
Abstract
Major depressive disorder (MDD) is predicted to become the second most common cause of disability in the near future. Exposure to glyphosate (Gly)-based herbicides has been linked to the onset of MDD. However, the underlying mechanisms remain unclear. The aim of this study was to investigate the potential molecular mechanisms of MDD induced by Gly using network toxicology approach. The MDD dataset GSE76826 from the Gene Expression Omnibus database was referenced to identify differentially expressed genes (DEGs) in peripheral blood leukocytes of MDD patients and controls. The potential intersection targets of Gly-induced MDD were screened by network toxicology. The intersection targets were used for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis and to construct protein–protein interaction networks. The binding potentials of hub targets with Gly were validated by molecular docking. In total, 1216 DEGs associated with Gly-induced MDD were identified. Subsequent network pharmacology further refined the search to 43 targets. GO and KEGG enrichment analyses revealed multiple signaling pathways involved in GLY-induced MDD. Six potential core targets (CD40, FOXO3, FOS, IL6, TP53, and VEGFA) were identified. Finally, molecular docking demonstrated that Gly exhibited strong binding affinity to the core targets. The results of this study identified potential molecular mechanisms underlying Gly induced MDD and provided new insights for prevention and treatment.
Similar content being viewed by others


Introduction
Major depressive disorder (MDD), commonly referred to as depression, has a significant impact on mental health, affecting approximately 3% of the global population, equating to approximately 216 million individuals [24]. Notably, individuals with depression may experience suicidal tendencies, underscoring the importance of early identification and prompt treatment of this debilitating condition [32]. MDD is anticipated to become the second most common cause of disability, particularly among the elderly population, by the year 2030 [14]. MDD is a complex disorder influenced by biological, genetic, and environmental factors [4].
Glyphosate (Gly) is a widely used active ingredient in herbicides, leading to widespread scrutiny of the potential toxicity of Gly-based formulations [22]. Worldwide, approximately 8.6 billion kilograms of Gly are applied to more than 350 million hectares annually [6]. Both in vivo and in vitro investigations have demonstrated toxic effects of Gly on both embryonic development and neurological systems [21]. Recent epidemiological investigations have demonstrated an association between Gly exposure and the incidence of MDD [11]. In particular, mice exposed to a low dose of Gly-based herbicide over a subchronic period exhibited behaviors resembling depression and anxiety [9]. In consideration of the wide use of Gly, potential harm to human health must be investigated, especially the potential molecular mechanisms underlying Gly-induced depression.
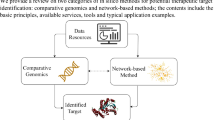
Therefore, the aim of the present study was to identify differentially expressed genes (DEGs) in peripheral blood leukocytes from MDD patients and controls. Furthermore, network toxicology and molecular docking strategies were used to screen potential targets of Gly exposure linked to the development of MDD. The results of this study indicate that exposure to Gly may be associated with the development of MDD and highlight the need for further investigation to elucidate the molecular mechanisms underlying this association. The study design and workflow are presented in Fig. 1.

Study design and workflow
Materials and methods
Genetic microarray data sources
The GSE76826 dataset was obtained from the Gene Expression Omnibus database (http://www.ncbi.nlm.nih.gov/). The study focused on nine older outpatients and inpatients (age ≥ 50 years) with MDD according to Diagnostic and Statistical Manual of Mental Disorders, 4th ed (DSM-IV) criteria. The severity of depression was assessed using the Hamilton Depression rating scale (SIGH-D) through structured interviews. In addition, 12 healthy individuals were included for comparison. Blood samples were collected, and RNA was extracted from peripheral blood leukocytes using the QIAmp RNA Blood Mini Kit. Microarray data were obtained based on the GPL17077 platform and conducted using the SurePrint G3 Human GE v2 8 × 60K Microarray (Agilent Technologies, Inc., Santa Clara, CA, USA). This study used publicly available gene data for analysis, and was therefore exempted from ethical approval.
