
Abstract
Background
Active metabolism of peripheral blood mononuclear cells (PBMC) could suggest their suitability for metabolomics studies. This study examined whether reductions in PBMCs and plasma lipoprotein-associated phospholipase A2 (Lp-PLA2) activities induced by dietary intervention affected the overall metabolic profiles of PBMC and plasma.
Methods
Eighty nonobese subjects aged 40–70 years (18.5 ≤ BMI < 30 kg/m2) with prediabetes or newly-diagnosed type-2 diabetes were assigned to consume either the usual refined-rice diet (control group, n = 40) or to replace refined rice with whole grains and legumes as carbohydrates (whole-grain group, n = 40) for three meals per day during the 12-week intervention. Fasting PBMC and plasma metabolomes were profiled using UPLC-LTQ-Orbitrap mass spectrometry.
Results
After 12 weeks, changes in fasting glucose, HbA1c, HOMA-IR, MDA, ox-LDL, LDL particle size, plasma Lp-PLA2 activity, and PBMC enzyme activity in the whole-grain group were significantly different from those in the control group before and after adjusting for baseline levels. The PBMC levels of L-leucine, oleamide, lysoPC (16:0), and lysoPC (18:0) in the whole-grain group showed greater reductions compared with those of the control group. Changes in plasma metabolites were not significantly different between the two groups. Changes in PBMC Lp-PLA2 activity positively correlated with changes in L-leucine, oleamide, lysoPC (16:0), lysoPC (18:0), glucose, and ox-LDL, and negatively correlated with changes in LDL particle size.
Conclusions
This study showed that dietary intervention in prediabetic or type-2 diabetic patients had a greater effect on PBMC Lp-PLA2 activity and metabolites compared with those of plasma metabolites.
Trial registration
Similar content being viewed by others

Background
Diabetes is an epidemic metabolic disorder; about 2.7 million Korean people (8.03 %) aged 30 years or older had type-2 diabetes (T2D) and 25.0 % of adults had prediabetes in 2013 according to Korean Diabetes Fact Sheet 2015. Diabetes-related mortality was steadily decreased since 2003 and ranked as the fifth leading cause of natural death [1]. Prediabetes can be indicated by either impaired fasting glucose (IFG) by the American Diabetes Association criteria [2] or impaired glucose tolerance (IGT) by World Health Organization criteria [3].
Lipoprotein-associated phospholipase A2 (Lp-PLA2) independently predicts T2D incidence and may be involved in its etiology [4]. In recent study, an inverse association was observed between protein intake and circulating Lp-PLA2 activity, suggesting that nutritional factors may influence Lp-PLA2 activity [5]. An intervention study that replaced refined rice with whole grains and legumes reduced blood glucose, insulin, Lp-PLA2 activity, and cardiovascular risk factors in patients with prediabetes or T2D [6]. The effects of this intervention diet on plasma and peripheral blood mononuclear cell (PBMC) metabolites have not been determined.
PBMCs include monocytes and lymphocytes which are blood cells having a round nucleus. These blood cells are a critical component in the immune system to fight infection and adapt to intruders. Monocytes have a key role in onset and development of inflammatory reactions by generating bioactive molecules such as Lp-PLA2 in response to inflammatory stimuli [7]. Lymphocytes are consist of three major types; T cells, B cells, and natural killer cells. T cells and B cells are the major cellular components of the adaptive immune response, whereas natural killer cells are a part of the innate immune system. The production and release of Lp-PLA2 by lymphocytes may become increased under inflammatory conditions [8]. Dietary intervention induces PBMC gene expression changes, including downregulating genes involved in inflammatory processes [9]. Therefore, changes in PBMC metabolites and Lp-PLA2 activity after dietary intervention could reflect dynamic responses, which are not detectable in plasma metabolomics analyses. The aim of this 12-week intervention study was to examine whether reductions in PBMC and plasma Lp-PLA2 activities induced by dietary intervention (replacement of refined rice with whole grains and legumes, and higher intake of vegetables) affected the overall metabolic profiles of PBMC and plasma in nonobese patients that exhibited IFG, IGT, or newly-diagnosed T2D.
