revealed heterogeneous variations in the CYBB gene
After suspecting X-linked CGD in male patients with abolished H2O2 production, we conducted genetic analysis to investigate possible CYBB gene alterations. The hemizygous mutations were identified in the CYBB gene through genetic analysis. Among the patients, three individuals (P1/P2 with c.925G > A/p. E309K mutation, and P5 with c.732T > G/p.C244W mutation demonstrated missense mutations, while 2 patients (P3 and P4) exhibited nonsense mutations (c.216T > A/p.C72X) in the CYBB gene (Table 1; Figs. 3, 4 and 5).These identified mutations were found to be located in in exon 3, exon 7, and exon 9 of the CYBB gene (Figs. 3C, 4 and 5 C). It is worth mentioning that missense mutation (c.925G > A/p. E309K) has been previously reported [22], while based on our search, novel genetic variations (c.925G > A/p and c.732T > G/p.C244W) were not found in genetic variation databases like ExAC, LSDB, 1000G, Ensemble, or CYBB mutation database, indicating that these mutations are novel.

(A), (B), and (C) showing family pedigree of P1/P2 chromatogram and location of mutation (c.925G > A/p. E309K) in exon 9 respectively while (D) and (E) representing residue p. E309 across the species and 3D structure analysis of wild and mutant type gp91phox respectively, (F), (G) and (H) displaying the prediction tools (Polyphen-2, Mutation Taster and SIFT). The black filled squares represent patients

(A), (B), and (C) showing family pedigree of P2/P3, chromatogram and location of mutation (c.216T > A/p.C72X) in exon 3 respectively while (D) and (E) indicating residue p.C72 across the species and 3D structure analysis of wild and mutant type p.C72X gp91phox respectively. (F) indicating the Mutation Taster result. The black filled squares represent patients

(A), (B), and (C) presenting family pedigree of P5, chromatogram and location of mutation (c.732T > G/p.C244W) in exon 7 respectively while (D) and (E) representing residue p.C244W across the species and 3D structure analysis of wild and mutant type p.C244W gp91phox respectively, (F), (G), and (H) representing the prediction tools (Polyphen-2, Mutation Taster and SIFT). The black filled squares represent patients
In Silico analyses predicting that sequence variations affect the NAPDH oxidase activity
The multiple alignments of the identified mutations i.e.C244W, E309K, and C72X revealed that they are in highly conserved motifs of the NOXs (Figs. 3D, 4D and 5D). This suggests that these mutations may have an impact on the function of gp91phox. The mutations such as E309K are residing in the FAD binding pocket, which could disrupt the FAD binding to the gp91phox and hence its functionality. Additionally, the nonsense mutation C72X lead to a truncated protein which may completely abolished NADPH oxidase activity. Computational models of gp91phox further support our functional and genetic analyses, showing the presence of a truncated protein (Figs. 3E, 4E and 5E).
Mutation pathogenicity tools evaluation suggesting that sequence alterations are pathogenic
To assess the pathogenicity of the identified mutations, the we utilized mutation prediction tools like Polyphen-2, Mutation Taster and SIFT. The results from these tools consistently indicated that the sequence variations found in our study are disease-causing and thus pathogenic, as shown in Fig. 3 (F), Fig. 4 (F-G), and Fig. 5 (F-G). The genetic variations observed in the CYBB gene are strongly associated with the presence of CGD, other possibilities cannot be completely ruled out at this time, such as the presence of a genetic mutation in a gene not yet linked to CGD. Further investigation is needed to gain a comprehensive understanding of the underlying mechanisms and potential contributions of other factors.
rhIFN-γ improved the phagocytic activity of monocytes derived macrophages
The study findings indicate that MDMs from patients with functional deficiency of gp91phox exhibit impaired control over the proliferation of Mycobacterium tuberculosis (M. tuberculosis). However, when these MDMs were treated with recombinant human interferon (rhIFN-γ), the MDMs derived from patients with gp91phox functional deficiency exhibited improved capability in controlling the proliferation of M. tuberculosis compared to MDMs from healthy controls (Fig. 6). This finding suggests that the administration of interferon gamma may have a positive impact on enhancing the immune response against M. tuberculosis in patients with gp91phox functional deficiency (Fig. 6). It is worth mentioning that our this experiment needs further validation using more rhIFN-γ treated MDMs from X-linked CGD patients with similar underlying mutations to characterize the exhibited improved capability in controlling the proliferation of M. tuberculosis.

