
Abstract
Currently, it is estimated that 15% of human neoplasms globally are caused by infectious agents, with new evidence emerging continuously. Multiple agents have been implicated in various forms of neoplasia, with viruses as the most frequent. In recent years, investigation on viral mechanisms underlying tumoral transformation in cancer development and progression are in the spotlight, both in human and veterinary oncology. Oncogenic viruses in veterinary medicine are of primary importance not only as original pathogens of pets, but also in the view of pets as models of human malignancies. Hence, this work will provide an overview of the main oncogenic viruses of companion animals, with brief notes of comparative medicine.
Similar content being viewed by others


Introduction
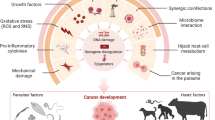
For many years, studies and observations on the animal world have contributed to our medical and scientific knowledge. For instance, porcine dissections were used to write the earliest anatomy textbooks around 900 A.D. [1], while rodents have been employed as models for cancer research for years [2]. Over time, animal models have yielded many general concepts in molecular oncology and hypotheses on the role of oncoviruses in tumor development [3]. Rous sarcoma virus, discovered by Rous in the early twentieth century, was the first oncogenic virus [4, 5] and a landmark in the oncovirus field. It led to the research on proto-oncogenes and the cancer-related action of tyrosine kinase. Nearly half a century later, in 1964, Epstein discovered the first human oncogenic virus, thanks to Rous's intuition [6]. Subsequently, it was discovered that oncoviruses are prevalent among both animals and humans and cause about 12% of human cancers [7,8,9,10]. Despite many similarities between animals and humans, there remain intrinsic differences that must be considered when they are used in cancer research [3]. However, the use of animals as models for human diseases is a crucial cornerstone of cancer research, both in the past and present, especially for prevention and therapy [3]. Over the years the use of animal models has been essential for medical understanding and advancement. To make only some examples, animals were extremely useful in the understanding process of human diseases, in the development of vaccines [11], antibiotics [12] and new surgical techniques [13, 14], in the identification and validation of new markers as well as in preclinical studies for the evaluation of new therapies [15]. The identification of a viral etiology for many types of cancers has several implications. It provides additional information on cancer development and progression and leads to the identification of new cellular targets for therapy. This knowledge is useful not only for virus-related cancers but also for those with no viral etiology. For example, the role of tumor suppressors p53 and pRb led to the development of several therapeutic approaches. Additionally, viral gene products themselves may serve as potential targets for therapy or may allow the development of screening and prevention strategies, as already happened with the Pap smear and vaccination against Human Papillomavirus (HPV)-related cervical cancer [16, 17]. Establishing a causal association between a virus and a specific cancer requires fulfilling most, if not all, of Hill's criteria [18], which demands a substantial number of epidemiological studies. For this reason, a limited number of viruses are currently considered as pathogens with oncogenic potential in humans. Among these, Epstein Barr Virus, Hepatitis B Virus (HBV); Hepatitis C Virus (HCV), Human Papilloma Virus (HPV), Human T-Lymphotropic Virus 1 (HTLV-1), Human Herpesvirus 8 (HHV-8), Merkel Cell Polyomavirus (MCP) and Mouse Mammary Tumor Virus (MMTV) are recognized [19, 20]. Furthermore, although there is a substantial body of literature on the correlation between viruses and cancer in domestic animals, it is often fragmented and lacks a comprehensive overview. Therefore, we consider it extremely useful to collect all data and review the various research studies carried out so far over the years. Modern oncology has focused on using companion animals as the best animal model for human diseases. Indeed, compared to rodents, they are phylogenetically closer to humans and show (i) similar genomic organization, (ii) similar cancer incidence, (iii) similar molecular pathological and clinical features, (iv) similar toxicity and therapeutic responses compared to the human counterpart. Of note, unlike laboratory rodents, they develop spontaneous cancer, and different from humans, they have a shorter life, making cancer progression often faster. Finally, they also share the environment and the socioeconomic factors with their owners [15]. All these features make the companion animals eligible to be the best candidates for animal models in cancer research, and for this reason we decided to focus this review on them. We will provide an overview of oncoviruses of veterinary importance in domestic animals, investigating their role as pathogens for both dogs and cats. In the first section of this review, we will describe the oncoviruses for which the oncogenic potential has already been proven in literature: Feline Leukemia Virus (FeLV), Feline Immunodeficiency Virus (FIV) and Papillomavirus (PV). In the second section, we will collect and discuss data about emerging potentially oncogenic viruses, for which a cause-effect relationship to cancer in domestic animals has not been established yet: Herpesvirus, Hepadnavirus, and Mouse Mammary Tumor Virus (MMTV). For all these viruses, the interest has arisen from evidence on the oncogenic potential of similar viruses in humans. For each virus discussed, we will provide data on their structure, life cycle, clinical presentation, transmission and oncogenic mechanisms in dogs and cats, when available, or in their original host when not, with brief comparative notes. An overview of the current knowledge about the association between viral agents and cancer in veterinary medicine is summarized in Fig. 1, together with a focus on the main oncogenic mechanisms proposed.

Overview of the oncoviruses in domestic animals and of their main oncogenic mechanisms. Created by Biorender.com
Feline leukemia virus
FeLV is an exogenous retrovirus of the genus Gammaretrovirus. Infection occurs worldwide, with a prevalence different in various geographical areas, ranging from less than 1% to 20% based on cat density [21]. FeLV is an enveloped single stranded RNA virus made up of three genes, the group specific antigen (gag), the polymerase (pol) and the envelope (env) genes. These three genes are flanked by long terminal repeats (LTRs) containing promoter and enhancer elements. On the basis of the env sequences, three main subtypes are described which differ in receptors and cell tropism: FeLV-A, FeLV-B, and FeLV-C. Only the predominant subtype, FeLV-A, can be transmitted from cat to cat, while the other two result from genomic alterations achieved through different mechanisms, but they are typically not further transmissible. FeLV-B occurs in 50% of infected cats and it is the result of a genome recombination between the FeLV-A subtype and the endogenous FeLV-related retroviruses. Endogenous retroviruses in cats are common genetic elements resulting from retroviral infection of ancestors. These elements cannot be transmitted exogenously but are inherited vertically by the germ line [22]. The infection with FeLV-B subtype can accelerate the development of lymphoma or increase the virus neuropathogenicity [22]. FeLV-C subtype arises rarely (1%) after a point mutation in the env gene sequence and causes fatal anemia in infected subjects [22, 23]. A more recent discovery involved another FeLV variant that arises through multiple mutations of the env gene in FeLV-A-infected subjects. This variant, with a selective tropism for T-lymphocytes, is known as FeLV-T and is associated with severe immunodeficiency [24,25,26].
The receptor for the FeLV-A subtype was recently recognized as a thiamine transport protein [27], while the FeLV-B subtype enters cells using Na-dependent inorganic phosphatase transporters [26, 28], the FeLV-C subtype uses a hemopoietic cell-associated transporter molecule [29, 30], and the FeLV-T subtype uses a coreceptor expressed on T-lymphocytes called feLIX [31].
The mechanism of infection by FeLV is similar to those of other retroviruses. Once the viral glycoprotein spikes on the envelope surface recognize their specific receptors on cells, the viral core can be internalized. The two copies of single-stranded RNA released in the host cell cytoplasm are reverse transcribed into DNA by reverse transcriptase. The viral DNA enters the nucleus and integrates into the host cell DNA, acting as a template for the production of new viral particles [32].
FeLV is mainly transmitted horizontally through the oronasal route of infection [33] by contact with virus-spreading secretions (e.g. saliva) from viremic subjects [34, 35]. This route of transmission makes overcrowded environments, such as cat colonies and catteries, the most infectious places for cats, due to the sharing of food, water, bowls and mutual grooming [31]. After contacts with viremic cats, FeLV initially replicates in lymphocytes and macrophages in local lymphoid tissue of the oropharynx [21, 31]. Viral particles spread through draining lymph node and blood to tissues rich in rapidly dividing cells [36]. The outcome of infection may be variable, depending on a number of factors, like immune status and age of the host, concentration and pathogenicity of the virus, route of exposure, and the presence of concomitant diseases [21, 31, 37]. If the immune system is able to contain viral spread through an appropriate immune response, cats may recover from infection, being known as “regressor subjects” [21, 32, 38]. This event, in natural circumstances, takes place in 1% up to 10% of exposed cats [23]. In the remaining subjects, the immune response does not succeed in controlling virus replication and infected cats develop persistent viremia. Viral particles spread to the target organs (spleen, thymus, lymph nodes, and salivary glands) and cats manifest nonspecific clinical signs, like pyrexia and lethargy, becoming infectious to others [21, 37]. Generally, this phase is transient, lasting about three weeks, during which cats are known as “transiently viremic” [21, 37]. Most of these cats succeed in overcoming infection and develop an immune response protecting them from further new exposure. In those who do not succeed in neutralizing infection, the involvement of bone marrow has been reported. The virus integrates its genome in haemopoietic precursor cells, and the progeny cells released will all be infected, with the development of high-level viremia [37]. A number of subjects recover from viremia, but remain “latently infected” because of the persisting integration of the virus in bone marrow stem cell genome, while those who do not recover remain “persistently infected” [32].
Subjects with persistent infection (but also latently infected ones after reactivation [39, 40] may develop FeLV-related diseases and have a poor prognosis [23, 31]. Only 20% to 50% of cats with progressive infection were reported to survive three years post diagnosis [23, 41, 42]
FeLV-associated diseases are not only lymphoma and leukemia, as suggested by the name of the virus, but they also include non-neoplastic diseases related to its pancytotropism and to the involvement of bone marrow, lymphoid system and rapidly dividing cells. Among these diseases, myeloproliferative and immune-mediated diseases, cytopenia (particularly anemia), enteritis, and reproductive disorders can be found, along with secondary infections due to immunosuppression [23, 31, 32, 43, 44]
Lymphoma is the most common lymphoid malignancy associated to progressive infection by FeLV [45] and it is reported to occur in 10–20% of progressive infections [46,47,48]. FeLV-associated lymphomas are mainly high grade, T-type lymphomas, while their anatomic localization is related to the age of the host. The thymus is the most frequently involved especially in young cats [23], while FeLV-induced alimentary lymphomas are also frequent, but generally related to older subjects [31]. Spinal and multicentric localization are the most frequently reported sites [23], with the latter being typical of young subjects [31].
FeLV is a simple retrovirus, therefore its genome does not carry oncogenes directly causing malignancy, but it is suspected to cause cancer through indirect mechanisms. One of the most frequent ways is by insertional mutagenesis, through which the virus influences the expression of genes by integrating its genome in specific sites of the host DNA [23]. Particularly, integration may occur near cellular proto-oncogenes, which can be upregulated by enhancer sequences and transcriptional promoters located in the U3 region of LTRs [49,50,51,52,53,54,55,56]. Molecular analyses demonstrated that the most common integration sites (CISs) in FeLV-associated lymphomas occur in the vicinity of six genes, namely c-myc, flvi-1, flvi-2, fit-1, pim-1, and flit-1 [53, 54, 57,58,59,60,61]. As evidence, mutational events in LTRs enhancing their functions [23, 62,63,64,65], as well as c-myc dysregulation [54], are frequently observed in FeLV-associated neoplasms. However, viral integration can also occur within sequences encoding tumor suppressing genes, interrupting their expression. In this case, both alleles should be inactivated to cause an effective gene loss of function, so this is an uncommon way [23].
Another mechanism responsible of the development of FeLV-related lymphomatous malignancies is transduction, thanks to which the virus acquires cellular oncogenes during its replication. In this eventuality, the oncogene will be transduced along with the viral genome [23, 61]. This mechanism is exploited by the FeLV-A subtype when it acquires cellular oncogenes like fes, fms, fgr, abi, and kit through transduction, giving rise to the so-called feline sarcoma viruses (FeSV), responsible of multicentric sarcomas [32, 66, 67]. FeSV belongs to the group of sarcoma viruses, recombinant viruses coming from the assembling of some leukemia virus genes and some host cell genes that Hardy, in 1981, defined as sarc [66]. Thus, FeSV genome is characterized by two distinct sequences subsets, one including sequences shared with FeLV helper virus and designated as com, the other including sequences coming from the transduction of host cell gene, known as src sequences, conferring transforming properties to the virus [68]. It has been highlighted that src sequences from sarcoma virus of the various species are close each other, thus suggesting that they came from the same ancient cellular genes that has been conserved for years in many species [66]. Furthermore, FeSV lacks pol gene and most of the env gene, thus it cannot synthetize reverse transcriptase enzyme and the proteins required for envelope production, like gp70 and p15E. For this reason, in nature FeSV exists as pseudotype virus with a FeLV helper [69,70,71]. Viral particles are composed by a FeSV RNA genome enclosed in a FeLV envelope. Thanks to the presence of envelope, viral particles can enter host cells through their receptors, within the cells the FeLV help reverse transcriptase allows the formation of both FeLV and FeSV proviral DNA, which are then integrated into the host cell DNA. In this way, viral genes may be expressed and the presence of src genes may allow malignant transformation of host cells, thus giving rise to sarcoma. Two distinct types of viral particles are then produced from these infected cells: FeLV viral particles, characterized by FeLV RNA and enclosed in FeLV envelope, and FeSV viral particles [66].
As last oncogenetic mechanism proposed, it has been hypothesized that FeLV may influence cellular genes through transactivation [72], by which transcripts from the U3 region may activate a signaling pathway commonly involved in oncogenesis. Moreover, the role of endogenous FeLV has recently been questioned, since two studies suggested that their presence was associated with better prognosis upon infection with exogenous FeLV [32]. Finally, we should also take into consideration that FeLV may contribute to the development of neoplasms simply by bringing about an immunosuppression state associated with viral infection which leads to the reduction of the immune surveillance and contributes to tumor cell survival [23].
Feline immunodeficiency virus
FIV is an enveloped RNA virus belonging to the Lentivirus genus of the Retroviridae family [73]. It was described for the first time in 1986 in a cattery in California [74] and is known to cause an immunodeficiency syndrome similar to human AIDS in cats worldwide [75], with a prevalence ranging from 1% in healthy domestic cats up to 47% in feral subjects, with a higher prevalence in sick cats [32, 76,77,78,79,80,81,82].
Like other retroviruses, the FIV provirus is bordered by LTRs and comprises gag, pol and env genes, as well as other regulatory and accessory genes [83]. Based on the env gene sequence, six different subtypes of FIV are recognized, from A to F, with A, B and probably C as the most widely distributed subtypes. Recombinant subtypes have also been documented and the existence of further subtypes has been suspected [73].
To enter the host cells, FIV uses CD134 as its primary receptor and CXCR4, a chemokine receptor, as a secondary one. CD134 is expressed in CD4 + T-lymphocytes, B-lymphocytes and activated macrophages. For this reason, they act as the first target cells for the virus [73]. The mechanism of infection is similar to FeLV and other retroviruses (see the previous section). Proviral DNA may be transcriptionally active or silent, based on cellular environment [73]. FIV is transmitted through biting by infected cats [84]. This transmission route makes free ranging intact male cats, prone to fighting behaviors, more exposed to infection [31, 74]. Differently from natural conditions, vertical transmission has also been demonstrated in the experimental field [31, 32, 85, 86]. Unlike Human Immunodeficiency Virus (HIV), venereal transmission has not been documented for FIV, but viral particles have been found in semen and experimental infection through the vagina has been demonstrated. Moreover, transmission through viral inoculation in bloodstream is thought to be an additional route [32, 73].
As HIV, FIV can progress through several stages of infection. Viral particles are inoculated through saliva, infect lymphoid and myelomonocytic cells, particularly CD4 + T lymphocytes, macrophages, and dendritic cells, integrate their genome, and then start to replicate and release new virions. In the first phase of viremia peak, early clinical signs are fever, anorexia, depression, leucopenia, gingivitis, and general lymphadenopathy [31, 75], which regress with the development of the host immune response. Within one week from infection, FIV-specific CD8 + T lymphocytes can be detected [87], while anti-FIV antibodies and virus-neutralizing antibodies are detected by three weeks after [88]. The development of a specific immune response causes the viral load to decrease until a steady state is reached, which marks the beginning of the latent asymptomatic phase. This stage is characterized by a slow and steady reduction of CD4 + T lymphocytes that may last for years, in some cases for the entire life of cats, while the infected animals are infected subclinical [31, 75]. However, an evolution to an end stage with clinical manifestation similar to AIDS for HIV-infected humans is also possible, with marked loss of CD4 + T lymphocytes, frequent bacterial respiratory, ocular and oral infections, enteritis, neurologic disorders, opportunistic infections, and development of cancers [31, 75]. Despite this possible evolution, infection is not necessarily life-threatening, and many studies demonstrated that life expectancy of FIV-infected cats is quite comparable to that of non-infected ones [42, 89, 90]. Even if the outcome of FIV infection is not predictable [90,91,92], it has been suggested that several factors influence its evolution, like genetic features and concurrent diseases [31]. The development of cancer is reported with an incidence from 1 to 21% in positive subjects. In addition to sporadic reports of myeloproliferative leukemia, mastocytoma, fibrosarcoma, and squamous cell carcinoma [93,94,95], lymphoma is the most common neoplasia associated with FIV [76, 84, 94,95,96,97,98,99,100,101]. Particularly, it has been reported that FIV-infected cats are 5 to 6 times (up to 80 times in case of FeLV/FIV coinfection) more susceptible to develop lymphoma compared to uninfected ones [48]. FIV-associated lymphomas are reported to be mainly B-type [100,101,102,103,104,105] with a prevalence from 40 to 87% [102,103,104,105], while T-cell lymphomas range between 0 and 28% [101, 103,104,105] and non-B, non-T lymphomas are uncommon [101, 103,104,105]. A possible explanation of the prevalence of B-type lymphomas is the rapid proliferation of B lymphocytes occurring in the early phase of infection [75], that statistically increases the chances of these cells to malignant transformation [106,107,108]. There is no anatomical predisposition, since FIV-associated lymphomas have been described in a multitude of organs [75], and neoplasms arise in cats ranging from 5 to 13 years [48, 93, 94, 101,102,103, 109]. In respect to oncogenic mechanisms, FIV is not likely to be directly involved in lymphomagenesis because its genome does not carry oncogenes, so it cannot induce neoplasms through acute transformation. Insertional mutagenesis mechanisms are also unlikely since, in this perspective, clonal integration of provirus should be recognized in lymphomas. Only two studies support this finding [105, 110, 111], while most of the other investigations trying to confirm insertional mutagenesis power of FIV have failed in their attempt [99, 101, 103]. Nowadays, the most accepted theory is that FIV induces lymphoma via indirect mechanisms, for instance through the reduction of immune surveillance, and consequent impairment of neoplastic cell removal secondary to virally-induced immune dysfunction [75].