Methods
Subjects and study design
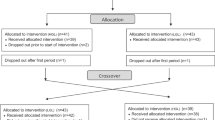
Nonobese subjects aged 40–70 years (18.5 ≤ BMI < 30 kg/m2) were recruited from the Health Service Center (HSC) at the Ilsan Hospital, Goyang, Korea, during January–June 2013. Based on the HSC data, subjects who had IFG (100 ≤ fasting glucose <126 mg/dL) or newly-diagnosed T2D (fasting glucose ≥126 mg/dL) were referred to the Department of Family Medicine or Internal Medicine. Exclusion criteria included: current and/or past history of cardiovascular disease; liver or kidney dysfunction; thyroid or pituitary disease. Subjects who were taking medications or supplements also were excluded. A total of 82 subjects were enrolled. The macronutrient composition of each subject’s usual diet corresponded to a typical diet with cooked refined rice. The purpose of the study was carefully explained to all participants, and written consent was obtained prior to their participation. The Institutional Review Board of the NHIC-sponsored Ilsan Hospital and Yonsei University provided ethical approval of the study protocol, which was performed according to the Helsinki Declaration.
The present study was performed in two phases, including a 2-week run-in phase consisting of the usual diet with refined rice, and a 12-week intervention phase. During the run-in period, two subjects who did not maintain their energy intake dropped out. The remaining 80 subjects were randomly subdivided into the two study groups, and were assigned to consume either the usual refined-rice diet (control group, n = 40) or to replace refined rice with whole grains and legumes as carbohydrates (whole-grain group, n = 40) for three meals per day during the 12-week intervention.
Assessment of dietary intake and physical activity level
All subjects were given written and verbal instructions by a registered dietitian on completion of a 3-day (2 week days and 1 weekend day) dietary record every 2 weeks throughout the study. On the dietary record sheet, subjects were instructed to weigh and record the food amount before and after ingestion. All participants were advised to continue their usual refined-rice diet during a 2-week run-in period. Baseline measurements were performed at the start of the run-in phase. After a run-in period, subjects in the control group maintained the usual refined-rice diet, whereas subjects in the whole-grain group replaced refined rice with a mix of 1/3 legumes, 1/3 barley, and 1/3 wild rice three times per day, and increased vegetable intake to at least 6 units (30–70 g/unit) per day for sufficient dietary fiber intake. The dietitian monitored subject compliance and body-weight changes during the whole study by performing biweekly visits or telephone interviews and all participants were encouraged to maintain their usual lifestyles. Dietary energy values and nutrient contents from 3-day food records were calculated using the CAN-pro 3.0 (Korean Nutrition Society, Seoul, Korea). Total energy expenditures (kcal/day) were calculated from activity patterns including basal metabolic rate, physical activity for 24 h [10], and specific food dynamic action. Basal metabolic rate for each subject was calculated with the Harris–Benedict equation [11].
Anthropometry and blood pressure analysis
Body weight and height of unclothed subjects without shoes were measured in the morning for calculating body mass index (BMI, kg/m2). Waist circumference was measured on standing subjects at the umbilical level after normal expiration. Blood pressure (BP) of seated subjects after a 20-min rest was measured in the left arm with an automatic BP monitor (FT-200S, Jawon Medical, Gyeongsan, Korea). After a 12-h fasting period, venous blood specimens were collected in EDTA-treated and plain tubes and centrifuged to yield plasma or serum, respectively, which were stored at −70 °C until analysis.
Clinical measurements
Fasting total cholesterol and triglyceride levels were analyzed using a Hitachi 7600 Autoanalyzer (Hitachi Ltd., Tokyo, Japan). ApoB-containing lipoproteins were precipitated with dextran-magnesium sulfate, and high density lipoprotein (HDL)-cholesterol concentrations in patient serum samples were measured enzymatically. For subjects with serum triglyceride levels <400 mg/dL, low density lipoprotein (LDL)-cholesterol concentrations were estimated indirectly using the Friedewald formula. For subjects with serum triglyceride levels ≥400 mg/dL, LDL-cholesterol concentrations were measured directly. Free fatty acids (FFA) were analyzed using the acyl-CoA synthetase − acyl-CoA oxidase enzymatic assay method and a Hitachi 7600 Autoanalyzer.
All subjects underwent an oral glucose-tolerance test at 0 and 12 weeks by ingesting a 75 g glucose solution after a 12-h overnight fast. Venous specimens were collected before glucose loading, at loading, and 30, 60, and 120-min after loading to determine serum glucose levels and responses. Fasting glucose levels were analyzed by the hexokinase method using a Hitachi 7600 Autoanalyzer. Insulin levels were measured using an immunoradiometric assay kit from DIAsource ImmunoAssays S.A. (Louvain, Belgium). Hemoglobin A1c (HbA1c) was measured by immonoturbidimetric analysis. Insulin resistance (IR) was calculated by the homeostasis-model assessment (HOMA) [12].