Identification of DEGs
The original data were recorded in the MINiML file format. The “normalize.quantiles” function from the “preprocessCore” package of the R software (version 3.4.1; https://www.r-project.org/) was used to standardize the microarray data. The normalized data platform provided annotation information, which was utilized to convert probes into gene symbols. All probes that matched multiple genes were eliminated from the datasets. The final expression value was calculated from the average expression value of genes that were identified by several probes. The criteria for MDD-related DEGs were |log2 fold change| ≥ 0.585 and a false discovery rate (FDR) < 0.05. Considering multiple testing, the Benjamini–Hochberg (BH) method was used in this study to adjust the p value to control for FDR. A difference was considered statistically significant if the BH-adjusted p value was less than 0.05.
Network toxicology analysis
The GeneCards database of human genes (http://www.genecards.org/), STITCH database of known and predicted interactions between chemicals and proteins (http://stitch.embl.de/), and the Comparative Toxicogenomics Database (CTD, http://ctdbase.org/) were searched to identify potential targets of Gly-induced MDD. The two-dimensional structures of Gly were obtained from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). The CTD screening conditions were set to default parameters with the target species set to Homo sapiens. Similarly, the target species for searching of the STITCH database was set to H. sapiens and the minimum required interaction score was set to 0.4. The Draw Venn Diagram tool (http://bioinformatics.psb.ugent.be/webtools/Venn/) was used to identify the intersecting targets of Gly that may contribute to MDD. A protein–protein interaction (PPI) network was constructed of the intersecting targets using the STRING database (https://string-db.org/), with a minimum required interaction score of 0.15 for the species H. sapiens, and visualized using Cytoscape 3.6.1 (https://cytoscape.org/), an open source software platform for visualizing complex networks.
Gene-set enrichment analysis
Gene-set enrichment analysis was performed using the Metascape gene annotation and analysis resource (https://metascape.org/gp/index.html) with reference to the Gene Ontology (GO) knowledgebase (http://geneontology.org/) and the Kyoto Encyclopedia of Genes and Genomes (KEGG; https://www.genome.jp/kegg/). The threshold of statistical significance was set to p < 0.05. The top 10 GO terms for biological process (BP), cellular component (CC), and molecular function (MF), in addition to the top 10 KEGG pathways were selected for further investigation.
Hub targets screening
The cytoHubba plugin (https://apps.cytoscape.org/apps/cytohubba) was used to identify important nodes/hubs based on eight topological algorithms [betweenness, bottleneck, closeness, degree, edge percolated component (EPC), maximal clique centrality (MCC), maximum neighborhood component (MNC), and stress]. Then, the Draw Venn Diagram tool was employed to identify the top 10 overlapping (hub) targets.
Molecular docking
To conduct molecular docking of Gly to the hub targets, the three-dimensional structure of Gly was retrieved from the PubChem database and converted into the Mol2 format using UCSF Chimera software version 1.16. In addition, the three-dimensional structures of the core targets were retrieved from the Worldwide Protein Data Bank (https://www.wwpdb.org/). Hydrogenation, dehydration, and ligand removal of the core targets were assessed with UCSF Chimera software (https://www.cgl.ucsf.edu/chimera/). The SwissDock online platform (http://www.swissdock.ch/) was utilized to perform molecular docking and to assess the binding activity of Gly and the hub targets. The results of the docking process were visualized using the UCSF Chimera software.
Results
DEG screening and network toxicological analysis
Overall, 1216 (4.8%) of the identified 25,298 mRNAs were DEGs, of which 776 were upregulated and 440 were downregulated (Fig. 2a and b, respectively). After removing duplicates, 472 targets of Gly were retrieved from the CTD, STITCH, and GeneCards databases. Venn analysis identified 43 intersection targets (32 upregulated and 11 downregulated; Fig. 2c), which were used to construct a PPI network using the STRING database, which included 43 nodes and 329 edges with an average node degree of 15.3 (Fig. 2d, Additional file 1: Table S1).