Papillomavirus
PV has become relevant in human medicine since it was proved that HPV type 16 and 18 are causative agents of cancer. Particularly, it has been shown that some genotypes, belonging to Alphapapillomavirus, are able to induce cervical cancer, as well as anogenital tumors and a percentage of head and neck squamous cell carcinomas mostly in the oropharyngeal area [112,113,114,115]. Following these findings, nowadays, human PVs (HPVs) are subdivided in “low-risk types”, causing asymptomatic or self-resolving conditions, and “high-risk types”, causing cancer [116]. There is more and more compelling evidence that canine papillomavirus (CPV) and feline papillomavirus (FPV) may also cause cancer in dogs and cats, respectively, but additional studies are required to investigate their role in tumor development [117]. Although viral etiology of warts has been recognized since 1907 [118], the first association between PV and cancer dates back to 1935, after one study on rabbits [119]. It was only in 1981 that PVs acquired great importance, when zur Hausen et al. proved that PV caused cervical cancer in humans [120]. PV was suspected as a viral etiological agent for cancer for the first time in 1969 for dogs [117] and in 1990 for cats [121], but it was only in 1994 that the first canine PV was sequenced [122], while the first feline PV was sequenced later, in 2002 [123, 124].
PV is a small, non-enveloped virus containing a double-stranded circular DNA genome, composed of three main regions: the long control region (LCR), the early region (ER) and the late region (LR). The LCR is involved in the modulation of viral replication and transcription. The ER and LR are composed of six and two open reading frames (ORF) respectively, in detail E1, E2, E4, E5, E6, E7 for ER and L1 and L2 for LR, encoding for their respective and homonymous proteins [125, 126]. Particularly, E5, E6 and E7 are oncoproteins responsible for the induction of cell proliferation and cancer development through different mechanisms. Differently from HPV and CPV, FPV doesn’t express E4 and E5 proteins [127]. Taxonomic classification of PVs is based on the highly conserved L1 ORF. If the similarity between L1 ORF is more than 60% then two PVs are classified within the same genus; if similarity is less than 90%, they are classified in different types [117, 128]. Types belonging to the same genus generally infect the same host species with similar presentation [128]. In dogs, twenty-three types of CPV, belonging to the Lambdapapillomavirus, Taupapillomavirus and Chipapillomavirus genera, are reported, while, in cats, six types of FPV have been detected within the Lambdapapillomavirus, Taupapillomavirus and Dyothetapapillomavirus genera [116].
PVs can be transmitted directly, through contact with infected subjects, or indirectly, via fomites [116, 117]. The infection begins when abrasions on mucocutaneous epithelium allow the virus to access the basal layer of cells. Here, binding between L1 protein and heparan sulfate proteoglycan receptors allows conformational changes in viral capsid, thus permitting contact between the L2 viral protein and a second receptor, located in the annexin A2 heterotetramer [129, 130]. PV internalization occurs through endocytosis: the L1-L2-viral DNA complex is transported at first to the Golgi network and then to the host cell nucleus [131]. Here, viral replication and transcription occur, thanks to cellular transcription factors (TFs), which are specific of differentiated epidermal cells [132]. Thus, as basal cells differentiate, viral genome is amplified, and early proteins are expressed. Particularly, E6 and E7 oncoproteins increase their expression and interact with p53 and pRB, with mechanisms explained in detail below, inhibiting apoptosis, causing cell cycle arrest and enhancing the progression from G1 to S phase in cell cycle [133, 134]. In the meantime, E1 and E2 expression amplifies viral genome replication and multiple viral DNA copies are produced [135]. In this way, PV infection persists since progeny cells inherit PV DNA and move to the suprabasal layer of the epithelium [116, 136, 137]. The production of infectious viral particles is possible only once L1 and L2 are expressed and produce the viral capsid around viral DNA. Then virions can be released into the environment thanks to the action of protein E4, which is responsible of viral particle-laden keratinocyte rupture [138,139,140].
Clinical presentation of PV infection may be variable. Generally, most HPV types do not cause clinical manifestations [141,142,143]; infection is subclinical for most dogs and cats as well [144,145,146]. Only a small percentage of infection cases exhibits an increase in keratinocytes replication hesitating in epithelial hyperplasia and papillomas [117]. This outcome is generally linked to the presence of certain PV types or to the lack of immune response. Indeed, even if the virus is confined in the outer epithelial layers and the immune response is weak, PV elicits both a humoral and cell-mediated response. The former is responsible of the production of IgG antibodies blocking further infection by the same PV-type, the latter is involved in resolution of an established infection [117]. Conversely, immunosuppression may favor the progression of PV infection into clinical disease, with different presentations: hyperplastic lesions, preneoplastic diseases and cancers.
Among hyperplastic lesions, PV is frequently associated with cutaneous and oromucosal warts. This clinical presentation is typical of young dogs [147], where CPV2 and CPV1, alone or in coinfection, and rarely CPV6, cause papillomas [148,149,150,151,152]. They usually arise as cauliflower-like lesions on areas subjected to trauma, like feet or around the face, lips, and ears [116, 117, 153,154,155] and are generally self-limiting. There are very few reports of CPV-induced warts transformed into SCC [156]. However, there are some reports of lesions that continued to increase in size, spreading to the haired skin [157] or progressing to SCC [158].
Surgical excision or cryotherapy is recommended if these lesions become too large and interfere with eating or breathing [159]. In cats, however, this clinical presentation is not so frequent: warts are suspected to be caused by FPV1. There are very few reports on this manifestation appearing on the nasal planum, on the eyelid or arising in clusters on the ventral side of tongue [116, 160, 161].
PV-related preneoplastic lesions in domestic animals are represented by pigmented viral plaques both in dogs and cats and by Bowenoid in situ carcinomas (BISCs) in cats alone. Of note, pigmented viral plaques and BISCs are generally described as different pathologies even if they probably represent different stages of the same entity. Particularly, it is the presence of more marked hyperplasia that makes the lesion bulge into the dermis, keratinocytes dysplasia or basal cell crowding help to identify BISCs [162].
PV-related preneoplastic lesions rarely develop both in middle-aged and older dogs and cats [163, 164], and are caused by Chipapillomavirus types, mainly CPV4 in dogs [165,166,167,168], and FPV2 or, less frequently, other Taupapillomavirus in cats [162, 169,170,171,172,173]. In both species, infection is generally subclinical, but immune disfunctions may cause inability to limit viral replication, leading to increased PV replication and thickening of the epidermis, with the development of lesions. In both species, breed predisposition has been proposed as linked to inherited deficiency in keratinocyte immunity. Particularly, predisposition has been highlighted in Vizla and Pug dogs [164, 165, 174, 175] and in Sphinx and Devon Rex cats [176, 177]. Plaques usually occur on limbs and face in dogs, while preneoplastic lesions generally arise on the head or neck in cats. Canine plaques have a benign course, where coalescence or extension to other areas are rare [163, 178] and spontaneous regression often occurs. Progression to SCC has been reported only once, during infection with specific HPV types, like CPV16 [149, 151, 179,180,181]. Conversely, in cats, these plaques rarely regress; quite oppositely, they usually progress to ulceration and more severe morbidity that has to be treated through cryotherapy, surgery or using specific cremes [182]. Progression to SCC has been reported, especially in Sphynx and Devon Rex cats [176, 177].
Due to the frequent subclinical course of CPV, even if some studies report the presence of CPV DNA in canine SCCs [183,184,185], its detection is challenging to understand. Conversely, several studies have linked FPV to SCC, basal cell carcinoma and Merkel cell carcinoma in cats [117, 186]. Particularly, literature suggested that an FPV etiology may be suspected in 75% of feline SCCs arisen in UV-protected areas, while in UV-exposed areas SCCs are mainly caused by UV rays and only 30% of them are linked to PV [187]. The most frequent SCC-associated type in cats is FPV2 [188,189,190,191,192,193] which is reported to cause tumors. A key role is played by viral E6 and E7 proteins, that dysregulate normal p53 function and degrade pRb, leading to impaired recognition of damaged DNA and disruption of important cell replication checkpoints [190, 194]. Despite FPV2 infects cats from the birth and lifelong, not all infected subjects develop SCC [195]. This evidence suggests that other factors may be involved in the development of this neoplasm. Moreover, it is not clear yet if PV-related cancer evolves necessarily from PV-induced plaques, or if it may arise on skin without precursor lesions [164, 176, 177]. Among the diseases recognized to be caused by PV there are also feline sarcoids. Like equine sarcoids, they are caused by aberrant infections with Bos taurus papillomavirus (BPV) [196, 197]. The etiological agent causing feline sarcoids is a delta papillomavirus previously known as FeSarPV. Nowadays it has been classified as BPV-14 and it has been highlighted is closer relation with BVP types 1, 2 and 13 [196, 197]. Feline sarcoids are mesenchymal neoplasms characterized by fibroblasts proliferation with epithelial hyperplasia and deep rete ridges [198]. They generally arise on face or digit, probably because these sites are frequently predisposed to trauma which allow viral penetration in the dermis [198]. Feline sarcoids are very rarely reported. One possible reason for these limited reports can be linked to misdiagnosis, since if only one portion of the tumor is submitted to histopathological analysis, without including epithelium, it is difficult to diagnose correctly. Moreover, as already discussed, this is a typical finding of cats living in country environment, which are often feral or subjects receiving less care than the housed ones [197]. Finally, sarcoids can spontaneously evolve in regressive lesions, further reducing the frequency of this finding [197].
The oncogenic mechanisms of PV are well known in humans, and there is strong evidence of similar pathways in dogs and cats. The integration of viral DNA is a preliminary and essential step in the oncogenetic process [199], as previously suggested by studies that linked it to both viral and host genome instability caused by oncoproteins E6 and E7, leading to double strand breaks [200]. For this reason, even though all viral genome is essential to the virus’s life cycle and regulation, oncoproteins E5, E6 and E7 have a key role in the process of cancer development [201].
It has been reported that E6 interacts with two different classes of molecules: proteins harboring the LxxLL motif and proteins with a PDZ domain. The ligase E6-associated protein belongs to the former class and, when associated to protein E6, is involved in p53 degradation through the proteasome, thus inducing cancer. This kind of interaction occurs both in low- and high-risk types. On the other hand, proteins containing PDZ domains are involved in many cell signaling pathways and their association to protein E6 is closely linked to malignant cell transformation [202]. Interaction of E6 with PDZ proteins is a prerogative of high-risk PV types, and, alone, it is sufficient to cause neoplasms through the induction of cell transformation [140]. These two mechanisms are common in human, canine and feline species, even though the ability of E6 to dysregulate p53 should be further investigated in dogs [140]. Furthermore, protein E6 has also been described to affect the production of antiviral cytokines in keratinocytes of immunodeficient dogs. In humans, E6 has also been reported to decrease Bax and Bak protein family content, preventing them from entering mitochondria and avoiding the activation of apoptosis [165].
In humans, E7 protein acts as oncogenic through proteasome-mediated pRB degradation [203]. pRB is a tumor suppressing protein whose degradation releases E2F and promotes the entry of the infected cells in the S phase of cell cycle. In cats, E7 acts in a similar way, while in dogs, E7 lacks the pRB binding site, and therefore, PV has been suggested to modify the cell cycle in other ways [140, 195]. E5 is the smallest and least studied PV oncoprotein and its oncogenic action is linked to its promoting activity on the expression of the epidermal growth factor receptor (EGFR). Moreover, in humans, E5 can bind endosomal vacuolar V-ATPase, affecting its activity and reducing vesicular transport [204]; it also promotes degradation of Bax, a proapoptotic protein, thus preventing apoptosis [205].
The case of BPV-14, etiological agent of feline sarcoids, deserves a separated discussion. Even if the oncogenic mechanism has not been fully understood yet, it has been recognized that E5 protein produced by delta BPVs has a key role in the neoplastic transformation process [196, 197]. This eventuality is linked to the binding and the activation of the platelet derived growth factor beta receptor (PDGFB-R), which is possible thanks to the presence of 4 specific amino acids within the E5 protein. This binding causes a cascade of events, like the activation of kinases and further mechanisms to support neoangiogenesis and immune evasion, leading to mesenchymal cell proliferation [206,207,208]. On the other hand, E6 protein lacks PDZ binding motif and E7 lacks retinoblastoma binding sites, further elements arguing in favor of an essential role of E5 protein in the BPV-14 caused oncogenic transformation [196].
Assuming that in recent years feline oral squamous cells carcinoma (FOSCC) are thought to be very similar to human head and neck squamous cells carcinoma (HNSCC) [15], and the role of FPV-2 as etiological factor for the development of these neoplasms is supported by several studies, new evidence suggests an emerging role of cats as spontaneous model for HPV related HNSCC [209]. The reasons for this speculation have their root in several pieces of evidence coming from a multitude of studies collected and revised by Altamura and Borzacchiello [210]. As already known, HPV-positive SCC are classified as distinct entities because they showed different molecular, genetic, and biological features than their HPV-negative counterpart. It is not yet clear whether the same is applicable to FOSCC. Given that this is one of the last pieces missing to the full confirmation of the reliability of such a model, Altamura and Borzacchiello launched an appeal for a collective effort in additional research and data collection on FPV [210]. Furthermore, a coordinated interest of researchers should also be promoted in the light to develop further preventing therapies, both for companion animals and humans. Indeed, it is noteworthy that prevention, one of the core principles of modern oncology, reached a milestone exactly in contrasting human papillomavirus, since a vaccination campaign has already been active for some years targeting adolescent girls. Vaccine effectiveness has been demonstrated to be high since the prevalence of HPV type 6/11/16/18 infection and, of consequence, the prevalence and incidence of genital warts, decreased in the targeting population [211], a great achievement that encourages the research effort on this area.
Gammaherpesvirus
With this paragraph, we will begin the discussion of viruses for which an oncogenic potential has already been confirmed in other species while they are currently under investigation in cats and dogs. For this reason, in this and the next two sections, we will start from solid data already present in the literature in the original species, while attempting to retrace the studies carried out in dogs and cats so far. Herpesviruses are the first viral agents described as oncogenic in human medicine and classified among class I carcinogens by the International Agency for Research on Cancer (IARC) [212]. They are double-stranded DNA viruses classified into three different subfamilies, Alfaherpesvirinae, Betaherpesvirinae and Gammaherpevirinae [213]. In humans, two Gammaharpesvirus are frequently associated with HIV-related neoplasms [214, 215], namely Epstein-Barr virus (EBV, Human Herpesvirus 4) and Kaposi’s sarcoma-associated herpesvirus (KSHV, Human Herpesvirus 8). KSHV belongs to the Rhadinovirus genus, it is responsible for epidemic Kaposi’s sarcoma, frequently associated with HIV infection [216, 217]. The EBV belongs to the Lymphocryptovirus genus [218]: nowadays, its causal role in transforming latently infected lymphocytes in HIV-positive patients is well described [214]. Although the infection is generally asymptomatic in immune-competent individuals during childhood [219, 220], infectious mononucleosis may be a manifestation in adolescents and early adults [221]. Furthermore, this virus has been linked to the development of some lymphoproliferative neoplasms, for instance Burkitt’s lymphoma, Hodgkin’s disease, diffuse large B cell lymphoma, T-cell lymphoma, [222, 223] and nasopharyngeal and gastric carcinoma [224, 225], since lymphoid and epithelial cells are both highly receptive and permissive for infection [226, 227]. From 1995 to the present days, a great number of studies suggests that EBV may also have a role in the development of breast cancer (BC) in women [228,229,230,231,232,233,234,235,236,237,238,239,240,241,242,243,244,245] alone or in association with other viruses [246, 247]. However, other authors did not succeed in confirming this association [229, 242, 248,249,250,251,252,253] and, despite the fact that epidemiological evidence points out a higher risk of BC in the presence of EBV, its role in the development of BC remains controversial [254]. A search for the involvement of EBV or an EBV-like virus in BC, as well as in similar feline and canine types of tumors, is under investigation.
EBV is characterized by a 184-kbp genome encoding more than 85 genes, including numerous oncogenes such as nuclear antigens (EBNA1, -2, -3A, -3B, -3, C, -LP), latent membrane proteins ((LMP)-1, -2A, -2B), and noncoding RNAs (EBERs and miRNAs) [255]. During infection, EBV alternates between two possible states, i.e. latent and lytic state. In the latent state, the virus genome exists as a closed circular plasmid DNA in the cell nucleus, incorporated with histones, and replicates at the same time as the host genome. It is inherited by progeny cells, and only a select number of genes are expressed, depending on infected tissue, state of cells, and immune condition [256,257,258,259].
Latency factors play a key role in cancer development as they promote cell proliferation. Three patterns of latency have been reported in humans. Burkitt lymphoma and gastric carcinoma generally undergo type I latency, where gene expression is limited to EBERs and EBNA1. Some Hodgkin lymphomas, nasopharyngeal carcinomas (NPC), and T/NK lymphomas express the previous genes together with LMP1 and LMP2 genes in the so-called type II latency. EBNA2, EBNA3, EBNA-LP, together with all the previous type I and type II genes, are typical of post-transplant lymphoproliferative disorders and lymphoblastoid cell lines (LCLs) in type III latency.
In the lytic cycle, all the viral genes are expressed at the same time to produce new virions. It has been suggested that, at the beginning of infection, EBV follows an abortive cycle, during which only the immediate-early and early genes are expressed without DNA replication. This short cycle is then silenced after a few weeks, and a latent phase begins. A number of cells may go back from the latent stage to an abortive phase and then become silenced again, while others may express late genes and replicate the viral genome, thus entering the complete lytic cycle, with production of new virions [260].