Whole blood was mixed with the same volume of RPMI 1640 (Gibco, Life Technologies, Gland Island, NY) and gently laid on a histopaque-1077 (Sigma-Aldrich, St. Louis, MO). The sample was then centrifuged at 1800 rpm for 20-min at 10 °C. After the separation, a thin layer of PBMCs was isolated and washed twice with RPMI 1640. The pellet was resuspended in RPMI 1640 with streptomycin. Isolated PBMCs were cultured in RPMI 1640 supplemented with 10 % fetal bovine serum (FBS), seeded in 12-well plates (1x106 cells/mL; SPL, Gyeonggi-do, Korea), and incubated at 37 °C with 5 % CO2 for 22.5 h. After a 22.5 h incubation, 10 % FBS was added and incubated for 24.5 h. At third day, PBMC supernatants were collected and stored at −80 °C until Lp-PLA2 activity levels were assayed. Lp-PLA2 activity in plasma and PBMC supernatants was measured by using a modification of a previously described high-throughput radiometric activity assay [13].
Serum high-sensitivity C-reactive protein (hs-CRP) was measured with an ADVIA 2400 Clinical Chemistry System (Siemens Ltd., Tarrytown, NY) using a commercially available, hs-CRP-Latex(II) X2 kit (Denka-Seiken Co., Ltd., Tokyo, Japan). Plasma malondialdehyde (MDA) was measured from thiobarbituric acid − reactive substances (TBARS) using the TBARS Assay Kit (ZeptoMetrix Co., Buffalo, NY). LDL particles were isolated by sequential flotation ultracentrifugation, and particle size distribution (1.019–1.063 g/mL) was examined using a pore-gradient lipoprotein system (CBS Scientific Company, San Diego, CA) on commercially available, non-denaturing gels containing a linear 2–16 % acrylamide gradient (CBS Scientific Company). Latex-bead (30 nm) conjugated thyroglobulin (17 nm), ferritin (12.2 nm), and catalase (10.4 nm) standards were used to estimate the relative band migration rates. Gels were scanned using a GS-800 Calibrated Imaging Densitometer (Bio-Rad Laboratories, Hercules, CA). Plasma oxidized (ox)-LDL was measured using an enzyme immunoassay (Mercodia AB, Uppsala, Sweden), and the resulting color reaction was determined at 450 nm on a Wallac Victor2 multilabel counter (Perkin-Elmer Life Sciences, Boston, MA).
Global (nontargeted) metabolic profiling of PBMC and plasma
PBMC and plasma extract sample preparation
Before analysis, 800 μL of 80 % acetonitrile was added to 100 μL of PBMC and plasma, mixed by vortexing, and centrifuged at 10,000 rpm for 5-min at 4 °C. The supernatant was dried with N2 (l), dissolved in 10 % methanol, mixed by vortexing, and centrifuged at 10,000 rpm for 5-min at 4 °C. The supernatant was transferred into a vial.
Ultra performance liquid chromatography
PBMC and Plasma extract samples (4 μL) were injected into an Acquity UPLC-BEH-C18 column (2.1 × 50 mm, 1.7 μm; Waters, Milford, MA) that was coupled in-line with a UPLC-LTQ-Orbitrap XL (Thermo Fisher Scientific, Waltham, MA). The injected samples were equilibrated with water containing 0.1 % formic acid. Samples were eluted with an acetonitrile gradient containing 0.1 % formic acid at a flow rate of 0.35 mL/min for 20-min. Metabolites were separated by UPLC, analyzed, and assigned by LTQ-Orbitrap-XL. The mass spectrometer (MS) was operated in ESI-positive mode. The spray voltage was 5 kV. The flow-rate nitrogen sheath gas and the auxiliary gas were 50 and 5 (arbitrary units). The capillary voltage (V), tube-lens voltage (V), and capillary temperature (°C) were kept constant at 35 V, 80 V, and 370 °C. Orbitrap data were collected in the range of m/z 50–1,000. MS/MS spectra of metabolites were obtained by a collision-energy ramp from 55–65 eV, and conducted with Xcalibur 2.1 and MS Frontier software (Thermo Fisher Scientific).