DEG screening and network toxicological analysis. a Volcano plot of DEGs between MDD patients and control. The hub targets were marked with black arrows. b Heatmap of DEGs between MDD patients and control. c The Venn map of Gly-related targets and MDD-related targets. d The PPI network. The width of each edge is proportional to the combined score, whereas the color (from light to dark) and size of each node are proportional to interaction strength
GO and KEGG enrichment analysis
GO and KEGG pathway enrichment analyses were conducted to investigate the potential mechanisms underlying Gly-induced MDD (Additional file 1: Table S2–3). The overlapping targets were enriched in the BP terms “positive regulation of cell death”, “positive regulation of apoptotic process”, and “regulation of stress-activated MAPK cascade”; the CC terms “presynaptic active zone”, “transcription regulator complex”, and “plasma membrane signaling receptor complex”; and the MF terms “kinase binding”, “growth factor receptor binding”, and “signaling receptor activator activity” (Fig. 3a–c). The KEGG pathway analysis revealed that the common targets were enriched in the terms “MAPK signaling pathway”, “Apoptosis”, and “PI3K-Akt signaling pathway” (Fig. 3d).

GO and KEGG enrichment analysis. a BP enrichment analysis, b CC enrichment analysis, c MF enrichment analysis, and d KEGG pathway enrichment analysis
Hub target identification
The top 10 targets were identified using eight topological analysis methods (betweenness, bottleneck, closeness, degree, EPC, MCC, MNC, and stress) (Fig. 4a–h, Additional file 1: Table S4-11). The top six hub targets, as determined by Venn analysis, included cluster of differentiation 40 (CD40), forkhead box O3 (FOXO3), Fos proto-oncogene, AP-1 transcription factor subunit (FOS), interleukin 6 (IL6), tumor protein p53 (TP53), and vascular endothelial growth factor A (VEGFA) (Fig. 4i). PPI networks of the top six hub targets were constructed with the STRING database (Fig. 4j).

Hub target identification. Top 10 targets calculated through a the betweenness method, b the bottleneck method, c the closeness method, d the degree method, e the EPC method, and f the MCC method, g the MNC method, and h the stress method. i A Venn map of the top 6 core targets. h The PPI network of the 6 targets
Molecular docking
Molecular docking analysis was performed to further investigate the ability of Gly to bind to the top six core targets. The binding energies of Gly with CD40, FOXO3, FOS, IL6, TP53, and VEGFA were − 6.63, − 6.45, − 6.12, − 6.88, − 6.88, and − 6.19 kcal/mol, respectively (Fig. 5a–f).

Molecular docking. a Gly-CD40, b Gly-FOS, c Gly-FOXO3, d Gly-IL6, e Gly-TP53, and f Gly-VEGFA
Discussion
Environmental factors have been reported to play a significant role in the onset of MDD, as the majority of cases are believed to occur randomly [37]. Epidemiological studies have suggested a possible association between exposure to pesticides and the development of MDD [5]. To better understand the molecular mechanisms underlying Gly-induced MDD, bioinformatics algorithms were used to explore aberrantly expressed mRNAs between MDD patients and controls, which identified 1216 DEGs (776 upregulated and 440 downregulated) from the GSE76826 dataset. Network pharmacology predicted 43 potential targets of Gly associated with the induction of MDD (32 upregulated and 11 downregulated). GO and KEGG analyses were performed to investigate the biological characteristics of these potential targets and six hub targets (CD40, FOXO3, FOS, IL6, TP53, and VEGFA) were identified using molecular docking.
CD40 is a transmembrane protein and a member of the tumor necrosis factor receptor superfamily that plays a crucial role in the initiation and maintenance of inflammatory reactions [16]. CD40-mediated aberrant neuroinflammation can increase permeability of the blood–brain barrier and subsequently damage neurons and glial cells [30]. A previous study suggested that newly diagnosed depression is associated with increased expression of platelet-derived CD40 [29]. In addition, depression-like behavior was associated with abnormal microglial expression of CD40 in a rat model of Alzheimer’s disease [17]. Gly has been demonstrated to induce death of human neuroblastoma SH-SY5Y cells through upregulation of CD40 expression [27]. Therefore, CD40 and related inflammatory signaling pathways are potential targets for the treatment of Gly-induced MDD.
FOXO3 is a crucial transcription factor that regulates various gene networks associated with cellular metabolism that contribute to various physiological and pathological processes [8], and plays a critical role in oxidative stress-induced neuronal death [39]. Recent studies have indicated an association between MDD and selective and persistent death of hippocampal neurons [23]. Although the role of FOXO3 in MDD remains unclear, a prior epidemiological study suggested that genetic variants of FOXO3 may be associated with depressive symptoms in older adults [26]. It has been reported that Gly exposure induced proliferation of breast cancer cells through upregulation of FOXO3 mRNA expression [33]. Collectively, these findings suggest that FOXO3 may play a key role in Gly-induced MDD, although further investigations are needed to clarify the underlying mechanisms.