However, the lytic stage is also involved in carcinogenesis, as it causes intense production of cytokines and growth factors that are exploited by neoplastic cells for their metabolism. In humans, EBV oncogenesis has been reported to occur through three different mechanisms. Firstly, the direct activity of viral oncogenes (such as LMP-1 and -2, EBV nuclear antigens (EBNAs), EBV-encoded nonconding RNAs (EBERs) and microRNAs) is implicated by eliciting growth-promoting signals or mitogen-activated protein kinase, or by silencing tumor suppressors, with several mechanisms which have not yet been fully understood [261,262,263,264,265,266,267,268]. Secondly, an indirect effect occurs through suppression of the host immune system (e.g., systemic immunosuppression during AIDS or associated with transplant, B cell lymphoma and disorders causing downregulation of MHC molecules) [217, 269, 270], or by exploitation of the inflammatory status, when cytokines and growth factors are released to promote neoplastic growth. Finally, EBV oncogenesis may also be linked to genetic or epigenetic alterations of host DNA, such as myc traslocation in Burkitt lymphoma [257, 272] or introducing mutations in suppressing genes leading to their inactivation [263, 267, 273]. For more comprehensive reviews on the topic see Murata et al., 2014 [260]
The close relationship between pets and humans suggests that an EBV-related infection might exist in domestic animals. In 2005, Chiou et al. found an 88% prevalence of anti-EBV antibodies in 36 pet dog blood samples and identified the presence of an EBV-specific BamHI W fragment in 71% of the corresponding 21 leukocyte DNA samples. In addition, through in situ hybridization (ISH), they also highlighted EBER in dog blood and bone marrow [274]. In 2010, Milman et al. detected anti-EBV antibodies in 43/112 in dog sera coming from UK, 67/104 in canine serum samples from US, EBV-like sequences in 1/33 canine palatine tonsil and succeeded in confirmed their presence in 38/100 cat blood samples, too [275]. However, EBV was detected molecularly in tissues from one dog, while RT-PCR did not succeed in detecting transcripts associated with lytic infection or latency, thus suggesting that pets are exposed to EBV or EBV-related viruses, but with no signs of persistent infection in the analyzed tissues. Interestingly, the prevalence of exposure is higher in pet cats rather than in stray ones, suggesting the owners as the main source of virus.
Later research focused on dogs with tumors. In 2012, Huang et al. reported the presence of anti-EBV antibody both in healthy subjects and in those with spontaneous lymphoma, together with the finding of extracellular viral particles similar to EBV in cultures of canine malignant B cells. This suggested that an EBV-like virus could be involved in some steps of canine lymphomagenesis [276]. A significant step forward was made in 2013 by Chiu et al., who restricted their study to dogs with tumors, particularly focusing on oral tumors, since it had already been proven that EBV is transmitted through saliva in humans [277,278,279]. They found 80% of EBV DNA prevalence in 10 samples from canine oral tumors, together with the presence of transcripts of EBER, of the viral bcl2 homologue BHRF1, and of LMP1. Moreover, they confirmed the presence of virions by transmission electron microscopy, highlighting a similarity in size between these virions and EBV viral particles [280, 281]. Furthermore, the similarity of the canine EBV-like virus LMP1 and the identity of canine BHRB1 both to the EBV counterparts argued in favor of a close relationship between canine and human viruses [279].
Despite all these encouraging results, another recent study by Waugh et al. [282] did not find an association between EBV or a related virus and 112 samples from canine lymphomas. However, through serological analysis, these authors confirmed that EBV or a closely related virus circulates in dogs, albeit with a lower prevalence than the one previously found. Although dogs with cancers were more likely to be seropositive than the others, the authors did not find higher antibody titers in subjects with lymphomas, compared with those bearing other types of neoplasms, nor did they find different seroprevalences between dogs with B- and T-cell type lymphomas, as expected. Starting from the idea that herpesviruses are extremely species-specific and rarely cross species barrier, they also set up multiple degenerated PCR assays to evaluate the presence of other herpesviruses, related but different from EBV, in canine samples. Again, they found no evidence of the involvement of a Gammaherpesvirus in common canine lymphoma. Recently, EBV was suggested as a putative etiological agent in the development of breast cancer [283]. For this reason, a recent study focused on the search for the putative involvement of an EBV-like Gammaherpesvirus in canine mammary tumors, once it was found to infect dogs (as mentioned above). However, only one sample was found to be positive for EBV genes. Although the authors suggested that their results could have been affected by technical factors related to DNA quantity or quality or other unclear factors, they had to conclude that there was not sufficient evidence of EBV involvement in the carcinogenesis of canine mammary tumors [284].
Due to the close parallel between HIV and FIV, and the frequent association between EBV and HIV infections, a similar causal role for a putative Gammaherpesvirus in cancer is under investigation in cats. A novel feline Gammaherpesvirus, the Felis catus Gammaherpesvirus 1 (FcaGHV-1,) has been recently discovered [285]. Subsequent studies have demonstrated a worldwide prevalence ranging from 5% up to 25% based on the geographical area considered [286, 287]. It has been hypothesized that this virus is transmitted horizontally, sharing its route of infection through biting with FIV [188], and that age, male sex and concurrent FeLV or FIV infection may be predisposing factors for cats [260,261,262]. Further studies are needed to confirm these observations.
In a study, FcaGHV-1 DNA was detected in 40.4% out of 104 FIV infected cats and it was estimated that a FIV-positive and haemoplasma-positive subjects was respectively 4, 5 [286, 287] and 16 times [288] more likely to be FcaGHV1 positive compared to the negative cats [288]. FcaGHV-1 viral particles have been also detected in the bone marrow and small intestine of cats with alimentary lymphoma [286]. In one of those cases, researchers succeeded in demonstrating and isolating the viral genome through quantitative PCR (qPCR) [213]. More recently, Aghazadeh et al. highlighted FcaGHV-1 DNA in one FIV-associated alimentary lymphoma through in situ hybridization (ISH) [289]. To sum up, even if the available data support a pathogenic role for this virus, at least in cats, further studies are needed to elucidate its involvement in oncogenesis.
Hepadnavirus
The discovery of a Hepatitis B-like virus in domestic animals is relatively recent. Its prototype is the Hepatitis B virus (HBV), which has strict tropism for the liver and causes chronic hepatitis and hepatocellular carcinoma (HCC) in humans [290]. In 2018, a new virus was detected for the first time in an Australian FIV-positive cat during transcriptomics studies [291]. This virus has been named Domestic Cat Hepadnavirus (DCH) and was classified as a circular, partially double stranded DNA virus belonging to the Orthohepadnavirus genus, Hepadnaviridae family. Similar to the others Hepadnaviruses, its genome encodes four overlapping reading frames (ORFs) for the polymerase (P), surface (S), core (C), and X proteins [291, 292]. Although DCH belongs to the same genus as HBV, the real function of DCH viral proteins should be elucidated due to the phylogenetic divergence between DCH and HBV [291, 293, 294].
HBV oncogenesis is a multifactorial process. The first mechanism of HBV-related HCC induction is linked to chronic inflammation. HBV infection induces chronic inflammation in the liver, leading to regulatory T cell (Treg) disfunction, increased cytokine production (TGF-ß, IL-4, IL-10, IL-12, IL-13) and alteration of specific signaling pathways that ultimately increase the risk of hepatocarcinogenesis. Furthermore, HBV can cause oxidative stress with increased ROS production and ROS-induced DNA damage, frequently associated with chromosomal aberrations and cellular transformation. HBV-DNA integrations have been detected close to certain gene targets, suggesting that viral DNA integration in the host genome may be another way through which HBV can drive malignant transformation. Lastly, other specific mechanisms involving selected viral proteins (HBs and HBx), microRNA expression, DNA methylation modifications, and related epigenetic changes have been associated with HCC development. For a more comprehensive review on the issue, see Stella et al. [295].
Over the years, molecular studies have been carried out in several countries to test the prevalence of DCH in cat populations. Recent studies highlighted that 6.5% sera of 123 Australian cats [289] and 10.8% of 390 Italian ones [296] were DCH-positive and showed active viremia. In Malaysia, DCH has been detected in 12.3% of 253 blood samples and 14.9% of 87 liver samples [297]. In Thailand, DCH was confirmed in 12.4% out of 209 sera and in 20% of both sera and organs from 15 necropsies [294]. A multicentric study including 86 liver biopsies samples from the United States, United Kingdom, Australia, and New Zealand confirmed the presence of DCH DNA in cats with selected pathologies [298]. In 2022, a Japanese PCR-based study found a lower prevalence of 0.78% in 139 feline blood samples [299]. Furthermore, as HBV and Hepatitis C virus co-infection in humans was hypothesized to increase the risk of uveitis [300], DCH was also analyzed in subjects with uveitis where the virus was detected in 2 out of 65 cats, while the healthy control group resulted to be negative [301].
Given the possibility of inter-species transmission, the evidence that some viruses circulate among both dogs and cats [302], and the presence of antibodies specific to HBV in canine sera from several studies [303,304,305,306], it was hypothesized that HBV-like viruses might also be harbored by dogs. In 2019, Hepadnavirus DNA was detected in dogs for the first time using PCR targeting the preS/S1 genomic region and the core gene of HBV. Hepadnavirus DNA was identified in 10% of sampled dogs [306]. In 2022, another study reported that 6.3% of canine sera samples tested positive by qPCR, and the virus became known as Domestic Dog Hepadnavirus (DDH) [307]. DDH genome showed 98.0% nucleotide identity at the whole genome level with the Italian DCH and 96.9% with the Australian one [308].
Over time, similarities have been described between Hepadnaviruses infecting domestic animals and HBV. Piewbang et al. suggested that the localization and maturation of Dane-like particles in liver of cats analyzed by transmission electron microscopy was ultra-structurally similar to those observed for HBV [294, 309, 310]. Moreover, different studies involving infected cats have reported an elevation in liver disease-related markers (such as serum alanine aminotransferase) similar to HBV-infected humans [296, 297, 311] leading to the suspicion that DCH in cats may reflect similar tropism and pathogenesis to that of HBV in humans [294]. This suspicion prompted Pesavento et al. [298] to focus their research on a putative correlation between DCH and liver damage in cats. They analyzed liver biopsies from healthy (n = 15) and diseased (n = 71) cats through PCR and ISH and reported that a conspicuous percentage of subjects with liver diseases already reported to be associated to HBV infection in humans were positive for DCH, namely 43% (6/14) of chronic hepatitis cases and 28% (8/29) of HCCs. Conversely, subjects with bile duct-associated liver disease and healthy ones were negative for DCH.
Histologic features of hepatitis and hepatic neoplasia in cats have been described as extremely similar to those observed in HBV-related diseases in humans, strengthening the speculation that DCH may be associated with hepatitis and HCC in cats. Similar alterations in liver disease-related markers have been highlighted in DDH-positive dogs as well [307], but Choi et al. [312] did not succeed in confirming a contribution of Hepadnavirus in either chronic hepatitis or HCC [312]. Data from all these studies suggest that DCH infection is associated with immunocompromised conditions in cats [291, 293, 295, 296]. Moreover, due to viral particle detection in serum, whole blood, heart, lungs, intestines, kidneys, and spleen, [291, 293, 295, 296] researchers hypothesize that DCH may spread from one cat to another through blood, but not through semen or percutaneously, as for HVB infection [308, 313]. These observations were also supported by the longitudinal observation of two DCH-positive cats that constantly yielded negative DCH amplification PCR results of oral, conjunctival, and preputial swabs [314]. Regarding fecal transmission, although Capozza et al. [314] found only negative rectal swabs in their longitudinal studies, Piewbang et al. [294] found high numbers of DCH genome copies in intestinal samples and the expression of viral protein through IHC in cells from intestinal villi of cats. However, due to the limited numbers of studies, knowledge on the pathobiology of DCH and DDH is inconclusive, and whether these viruses are apathogenic or may have a potential role in certain clinical diseases remains to be elucidated.
Mouse mammary tumor virus
MMTV infection is presently considered a proven risk factor for the development of mammary carcinoma in mice. Its discovery dates back to 1936, when John Bittner highlighted that mice with mammary carcinoma could transmit a factor capable of causing the development of the same cancer in the offspring that fed on infected milk [315]. This agent, known from the beginning as “milk factor”, was classified as a retrovirus in 1966 and later became known as Mouse Mammary Tumor Virus. Since its discovery, speculation has been made about the existence of a similar viral agent linked to human breast cancer (BC) first, and to neoplasms of domestic animals later. MMTV is a Betaretrovirus of the Retroviridae family. While it was initially classified as a simple retrovirus, now it is classified as a complex one since it has been discovered that its genome encodes not only structural proteins, but also at least three regulatory and accessory proteins. Between two long terminal repeats (LTRs) located at 5ʹ and 3ʹ like for all retroviruses, there are 4 ORFs encoding gag proteins, protease (PR), pol proteins and env proteins. MMTV LTRs encode two additional genes, sag and rem, which are translated in a superantigen and an RNA exporting protein, respectively. Sag is a transmembrane protein with an essential role in tumor development, since it is responsible of the efficient transmission of viral particles from the gut to the mammary gland: its action is essential to the lymphocytes amplification of cognate cells deriving from the Sag-MHC complex recognition by specific T-cells, but it is not required for the initial infection [316]. Rem, which should be associated with a Rem-responsive element located on MMTV RNA, is responsible for the transport of unspliced viral RNAs [317]. Furthermore, MMTV LTRs are also important since they encode hormone response elements (HREs) upregulating viral production [318, 319], in addition to negative regulatory elements (NREs) inhibiting viral transcription [320], and a transcription enhancer factor-1 (TEF-1) binding site, involved in tumorigenesis [321].
Over time, an MMTV variant, the type B leukemogenic virus (TBLV), was discovered. This is involved in the development of lymphomas, particularly T-type ones, in mice [322, 323]. It is the result of defined LTR modifications, such as the loss of NREs, multiplication of regulatory elements, and the coding of T cell-specific transcriptional enhancers [324, 325]. These modifications cause the virus to shift its ability from causing mammary cancer to inducing lymphoma.
The viral cycle starts when exogenous virions in nursing mice milk reach the gut of suckling pups, infecting first dendritic cells and B lymphocytes in the Peyer’s patches. These cells process the Sag antigen and expose it on their surface, in the context of the major histocompatibility complex (MHC) class II. Sag-activated T-lymphocytes, in turn, further stimulate the proliferation of dendritic cells, B- and T-lymphocytes, and establish a reservoir of both permissive and infected cells. Hence, the virus exploits immune cells to reach its target, the mammary gland, where infection of mammary epithelial cells occurs during hormonal stimulation typical of puberty and pregnancy, when they are prone to divide [316]. For this reason, there is a period of latency between the time of ingestion of infected milk and tumor development. Once hormonal stimulation starts, MMTV binds to host cellular Transferrin Receptor 1 (TfR1) and passes the cell membrane within a low-pH endosome. Inside the cell, the viral genome is reverse-transcribed and delivered to the nucleus, where integration into the host genome as proviral DNA occurs [316]. To produce virions, viral RNA has to be translated, and both translation products and progeny RNAs are assembled into virions and released through budding from the host cells. On the other hand, amplification of the virus is necessary for tumors to develop. It is believed that MMTV infects multipotent mammary stem cells during hormonal circumstances, increasing the regenerative activity of the mammary gland [326, 327]. At first, MMTV causes a hormone-dependent hyperplastic alveolar nodule (HAN), followed by a hormone-independent phase of tumorigenesis [316, 328]. It has been suggested that MMTV-induced mammary carcinoma is a monoclonal tumor, with all neoplastic cells originating from one individually transformed and expanded stem cell [329,330,331,332,333,334]. MMTV is classified as a non-acutely transforming retrovirus, since it does not contain oncogenes. To develop cancer, the virus has to integrate its genome close to cellular proto-oncogenes, which are activated and overexpressed through the action of LTR promoters or enhancers. Particularly, the most common integration sites (CIS) involved in tumorigenesis of MMTV-related mammary tumors are members of wnt, fgf, rspo, notch4/int3, and eIF3e/int6 [316, 334,335,336]. Activation of multiple genes is required for tumorigenesis [316], leading to disruption of cell signaling pathways, dysregulation in cell proliferation and differentiation and, finally, to tumor development. TBLV, the MMTV variant causing lymphoma, has some structural differences in U3 region of LTRs allowing a different target. Particularly, TBLV U3 region is characterized by a deletion of NREs and multiple flanking sequences encoding for lymphocyte-specific transcriptional enhancers [324]. These structural modifications are per se sufficient to cause lymphoma, rather than mammary carcinoma [325], although the mechanisms of viral oncogenesis remain the same [337,338,339,340]. TBLV CIS are members of myc, rorc, notch1 and tblvi1 gene families. Furthermore, it has been suggested that, differently from mammary tumors, MMTV lymphomas are polyclonal neoplasms and require additional infection and proviral integration events for their development [325].
Although a considerable number of studies have been conducted, evidence of the hypothetical involvement of an MMTV-like virus in neoplasms of human and domestic animals is still weak, as reported by Amarante et al., Szabo et al., and Parisi et al. [341,342,343]. Specifically, the suggestion of such etiology in the development of BC has been the subject of a long controversy between researchers who support this hypothesis and those who do not [341]. However, over time, compelling evidence has accumulated both in vivo and in vitro, and it is now generally accepted that an MMTV-like virus, known as Human Mammary Tumor Virus (HMTV), is associated with BC. The mechanisms of oncogenesis and common viral integration sites have been studied [343].
It has been highlighted that MMTV infects a variety of murine organs, such as kidney, salivary glands, and the male genital system [344,345,346], and that TBLV is involved in T-cell lymphoma development [322, 323, 347, 348]. Similarly, in humans, sequences of a virus very closely resembling MMTV have been found in other tissues, in addition to T cells, which has given rise to the hypothesis that an MMTV-like virus may be also associated with human lymphomas [349,350,351], hepatic carcinomas [352], and other diseases affecting the liver, such as primary biliary cirrhosis [353,354,355]. MMTV-like virus has been associated not only with T-cell lymphomas, as in mice [356,357,358], but also with B-type lymphoma. Moreover, MMTV-like sequences have been found in other neoplasms, such as those in the ovary, prostate, skin [359], and endometrial carcinoma [360], although the role of the virus in the development of these human cancers still needs to be clarified. Viral sequences have also been found in saliva, blood, and milk. It has been suggested that the presence of the virus in saliva may follow inter-human spread [361].
In an effort to provide further insight into the epidemiology of this virus, Stewart et al. [362, 363] noted that North West Europe, the European area with the highest incidence of BC, matched geographically with areas where a particular mouse strain, Mus domesticus, was a resident species. This evidence led to the suspicion that the virus could spread from mice to humans using domestic animals, such as cats and dogs, as intermediary hosts. In recent years, veterinary medicine studies focused on the search for a putative MMTV-like virus in dogs and cats. The first evidence of MMTV-like sequences was found in 2005, when MMTV-like sequences were amplified from the thymus of a kitten and the spleen of an adult cat [364]. In 2012, Hsu et al. investigated mammary tumors from 145 dogs and 11 cats, targeting MMTV-like env and LTR sequences. They found a prevalence of 3.49% (3/145) and 18.6% (16/145) for env and LTR sequences in dogs, respectively, and 22.22% (2/11) for both targets in cats [365]. The amplificon sequences shared 94% and 98% similarity with MMTV and HMTV, respectively. Additionally, they detected the presence of these sequences in normal mammary tissue from both cats and dogs. Two subsequent studies confirmed the presence of an MMTV-like sequence sharing high homology with MMTV and HMTV in feline mammary tumors, with a prevalence of 7% (7/86) [366] and 12.5% (3/24) [367], but not in healthy tissue from both cats and dogs, nor in canine mammary tumors. Preliminary research on feline lymphomas showed that MMTV-like sequences may also be found also in this kind of neoplasm [368]. Particularly, the authors found a 9.4% prevalence (5/53) of MMTV-like sequences and a strong association with lymphoma localized in the nasal cavity, since two out of the three nasal lymphomas included in the study were found to be positive. However, all these are only preliminary results on an interesting topic that requires further investigation. Due to the evidence of MMTV involvement in carcinogenesis in humans and mice, any similar etiological agents should be identified in domestic animals, to develop coordinated therapies and prevention measures and to investigate their hypothetical zoonotic potential.