Defective control of the proliferation of M. tuberculosis by MDMs from gp91phox functional deficient patients is improved by rhIFN-γ. The results of untreated (-) and rhIFN-γ treated (rhIFN-γ) MDMs are shown (A) Phagocytosis, (B) M. tuberculosis growth index. The bacterial proliferation index was determined based on the ratio of the CFU numbers on day 6 to the CFU number on day 0 after the challenge (Control N = 5 and Patients N = 5). - indicating without rhIFN-γ treatment of MDMs
Discussion
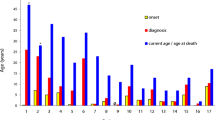
We conducted an analysis of underlying mutations, specifically (c.925G > A/ p.E309K) in exon number 9, (c.216T > A/p.C72X) in exon 3, (c.732T > G/p.C244W) in exon 7, within the CYBB gene. These mutations were observed in individuals who exhibited various clinical manifestations, notably severe bacterial and fungal infections, including mycobacteriosis. Our investigation encompassed genomic, in silico, and functional analyses, which indicated that these novel genetic variants contribute to the pathogenesis process by potentially affecting the NADPH oxidase activity. The average age of the XCGD patients in our study was 9 years old at the time of diagnosis. Our data suggest that the residues involved in the sequence variations are highly conserved across different species and are typically absent in healthy individuals within the same population as the patients. Nonetheless, further research is necessary to establish a connection between these novel mutations and the mechanisms underlying pathogenesis, particularly in relation to the abnormal function of NADPH oxidase.
The clinical features of X-linked NOX2-deficient CGD are presented with earlier onset of recurrent microbial and fungal infections and aberrant inflammation leading to early identification of the disease during childhood [24, 25]. In accordance with this, the patients in our study established characteristic CGD phenotypes during infancy. In various cases of gp91phox deficiency, the mycobacterial infections like tuberculosis have been reported [3, 26]. Importantly, the immunization of newborns in Pakistan are mandatory with BCG at birth against the protection of TB. In fact, one of our patients exhibited adverse BCG vaccination reaction and later developed tuberculosis. BCGitis occurred in 23.53% of our patients with tuberculosis, which is almost similar to the other published data [27, 28] and lower than Iranian patients (56%) [29]. It is presumed that such differences could be due to the type of BCG strain, age at the time of vaccination, amount of BCG dosages and type of mutations. Pulmonary infections were the common clinical phenotype of XCGD in our patients, which is in accordance with previous reports [8, 24, 29, 30].
Staphylococci and Aspergillus are the main causal agents found in CGD [31], which were also the causal pathogens in some of our CGD patients. In various countries, the limited medical awareness about primary immunodeficiencies (PID) is the most apparent reason of late diagnosis of the disease [10, 11, 32,33,34]. Globally, the average age of diagnosis for patients with XCGD is 5 years [25], in contrast to this, our cohort exhibited a slightly higher mean age of diagnosis, which was 9 years, suggesting late diagnosis of CGD in Pakistan. Other factors may also contribute to the late identification of the CGD in our patients since clinicians in Pakistan tend to focus on signs and symptoms (e.g., diarrhea, sinusitis, TB, pneumonia, and bronchitis) rather than the underlying PID.
Various subtypes of X-linked chronic granulomatous disease (XCGD) can arise from different types of mutations in the CYBB gene. The specific mutations can give rise to distinct variants of XCGD with varying clinical presentations and severity [32]. In our cohort of XCGD patients, mutations in CYBB gene led to the two variants of XCGD (X91o and X91−). Similarly, missense mutations in the CYBB gene of P1, 2 and P5, caused low expression of NOX2 (X91− variant) with impaired activity of NADPH oxidase, while nonsense mutations in CYBB gene of P3 and P4 resulted in the absence of NOX2 expression (X910 variant).The identification of mutations: c.925G > A/p.E309K in our two siblings (P1/P2) has been previously reported to affect both ROS production and gp91phox expression, as in accordance with [22, 23]. Most reported mutations are found throughout the 13 exons of the CYBB gene [35].