FOS is a transcription factor that is rapidly and transiently activated [15]. Hence, numerous studies have advocated the use of FOS as an indicator of stimulus-triggered modifications in brain function [15]. Many studies have reported altered expression of FOS in models of depression. Blue light deprivation is reported to produce a depression-like response in Mongolian gerbils accompanied by decreased FOS expression in brain tissues [20]. Another study reported that rice wine can potentially reduce stress-induced depression-like behaviors and FOS expression in rats [28]. In addition, FOS was associated with Gly-induced hematotoxicity via dysregulation of hematopoietic stem cell function [19] and Gly-induced anxiety and depression-like behavior in mice was accompanied by increased FOS expression in brain tissues [1]. However, it remains unclear whether FOS is a key molecule in Gly-induced MDD.
Increased expression of the cytokine IL6, in both peripheral and central systems, is believed to play a vital role in stress reactions and depressive disorders, especially as a comorbidity with physical illnesses [13, 40, 41]. Elevated IL6 activity can potentially induce depression via stimulation of the hypothalamic–pituitary–adrenal axis or modulation of neurotransmitter metabolism [36]. Hence, manipulation of IL-6 activity could benefit individuals with MDD and inflammatory characteristics [36]. Perinatal exposure to Gly significantly increased IL6 expression and caused liver damage in rat offspring [31]. In addition, Gly induced an inflammatory response mediated by elevated IL6 and altered the microbial composition in the rat intestine [34]. Taken together, these findings suggest that IL6 is a potential key molecule in the onset of Gly-induced MDD.
TP53 acts as a transcription factor that regulates the expression of almost 500 target genes that participate in various cellular processes, including cell death, the cell cycle, cell senescence, DNA repair, and metabolism [35]. In addition, TP53 is involved in regulation of the mitogen-activated protein kinase signaling pathway, which plays an important role in the development and progression of MDD [12]. Epidemiological studies have shown that genetic polymorphisms of TP53 are strongly associated with MDD and the minor allele 72 C may be a protective factor against MDD [25]. Gly has been reported to influence TP53 expression in human peripheral blood mononuclear cells by increasing methylation of the TP53 promoter [38]. Recent research of TP53 has mainly focused on tumor formation and development. However, the results of the present study found that TP53 may play a key role in Gly-induced MDD. Therefore, further studies are warranted to investigate the underlying mechanisms.
VEGF is elevated in damaged vessels and neurons, abnormal branching, and chronic inflammatory conditions, such as cardiovascular disease and depression [2, 10]. In a previous study, serum VEGFA mRNA and protein levels were significantly elevated in patients with MDD, suggesting a potentially important role in the pathogenesis of depression [7]. Another study found increased mRNA levels of VEGFA in peripheral blood cells and serum in patients with recurrent depression as compared to healthy controls, and suggested that VEGFA gene polymorphism could be a prognostic factor for the development of recurrent depression [18]. Furthermore, it has been reported that exposure to Gly during pregnancy reduces placental vascular density and cell proliferation via interference with VEGFA expression and subsequent impaired barrier function and nutrient transport in the placenta of newborn piglets [3]. These findings combined with the results of the present study indicate that VEGFA may play an important role in the onset of Gly-induced MDD.
Conclusion
Network toxicology analyses of the molecular pathways associated with Gly-induced MDD identified CD40, FOXO3, FOS, IL6, TP53, and VEGFA as potential targets. The findings of this preliminary examination of the correlation between exposure to Gly-based herbicides and MDD offer useful references for future research. Nevertheless, further in vivo and in vitro studies are needed to validate these results.
Availability of data and materials
All data in this study are available from the corresponding authors upon request.
References
Ait Bali Y, et al. Anxiety and gene expression enhancement in mice exposed to glyphosate-based herbicide. Toxics. 2022;10(5):226.
Anisman H. Inflaming depression. J Psychiatry Neurosci. 2011;36(5):291–5.