Conclusions
Viral oncology is an emergent topic continually under research. Knowledge about viruses involved in the tumor formation process in dogs and cats is essential, both as pathogens for their original host and as animal models for human diseases. Regarding the last aspect, particular attention should be paid to PV-positive FOSCCs, due to the most recent evidence suggesting that the cat may be a potential animal model for PV-induced HNSCCs in humans. However, knowledge on oncogenic viruses of companion animals relies almost entirely, although with some exceptions, on individual studies and research carried out by isolated working groups, lacking in some cases an overall picture. Moreover, in this field, a joint effort of veterinary and human medicine is crucial since the results obtained by investigating the molecular mechanisms may often be of interest to both species. With this work we would like to give an overview of the progress made in canine and feline oncovirus field, also highlighting the most relevant aspects of their human counterpart. Further studies on the pathophysiological mechanisms underlying the interactions between viruses and cancer are encouraged. They would support the hypothesis of a role of putative oncoviruses in tumor formation, solidify knowledge about already proven oncogenic viruses, and develop new medical treatments in human and veterinary medicine.
Availability of data and materials
Not applicable.
References
Ferraris ZA, Ferraris VA. The women of Salerno: contribution to the origins of surgery from medieval Italy. Ann Thorac Surg. 1997;64(6):1855–7.
Kattner P, Zeiler K, Herbener VJ, et al. What animal cancers teach us about human biology. Theranostics. 2021;11(14):6682–702. https://doi.org/10.7150/thno.56623.
Truyen U, Löchelt M. Relevant oncogenic viruses in veterinary medicine: original pathogens and animal models for human disease. Contrib Microbiol. 2006;13:101–17. https://doi.org/10.1159/000092968.
Rous P. A transmissible avian neoplasm. (Sarcoma of the common fowl). J Exp Med. 1910;12(5):696–705. https://doi.org/10.1084/jem.12.5.696.
Rous P. A sarcoma of the fowl transmissible by an agent separable from the tumor cells. J Exp Med. 1911;13(4):397–411. https://doi.org/10.1084/jem.13.4.397.
Epstein MA, Achong BG, Barr YM. Virus particles in cultured lymphoblasts from Burkitt’s lymphoma. Lancet. 1964;1(7335):702–3. https://doi.org/10.1016/s0140-6736(64)91524-7.
Boyle P, Levin B. World cancer report 2008. Lyon, France: International Agency for Research on Cancer; 2008.
Bouvard V, Baan R, Straif K, et al. A review of human carcinogens–part B: biological agents. Lancet Oncol. 2009;10(4):321–2. https://doi.org/10.1016/s1470-2045(09)70096-8.
de Martel C, Ferlay J, Franceschi S, et al. Global burden of cancers attributable to infections in 2008: a review and synthetic analysis. Lancet Oncol. 2012;13(6):607–15. https://doi.org/10.1016/S1470-2045(12)70137-7.
Mui UN, Haley CT, Tyring SK. Viral oncology: molecular biology and pathogenesis. J Clin Med. 2017;6(12):111. https://doi.org/10.3390/jcm6120111.
Sabin AB. Oral poliovirus vaccine. History of its development and prospects for eradication of poliomyelitis. JAMA. 1965;194(8):872–6. https://doi.org/10.1001/jama.194.8.872.
King AJ. The use of animal models in diabetes research. Br J Pharmacol. 2012;166(3):877–94. https://doi.org/10.1111/j.1476-5381.2012.01911.x.
Litynski GS. Laparoscopy–the early attempts: spotlighting Georg Kelling and Hans Christian Jacobaeus. JSLS. 1997;1(1):83–5.
Haddad FS. Ibn Zuhr and experimental tracheostomy and tracheotomy. J Am Coll Surg. 2004;199(4):665. https://doi.org/10.1016/j.jamcollsurg.2004.06.015.
Giuliano A. Companion animal model in translational oncology; feline oral squamous cell carcinoma and canine oral melanoma. Biology. 2021;11(1):54. https://doi.org/10.3390/biology11010054.
Schiffman M, Wentzensen N, Wacholder S, Kinney W, Gage JC, Castle PE. Human papillomavirus testing in the prevention of cervical cancer. J Natl Cancer Inst. 2011;103(5):368–83. https://doi.org/10.1093/jnci/djq562.
Schiller JT, Lowy DR. Virus infection and human cancer: an overview. Recent Results Cancer Res. 2014;193:1–10. https://doi.org/10.1007/978-3-642-38965-8_1.
Hill A. Statistical evidence and inference. In: Principles of medical statistics, 9th ed. Oxford; 1971.
Chang Y, Moore PS, Weiss RA. Human oncogenic viruses: nature and discovery. Philos Trans R Soc Lond B Biol Sci. 2017;372(1732):20160264. https://doi.org/10.1098/rstb.2016.0264.
Lawson JS, Glenn WK. Mouse mammary tumour virus (MMTV) in human breast cancer-the value of bradford hill criteria. Viruses. 2022;14(4):721. https://doi.org/10.3390/v14040721.PMID:35458452;PMCID:PMC9028876.
Lutz H, Addie D, Belák S, et al. Feline leukaemia. ABCD guidelines on prevention and management. J Feline Med Surg. 2009;11(7):565–74. https://doi.org/10.1016/j.jfms.2009.05.005.
Roy-Burman P. Endogenous env elements: partners in generation of pathogenic feline leukemia viruses. Virus Genes. 1995;11(2–3):147–61. https://doi.org/10.1007/BF01728655.
Beatty J. Viral causes of feline lymphoma: retroviruses and beyond. Vet J. 2014;201(2):174–80. https://doi.org/10.1016/j.tvjl.2014.05.026.
Donahue PR, Quackenbush SL, Gallo MV, et al. Viral genetic determinants of T-cell killing and immunodeficiency disease induction by the feline leukemia virus FeLV-FAIDS. J Virol. 1991;65(8):4461–9. https://doi.org/10.1128/JVI.65.8.4461-4469.1991.
Anderson MM, Lauring AS, Burns CC, Overbaugh J. Identification of a cellular cofactor required for infection by feline leukemia virus. Science. 2000;287(5459):1828–30. https://doi.org/10.1126/science.287.5459.1828.
Anderson MM, Lauring AS, Robertson S, Dirks C, Overbaugh J. Feline Pit2 functions as a receptor for subgroup B feline leukemia viruses. J Virol. 2001;75(22):10563–72. https://doi.org/10.1128/JVI.75.22.10563-10572.2001.
Mendoza R, Anderson MM, Overbaugh J. A putative thiamine transport protein is a receptor for feline leukemia virus subgroup A. J Virol. 2006;80(7):3378–85. https://doi.org/10.1128/JVI.80.7.3378-3385.2006.
Takeuchi Y, Vile RG, Simpson G, O’Hara B, Collins MK, Weiss RA. Feline leukemia virus subgroup B uses the same cell surface receptor as gibbon ape leukemia virus. J Virol. 1992;66(2):1219–22. https://doi.org/10.1128/JVI.66.2.1219-1222.1992.
Quigley JG, Burns CC, Anderson MM, et al. Cloning of the cellular receptor for feline leukemia virus subgroup C (FeLV-C), a retrovirus that induces red cell aplasia. Blood. 2000;95(3):1093–9.
Tailor CS, Willett BJ, Kabat D. A putative cell surface receptor for anemia-inducing feline leukemia virus subgroup C is a member of a transporter superfamily. J Virol. 1999;73(8):6500–5. https://doi.org/10.1128/JVI.73.8.6500-6505.1999.
Dunham SP, Graham E. Retroviral infections of small animals. Vet Clin North Am Small Anim Pract. 2008;38(4):879–ix. https://doi.org/10.1016/j.cvsm.2008.03.005.
Rolph KE, Cavanaugh RP. Infectious causes of neoplasia in the domestic cat. Vet Sci. 2022;9(9):467. https://doi.org/10.3390/vetsci9090467.
Hardy WD Jr, Hess PW, MacEwen EG, et al. Biology of feline leukemia virus in the natural environment. Cancer Res. 1976;36(2 pt 2):582–8.
Jarrett WF, Crawford EM, Martin WB, Davie F. A virus-like particle associated with leukemia (lymphosarcoma). Nature. 1964;202:567–9. https://doi.org/10.1038/202567a0.
Jarrett WF, Martin WB, Crighton GW, Dalton RG, Stewart MF. Transmission experiments with leukemia (lymphosarcoma). Nature. 1964;202:566–7. https://doi.org/10.1038/202566a0.
Hardy WD. Haematopoietic tumours of cats. J Am Anim Hosp Assoc. 1981;17:921–40.
Hartmann K. Clinical aspects of feline immunodeficiency and feline leukemia virus infection. Vet Immunol Immunopathol. 2011;143(3–4):190–201. https://doi.org/10.1016/j.vetimm.2011.06.003.
Hartmann, K. Feline leukemia virus infection. In: Greene CE, editor. Infectious diseases of the dog and cat, 4th ed. Elsevier Saunders: St Louis; 2012. p. 108–36.
Rojko JL, Hoover EA, Quackenbush SL, Olsen RG. Reactivation of latent feline leukaemia virus infection. Nature. 1982;298(5872):385–8. https://doi.org/10.1038/298385a0.
Pacitti AM, Jarrett O, Hay D. Transmission of feline leukaemia virus in the milk of a non-viraemic cat. Vet Rec. 1986;118(14):381–4. https://doi.org/10.1136/vr.118.14.381.
McClelland AJ, Hardy WD, Zuckerman EE. Prognosis of healthy feline leukemia virus infected cats. In: Hardy WD, Essex M, McClelland AJ, editors. Feline leukemia virus, vol. 4. North Holland: Elsevier; 1980. p. 121–6.
Levy JK, Scott HM, Lachtara JL, Crawford PC. Seroprevalence of feline leukemia virus and feline immunodeficiency virus infection among cats in North America and risk factors for seropositivity. J Am Vet Med Assoc. 2006;228(3):371–6. https://doi.org/10.2460/javma.228.3.371.
Hardy WD Jr. Immunopathology induced by the feline leukemia virus. Springer Semin Immunopathol. 1982;5(1):75–106. https://doi.org/10.1007/BF00201958.
Hoover EA, Mullins JI. Feline leukemia virus infection and diseases. J Am Vet Med Assoc. 1991;199(10):1287–97.
Stewart MA, Forrest D, McFarlane R, Onions D, Wilkie N, Neil JC. Conservation of the c-myc coding sequence in transduced feline v-myc genes. Virology. 1986;154(1):121–34. https://doi.org/10.1016/0042-6822(86)90435-6.
Hardy WD Jr, McClelland AJ, Zuckerman EE, et al. Development of virus non-producer lymphosarcomas in pet cats exposed to FeLv. Nature. 1980;288(5786):90–2. https://doi.org/10.1038/288090a0.
Reinacher M. Diseases associated with spontaneous feline leukemia virus (FeLV) infection in cats. Vet Immunol Immunopathol. 1989;21(1):85–95. https://doi.org/10.1016/0165-2427(89)90132-3.
Shelton GH, Grant CK, Cotter SM, Gardner MB, Hardy WD Jr, DiGiacomo RF. Feline immunodeficiency virus and feline leukemia virus infections and their relationships to lymphoid malignancies in cats: a retrospective study (1968–1988). J Acquir Immune Defic Syndr. 1990;3(6):623–30.
Forrest D, Onions D, Lees G, Neil JC. Altered structure and expression of c-myc in feline T-cell tumours. Virology. 1987;158(1):194–205. https://doi.org/10.1016/0042-6822(87)90253-4.
Miura T, Tsujimoto H, Fukasawa M, et al. Structural abnormality and over-expression of the myc gene in feline leukemias. Int J Cancer. 1987;40(4):564–9. https://doi.org/10.1002/ijc.2910400422.
Miura T, Shibuya M, Tsujimoto H, Fukasawa M, Hayami M. Molecular cloning of a feline leukemia provirus integrated adjacent to the c-myc gene in a feline T-cell leukemia cell line and the unique structure of its long terminal repeat. Virology. 1989;169(2):458–61. https://doi.org/10.1016/0042-6822(89)90172-4.
Levy LS, Gardner MB, Casey JW. Isolation of a feline leukaemia provirus containing the oncogene myc from a feline lymphosarcoma. Nature. 1984;308(5962):853–6. https://doi.org/10.1038/308853a0.
Levy LS, Lobelle-Rich PA, Overbaugh J, Abkowitz JL, Fulton R, Roy-Burman P. Coincident involvement of flvi-2, c-myc, and novel env genes in natural and experimental lymphosarcomas induced by feline leukemia virus. Virology. 1993;196(2):892–5. https://doi.org/10.1006/viro.1993.1553.
Tsatsanis C, Fulton R, Nishigaki K, et al. Genetic determinants of feline leukemia virus-induced lymphoid tumors: patterns of proviral insertion and gene rearrangement. J Virol. 1994;68(12):8296–303. https://doi.org/10.1128/JVI.68.12.8296-8303.1994.
Neil JC, Hughes D, McFarlane R, et al. Transduction and rearrangement of the myc gene by feline leukaemia virus in naturally occurring T-cell leukaemias. Nature. 1984;308(5962):814–20. https://doi.org/10.1038/308814a0.
Rohn JL, Linenberger ML, Hoover EA, Overbaugh J. Evolution of feline leukemia virus variant genomes with insertions, deletions, and defective envelope genes in infected cats with tumors. J Virol. 1994;68(4):2458–67. https://doi.org/10.1128/JVI.68.4.2458-2467.1994.
Levesque KS, Bonham L, Levy LS. flvi-1, a common integration domain of feline leukemia virus in naturally occurring lymphomas of a particular type. J Virol. 1990;64(7):3455–62. https://doi.org/10.1128/JVI.64.7.3455-3462.1990.
Levy LS, Lobelle-Rich PA. Insertional mutagenesis of flvi-2 in tumors induced by infection with LC-FeLV, a myc-containing strain of feline leukemia virus. J Virol. 1992;66(5):2885–92. https://doi.org/10.1128/JVI.66.5.2885-2892.1992.
Levy LS, Lobelle-Rich PA, Overbaugh J. flvi-2, a target of retroviral insertional mutagenesis in feline thymic lymphosarcomas, encodes bmi-1. Oncogene. 1993;8(7):1833–8.
Tsujimoto H, Fulton R, Nishigaki K, et al. A common proviral integration region, fit-1, in T-cell tumors induced by myc-containing feline leukemia viruses. Virology. 1993;196(2):845–8. https://doi.org/10.1006/viro.1993.1544.
Fujino Y, Ohno K, Tsujimoto H. Molecular pathogenesis of feline leukemia virus-induced malignancies: insertional mutagenesis. Vet Immunol Immunopathol. 2008;123(1–2):138–43. https://doi.org/10.1016/j.vetimm.2008.01.019.
Fulton R, Plumb M, Shield L, Neil JC. Structural diversity and nuclear protein binding sites in the long terminal repeats of feline leukemia virus. J Virol. 1990;64(4):1675–82. https://doi.org/10.1128/JVI.64.4.1675-1682.1990.
Neil J, Fulton R, Rigby M, Stewart M. Feline leukaemia virus: generation of pathogenic and oncogenic variants. In: Retroviral insertion and oncogene activation, Springer; 1991. p. 67–93.
Rohn JL, Overbaugh J. In vivo selection of long terminal repeat alterations in feline leukemia virus-induced thymic lymphomas. Virology. 1995;206(1):661–5. https://doi.org/10.1016/s0042-6822(95)80085-9.
Chandhasin C, Lobelle-Rich P, Levy LS. Feline leukaemia virus LTR variation and disease association in a geographical and temporal cluster. J Gen Virol. 2004;85(Pt 10):2937–42. https://doi.org/10.1099/vir.0.80149-0.
Hardy WD Jr. The feline sarcoma viruses. J Am Anim Hosp Assoc. 1981;17:981–97.
Bonham L, Lobelle-Rich PA, Henderson LA, Levy LS. Transforming potential of a myc-containing variant of feline leukemia virus in vitro in early-passage feline cells. J Virol. 1987;61(10):3072–81. https://doi.org/10.1128/JVI.61.10.3072-3081.1987.
Frankel AE, Gilbert JH, Porzig KJ, Scolnick EM, Aaronson SA. Nature and distribution of feline sarcoma virus nucleotide sequences. J Virol. 1979;30(3):821–7. https://doi.org/10.1128/JVI.30.3.821-827.1979.
Sarma PS, Baskar JF, Gilden RV, Gardner MB, Huebner RJ. In vitro isolation and characterization of the GA strain of feline sarcoma virus. Proc Soc Exp Biol Med. 1971A;137(4):1333–6. https://doi.org/10.3181/00379727-137-35783.
Sarma PS, Log T, Theilen GH. ST feline sarcoma virus. Biological characteristics and in vitro propagation. Proc Soc Exp Biol Med. 1971;137(4):1444–8.
Sarma PS, Sharar AL, McDonough S. The SM strain of feline sarcoma virus. Biologic and antigenic characterization of virus. Proc Soc Exp Biol Med. 1972;140(4):1365–8. https://doi.org/10.3181/00379727-140-36675.
Abujamra AL, Faller DV, Ghosh SK. Mutations that abrogate transactivational activity of the feline leukemia virus long terminal repeat do not affect virus replication. Virology. 2003;309(2):294–305. https://doi.org/10.1016/s0042-6822(03)00069-2.
Sykes JE. Feline immunodeficiency virus infection. Canine Feline Infect Dis. 2014. https://doi.org/10.1016/B978-1-4377-0795-3.00021-1.
Pedersen NC, Ho EW, Brown ML, Yamamoto JK. Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science. 1987;235(4790):790–3. https://doi.org/10.1126/science.3643650.
Magden E, Quackenbush SL, VandeWoude S. FIV associated neoplasms–a mini-review. Vet Immunol Immunopathol. 2011;143(3–4):227–34. https://doi.org/10.1016/j.vetimm.2011.06.016.
Gruffydd-Jones TJ, Hopper CD, Harbour DA, Lutz H. Serological evidence of feline immunodeficiency virus infection in UK cats from 1975–76. Vet Rec. 1988;123(22):569–70. https://doi.org/10.1136/vr.123.22.569.