Accordingly, we identified the mutations, (c.216T > A/p.C72X, c.216T > A/p.C72X, c.925G > A/ p.E309K and c.732T > G/p.C244W) which are residing in exon 3, exon 7, and exon 9 of CYBB gene respectively. Our in silico and functional analyses suggest that these novel mutations (c.216T > A/p.C72X, c.216T > A/p.C72X, and c.732T > G/p.C244W) affect the respiratory burst of NADPH oxidase. However, transgenic experiments are necessary to determine the impact of these new variants on the assembly of NOX2 with other proteins and the functions of NADPH oxidase. Bionda and her colleagues employed a similar strategy to effectively exploit missense mutations in the CYBB gene [36]. However, in the case of our CGD patients, rhIFN-𝛾 was not used as an immuno adjuvant. Treatment options for CGD typically include antibacterial prophylaxis (trimethoprim-sulfamethoxazole), antifungal prophylaxis (itraconazole), recombinant human interferon gamma, (rhIFN) and Hematopoietic stem cell transplant (HSCT). All these have been shown to aid in the recovery from infections in CGD [37,38,39,40,41,42]. In our cohort, CGD patients utilized antibacterial and antifungal prophylaxis, but they did not receive rhIFN as immuno adjuvant nor undergo HSCT. However, in vitro treatment of MDMs from gp91phox deficient patients with rhIFN-𝛾 improved their ability to control growth of M. tuberculosis, which aligns with previous reports [43, 44]. Importantly, we suggest that further validation of our result regarding the in vitro treatment of rhIFN-𝛾 should involve more MDMs from gp91phox functional deficient patients with similar underlying mutations.
Conclusion
We conclude that mutations in the CYBB gene cause X-linked CGD, and thus expanding the clinical and genetic spectrum of the disease. Additionally, early diagnosis of CGD is crucial for timely intervention, enabling physicians to treat infections before they escalate to a severe life-threatening stage.
Data Availability
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
Abbreviations
- CGD:
-
chronic granulomatous disease
- CYBB :
-
Cytochrome B-245 beta chain
- DHR:
-
dihydrorhodamine
- HEC:
-
higher education commission
- HIV:
-
human immunodeficiency virus
- H2O2 :
-
hydrogen peroxide
- NADPH:
-
oxidase nicotinamide adenine dinucleotide phosphate oxidase
- PBS:
-
phosphate buffered saline
- PCR:
-
Polymerase Chain Reaction
- PID:
-
primary immunodeficiencies
- PMA:
-
phorbol myristate acetic acid
- ROS:
-
reactive oxygen species
- SIFT:
-
sorting intolerant from tolerant
- XCGD:
-
X-linked chronic granulomatous disease
References
de Oliveira-Junior EB, Bustamante J, Newburger PE, Condino-Neto A. The human NADPH oxidase: primary and secondary defects impairing the respiratory burst function and the microbicidal ability of phagocytes. Scand J Immunol. 2011;73(5):420–7.
Lambeth JD. NOX enzymes and the biology of reactive oxygen. Nat Rev Immunol. 2004;4(3):181–9.
Deffert C, Cachat J, Krause KH. Phagocyte NADPH oxidase, chronic granulomatous disease and mycobacterial infections. Cell Microbiol. 2014;16(8):1168–78.
El-Benna J, Dang PM, Gougerot-Pocidalo MA, Marie JC, Braut-Boucher F. p47phox, the phagocyte NADPH oxidase/NOX2 organizer: structure, phosphorylation and implication in diseases. Exp Mol Med. 2009;41(4):217–25.
Tkalcevic J, Novelli M, Phylactides M, Iredale JP, Segal AW, Roes J. Impaired immunity and enhanced resistance to endotoxin in the absence of neutrophil elastase and cathepsin G. Immunity. 2000;12(2):201–10.
Segal BH, Leto TL, Gallin JI, Malech HL, Holland SM. Genetic, biochemical, and clinical features of chronic granulomatous disease. Medicine. 2000;79(3):170–200.
Holland SM. Chronic granulomatous disease. Hematol Oncol Clin N Am. 2013;27(1):89–99. viii.
Khan TA, Kalsoom K, Iqbal A, Asif H, Rahman H, Farooq SO, et al. A novel missense mutation in the NADPH binding domain of CYBB abolishes the NADPH oxidase activity in a male patient with increased susceptibility to infections. Microb Pathog. 2016;100:163–9.
Stasia MJ, Bordigoni P, Floret D, Brion JP, Bost-Bru C, Michel G, et al. Characterization of six novel mutations in the CYBB gene leading to different sub-types of X-linked chronic granulomatous disease. Hum Genet. 2005;116(1–2):72–82.
Leiva LE, Zelazco M, Oleastro M, Carneiro-Sampaio M, Condino-Neto A, Costa-Carvalho BT, et al. Primary immunodeficiency diseases in Latin America: the second report of the LAGID registry. J Clin Immunol. 2007;27(1):101–8.
Wood P, Stanworth S, Burton J, Jones A, Peckham DG, Green T, et al. Recognition, clinical diagnosis and management of patients with primary antibody deficiencies: a systematic review. Clin Exp Immunol. 2007;149(3):410–23.
Hasui M, Hirabayashi Y, Kobayashi Y. Simultaneous measurement by flow cytometry of phagocytosis and hydrogen peroxide production of neutrophils in whole blood. J Immunol Methods. 1989;117(1):53–8.