Bai G, et al. Perinatal exposure to glyphosate-based herbicides impairs progeny health and placental angiogenesis by disturbing mitochondrial function. Environ Int. 2022;170: 107579.
Bakusic J, et al. Increased methylation of NR3C1 and SLC6A4 is associated with blunted cortisol reactivity to stress in major depression. Neurobiol Stress. 2020;13: 100272.
Barnett JA, et al. Is the use of glyphosate in modern agriculture resulting in increased neuropsychiatric conditions through modulation of the gut-brain-microbiome axis. Front Nutr. 2022;9: 827384.
Belbin FE, et al. Plant circadian rhythms regulate the effectiveness of a glyphosate-based herbicide. Nat Commun. 2019;10(1):3704.
Berent D, et al. Vascular endothelial growth factor A gene expression level is higher in patients with major depressive disorder and not affected by cigarette smoking, hyperlipidemia or treatment with statins. Acta Neurobiol Exp (Wars). 2014;74(1):82–90.
Bernardo VS, et al. FoxO3 and oxidative stress: a multifaceted role in cellular adaptation. J Mol Med (Berl). 2023;101(1–2):83–99.
Bicca DF, et al. A subchronic low-dose exposure of a glyphosate-based herbicide induces depressive and anxious-like behavior in mice: quercetin therapeutic approach. Environ Sci Pollut Res Int. 2021;28(47):67394–403.
Chancharoenthana W, et al. Enhanced vascular endothelial growth factor and inflammatory cytokine removal with online hemodiafiltration over high-flux hemodialysis in sepsis-related acute kidney injury patients. Ther Apher Dial. 2013;17(5):557–63.
Chang ET, et al. Systematic literature review of the epidemiology of glyphosate and neurological outcomes. Int Arch Occup Environ Health. 2023;96(1):1–26.
Chen Y, et al. Identification of potential Mitogen-Activated Protein Kinase-related key genes and regulation networks in molecular subtypes of major depressive disorder. Front Psychiatry. 2022;13:1004945.
Chourbaji S, et al. IL-6 knockout mice exhibit resistance to stress-induced development of depression-like behaviors. Neurobiol Dis. 2006;23(3):587–94.
Chu SF, et al. Low corticosterone levels attenuate late life depression and enhance glutamatergic neurotransmission in female rats. Acta Pharmacol Sin. 2021;42(6):848–60.
Cruz-Mendoza F, et al. Immediate Early gene c-fos in the brain: focus on glial cells. Brain Sci. 2022;12(6):687.
Elgueta R, et al. Molecular mechanism and function of CD40/CD40L engagement in the immune system. Immunol Rev. 2009;229(1):152–72.
Farhangian M, et al. Intranasal interferon-beta alleviates anxiety and depressive-like behaviors by modulating microglia polarization in an Alzheimer’s disease model. Neurosci Lett. 2023;792: 136968.
Gałecki P, et al. Vascular endothelial growth factor gene (VEGFA) polymorphisms may serve as prognostic factors for recurrent depressive disorder development. Prog Neuropsychopharmacol Biol Psychiatry. 2013;45:117–24.
He Y, et al. Hematological effects of glyphosate in mice revealed by traditional toxicology and transcriptome sequencing. Environ Toxicol Pharmacol. 2022;92: 103866.
Hu H, et al. Blue light deprivation produces depression-like responses in Mongolian gerbils. Front Psychiatry. 2020;11:233.
Ji H, et al. Differential microRNA expression in the prefrontal cortex of mouse offspring induced by glyphosate exposure during pregnancy and lactation. Exp Ther Med. 2018;15(3):2457–67.
Kier LD. Review of genotoxicity biomonitoring studies of glyphosate-based formulations. Crit Rev Toxicol. 2015;45(3):209–18.
Lee AL, et al. Stress and depression: possible links to neuron death in the hippocampus. Bipolar Disord. 2002;4(2):117–28.
Liu G, et al. P11 loss-of-function is associated with decreased cell proliferation and neurobehavioral disorders in mice. Int J Biol Sci. 2019;15(7):1383–95.
Mahmood S, et al. Association of EGF, IGFBP-3 and TP53 gene polymorphisms with major depressive disorder in Slovak population. Cent Eur J Public Health. 2016;24(3):223–30.