Grindem CB, Corbett WT, Ammerman BE, Tomkins MT. Seroepidemiologic survey of feline immunodeficiency virus infection in cats of Wake County, North Carolina. J Am Vet Med Assoc. 1989;194(2):226–8.
Furuya T, Kawaguchi Y, Miyazawa T, et al. Existence of feline immunodeficiency virus infection in Japanese cat population since 1968. Nihon Juigaku Zasshi. 1990;52(4):891–3. https://doi.org/10.1292/jvms1939.52.891.
Bandecchi P, Matteucci D, Baldinotti F, et al. Prevalence of feline immunodeficiency virus and other retroviral infections in sick cats in Italy. Vet Immunol Immunopathol. 1992;31(3–4):337–45. https://doi.org/10.1016/0165-2427(92)90020-q.
Ueland K, Lutz H. Prevalence of feline leukemia virus and antibodies to feline immunodeficiency virus in cats in Norway. Zentralbl Veterinarmed B. 1992;39(1):53–8. https://doi.org/10.1111/j.1439-0450.1992.tb01137.x.
Hartmann K. Feline immunodeficiency virus infection: an overview. Vet J. 1998;155(2):123–37. https://doi.org/10.1016/s1090-0233(98)80008-7.
Hosie MJ, Addie D, Belák S, et al. Feline immunodeficiency. ABCD guidelines on prevention and management. J Feline Med Surg. 2009;11(7):575–84. https://doi.org/10.1016/j.jfms.2009.05.006.
Elder JH, Sundstrom M, de Rozieres S, de Parseval A, Grant CK, Lin YC. Molecular mechanisms of FIV infection. Vet Immunol Immunopathol. 2008;123(1–2):3–13. https://doi.org/10.1016/j.vetimm.2008.01.007.
Yamamoto JK, Hansen H, Ho EW, et al. Epidemiologic and clinical aspects of feline immunodeficiency virus infection in cats from the continental United States and Canada and possible mode of transmission. J Am Vet Med Assoc. 1989;194(2):213–20.
Addie D, Belák S, Boucraut-Baralon C, et al. Feline infectious peritonitis. ABCD guidelines on prevention and management. J Feline Med Surg. 2009;11(7):594–604. https://doi.org/10.1016/j.jfms.2009.05.008.
Sellon RK, Hartmann K. Feline immunodeficiency virus infection. In: Greene CE, editor. Infectious diseases of the dog and cat. 4th ed. St Louis: Elsevier Saunders; 2012. p. 136–49.
Beatty JA, Willett BJ, Gault EA, Jarrett O. A longitudinal study of feline immunodeficiency virus-specific cytotoxic T lymphocytes in experimentally infected cats, using antigen-specific induction. J Virol. 1996;70(9):6199–206. https://doi.org/10.1128/JVI.70.9.6199-6206.1996.
Fevereiro M, Roneker C, Laufs A, Tavares L, de Noronha F. Characterization of two monoclonal antibodies against feline immunodeficiency virus gag gene products and their application in an assay to evaluate neutralizing antibody activity. J Gen Virol. 1991;72(Pt 3):617–22. https://doi.org/10.1099/0022-1317-72-3-617.
Ravi M, Wobeser GA, Taylor SM, Jackson ML. Naturally acquired feline immunodeficiency virus (FIV) infection in cats from western Canada: prevalence, disease associations, and survival analysis. Can Vet J. 2010;51(3):271–6.
Liem BP, Dhand NK, Pepper AE, Barrs VR, Beatty JA. Clinical findings and survival in cats naturally infected with feline immunodeficiency virus. J Vet Intern Med. 2013;27(4):798–805. https://doi.org/10.1111/jvim.12120.
Castro KG, et al. 1993 revised classification system for HIV infection and expanded surveillance case definition for AIDS among adolescents and adults. Morbid Mortal Week Rep. 1992; 41.
Gleich S, Hartmann K. Hematology and serum biochemistry of feline immunodeficiency virus-infected and feline leukemia virus-infected cats. J Vet Intern Med. 2009;23(3):552–8. https://doi.org/10.1111/j.1939-1676.2009.0303.x.
Ishida T, Washizu T, Toriyabe K, Motoyoshi S, Tomoda I, Pedersen NC. Feline immunodeficiency virus infection in cats of Japan. J Am Vet Med Assoc. 1989;194(2):221–5.
Shelton GH, Waltier RM, Connor SC, Grant CK. Prevalence of feline immunodeficiency virus and feline leukemia virus infections on pet cats. J Am Anim Hosp Assoc. 1989;25:7–12.
Hutson CA, Rideout BA, Pedersen NC. Neoplasia associated with feline immunodeficiency virus infection in cats of southern California. J Am Vet Med Assoc. 1991;199(10):1357–62.
Alexander R, Robinson WF, Mill JN, Sherry CR, Sherard E, Paterson AJ, Shaw SE, Clark WT, Hollingsworth T. Isolation of feline immunodeficiency virus from three cats with lymphoma. Aust Vet Pract. 1989;19:93–7.
Hopper CD, Sparkes AH, Gruffydd-Jones TJ, et al. Clinical and laboratory findings in cats infected with feline immunodeficiency virus. Vet Rec. 1989;125(13):341–6. https://doi.org/10.1136/vr.125.13.341.
Shelton GH, McKim KD, Cooley PL, Dice PF, Russell RG, Grant CK. Feline leukemia virus and feline immunodeficiency virus infections in a cat with lymphoma. J Am Vet Med Assoc. 1989;194(2):249–52.
Callanan JJ, McCandlish IA, O’Neil B, et al. Lymphosarcoma in experimentally induced feline immunodeficiency virus infection. Vet Rec. 1992;130(14):293–5. https://doi.org/10.1136/vr.130.14.293.
Poli A, Abramo F, Baldinotti F, Pistello M, Da Prato L, Bendinelli M. Malignant lymphoma associated with experimentally induced feline immunodeficiency virus infection. J Comp Pathol. 1994;110(4):319–28. https://doi.org/10.1016/s0021-9975(08)80309-x.
Terry A, Callanan JJ, Fulton R, Jarrett O, Neil JC. Molecular analysis of tumours from feline immunodeficiency virus (FIV)-infected cats: an indirect role for FIV? Int J Cancer. 1995;61(2):227–32. https://doi.org/10.1002/ijc.2910610215.
Callanan JJ, Jones BA, Irvine J, Willett BJ, McCandlish IA, Jarrett O. Histologic classification and immunophenotype of lymphosarcomas in cats with naturally and experimentally acquired feline immunodeficiency virus infections. Vet Pathol. 1996;33(3):264–72. https://doi.org/10.1177/030098589603300302.
Endo Y, Cho KW, Nishigaki K, et al. Molecular characteristics of malignant lymphomas in cats naturally infected with feline immunodeficiency virus. Vet Immunol Immunopathol. 1997;57(3–4):153–67. https://doi.org/10.1016/s0165-2427(97)00004-4.
Gabor LJ, Love DN, Malik R, Canfield PJ. Feline immunodeficiency virus status of Australian cats with lymphosarcoma. Aust Vet J. 2001;79(8):540–5. https://doi.org/10.1111/j.1751-0813.2001.tb10742.x.
Wang J, Kyaw-Tanner M, Lee C, Robinson WF. Characterisation of lymphosarcomas in Australian cats using polymerase chain reaction and immunohistochemical examination. Aust Vet J. 2001;79(1):41–6. https://doi.org/10.1111/j.1751-0813.2001.tb10639.x.
Callanan JJ, Racz P, Thompson H, Jarrett O. Morphological characterization of the lymph node changes in feline immunodeficiency virus infection as an animal model of AIDS. In: Racz P, Letvin NL, Gluckmasn JC, editors. Animal models of HIV and other retroviral infections. Basel, Switzerland: Karger Publishing; 1993. p. 115–36.
Magden E, Miller C, MacMillan M, et al. Acute virulent infection with feline immunodeficiency virus (FIV) results in lymphomagenesis via an indirect mechanism. Virology. 2013;436(2):284–94. https://doi.org/10.1016/j.virol.2012.12.003.
Murphy BG, Eckstrand C, Castillo D, et al. Multiple, independent T Cell lymphomas arising in an experimentally FIV-infected cat during the terminal stage of infection. Viruses. 2018;10(6):280. https://doi.org/10.3390/v10060280.
Court EA, Watson AD, Peaston AE. Retrospective study of 60 cases of feline lymphosarcoma. Aust Vet J. 1997;75(6):424–7. https://doi.org/10.1111/j.1751-0813.1997.tb14347.x.
Beatty JA, Callanan JJ, Terry A, Jarrett O, Neil JC. Molecular and immunophenotypical characterization of a feline immunodeficiency virus (FIV)-associated lymphoma: a direct role for FIV in B-lymphocyte transformation? J Virol. 1998;72(1):767–71. https://doi.org/10.1128/JVI.72.1.767-771.1998.
Beatty J, Terry A, MacDonald J, et al. Feline immunodeficiency virus integration in B-cell lymphoma identifies a candidate tumor suppressor gene on human chromosome 15q15. Cancer Res. 2002;62(24):7175–80.
Fakhry C, Gillison ML. Clinical implications of human papillomavirus in head and neck cancers. J Clin Oncol. 2006;24(17):2606–11. https://doi.org/10.1200/JCO.2006.06.1291.
zur HH. Human papillomaviruses in the pathogenesis of anogenital cancer. Virology. 1991;184(1):9–13. https://doi.org/10.1016/0042-6822(91)90816-t.
Chaturvedi AK, Engels EA, Pfeiffer RM, et al. Human papillomavirus and rising oropharyngeal cancer incidence in the United States. J Clin Oncol. 2011;29(32):4294–301. https://doi.org/10.1200/JCO.2011.36.4596.
Estêvão D, Costa NR, Gil da Costa RM, Medeiros R. Hallmarks of HPV carcinogenesis: The role of E6, E7 and E5 oncoproteins in cellular malignancy. Biochim Biophys Acta Gene Regul Mech. 2019;1862(2):153–162. doi:https://doi.org/10.1016/j.bbagrm.2019.01.001
Munday JS, Knight CG, Luff JA. Papillomaviral skin diseases of humans, dogs, cats and horses: aA comparative review. Part 1: papillomavirus biology and hyperplastic lesions. Vet J. 2022;288:105897. https://doi.org/10.1016/j.tvjl.2022.105897.
Munday JS, French A, Thomson N. Detection of DNA sequences from a novel papillomavirus in a feline basal cell carcinoma. Vet Dermatol. 2017;28(2):236-e60. https://doi.org/10.1111/vde.12402.
Ciuffo G. Imnesto positivo con filtrato di verruca volgare. Giorn Ital Mal Venereol. 1907;48:12–7.
Rous P, Beard JW. The progression to carcinoma of virus-induced rabbit papillomas (shope). J Exp Med. 1935;62(4):523–48. https://doi.org/10.1084/jem.62.4.523.
zur HH. Papillomaviruses in the causation of human cancers - a brief historical account. Virology. 2009;384(2):260–5. https://doi.org/10.1016/j.virol.2008.11.046.
Carney HC, England JJ, Hodgin EC, Whiteley HE, Adkison DL, Sundberg JP. Papillomavirus infection of aged Persian cats. J Vet Diagn Invest. 1990;2(4):294–9. https://doi.org/10.1177/104063879000200407.
Delius H, Van Ranst MA, Jenson AB, zur Hausen H, Sundberg JP. Canine oral papillomavirus genomic sequence: a unique 1.5-kb intervening sequence between the E2 and L2 open reading frames. Virology. 1994;204(1):447–52. https://doi.org/10.1006/viro.1994.1552.
Tachezy R, Duson G, Rector A, Jenson AB, Sundberg JP, Van Ranst M. Cloning and genomic characterization of Felis domesticus papillomavirus type 1. Virology. 2002;301(2):313–21. https://doi.org/10.1006/viro.2002.1566.
Terai M, Burk RD. Felis domesticus papillomavirus, isolated from a skin lesion, is related to canine oral papillomavirus and contains a 1.3 kb non-coding region between the E2 and L2 open reading frames. J Gen Virol. 2002;83(Pt 9):2303–7. https://doi.org/10.1099/0022-1317-83-9-2303.
Munday JS, Pasavento P. Papillomaviridae and polyomaviridae. In: MacLachlan NJ, Dubovi EJ, editors. Fenner’s veterinary virology. 5th ed. London: Academic Press; 2017. p. 229–43.
Rector A, Van Ranst M. Animal papillomaviruses. Virology. 2013;445(1–2):213–23. https://doi.org/10.1016/j.virol.2013.05.007.
Munday JS, Thomson NA. Papillomaviruses in domestic cats. Viruses. 2021;13(8):1664. https://doi.org/10.3390/v13081664.
Bernard HU, Burk RD, Chen Z, van Doorslaer K, zur Hausen H, de Villiers EM. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology. 2010;401(1):70–9. https://doi.org/10.1016/j.virol.2010.02.002.
Woodham AW, Da Silva DM, Skeate JG, et al. The S100A10 subunit of the annexin A2 heterotetramer facilitates L2-mediated human papillomavirus infection. PLoS ONE. 2012;7(8):e43519. https://doi.org/10.1371/journal.pone.0043519.
Aksoy P, Gottschalk EY, Meneses PI. HPV entry into cells. Mutat Res Rev Mutat Res. 2017;772:13–22. https://doi.org/10.1016/j.mrrev.2016.09.004.
Bergant Marušič M, Ozbun MA, Campos SK, Myers MP, Banks L. Human papillomavirus L2 facilitates viral escape from late endosomes via sorting nexin 17. Traffic. 2012;13(3):455–67. https://doi.org/10.1111/j.1600-0854.2011.01320.x.
Ribeiro AL, Caodaglio AS, Sichero L. Regulation of HPV transcription. Clinics. 2018;73(suppl 1):e486s. https://doi.org/10.6061/clinics/2018/e486s.
Thomas M, Pim D, Banks L. The role of the E6–p53 interaction in the molecular pathogenesis of HPV. Oncogene. 1999;18(53):7690–700. https://doi.org/10.1038/sj.onc.1202953.
Zhang B, Chen W, Roman A. The E7 proteins of low- and high-risk human papillomaviruses share the ability to target the pRB family member p130 for degradation. Proc Natl Acad Sci U S A. 2006;103(2):437–42. https://doi.org/10.1073/pnas.0510012103.
Schiller JT, Day PM, Kines RC. Current understanding of the mechanism of HPV infection. Gynecol Oncol. 2010;118(1 Suppl):S12–7. https://doi.org/10.1016/j.ygyno.2010.04.004.
Klumpp DJ, Laimins LA. Differentiation-induced changes in promoter usage for transcripts encoding the human papillomavirus type 31 replication protein E1. Virology. 1999;257(1):239–46. https://doi.org/10.1006/viro.1999.9636.
McBride AA. Human papillomaviruses: diversity, infection and host interactions. Nat Rev Microbiol. 2022;20(2):95–108. https://doi.org/10.1038/s41579-021-00617-5.
Doorbar J, Quint W, Banks L, et al. The biology and life-cycle of human papillomaviruses. Vaccine. 2012;30(Suppl 5):F55–70. https://doi.org/10.1016/j.vaccine.2012.06.083.
Graham SV. The human papillomavirus replication cycle, and its links to cancer progression: a comprehensive review. Clin Sci. 2017;131(17):2201–21. https://doi.org/10.1042/CS20160786.
Cruz-Gregorio A, Aranda-Rivera AK, Pedraza-Chaverri J. Pathological similarities in the development of papillomavirus-associated cancer in humans, dogs, and cats. Animals. 2022;12(18):2390. https://doi.org/10.3390/ani12182390.
Antonsson A, Forslund O, Ekberg H, Sterner G, Hansson BG. The ubiquity and impressive genomic diversity of human skin papillomaviruses suggest a commensalic nature of these viruses. J Virol. 2000;74(24):11636–41. https://doi.org/10.1128/jvi.74.24.11636-11641.2000.
Antonsson A, Erfurt C, Hazard K, et al. Prevalence and type spectrum of human papillomaviruses in healthy skin samples collected in three continents. J Gen Virol. 2003;84(Pt 7):1881–6. https://doi.org/10.1099/vir.0.18836-0.
Doorbar J. The papillomavirus life cycle. J Clin Virol. 2005;32(Suppl 1):S7–15. https://doi.org/10.1016/j.jcv.2004.12.006.
Munday JS, Witham AI. Frequent detection of papillomavirus DNA in clinically normal skin of cats infected and noninfected with feline immunodeficiency virus. Vet Dermatol. 2010;21(3):307–10. https://doi.org/10.1111/j.1365-3164.2009.00811.x.
Lange CE, Zollinger S, Tobler K, Ackermann M, Favrot C. Clinically healthy skin of dogs is a potential reservoir for canine papillomaviruses. J Clin Microbiol. 2011;49(2):707–9. https://doi.org/10.1128/JCM.02047-10.
Thomson NA, Dunowska M, Munday JS. The use of quantitative PCR to detect Felis catus papillomavirus type 2 DNA from a high proportion of queens and their kittens. Vet Microbiol. 2015;175(2–4):211–7. https://doi.org/10.1016/j.vetmic.2014.11.028.
Oğuzoğlu TÇ, Timurkan MÖ, Koç BT, Alkan F. Comparison of genetic characteristics of canine papillomaviruses in Turkey. Infect Genet Evol. 2017;55:372–6. https://doi.org/10.1016/j.meegid.2017.10.010.
Sundberg JP, Smith EK, Herron AJ, Jenson AB, Burk RD, Van Ranst M. Involvement of canine oral papillomavirus in generalized oral and cutaneous verrucosis in a Chinese Shar Pei dog. Vet Pathol. 1994;31(2):183–7. https://doi.org/10.1177/030098589403100204.
Lange CE, Tobler K, Ackermann M, Panakova L, Thoday KL, Favrot C. Three novel canine papillomaviruses support taxonomic clade formation. J Gen Virol. 2009;90(Pt 11):2615–21. https://doi.org/10.1099/vir.0.014498-0.
Lange CE, Jennings SH, Diallo A, Lyons J. Canine papillomavirus types 1 and 2 in classical papillomas: high abundance, different morphological associations and frequent co-infections. Vet J. 2019;250:1–5. https://doi.org/10.1016/j.tvjl.2019.05.016.
Chang CY, Chen WT, Haga T, et al. The detection and association of canine papillomavirus with benign and malignant skin lesions in dogs. Viruses. 2020;12(2):170. https://doi.org/10.3390/v12020170.
Orlandi M, Mazzei M, Vascellari M, et al. Localization and genotyping of canine papillomavirus in canine inverted papillomas. J Vet Diagn Invest. 2021;33(6):1069–78. https://doi.org/10.1177/10406387211035799.