Köker MY, Sanal O, De Boer M, Tezcan I, Metin A, Ersoy F, et al. Mutations of chronic granulomatous disease in turkish families. Eur J Clin Invest. 2007;37(7):589–95.
Wada T, Muraoka M, Toma T, Imai T, Shigemura T, Agematsu K, et al. Rapid detection of intracellular p47phox and p67phox by flow cytometry; useful screening tests for chronic granulomatous disease. J Clin Immunol. 2013;33(4):857–64.
Ali SK, Doss CG, Kumar DT, Zhu H. CoagVDb: a comprehensive database for coagulation factors and their associated SAPs. Biol Res. 2015;48(1):35.
Girotto G, Scheffer DI, Morgan A, Vozzi D, Rubinato E, Di Stazio M, et al. PSIP1/LEDGF: a new gene likely involved in sensorineural progressive hearing loss. Sci Rep. 2015;5:18568.
Schiemann AH, Stowell KM. Comparison of pathogenicity prediction tools on missense variants in RYR1 and CACNA1S associated with malignant hyperthermia. Br J Anaesth. 2016;117(1):124–8.
Martins-Pinheiro M, Lima WC, Asif H, Oller CA, Menck CF. Evolutionary and Functional Relationships of the dha regulon by genomic context analysis. PLoS ONE. 2016;11(3):e0150772.
Magnani F, Nenci S, Millana Fananas E, Ceccon M, Romero E. Crystal structures and atomic model of NADPH oxidase. 2017;114(26):6764–9.
Sali A, Blundell TL. Comparative protein modelling by satisfaction of spatial restraints. J Mol Biol. 1993;234(3):779–815.
Esquivel-Solís H, Quiñones-Falconi F, Zarain-Herzberg A, Amieva-Fernández RI, López-Vidal Y. Impaired activation of Stat1 and c-Jun as a possible defect in macrophages of patients with active tuberculosis. Clin Exp Immunol. 2009;158(1):45–54.
Roos D, Kuhns DB, Maddalena A, Roesler J, Lopez JA, Ariga T, et al. Hematologically important mutations: X-linked chronic granulomatous disease (third update). Blood Cells Mol Dis. 2010;45(3):246–65.
Ishibashi F, Nunoi H, Endo F, Matsuda I, Kanegasaki S. Statistical and mutational analysis of chronic granulomatous disease in Japan with special reference to gp91-phox and p22-phox deficiency. Hum Genet. 2000;106(5):473–81.
n den Berg JM, van Koppen E, Ahlin A, Belohradsky BH, Bernatowska E, Corbeel L, et al. Chronic granulomatous disease: the european experience. PLoS ONE. 2009;4(4):e5234.
Winkelstein JA, Marino MC, Johnston RB Jr, Boyle J, Curnutte J, Gallin JI, et al. Chronic granulomatous disease. Report on a national registry of 368 patients. Medicine. 2000;79(3):155–69.
Filipe-Santos O, Bustamante J, Haverkamp MH, Vinolo E, Ku CL, Puel A, et al. X-linked susceptibility to mycobacteria is caused by mutations in NEMO impairing CD40-dependent IL-12 production. J Exp Med. 2006;203(7):1745–59.
Al-Hammadi S, Yahya AM, Al-Amri A, Shibli A, Balhaj GB, Tawil MI, et al. Case Report: BCG-Triggered Hemophagocytic Lymphohistiocytosis in an infant with X-Linked recessive mendelian susceptibility to Mycobacterial Disease due to a variant of chronic Granulomatous Disease. Front Pead. 2021;9:687538.
Köker MY, Camcıoğlu Y, van Leeuwen K, Kılıç S, Barlan I, Yılmaz M, et al. Clinical, functional, and genetic characterization of chronic granulomatous disease in 89 turkish patients. J Allergy Clin Immunol. 2013;132(5):1156–63e5.
Fattahi F, Badalzadeh M, Sedighipour L, Movahedi M, Fazlollahi MR, Mansouri SD, et al. Inheritance pattern and clinical aspects of 93 iranian patients with chronic granulomatous disease. J Clin Immunol. 2011;31(5):792–801.
Agudelo-Flórez P, Prando-Andrade CC, López JA, Costa-Carvalho BT, Quezada A, Espinosa FJ, et al. Chronic granulomatous disease in latin american patients: clinical spectrum and molecular genetics. Pediatr Blood Cancer. 2006;46(2):243–52.
Morel F. [Molecular aspects of chronic granulomatous disease. “the NADPH oxidase complex”]. Bull de l’Academie nationale de Med. 2007;191(2):377–. – 90; discussion 90 – 2.