Data processing and identification of metabolites
All MS data including retention times, m/z, and ion intensities were extracted by SIEVE software (Thermo Fisher Scientific) incorporated into the instrument, and the resulting MS data were assembled into a matrix. SIEVE parameters were set as follows: m/z range 50–1,000; m/z width 0.02; retention time width 2.5; and m/z tolerance 0.005. Metabolites were searched using the following databases: ChemSpider (www.chemspider.com), Human Metabolome (www.hmdb.ca), Lipid MAPS (www.lipidmaps.org), KEGG (www.genome.jp/kegg), and MassBank (www.massbank.jp). Selected metabolites were confirmed by retention times and mass spectra of standard samples.
Statistical analyses
Statistical analyses were performed using SPSS v. 21.0 (IBM SPSS Statistics 21, Chicago, IL). Skewed variables were logarithmically transformed for statistical analyses. A two-tailed P-value of <0.05 was considered statistically significant. Differences in biochemical variables between two groups at baseline and follow-up were tested using Student’s independent t-test. General linear model tests were applied to compare parameter changes between the two groups by adjusting for baseline values. Paired t-tests were used to evaluate differences between baseline and follow-up levels in each group. Pearson’s and partial correlation coefficients were used to examine the relationships between variables over time. False discovery rate − corrected q-values were computed using the R package ‘fdrtool’. Heat map was created to visualize and evaluate correlations among metabolites and conventional risk factors in study populations.
Multivariate statistical analysis was performed using SIMCA-P+ software version 12.0 (Umetrics, Umeå, Sweden). Partial least-squares discriminant analysis (PLS-DA) was used as the classification method for modeling the discrimination between groups by visualizing the score scatter plot or S-plot using the first and second PLS components. The goodness-of-fit was quantified by R 2 Y, whereas the predictive ability was quantified by Q 2 Y. Generally, R 2 Y describes how well the data in the training set were mathematically reproduced and varied between 0 and 1 (a value of 1 indicated a model with a perfect fit). Models with Q 2 Y ≥0.5 were considered to have good predictive capabilities.
Results
Clinical characteristics, lipid profiles, and nutrient intake
There were no significant differences between two groups in baseline characteristics including age, gender, smoking, and drinking (data not shown). At baseline, there were no significant differences between two groups in BMI, waist:hip ratio (WHR), systolic BP, diastolic BP, serum triglyceride, total cholesterol, LDL-cholesterol, HDL-cholesterol, FFA, and hs-CRP. BMI, WHR, BP, serum lipid profiles, hs-CRP, total energy expenditure, and total energy intake were similar before and after the study in both groups (data not shown).
Replacement with whole grains and legumes caused significant increase in percent energy intake of protein and fat, and significant decrease in percent energy intake of carbohydrate. The percent energy intake of protein, fat, and carbohydrate significantly differed between the two groups before adjusting for baseline values. The whole-grain group had significant increases in fiber intake and polyunsaturated-to-saturated fatty acids ratio compared with baseline values. After 12-week, the whole-grain group had lower percent energy of carbohydrate, higher percent calorie of protein and fat, and fiber intake than control group (Table 1).
Fasting glucose, insulin, and malondialdehyde
At the end of the study, glucose, HbA1c, and MDA concentrations significantly increased in the control group, whereas glucose, glucose AUC (area under the curve), HbA1c, HOMA-IR, insulin, and MDA significantly decreased in the whole-grain group (Table 1). Changes in glucose, glucose AUC, HbA1c, HOMA-IR, and MDA in the whole-grain group were significantly different from those in the control group before and after adjusting for baseline levels. Post-treatment glucose, HOMA-IR, and MDA in the whole-grain group were significantly lower than those in the control group (Table 1).
Plasma ox-LDL, LDL particle size, Lp-PLA2 activity in plasma and unstimulated PBMC
At the end of the study, the whole-grain group had lower ox-LDL and Lp-PLA2 activity in PBMC and larger LDL particle size, whereas the control group had higher Lp-PLA2 activity in PBMC (Table 1). These changes in ox-LDL, LDL particle size, plasma Lp-PLA2 activity, and PBMC Lp-PLA2 activity in the whole-grain group were significantly different from those in the control group before and after adjusting for baseline levels. The post-treatment whole-grain group had lower ox-LDL and PBMC Lp-PLA2 activity, and larger LDL particle size than control group (Table 1).