Gross TL, Ihrke PJ, Walder EJ, Affolter VK. Skin diseases of the dog and cat: clinical and histopathologic diagnosis. 2nd ed. Oxford: Blackwell Science; 2005.
Munday JS, French AF, MacNamara AR. The development of multiple cutaneous inverted papilloma following ovariohysterectomy in a dog. N Z Vet J. 2010;58(3):168–71. https://doi.org/10.1080/00480169.2010.67519.
Gould AP, Coyner KS, Trimmer AM, Tater K, Rishniw M. Canine pedal papilloma identification and management: a retrospective series of 44 cases. Vet Dermatol. 2021;32(5):509-e141. https://doi.org/10.1111/vde.12999.
Thaiwong T, Sledge DG, Wise AG, Olstad K, Maes RK, Kiupel M. Malignant transformation of canine oral papillomavirus (CPV1)-associated papillomas in dogs: an emerging concern? Papillomavirus Res. 2018;6:83–9. https://doi.org/10.1016/j.pvr.2018.10.007.
Nicholls PK, Klaunberg BA, Moore RA, et al. Naturally occurring, nonregressing canine oral papillomavirus infection: host immunity, virus characterization, and experimental infection. Virology. 1999;265(2):365–74. https://doi.org/10.1006/viro.1999.0060.
Regalado Ibarra AM, Legendre L, Munday JS. Malignant transformation of a canine papillomavirus type 1-induced persistent oral papilloma in a 3-year-old dog. J Vet Dent. 2018;35(2):79–95. https://doi.org/10.1177/0898756418774575.
Richman AW, Kirby AL, Rosenkrantz W, Muse R. Persistent papilloma treated with cryotherapy in three dogs. Vet Dermatol. 2017;28(6):625-e154. https://doi.org/10.1111/vde.12469.
Carpenter JL, Kreider JW, Alroy J, Schmidt GM. Cutaneous xanthogranuloma and viral papilloma on an eyelid of a cat. Vet Dermatol. 1992;3:187–90.
Munday JS, Hanlon EM, Howe L, Squires RA, French AF. Feline cutaneous viral papilloma associated with human papillomavirus type 9. Vet Pathol. 2007;44(6):924–7. https://doi.org/10.1354/vp.44-6-924.
Munday JS, Thomson NA, Luff JA. Papillomaviruses in dogs and cats. Vet J. 2017;225:23–31. https://doi.org/10.1016/j.tvjl.2017.04.018.
Munday JS, Lam ATH, Sakai M. Extensive progressive pigmented viral plaques in a Chihuahua dog. Vet Dermatol. 2022;33(3):252–4. https://doi.org/10.1111/vde.13056--B.
Munday JS, Knight CG, Luff JA. Papillomaviral skin diseases of humans, dogs, cats and horses: a comparative review. Part 2: pre-neoplastic and neoplastic diseases. Vet J. 2022;288:105898. https://doi.org/10.1016/j.tvjl.2022.105898.
Tobler K, Lange C, Carlotti DN, Ackermann M, Favrot C. Detection of a novel papillomavirus in pigmented plaques of four pugs. Vet Dermatol. 2008;19(1):21–5. https://doi.org/10.1111/j.1365-3164.2007.00640.x.
Lange CE, Tobler K, Lehner A, Vetsch E, Favrot C. A case of a canine pigmented plaque associated with the presence of a Chi-papillomavirus. Vet Dermatol. 2012;23(1):76-e19. https://doi.org/10.1111/j.1365-3164.2011.01007.x.
Yuan H, Luff J, Zhou D, et al. Complete genome sequence of canine papillomavirus type 9. J Virol. 2012;86(10):5966. https://doi.org/10.1128/JVI.00543-12.
Luff J, Mader M, Britton M, et al. Complete genome sequence of canine papillomavirus type 16. Genome Announc. 2015;3(3):e00404-15. https://doi.org/10.1128/genomeA.00404-15.
Nespeca G, Grest P, Rosenkrantz WS, Ackermann M, Favrot C. Detection of novel papillomaviruslike sequences in paraffin-embedded specimens of invasive and in situ squamous cell carcinomas from cats. Am J Vet Res. 2006;67(12):2036–41. https://doi.org/10.2460/ajvr.67.12.2036.
Lange CE, Tobler K, Markau T, et al. Sequence and classification of FdPV2, a papillomavirus isolated from feline Bowenoid in situ carcinomas. Vet Microbiol. 2009;137(1–2):60–5. https://doi.org/10.1016/j.vetmic.2009.01.002.
Demos LE, Munday JS, Lange CE, Bennett MD. Use of fluorescence in situ hybridization to detect Felis catus papillomavirus type 2 in feline Bowenoid in situ carcinomas. J Feline Med Surg. 2019;21(6):575–80. https://doi.org/10.1177/1098612X18795919.
Kok MK, Yamashita-Kawanishi N, Chambers JK, et al. Pathologic characterization of Felis catus papillomavirus type 5 (FcaPV-5)-associated viral plaques and Bowenoid in situ carcinoma in a Domestic Shorthair cat. J Vet Med Sci. 2019;81(5):660–6. https://doi.org/10.1292/jvms.18-0771.
Vascellari M, Mazzei M, Zanardello C, et al. Felis catus papillomavirus types 1, 2, 3, 4, and 5 in feline Bowenoid in situ carcinoma: an in situ hybridization study. Vet Pathol. 2019;56(6):818–25. https://doi.org/10.1177/0300985819859874.
Nagata M, Nanko H, Moriyama A, Washizu T, Ishida T. Pigmented plaques associated with papillomavirus infection in dogs: is this epidermodysplasia verruciformis? Vet Dermatol. 1995;6(4):179–86. https://doi.org/10.1111/j.1365-3164.1995.tb00063.x.
Hansen N, Nicholas N, Pack G, et al. Progressive cutaneous viral pigmented plaques in three Hungarian Vizslas and the response of lesions to topical tigilanol tiglate gel. Vet Med Sci. 2017;4(1):53–62. https://doi.org/10.1002/vms3.85.
Ravens PA, Vogelnest LJ, Tong LJ, Demos LE, Bennett MD. Papillomavirus-associated multicentric squamous cell carcinoma in situ in a cat: an unusually extensive and progressive case with subsequent metastasis. Vet Dermatol. 2013;24(6):642-e162. https://doi.org/10.1111/vde.12080.
Munday JS, Benfell MW, French A, Orbell GM, Thomson N. Bowenoid in situ carcinomas in two Devon Rex cats: evidence of unusually aggressive neoplasm behaviour in this breed and detection of papillomaviral gene expression in primary and metastatic lesions. Vet Dermatol. 2016;27(3):215-e55. https://doi.org/10.1111/vde.12319.
Knight EC, Munday JS, Stone BM, Shipstone MA. Carbon dioxide laser treatment of extensive pigmented viral plaque lesions in a golden retriever dog. Vet Dermatol. 2016;27(5):442-e117. https://doi.org/10.1111/vde.12350.
Munday JS, O’Connor KI, Smits B. Development of multiple pigmented viral plaques and squamous cell carcinomas in a dog infected by a novel papillomavirus. Vet Dermatol. 2011;22(1):104–10. https://doi.org/10.1111/j.1365-3164.2010.00913.x.
Luff J, Mader M, Rowland P, Britton M, Fass J, Yuan H. Viral genome integration of canine papillomavirus 16. Papillomavirus Res. 2019;7:88–96. https://doi.org/10.1016/j.pvr.2019.02.002.
Alves CDBT, Weber MN, Guimarães LLB, et al. Canine papillomavirus type 16 associated to squamous cell carcinoma in a dog: virological and pathological findings. Braz J Microbiol. 2020;51(4):2087–94. https://doi.org/10.1007/s42770-020-00310-4.
Gill VL, Bergman PJ, Baer KE, Craft D, Leung C. Use of imiquimod 5% cream (Aldara) in cats with multicentric squamous cell carcinoma in situ: 12 cases (2002–2005). Vet Comp Oncol. 2008;6(1):55–64. https://doi.org/10.1111/j.1476-5829.2007.00144.x.
Teifke JP, Löhr CV, Shirasawa H. Detection of canine oral papillomavirus-DNA in canine oral squamous cell carcinomas and p53 overexpressing skin papillomas of the dog using the polymerase chain reaction and non-radioactive in situ hybridization. Vet Microbiol. 1998;60(2–4):119–30. https://doi.org/10.1016/s0378-1135(98)00151-5.
Zaugg N, Nespeca G, Hauser B, Ackermann M, Favrot C. Detection of novel papillomaviruses in canine mucosal, cutaneous and in situ squamous cell carcinomas. Vet Dermatol. 2005;16(5):290–8. https://doi.org/10.1111/j.1365-3164.2005.00467.x.
Waropastrakul S, Munday JS, French AF. Infrequent detection of papillomaviral DNA within canine cutaneous squamous cell carcinomas, haemangiosarcomas and healthy skin on the ventrum of dogs. Vet Dermatol. 2012;23(3):197-e41. https://doi.org/10.1111/j.1365-3164.2012.01043.x.
Ito S, Chambers JK, Sumi A, et al. Involvement of Felis catus papillomavirus type 2 in the tumorigenesis of feline Merkel cell carcinoma. Vet Pathol. 2022;59(1):63–74. https://doi.org/10.1177/03009858211045440.
Munday JS, Gibson I, French AF. Papillomaviral DNA and increased p16CDKN2A protein are frequently present within feline cutaneous squamous cell carcinomas in ultraviolet-protected skin. Vet Dermatol. 2011;22(4):360–6. https://doi.org/10.1111/j.1365-3164.2011.00958.x.
O’Neill SH, Newkirk KM, Anis EA, Brahmbhatt R, Frank LA, Kania SA. Detection of human papillomavirus DNA in feline premalignant and invasive squamous cell carcinoma. Vet Dermatol. 2011;22(1):68–74. https://doi.org/10.1111/j.1365-3164.2010.00912.x.
Munday JS, Aberdein D. Loss of retinoblastoma protein, but not p53, is associated with the presence of papillomaviral DNA in feline viral plaques, Bowenoid in situ carcinomas, and squamous cell carcinomas. Vet Pathol. 2012;49(3):538–45. https://doi.org/10.1177/0300985811419534.
Munday JS, French AF, Gibson IR, Knight CG. The presence of p16 CDKN2A protein immunostaining within feline nasal planum squamous cell carcinomas is associated with an increased survival time and the presence of papillomaviral DNA. Vet Pathol. 2013;50(2):269–73. https://doi.org/10.1177/0300985812452582.
Thomson NA, Munday JS, Dittmer KE. Frequent detection of transcriptionally active Felis catus papillomavirus 2 in feline cutaneous squamous cell carcinomas. J Gen Virol. 2016;97(5):1189–97. https://doi.org/10.1099/jgv.0.000416.
Altamura G, Power K, Martano M, et al. Felis catus papillomavirus type-2 E6 binds to E6AP, promotes E6AP/p53 binding and enhances p53 proteasomal degradation. Sci Rep. 2018;8(1):17529. https://doi.org/10.1038/s41598-018-35723-7.
Yamashita-Kawanishi N, Chang CY, Chambers JK, et al. Comparison of prevalence of Felis catus papillomavirus type 2 in squamous cell carcinomas in cats between Taiwan and Japan. J Vet Med Sci. 2021;83(8):1229–33. https://doi.org/10.1292/jvms.21-0153.
Altamura G, Corteggio A, Pacini L, et al. Transforming properties of Felis catus papillomavirus type 2 E6 and E7 putative oncogenes in vitro and their transcriptional activity in feline squamous cell carcinoma in vivo. Virology. 2016;496:1–8. https://doi.org/10.1016/j.virol.2016.05.017.
Thomson NA, Thomas DG, Weidgraaf K, Munday JS. Felis catus papillomavirus type 2 DNA loads on kittens are transient and do not reflect their susceptibility to infection. J Feline Med Surg. 2018;20(4):332–8. https://doi.org/10.1177/1098612X17710382.
Munday JS, Thomson N, Dunowska M, Knight CG, Laurie RE, Hills S. Genomic characterisation of the feline sarcoid-associated papillomavirus and proposed classification as Bos taurus papillomavirus type 14. Vet Microbiol. 2015;177(3–4):289–95. https://doi.org/10.1016/j.vetmic.2015.03.019.
Greenwood S, Campbell O, Movasseghi AR. Oral sarcoid in a cat. Can Vet J. 2019;60(5):485–9.
Schulman FY, Krafft AE, Janczewski T. Feline cutaneous fibropapillomas: clinicopathologic findings and association with papillomavirus infection. Vet Pathol. 2001;38(3):291–6. https://doi.org/10.1354/vp.38-3-291.
Georgescu SR, Mitran CI, Mitran MI, et al. New insights in the pathogenesis of HPV infection and the associated carcinogenic processes: the role of chronic inflammation and oxidative stress. J Immunol Res. 2018;2018:5315816. https://doi.org/10.1155/2018/5315816.
Oyervides-Muñoz MA, Pérez-Maya AA, Rodríguez-Gutiérrez HF, et al. Understanding the HPV integration and its progression to cervical cancer. Infect Genet Evol. 2018;61:134–44. https://doi.org/10.1016/j.meegid.2018.03.003.
Roden RBS, Stern PL. Opportunities and challenges for human papillomavirus vaccination in cancer. Nat Rev Cancer. 2018;18(4):240–54. https://doi.org/10.1038/nrc.2018.13.
Yuan H, Ghim S, Newsome J, et al. An epidermotropic canine papillomavirus with malignant potential contains an E5 gene and establishes a unique genus. Virology. 2007;359(1):28–36. https://doi.org/10.1016/j.virol.2006.08.029.
Boyer SN, Wazer DE, Band V. E7 protein of human papilloma virus-16 induces degradation of retinoblastoma protein through the ubiquitin-proteasome pathway. Cancer Res. 1996;56(20):4620–4.
Di Domenico F, Foppoli C, Blarzino C, et al. Expression of human papilloma virus type 16 E5 protein in amelanotic melanoma cells regulates endo-cellular pH and restores tyrosinase activity. J Exp Clin Cancer Res. 2009;28(1):4. https://doi.org/10.1186/1756-9966-28-4.
Oh JM, Kim SH, Cho EA, Song YS, Kim WH, Juhnn YS. Human papillomavirus type 16 E5 protein inhibits hydrogen-peroxide-induced apoptosis by stimulating ubiquitin-proteasome-mediated degradation of Bax in human cervical cancer cells. Carcinogenesis. 2010;31(3):402–10. https://doi.org/10.1093/carcin/bgp318.
Borzacchiello G, Mogavero S, De Vita G, Roperto S, Della Salda L, Roperto F. Activated platelet-derived growth factor beta receptor expression, PI3K-AKT pathway molecular analysis, and transforming signals in equine sarcoids. Vet Pathol. 2009;46(4):589–97. https://doi.org/10.1354/vp.08-VP-0191-B-FL.
Munday JS. Bovine and human papillomaviruses: a comparative review. Vet Pathol. 2014;51(6):1063–75. https://doi.org/10.1177/0300985814537837.
Araldi RP, Assaf SMR, Carvalho RF, et al. Papillomaviruses: a systematic review. Genet Mol Biol. 2017;40(1):1–21. https://doi.org/10.1590/1678-4685-GMB-2016-0128.
Altamura G, Borzacchiello G. HPV related head and neck squamous cell carcinoma: new evidences for an emerging spontaneous animal model. Oral Oncol. 2019;88:84. https://doi.org/10.1016/j.oraloncology.2018.11.027.
Altamura G, Borzacchiello G. Feline oral squamous cell carcinoma and Felis catus papillomavirus: is it time to walk the path of human oncology? Front Vet Sci. 2023;10:1148673. https://doi.org/10.3389/fvets.2023.1148673.
Kamolratanakul S, Pitisuttithum P. Human papillomavirus vaccine efficacy and effectiveness against cancer. Vaccines. 2021;9(12):1413. https://doi.org/10.3390/vaccines9121413.PMID:34960159;PMCID:PMC8706722.
Centers for Disease Control. 1993 revised classification system for HIV infection and expanded surveillance case definition for AIDS among adolescents and adults. MMWR Recomm Rep. 1992;41:1–19.
Kaye S, Wang W, Miller C, et al. Role of feline immunodeficiency virus in lymphomagenesis-going alone or colluding? ILAR J. 2016;57(1):24–33. https://doi.org/10.1093/ilar/ilv047.
Cesarman E. Gammaherpesvirus and lymphoproliferative disorders in immunocompromised patients. Cancer Lett. 2011;305(2):163–74. https://doi.org/10.1016/j.canlet.2011.03.003.
Rahman R, Gopinath D, Buajeeb W, Poomsawat S, Johnson NW. Potential role of Epstein-Barr virus in oral potentially malignant disorders and oral squamous cell carcinoma: a scoping review. Viruses. 2022;14(4):801. https://doi.org/10.3390/v14040801.
Du MQ, Bacon CM, Isaacson PG. Kaposi sarcoma-associated herpesvirus/human herpesvirus 8 and lymphoproliferative disorders. J Clin Pathol. 2007;60(12):1350–7. https://doi.org/10.1136/jcp.2007.047969.
Bruce AG, Bielefeldt-Ohmann H, Barcy S, et al. Macaque homologs of EBV and KSHV show uniquely different associations with simian AIDS-related lymphomas. PLoS Pathog. 2012;8(10):e1002962. https://doi.org/10.1371/journal.ppat.1002962.
Carbone A, Cesarman E, Spina M, Gloghini A, Schulz TF. HIV-associated lymphomas and gamma-herpesviruses. Blood. 2009;113(6):1213–24. https://doi.org/10.1182/blood-2008-09-180315.
Rickinson AB, Moss DJ. Human cytotoxic T lymphocyte responses to Epstein-Barr virus infection. Annu Rev Immunol. 1997;15:405–31. https://doi.org/10.1146/annurev.immunol.15.1.405. (PMID: 9143694).
Callan MF, Tan L, Annels N, et al. Direct visualization of antigen-specific CD8+ T cells during the primary immune response to Epstein-Barr virus In vivo. J Exp Med. 1998;187(9):1395–402. https://doi.org/10.1084/jem.187.9.1395.
Baer R, Bankier AT, Biggin MD, et al. DNA sequence and expression of the B95–8 Epstein-Barr virus genome. Nature. 1984;310(5974):207–11. https://doi.org/10.1038/310207a0.
Grywalska E, Rolinski J. Epstein-Barr virus-associated lymphomas. Semin Oncol. 2015;42(2):291–303. https://doi.org/10.1053/j.seminoncol.2014.12.030.
Hsu JL, Glaser SL. Epstein-barr virus-associated malignancies: epidemiologic patterns and etiologic implications. Crit Rev Oncol Hematol. 2000;34(1):27–53. https://doi.org/10.1016/s1040-8428(00)00046-9.