Hussain W, Batool A, Ahmed TA, Bashir MM. Severe combined immunodeficiency due to adenosine deaminase deficiency. JPMA The Journal of the Pakistan Medical Association. 2012;62(3):297–9.
Khan MO, Memon AI, Zubairi AB, Imam AA. Undetected common variable immune deficiency in a young adult of pakistani descent. J Ayub Med Coll Abbottabad. 2011;23(1):165–6.
Saleem AF, Ali Khawaja RD, Shaikh AS, Ali SA, Mehdi Zaidi AK. Severe combined immune deficiency syndrome. J Coll Physicians Surgeons–Pakistan : JCPSP. 2013;23(8):570–3.
von Goessel H, Hossle JP, Seger R, Gungor T. Characterization of 17 new cases of X-linked chronic granulomatous disease with seven novel mutations in the CYBB gene. Exp Hematol. 2006;34(4):528–35.
Bionda C, Li XJ, van Bruggen R, Eppink M, Roos D, Morel F, et al. Functional analysis of two-amino acid substitutions in gp91 phox in a patient with X-linked flavocytochrome b558-positive chronic granulomatous disease by means of transgenic PLB-985 cells. Hum Genet. 2004;115(5):418–27.
Marciano BE, Wesley R, De Carlo ES, Anderson VL, Barnhart LA, Darnell D, et al. Long-term interferon-gamma therapy for patients with chronic granulomatous disease. Clin Infect Dis. 2004;39(5):692–9.
Gallin JI, Alling DW, Malech HL, Wesley R, Koziol D, Marciano B, et al. Itraconazole to prevent fungal infections in chronic granulomatous disease. N Engl J Med. 2003;348(24):2416–22.
Weening RS, Kabel P, Pijman P, Roos D. Continuous therapy with sulfamethoxazole-trimethoprim in patients with chronic granulomatous disease. J Pediatr. 1983;103(1):127–30.
Margolis DM, Melnick DA, Alling DW, Gallin JI. Trimethoprim-sulfamethoxazole prophylaxis in the management of chronic granulomatous disease. J Infect Dis. 1990;162(3):723–6.
Oikonomopoulou Z, Shulman S, Mets M, Katz B. Correction to: chronic granulomatous disease: an updated experience, with emphasis on newly recognized features. 2022;42(7):1420.
Gunderman L, Brown J, Chaudhury S, O’Gorman M, Fuleihan R, Khanolkar A. Co-occurring X-Linked agammaglobulinemia and X-Linked chronic Granulomatous Disease: two isolated pathogenic variants in one patient. 2023;11(3).
Gao XF, Yang ZW, Li J. Adjunctive therapy with interferon-gamma for the treatment of pulmonary tuberculosis: a systematic review. Int J Infect Dis. 2011;15(9):e594–600.
Khan TA, Schimke LF, Amaral EP, Ishfaq M, Barbosa Bonfim CC, Rahman H, et al. Interferon-gamma reduces the proliferation of M. tuberculosis within macrophages from a patient with a novel hypomorphic NEMO mutation. Pediatr Blood Cancer. 2016;63(10):1863–6.
Acknowledgements
All authors thank to all doctors of DHQ hospitals Khyber Pakhtunkhwa and the participants/patients for providing clinical information/histories.
Funding
The whole PhD study practically as well as the open access funding provided by The Jeffrey Modell Foundation, USA and Higher Education Commission (HEC), Pakistan through project No:10352/KPK/NRPU/R&D/HEC/2017. We acknowledge and extend our gratitude to the aforementioned funding organizations.
Author information
Authors and Affiliations
Contributions
IG: performed the design of the study and experimental work. TAK: performed the statistical analysis and shared in writing manuscript. NA: collected data and did clinical assessment for the studied group. NG: collected data and shared in writing and finalizing manuscript. RA: analyzed the gene sequences of the patients, SNK, checked out the whole manuscript and finalize the writing of manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate.
The experimental protocols were approved by the ethical committee of Kohat University of Science and Technology (KUST) under reference number (KUST/Ethical committee/18 − 2). All methods were performed in accordance with the ethical standards as laid down in the Declaration of Helsinki and its later amendments. Written informed consent was obtained from all participants.
Consent for publication.
Written informed consent for publication was obtained.
Competing interests.
All the authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gul, I., Khan, T.A., Akbar, N.u. et al. Novel mutations in CYBB Gene Cause X-linked chronic Granulomatous Disease in Pakistani patients. Ital J Pediatr 49, 95 (2023). https://doi.org/10.1186/s13052-023-01496-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13052-023-01496-7