Margrett JA, et al. Novel functional, health, and genetic determinants of cognitive terminal decline: Kuakini Honolulu Heart Program/Honolulu-Asia aging study. J Gerontol A Biol Sci Med Sci. 2022;77(8):1525–33.
Martínez MA, et al. Use of human neuroblastoma SH-SY5Y cells to evaluate glyphosate-induced effects on oxidative stress, neuronal development and cell death signaling pathways. Environ Int. 2020;135: 105414.
Nakatani Y, et al. Japanese Rice Wine can reduce psychophysical stress-induced depression-like behaviors and Fos expression in the trigeminal subnucleus caudalis evoked by masseter muscle injury in the rats. Biosci Biotechnol Biochem. 2019;83(1):155–65.
Neubauer H, et al. Newly diagnosed depression is associated with increased beta-thromboglobulin levels and increased expression of platelet activation markers and platelet derived CD40-CD40L. J Psychiatr Res. 2013;47(7):865–71.
Ots HD, et al. CD40-CD40L in neurological disease. Int J Mol Sci. 2022;23(8):4115.
Rieg C, et al. Perinatal exposure to a glyphosate pesticide formulation induces offspring liver damage. Toxicol Appl Pharmacol. 2022;454: 116245.
Segoviano-Mendoza M, et al. Hypocholesterolemia is an independent risk factor for depression disorder and suicide attempt in Northern Mexican population. BMC Psychiatry. 2018;18(1):7.
Stur E, et al. Glyphosate-based herbicides at low doses affect canonical pathways in estrogen positive and negative breast cancer cell lines. PLoS ONE. 2019;14(7): e0219610.
Tang Q, et al. Glyphosate exposure induces inflammatory responses in the small intestine and alters gut microbial composition in rats. Environ Pollut. 2020;261: 114129.
Thomas AF, et al. Of the many cellular responses activated by TP53, which ones are critical for tumour suppression. Cell Death Differ. 2022;29(5):961–71.
Ting EY, et al. Role of interleukin-6 in depressive disorder. Int J Mol Sci. 2020;21(6):2194.
van den Bosch M, Meyer-Lindenberg A. Environmental exposures and depression: biological mechanisms and epidemiological evidence. Annu Rev Public Health. 2019;40:239–59.
Woźniak E, et al. Glyphosate affects methylation in the promoter regions of selected tumor suppressors as well as expression of major cell cycle and apoptosis drivers in PBMCs (in vitro study). Toxicol In Vitro. 2020;63: 104736.
Xie Q, et al. Lysine methylation of FOXO3 regulates oxidative stress-induced neuronal cell death. EMBO Rep. 2012;13(4):371–7.
Ye G, et al. Association between increased serum interleukin-6 levels and sustained attention deficits in patients with major depressive disorder. Psychol Med. 2018;48(15):2508–14.
Zhang JC, et al. Blockade of interleukin-6 receptor in the periphery promotes rapid and sustained antidepressant actions: a possible role of gut-microbiota-brain axis. Transl Psychiatry. 2017;7(5): e1138.
Funding
This study was supported by the Xuzhou Science and Technology Project (KC22026), the Natural Science Foundation for Colleges and Universities in Jiangsu Province (22KJB330010), and the National Natural Science Foundation of China (82203994 and 82204139).
Author information
Authors and Affiliations
Contributions
JL and HB wrote the main manuscript text and J.L. prepared figures 1–5. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no potential conflicts of interest to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Information of PPI network. Table S2. GO enrichment analysis. Table S3. KEGG enrichment analysis. Table S4. Top 10 targets calculated by betweenness method. Table S5. Top 10 targets calculated by bottleneck method. Table S6. Top 10 targets calculated by closeness method. Table S7. Top 10 targets calculated by degree method. Table S8. Top 10 targets calculated by EPC method. Table S9. Top 10 targets calculated by MCC method. Table S10. Top 10 targets calculated by MNC method. Table S11. Top 10 targets calculated by stress method
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, J., Bi, H. Clarification of the molecular mechanisms underlying glyphosate-induced major depressive disorder: a network toxicology approach. Ann Gen Psychiatry 23, 8 (2024). https://doi.org/10.1186/s12991-024-00491-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12991-024-00491-4