Young LS, Dawson CW. Epstein-Barr virus and nasopharyngeal carcinoma. Chin J Cancer. 2014;33(12):581–90. https://doi.org/10.5732/cjc.014.10197.
Shinozaki-Ushiku A, Kunita A, Fukayama M. Update on Epstein-Barr virus and gastric cancer (review). Int J Oncol. 2015;46(4):1421–34. https://doi.org/10.3892/ijo.2015.2856.
Lemon SM, Hutt LM, Shaw JE, Li JL, Pagano JS. Replication of EBV in epithelial cells during infectious mononucleosis. Nature. 1977;268(5617):268–70. https://doi.org/10.1038/268268a0.
Sixbey JW, Nedrud JG, Raab-Traub N, Hanes RA, Pagano JS. Epstein-Barr virus replication in oropharyngeal epithelial cells. N Engl J Med. 1984;310(19):1225–30. https://doi.org/10.1056/NEJM198405103101905.
Labrecque LG, Barnes DM, Fentiman IS, Griffin BE. Epstein-Barr virus in epithelial cell tumors: a breast cancer study. Cancer Res. 1995;55(1):39–45.
Mazouni C, Fina F, Romain S, et al. Epstein-Barr virus as a marker of biological aggressiveness in breast cancer. Br J Cancer. 2011;104(2):332–7. https://doi.org/10.1038/sj.bjc.6606048.
Fina F, Romain S, Ouafik L, et al. Frequency and genome load of Epstein-Barr virus in 509 breast cancers from different geographical areas. Br J Cancer. 2001;84(6):783–90. https://doi.org/10.1054/bjoc.2000.1672.
Kijima Y, Hokita S, Takao S, et al. Epstein-Barr virus involvement is mainly restricted to lymphoepithelial type of gastric carcinoma among various epithelial neoplasms. J Med Virol. 2001;64(4):513–8. https://doi.org/10.1002/jmv.1079.
Kalkan A, Ozdarendeli A, Bulut Y, Yekeler H, Cobanoglu B, Doymaz MZ. Investigation of Epstein-Barr virus DNA in formalin-fixed and paraffin- embedded breast cancer tissues. Med Princ Pract. 2005;14(4):268–71. https://doi.org/10.1159/000085748.
Preciado MV, Chabay PA, De Matteo EN, et al. Epstein-Barr virus in breast carcinoma in Argentina. Arch Pathol Lab Med. 2005;129(3):377–81. https://doi.org/10.5858/2005-129-377-EVIBCI.
Joshi D, Quadri M, Gangane N, Joshi R, Gangane N. Association of Epstein Barr virus infection (EBV) with breast cancer in rural Indian women. PLoS ONE. 2009;4(12):e8180. https://doi.org/10.1371/journal.pone.0008180.
Lorenzetti MA, De Matteo E, Gass H, et al. Characterization of Epstein Barr virus latency pattern in argentine breast carcinoma. PLoS ONE. 2010;5(10):e13603. https://doi.org/10.1371/journal.pone.0013603.
Aguayo F, Khan N, Koriyama C, et al. Human papillomavirus and Epstein-Barr virus infections in breast cancer from chile. Infect Agent Cancer. 2011;6(1):7. https://doi.org/10.1186/1750-9378-6-7.
Zekri AR, Bahnassy AA, Mohamed WS, et al. Epstein-Barr virus and breast cancer: epidemiological and molecular study on Egyptian and Iraqi women. J Egypt Natl Canc Inst. 2012;24(3):123–31. https://doi.org/10.1016/j.jnci.2012.06.001.
Yahia ZA, Adam AA, Elgizouli M, et al. Epstein Barr virus: a prime candidate of breast cancer aetiology in Sudanese patients. Infect Agent Cancer. 2014;9(1):9. https://doi.org/10.1186/1750-9378-9-9.
Richardson AK, Currie MJ, Robinson BA, et al. Cytomegalovirus and Epstein-Barr virus in breast cancer. PLoS ONE. 2015;10(2):e0118989. https://doi.org/10.1371/journal.pone.0118989.
El-Naby NEH, Hassan Mohamed H, Mohamed Goda A, El Sayed MA. Epstein-Barr virus infection and breast invasive ductal carcinoma in Egyptian women: a single center experience. J Egypt Natl Canc Inst. 2017;29(2):77–82. https://doi.org/10.1016/j.jnci.2017.02.002.
Fessahaye G, Elhassan AM, Elamin EM, Adam AAM, Ghebremedhin A, Ibrahim ME. Association of Epstein - Barr virus and breast cancer in Eritrea. Infect Agent Cancer. 2017;12:62. https://doi.org/10.1186/s13027-017-0173-2.
Glaser SL, Ambinder RF, DiGiuseppe JA, Horn-Ross PL, Hsu JL. Absence of Epstein-Barr virus EBER-1 transcripts in an epidemiologically diverse group of breast cancers. Int J Cancer. 1998;75(4):555–8. https://doi.org/10.1002/(sici)1097-0215(19980209)75:4%3c555::aid-ijc10%3e3.0.co;2-8.
Pai T, Gupta S, Gurav M, et al. Evidence for the association of Epstein-Barr Virus in breast cancer in Indian patients using in-situ hybridization technique. Breast J. 2018;24(1):16–22. https://doi.org/10.1111/tbj.12828.
Sharifpour C, Makvandi M, Samarbafzadeh A, et al. Frequency of Epstein-Barr virus DNA in formalin-fixed paraffin-embedded tissue of patients with ductal breast carcinoma. Asian Pac J Cancer Prev. 2019;20(3):687–92. https://doi.org/10.31557/APJCP.2019.20.3.687.
Golrokh Mofrad M, Kazeminezhad B, Faghihloo E. Prevalence of Epstein-Barr virus (EBV) in Iranian breast carcinoma patients. Asian Pac J Cancer Prev. 2020;21(1):133–7. https://doi.org/10.31557/APJCP.2020.21.1.133.
Hippocrate A, Oussaief L, Joab I. Possible role of EBV in breast cancer and other unusually EBV-associated cancers. Cancer Lett. 2011;305(2):144–9. https://doi.org/10.1016/j.canlet.2010.11.007.
Gupta I, Ulamec M, Peric-Balja M, et al. Presence of high-risk HPVs, EBV, and MMTV in human triple-negative breast cancer. Hum Vaccin Immunother. 2021;17(11):4457–66. https://doi.org/10.1080/21645515.2021.1975452.
Deshpande CG, Badve S, Kidwai N, Longnecker R. Lack of expression of the Epstein-Barr Virus (EBV) gene products, EBERs, EBNA1, LMP1, and LMP2A, in breast cancer cells. Lab Invest. 2002;82(9):1193–9. https://doi.org/10.1097/01.lab.0000029150.90532.24.
Herrmann K, Niedobitek G. Lack of evidence for an association of Epstein-Barr virus infection with breast carcinoma. Breast Cancer Res. 2003;5(1):R13–7. https://doi.org/10.1186/bcr561.
Murray PG, Lissauer D, Junying J, et al. Reactivity with A monoclonal antibody to Epstein-Barr virus (EBV) nuclear antigen 1 defines a subset of aggressive breast cancers in the absence of the EBV genome. Cancer Res. 2003;63(9):2338–43.
Perrigoue JG, den Boon JA, Friedl A, Newton MA, Ahlquist P, Sugden B. Lack of association between EBV and breast carcinoma. Cancer Epidemiol Biomarkers Prev. 2005;14(4):809–14. https://doi.org/10.1158/1055-9965.EPI-04-0763.
Kadivar M, Monabati A, Joulaee A, Hosseini N. Epstein-Barr virus and breast cancer: lack of evidence for an association in Iranian women. Pathol Oncol Res. 2011;17(3):489–92. https://doi.org/10.1007/s12253-010-9325-z.
Dowran R, Joharinia N, Safaei A, et al. No detection of EBV, BKV and JCV in breast cancer tissue samples in Iran. BMC Res Notes. 2019;12(1):171. https://doi.org/10.1186/s13104-019-4178-3.
Arias-Calvachi C, Blanco R, Calaf GM, Aguayo F. Epstein-Barr Virus association with breast cancer: evidence and perspectives. Biology. 2022;11(6):799. https://doi.org/10.3390/biology11060799.
Young LS, Arrand JR, Murray PG, et al. EBV gene expression and regulation. In: Arvin A, et al., editors. Human herpesviruses: biology, therapy, and immunoprophylaxis. Cambridge University Press; 2007.
Küppers R. B cells under influence: transformation of B cells by Epstein-Barr virus. Nat Rev Immunol. 2003;3(10):801–12. https://doi.org/10.1038/nri1201.
Rowe M, Kelly GL, Bell AI, Rickinson AB. Burkitt’s lymphoma: the Rosetta Stone deciphering Epstein-Barr virus biology. Semin Cancer Biol. 2009;19(6):377–88. https://doi.org/10.1016/j.semcancer.2009.07.004.
Takacs M, Banati F, Koroknai A, et al. Epigenetic regulation of latent Epstein-Barr virus promoters. Biochim Biophys Acta. 2010;1799(3–4):228–35. https://doi.org/10.1016/j.bbagrm.2009.10.005.
Heuts F, Rottenberg ME, Salamon D, et al. T cells modulate Epstein-Barr virus latency phenotypes during infection of humanized mice. J Virol. 2014;88(6):3235–45. https://doi.org/10.1128/JVI.02885-13.
Murata T, Sato Y, Kimura H. Modes of infection and oncogenesis by the Epstein-Barr virus. Rev Med Virol. 2014;24(4):242–53. https://doi.org/10.1002/rmv.1786.
Portis T, Ikeda M, Longnecker R. Epstein-Barr virus LMP2A: regulating cellular ubiquitination processes for maintenance of viral latency? Trends Immunol. 2004;25(8):422–6. https://doi.org/10.1016/j.it.2004.05.009.
Soni V, Cahir-McFarland E, Kieff E. LMP1 TRAFficking activates growth and survival pathways. Adv Exp Med Biol. 2007;597:173–87. https://doi.org/10.1007/978-0-387-70630-6_14.
Maruo S, Zhao B, Johannsen E, Kieff E, Zou J, Takada K. Epstein-Barr virus nuclear antigens 3C and 3A maintain lymphoblastoid cell growth by repressing p16INK4A and p14ARF expression. Proc Natl Acad Sci U S A. 2011;108(5):1919–24. https://doi.org/10.1073/pnas.1019599108.
Marquitz AR, Raab-Traub N. The role of miRNAs and EBV BARTs in NPC. Semin Cancer Biol. 2012;22(2):166–72. https://doi.org/10.1016/j.semcancer.2011.12.001.
Takada K. Role of EBER and BARF1 in nasopharyngeal carcinoma (NPC) tumorigenesis. Semin Cancer Biol. 2012;22(2):162–5. https://doi.org/10.1016/j.semcancer.2011.12.007.
Lopes LF, Ruiz Miyazawa KW, de Almeida ER, et al. Epstein-Barr virus (EBV) microRNAs: involvement in cancer pathogenesis and immunopathology. Int Rev Immunol. 2013;32(3):271–81. https://doi.org/10.3109/08830185.2012.748053.
Skalska L, White RE, Parker GA, et al. Induction of p16(INK4a) is the major barrier to proliferation when Epstein-Barr virus (EBV) transforms primary B cells into lymphoblastoid cell lines. PLoS Pathog. 2013;9(2):e1003187. https://doi.org/10.1371/journal.ppat.1003187.
Vereide DT, Seto E, Chiu YF, et al. Epstein-Barr virus maintains lymphomas via its miRNAs. Oncogene. 2014;33(10):1258–64. https://doi.org/10.1038/onc.2013.71.
Levitskaya J, Coram M, Levitsky V, et al. Inhibition of antigen processing by the internal repeat region of the Epstein-Barr virus nuclear antigen-1. Nature. 1995;375(6533):685–8. https://doi.org/10.1038/375685a0.
Carbone A. AIDS-related non-Hodgkin’s lymphomas: from pathology and molecular pathogenesis to treatment. Hum Pathol. 2002;33(4):392–404. https://doi.org/10.1053/hupa.2002.124723.
Croft NP, Shannon-Lowe C, Bell AI, et al. Stage-specific inhibition of MHC class I presentation by the Epstein-Barr virus BNLF2a protein during virus lytic cycle. PLoS Pathog. 2009;5(6):e1000490. https://doi.org/10.1371/journal.ppat.1000490.
Gruhne B, Kamranvar SA, Masucci MG, Sompallae R. EBV and genomic instability–a new look at the role of the virus in the pathogenesis of Burkitt’s lymphoma. Semin Cancer Biol. 2009;19(6):394–400. https://doi.org/10.1016/j.semcancer.2009.07.005.
Uozaki H, Fukayama M. Epstein-Barr virus and gastric carcinoma–viral carcinogenesis through epigenetic mechanisms. Int J Clin Exp Pathol. 2008;1(3):198–216.
Chiou SH, Chow KC, Yang CH, Chiang SF, Lin CH. Discovery of Epstein-Barr virus (EBV)-encoded RNA signal and EBV nuclear antigen leader protein DNA sequence in pet dogs. J Gen Virol. 2005;86(Pt 4):899–905. https://doi.org/10.1099/vir.0.80792-0.
Milman G, Smith KC, Erles K. Serological detection of Epstein-Barr virus infection in dogs and cats. Vet Microbiol. 2011;150(1–2):15–20. https://doi.org/10.1016/j.vetmic.2010.12.013.
Huang SH, Kozak PJ, Kim J, et al. Evidence of an oncogenic gammaherpesvirus in domestic dogs. Virology. 2012;427(2):107–17. https://doi.org/10.1016/j.virol.2012.02.013.
Macsween KF, Crawford DH. Epstein-Barr virus-recent advances. Lancet Infect Dis. 2003;3(3):131–40. https://doi.org/10.1016/s1473-3099(03)00543-7.
Chiu HC, Chow KC, Fan YH, et al. Expression of EBV-encoded oncogenes and EBV-like virions in multiple canine tumors. Vet Microbiol. 2013;163(1–2):79–89. https://doi.org/10.1016/j.vetmic.2012.12.022.
Mollbrink A, Falk KI, Linde A, Barkholt L. Monitoring EBV DNA in saliva for early diagnosis of EBV reactivation in solid tumour patients after allogeneic haematopoietic SCT. Bone Marrow Transplant. 2009;44(4):259–61. https://doi.org/10.1038/bmt.2009.5.
Vazirabadi G, Geiger TR, Coffin Iii WF, Martin JM. Epstein-Barr virus latent membrane protein-1 (LMP-1) and lytic LMP-1 localization in plasma membrane-derived extracellular vesicles and intracellular virions. J Gen Virol. 2003;84(Pt 8):1997–2008. https://doi.org/10.1099/vir.0.19156-0.
Wang WH, Chang LK, Liu ST. Molecular interactions of Epstein-Barr virus capsid proteins. J Virol. 2011;85(4):1615–24. https://doi.org/10.1128/JVI.01565-10.
Waugh EM, Gallagher A, McAulay KA, et al. Gammaherpesviruses and canine lymphoma: no evidence for direct involvement in commonly occurring lymphomas. J Gen Virol. 2015;96(Pt 7):1863–72. https://doi.org/10.1099/vir.0.000106.
Lawson JS, Salmons B, Glenn WK. Oncogenic viruses and breast cancer: mouse mammary tumor virus (MMTV), Bovine leukemia virus (BLV), human papilloma virus (HPV), and Epstein-Barr virus (EBV). Front Oncol. 2018;8:1. https://doi.org/10.3389/fonc.2018.00001.
Roa López GA, Suárez JJ, Barato P, García NV. Lack of association between Epstein-Barr virus and mammary tumours in dogs. J Vet Res. 2018;62(3):309–15. https://doi.org/10.2478/jvetres-2018-0045.
Troyer RM, Beatty JA, Stutzman-Rodriguez KR, et al. Novel gammaherpesviruses in North American domestic cats, bobcats, and pumas: identification, prevalence, and risk factors. J Virol. 2014;88(8):3914–24. https://doi.org/10.1128/JVI.03405-13.
Beatty JA, Troyer RM, Carver S, et al. Felis catus gammaherpesvirus 1; a widely endemic potential pathogen of domestic cats. Virology. 2014;460–461:100–7. https://doi.org/10.1016/j.virol.2014.05.007.
Ertl R, Korb M, Langbein-Detsch I, Klein D. Prevalence and risk factors of gammaherpesvirus infection in domestic cats in Central Europe. Virol J. 2015;12:146. https://doi.org/10.1186/s12985-015-0381-6.
McLuckie A, Tasker S, Dhand NK, Spencer S, Beatty JA. High prevalence of Felis catus gammaherpesvirus 1 infection in haemoplasma-infected cats supports co-transmission. Vet J. 2016;214:117–21. https://doi.org/10.1016/j.tvjl.2016.06.001.
Aghazadeh M, Shi M, Pesavento PA, et al. Transcriptome analysis and in situ hybridization for FcaGHV1 in feline lymphoma. Viruses. 2018;10(9):464. https://doi.org/10.3390/v10090464.
Guidotti LG, Isogawa M, Chisari FV. Host-virus interactions in hepatitis B virus infection. Curr Opin Immunol. 2015;36:61–6. https://doi.org/10.1016/j.coi.2015.06.016.
Aghazadeh M, Shi M, Barrs VR, et al. A novel hepadnavirus identified in an immunocompromised domestic cat in Australia. Viruses. 2018;10(5):269. https://doi.org/10.3390/v10050269.
Magnius L, Mason WS, Taylor J, et al. ICTV virus taxonomy profile: hepadnaviridae. J Gen Virol. 2020;101(6):571–2. https://doi.org/10.1099/jgv.0.001415.
Ko K, Takahashi K, Nagashima S, et al. Existence of hepatitis B virus surface protein mutations and other variants: demand for hepatitis B infection control in Cambodia. BMC Infect Dis. 2020;20(1):305. https://doi.org/10.1186/s12879-020-05025-3.
Piewbang C, Wardhani SW, Chaiyasak S, et al. Insights into the genetic diversity, recombination, and systemic infections with evidence of intracellular maturation of hepadnavirus in cats. PLoS ONE. 2020;15(10):e0241212. https://doi.org/10.1371/journal.pone.0241212.
Stella L, Santopaolo F, Gasbarrini A, Pompili M, Ponziani FR. Viral hepatitis and hepatocellular carcinoma: from molecular pathways to the role of clinical surveillance and antiviral treatment. World J Gastroenterol. 2022;28(21):2251–81. https://doi.org/10.3748/wjg.v28.i21.2251.
Lanave G, Capozza P, Diakoudi G, et al. Identification of hepadnavirus in the sera of cats. Sci Rep. 2019;9(1):10668. https://doi.org/10.1038/s41598-019-47175-8.
Anpuanandam K, Selvarajah GT, Choy MMK, et al. Molecular detection and characterisation of Domestic Cat Hepadnavirus (DCH) from blood and liver tissues of cats in Malaysia. BMC Vet Res. 2021;17(1):9. https://doi.org/10.1186/s12917-020-02700-0.
Pesavento PA, Jackson K, Hampson TSTTB, Munday JS, Barrs VR, Beatty JA. A novel hepadnavirus is associated with chronic hepatitis and hepatocellular carcinoma in cats. Viruses. 2019;11(10):969. https://doi.org/10.3390/v11100969.
Takahashi K, Kaneko Y, Shibanai A, et al. Identification of domestic cat hepadnavirus from a cat blood sample in Japan. J Vet Med Sci. 2022;84(5):648–52. https://doi.org/10.1292/jvms.22-0010.
Tien PT, Lin CJ, Tsai YY, et al. Relationship between uveitis, different types of viral hepatitis, and liver cirrhosis: a 12-Year Nationwide Population-Based Cohort Study. Retina. 2016;36(12):2391–8. https://doi.org/10.1097/IAE.0000000000001103.
Jeanes EC, Wegg ML, Mitchell JA, Priestnall SL, Fleming L, Dawson C. Comparison of the prevalence of Domestic Cat Hepadnavirus in a population of cats with uveitis and in a healthy blood donor cat population in the United Kingdom. Vet Ophthalmol. 2022;25(2):165–72. https://doi.org/10.1111/vop.12956.
Decaro N, Buonavoglia D, Desario C, et al. Characterisation of canine parvovirus strains isolated from cats with feline panleukopenia. Res Vet Sci. 2010;89(2):275–8. https://doi.org/10.1016/j.rvsc.2010.03.001.
Hoofnagle JH, Schafer DF, Ferenci P, et al. Antibody to hepatitis B surface antigen in nonprimate animal species. Gastroenterology. 1983;84(6):1478–82.
Chen DS, Sung JL, Lai MY, Liu MR. Antibody to hepatitis B surface antigen in dogs in Taiwan. Gastroenterology. 1984;86(5 Pt 1):1001–3.
Vieira YR, Portilho MM, Oliveira FF, et al. Evaluation of HBV-like circulation in wild and farm animals from Brazil and Uruguay. Int J Environ Res Public Health. 2019;16(15):2679. https://doi.org/10.3390/ijerph16152679.
Al-Jumaa ZM, Ajaj EA, Dahl MO. Seroprevalence of canine hepatitis in stray dogs in Nineveh Province. Iraq Vet World. 2020;13(11):2326–9. https://doi.org/10.14202/vetworld.2020.2326-2329.
Diakoudi G, Capozza P, Lanave G, et al. A novel hepadnavirus in domestic dogs. Sci Rep. 2022;12(1):2864. https://doi.org/10.1038/s41598-022-06842-z.
Shofa M, Kaneko Y, Takahashi K, Okabayashi T, Saito A. Global Prevalence of domestic cat hepadnavirus: an emerging threat to cats’ health? Front Microbiol. 2022;13:938154. https://doi.org/10.3389/fmicb.2022.938154.
Kamimura T, Yoshikawa A, Ichida F, Sasaki H. Electron microscopic studies of Dane particles in hepatocytes with special reference to intracellular development of Dane particles and their relation with HBeAg in serum. Hepatology. 1981;1(5):392–7. https://doi.org/10.1002/hep.1840010504.
Sakamoto Y, Yamada G, Mizuno M, et al. Full and empty particles of hepatitis B virus in hepatocytes from patients with HBsAg-positive chronic active hepatitis. Lab Invest. 1983;48(6):678–82.
Kim WR, Flamm SL, Di Bisceglie AM, Bodenheimer HC. Public policy committee of the american association for the study of liver disease serum activity of alanine aminotransferase (ALT) as an indicator of health and disease. Hepatology. 2008;47(4):1363–70. https://doi.org/10.1002/hep.22109.
Choi YR, Chen M-C, Carrai M, Rizzo F, Chai Y, Tse M et al. Does a hepadnavirus contribute to chronic hepatitis or hepatocellular carcinoma in dogs? In: Paper presented at 6th international society for companion animal infectious diseases symposium (ISCAID 2022), Glasgow, United Kingdom; 2022.
Hou J, Liu Z, Gu F. Epidemiology and prevention of hepatitis B virus infection. Int J Med Sci. 2005;2:50–7. https://doi.org/10.7150/IJMS.2.50.
Capozza P, Lanave G, Diakoudi G, et al. A longitudinal observational study in two cats naturally-infected with hepadnavirus. Vet Microbiol. 2021;254:108999. https://doi.org/10.1016/j.vetmic.2021.108999.
Bittner JJ. Some possible effects of nursing on the mammary gland tumor incidence in mice. Science. 1936;84(2172):162. https://doi.org/10.1126/science.84.2172.162.
Ross SR. Mouse mammary tumor virus molecular biology and oncogenesis. Viruses. 2010;2(9):2000–12. https://doi.org/10.3390/v2092000.
Wei WZ, Gill RF, Wang H. Mouse mammary tumor virus associated antigens and superantigens–immuno-molecular correlates of neoplastic progression. Semin Cancer Biol. 1993;4(3):205–13.
McGrath CM, Jones RF. Hormonal induction of mammary tumor viruses and its implications for carcinogenesis. Cancer Res. 1978;38(11 Pt 2):4112–25.
Liu B, Wang Y, Melana SM, et al. Identification of a proviral structure in human breast cancer. Cancer Res. 2001;61(4):1754–9.
Maeda T, Maeda M, Stewart AF. TEF-1 transcription factors regulate activity of the mouse mammary tumor virus LTR. Biochem Biophys Res Commun. 2002;296(5):1279–85. https://doi.org/10.1016/s0006-291x(02)02085-5.
Zhou Y, Huang T, Cheng AS, Yu J, Kang W, To KF. The TEAD family and its oncogenic role in promoting tumorigenesis. Int J Mol Sci. 2016;17(1):138. https://doi.org/10.3390/ijms17010138.
Ball JK, Dekaban GA, McCarter JA, Loosmore SM. Molecular biological characterization of a highly leukaemogenic virus isolated from the mouse III Identity with mouse mammary tumour virus. J Gen Virol. 1983;64(Pt 10):2177–90. https://doi.org/10.1099/0022-1317-64-10-2177.
Ball JK, Arthur LO, Dekaban GA. The involvement of a type-B retrovirus in the induction of thymic lymphomas. Virology. 1985;140(1):159–72. https://doi.org/10.1016/0042-6822(85)90455-6.
Mertz JA, Mustafa F, Meyers S, Dudley JP. Type B leukemogenic virus has a T-cell-specific enhancer that binds AML-1. J Virol. 2001;75(5):2174–84. https://doi.org/10.1128/JVI.75.5.2174-2184.2001.
Broussard DR, Mertz JA, Lozano M, Dudley JP. Selection for c-myc integration sites in polyclonal T-cell lymphomas. J Virol. 2002;76(5):2087–99. https://doi.org/10.1128/jvi.76.5.2087-2099.2002.
Nicoll CS, Tucker HA. Estimates of parenchymal, stromal, and lymph node deoxyribonucleic acid in mammary glands of C3H/Crgl-2 mice. Life Sci. 1965;4(9):993–1001. https://doi.org/10.1016/0024-3205(65)90203-1.
Cardiff RD, Anver MR, Gusterson BA, et al. The mammary pathology of genetically engineered mice: the consensus report and recommendations from the Annapolis meeting. Oncogene. 2000;19(8):968–88. https://doi.org/10.1038/sj.onc.1203277.
DeOme KB, Miyamoto MJ, Osborn RC, Guzman RC, Lum K. Effect of parity on recovery of inapparent nodule-transformed mammary gland cells in vivo. Cancer Res. 1978;38(11 Pt 2):4050–3.
Kordon EC, Smith GH, Callahan R, Gallahan D. A novel non-mouse mammary tumor virus activation of the Int-3 gene in a spontaneous mouse mammary tumor. J Virol. 1995;69(12):8066–9. https://doi.org/10.1128/JVI.69.12.8066-8069.1995.
Young LJ, Medina D, DeOme KB, Daniel CW. The influence of host and tissue age on life span and growth rate of serially transplanted mouse mammary gland. Exp Gerontol. 1971;6(1):49–56. https://doi.org/10.1016/0531-5565(71)90048-9.
Cohen JC, Majors JE, Varmus HE. Organization of mouse mammary tumor virus-specific DNA endogenous to BALB/c mice. J Virol. 1979;32(2):483–96. https://doi.org/10.1128/JVI.32.2.483-496.1979.
Cohen JC, Shank PR, Morris VL, Cardiff R, Varmus HE. Integration of the DNA of mouse mammary tumor virus in virus-infected normal and neoplastic tissue of the mouse. Cell. 1979;16(2):333–45. https://doi.org/10.1016/0092-8674(79)90010-2.
Kordon EC, Smith GH. An entire functional mammary gland may comprise the progeny from a single cell. Development. 1998;125(10):1921–30. https://doi.org/10.1242/dev.125.10.1921.
Callahan R, Smith GH. Common integration sites for MMTV in viral induced mouse mammary tumors. J Mammary Gland Biol Neoplasia. 2008;13(3):309–21. https://doi.org/10.1007/s10911-008-9092-6.
Callahan R, Smith GH. MMTV-induced mammary tumorigenesis: gene discovery, progression to malignancy and cellular pathways. Oncogene. 2000;19(8):992–1001. https://doi.org/10.1038/sj.onc.1203276.
Dudley JP, Golovkina TV, Ross SR. Lessons learned from mouse mammary tumor virus in animal models. ILAR J. 2016;57(1):12–23. https://doi.org/10.1093/ilar/ilv044.
Nusse R, Varmus HE. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell. 1982;31(1):99–109. https://doi.org/10.1016/0092-8674(82)90409-3.
Peters G, Brookes S, Smith R, Dickson C. Tumorigenesis by mouse mammary tumor virus: evidence for a common region for provirus integration in mammary tumors. Cell. 1983;33(2):369–77. https://doi.org/10.1016/0092-8674(83)90418-x.
Nusse R. Insertional mutagenesis in mouse mammary tumorigenesis. Curr Top Microbiol Immunol. 1991;171:43–65. https://doi.org/10.1007/978-3-642-76524-7_3.
Clausse N, Baines D, Moore R, Brookes S, Dickson C, Peters G. Activation of both Wnt-1 and Fgf-3 by insertion of mouse mammary tumor virus downstream in the reverse orientation: a reappraisal of the enhancer insertion model. Virology. 1993;194(1):157–65. https://doi.org/10.1006/viro.1993.1245.
Amarante MK, de Sousa PN, Vitiello GAF, Watanabe MAE. Involvement of a mouse mammary tumor virus (MMTV) homologue in human breast cancer: evidence for, against and possible causes of controversies. Microb Pathog. 2019;130:283–94. https://doi.org/10.1016/j.micpath.2019.03.021.
Szabo S, Haislip AM, Garry RF. Of mice, cats, and men: is human breast cancer a zoonosis? Microsc Res Tech. 2005;68(3–4):197–208. https://doi.org/10.1002/jemt.20232.
Parisi F, Freer G, Mazzanti CM, Pistello M, Poli A. Mouse mammary tumor virus (MMTV) and MMTV-like viruses: an in-depth look at a controversial issue. Viruses. 2022;14(5):977. https://doi.org/10.3390/v14050977.
Muhlbock O. Mammary tumor-agent in the sperm of high-cancer-strain male mice. J Natl Cancer Inst. 1950;10(4):861–4.
Tsubura Y, Imai S, Morimoto J, Hilgers J. Strain difference in the expression of mammary tumor virus antigen in the male genital organs of mice during aging. Gan. 1981;72(3):424–9.
Imai S, Okumoto M, Iwai M, et al. Distribution of mouse mammary tumor virus in Asian wild mice. J Virol. 1994;68(5):3437–42. https://doi.org/10.1128/JVI.68.5.3437-3442.1994.
Lee WT, Prakash O, Klein D, Sarkar NH. Structural alterations in the long terminal repeat of an acquired mouse mammary tumor virus provirus in a T-cell leukemia of DBA/2 mice. Virology. 1987;159(1):39–48. https://doi.org/10.1016/0042-6822(87)90345-x.
Yanagawa S, Murakami A, Tanaka H. Extra mouse mammary tumor proviruses in DBA/2 mouse lymphomas acquire a selective advantage in lymphocytes by alteration in the U3 region of the long terminal repeat. J Virol. 1990;64(6):2474–83.
Cotterchio M, Nadalin V, Sauer M. Human breast cancer and lymphomas may share a common aetiology involving Mouse Mammary Tumour Virus (MMTV). Med Hypotheses. 2002;59(4):492–4. https://doi.org/10.1016/s0306-9877(02)00204-9.
Etkind P, Du J, Khan A, Pillitteri J, Wiernik PH. Mouse mammary tumor virus-like ENV gene sequences in human breast tumors and in a lymphoma of a breast cancer patient. Clin Cancer Res. 2000;6(4):1273–8.
Etkind PR, Stewart AF, Dorai T, Purcell DJ, Wiernik PH. Clonal isolation of different strains of mouse mammary tumor virus-like DNA sequences from both the breast tumors and non-Hodgkin’s lymphomas of individual patients diagnosed with both malignancies. Clin Cancer Res. 2004;10(17):5656–64. https://doi.org/10.1158/1078-0432.CCR-03-0364.
Johal H, Scott GM, Jones R, Camaris C, Riordan S, Rawlinson WD. Mouse mammary tumour virus-like virus (MMTV-LV) is present within the liver in a wide range of hepatic disorders and unrelated to nuclear p53 expression or hepatocarcinogenesis. J Hepatol. 2009;50(3):548–54. https://doi.org/10.1016/j.jhep.2008.10.026.
Lushnikova AA, Kriukova IN, Malivanova TF, Makhov PB, Polevaia EB. Korreliatsiia mezhdu ékspressieĭ antigena, immunologicheski rodstvennogo gp52, i transkriptsieĭ gomologichnykh ENV MMTV posledovatel’nosteĭ DNK v limfotsitakh perifericheskoĭ krovi bol’nykh rakom molochnoĭ zhelezy [Correlation between expression of antigen immunologically related to gp52 MMTV and transcription of homologous ENV MMTV DNA sequences in peripheral blood lymphocytes from breast cancer patients]. Mol Gen Mikrobiol Virusol. 1998;3:33–6.
Mason AL, Xu L, Guo L, et al. Detection of retroviral antibodies in primary biliary cirrhosis and other idiopathic biliary disorders. Lancet. 1998;351(9116):1620–4. https://doi.org/10.1016/S0140-6736(97)10290-2.
Wang W, Indik S, Wasilenko ST, et al. Frequent proviral integration of the human betaretrovirus in biliary epithelium of patients with autoimmune and idiopathic liver disease. Aliment Pharmacol Ther. 2015;41(4):393–405. https://doi.org/10.1111/apt.13054.
Michalides R, Wagenaar E, Hilkens J, Hilgers J, Groner B, Hynes NE. Acquisition of proviral DNA of mouse mammary tumor virus in thymic leukemia cells from GR mice. J Virol. 1982;43(3):819–29. https://doi.org/10.1128/JVI.43.3.819-829.1982.
Dekaban GA, Ball JK. Integration of type B retroviral DNA in virus-induced primary murine thymic lymphomas. J Virol. 1984;52(3):784–92. https://doi.org/10.1128/JVI.52.3.784-792.1984.
Dudley JP, Arfsten A, Hsu CL, Kozak C, Risser R. Molecular cloning and characterization of mouse mammary tumor proviruses from a T-cell lymphoma. J Virol. 1986;57(1):385–8. https://doi.org/10.1128/JVI.57.1.385-388.1986.
Johal H, Faedo M, Faltas J, et al. DNA of mouse mammary tumor virus-like virus is present in human tumors influenced by hormones. J Med Virol. 2010;82(6):1044–50. https://doi.org/10.1002/jmv.21754.
Deligdisch L, Marin T, Lee AT, et al. Human mammary tumor virus (HMTV) in endometrial carcinoma. Int J Gynecol Cancer. 2013;23(8):1423–8. https://doi.org/10.1097/IGC.0b013e3182980fc5.
Mazzanti CM, Lessi F, Armogida I, et al. Human saliva as route of inter-human infection for mouse mammary tumor virus. Oncotarget. 2015;6(21):18355–63.
Stewart TH, Sage RD, Stewart AF, Cameron DW. Breast cancer incidence highest in the range of one species of house mouse. Mus domesticus Br J Cancer. 2000;82(2):446–51.
Stewart AFR, Chen HH. Revisiting the MMTV zoonotic hypothesis to account for geographic variation in breast cancer incidence. Viruses. 2022;14(3):559.
Szabo S, Haislip AM, Traina-Dorge V, et al. Human, rhesus macaque, and feline sequences highly similar to mouse mammary tumor virus sequences. Microsc Res Tech. 2005;68(3–4):209–21. https://doi.org/10.1002/jemt.20233.
Hsu WL, Lin HY, Chiou SS, et al. Mouse mammary tumor virus-like nucleotide sequences in canine and feline mammary tumors. J Clin Microbiol. 2010;48(12):4354–62.
Civita P, Menicagli M, Scopelliti C, et al. Mouse mammary tumour virus-like env nucleotide and p14 signal peptide are present in feline mammary carcinomas, but not in neoplastic or dysplastic canine mammary lesions. PLoS ONE. 2018;13(7):e0200839. https://doi.org/10.1371/journal.pone.0200839.
Parisi F, Muscatello LV, Civita P, et al. Pathological features and molecular phenotype of MMTV like-positive feline mammary carcinomas. Animals. 2021;11(10):2821. https://doi.org/10.3390/ani11102821.
Parisi F, Lessi F, Menicagli M, et al. Presence of a mouse mammary tumour virus-like in feline lymphomas: a preliminary study. Infect Agent Cancer. 2022;17(1):35. https://doi.org/10.1186/s13027-022-00449-9.
Funding
This research received no external funding.
Author information
Authors and Affiliations
Contributions
Conceptualization, FP, MP and AP; development the concepts, FP, GF; writing—original draft preparation, FP; writing—review and editing, NF, FM, GF, MP, and AP; supervision, AP; All authors have read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Parisi, F., Fonti, N., Millanta, F. et al. Exploring the link between viruses and cancer in companion animals: a comprehensive and comparative analysis. Infect Agents Cancer 18, 40 (2023). https://doi.org/10.1186/s13027-023-00518-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13027-023-00518-7