Metabolic profiling of PBMC and plasma using UPLC-LTQ-orbitrap MS
Nontargeted metabolic pattern analysis
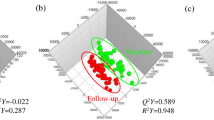
MS data of PBMC and plasma metabolites obtained at baseline and follow-up were analyzed with PLS-DA score scatter plot for the following two combinations: 1) control and whole-grain groups at baseline, control group at follow-up, and whole-grain group at follow-up (Fig. 1a, PBMC; Fig. 1c, plasma); and 2) control and whole-grain groups at follow-up (Fig. 1b, PBMC; Fig. 1d, plasma). The PBMC metabolite PLS-DA score scatter plot showed distinct clustering and clear separation for the following subjects: control and whole-grain groups at baseline, control group at follow-up, and whole-grain group at follow-up [R 2 X(cum) = 0.124, R 2 Y(cum) = 0.34, Q 2 Y(cum) = 0.218] (Fig. 1a). These distinct clusters indicate that PBMC profiling detects metabolic changes induced by dietary intervention. The PBMC metabolite PLS-DA score scatter plot showed distinct clustering for control and whole-grain groups at follow-up [R 2 X(cum) = 0.125, R 2 Y(cum) = 0.809, Q 2 Y(cum) = 0.525] (Fig. 1b).

Identification of PBMC and plasma metabolites that were significantly altered at 12-week follow-up. a Partial least squares discriminant analysis score scatter plot (PLS-DA score scatter plot) of PBMC metabolites at baseline (n = 80), whole-grain group at follow-up (n = 40), and control group at follow-up (n = 40). b PLS-DA score scatter plot of PBMC metabolites for whole-grain group at follow-up (n = 40) and control group at follow-up (n = 40). c PLS-DA PLS-DA score scatter plot of plasma metabolites at baseline (n = 80), whole-grain group at follow-up (n = 40), and control group at follow-up (n = 40). d PLS-DA score scatter plot of plasma metabolites for whole-grain group at follow-up (n = 40) and control group at follow-up (n = 40)
The plasma metabolite PLS-DA score scatter plot were not as clearly clustered as those for PBMC metabolites [R 2 X(cum) = 0.124, R 2 Y(cum) = 0.376, Q 2 Y(cum) = 0.127] (Fig. 1c). For control and whole-grain groups at follow-up, the plasma metabolite PLS-DA score scatter plot were not as clearly clustered as those for PBMC metabolites [R 2 X(cum) = 0.201, R 2 Y(cum) = 0.704, Q 2 Y(cum) = 0.414] (Fig. 1d). To identify metabolites that differentially determined data at baseline and follow-up, S-plots of p(1) and p(corr) (1) were generated using centroid scaling. The S-plots revealed that metabolites with higher or lower p(corr) values more clearly discriminated between the two groups.
Identification of PBMC metabolites
Of 1,923 PBMC metabolites, those that correlated with separation between the groups were identified by the variable important in the projection (VIP) parameter; VIP values >1.0 were highly relevant for group differences. 51 metabolites had VIP >1.0; 10 of these were previously identified and 41 were unknown. Those 10 PBMC metabolites at baseline and follow-up are shown in Table 2. There were no significant differences in baseline metabolites between two groups. After follow-up, the control group showed significant changes in six PBMC metabolite levels, whereas the whole-grain group showed significant changes in seven PBMC metabolite levels (Table 2).
We compared PBMC metabolite changes between two groups. The whole-grain group had greater reductions in L-leucine (q = 0.031), oleamide (q = 0.032), lysoPC (16:0) (q = 0.003), and lysoPC (18:0) (q = 0.003) (Table 2). At follow-up, the whole-grain group had higher peak intensities of L-pyroglutamic acid and ribothymidine, and lower peak intensities of palmitic amide, oleamide, and lysoPCs, compared with those of the control group (Table 2).
Identification of plasma metabolites
Of 4,121 plasma metabolites, those that correlated with separation between the groups were selected by VIP >1.0. 122 plasma metabolites were selected; 20 were previously identified and 102 were unknown (Table 3). There were no significant differences in baseline between two groups. After follow-up, C17 sphinganine significantly increased in the control group, also, there were no significant differences in metabolites between two groups, and no significant differences in metabolite changes with respect to baseline (Table 3).
Correlations among fasting glucose, plasma and PBMC Lp-PLA2 activities, biochemical parameters, and major PBMC metabolites
The correlation matrix of changes in glucose, Lp-PLA2 activities in plasma and PBMC, biochemical parameters, and major PBMC metabolites was computed (Fig. 2). Analysis of metabolic changes including all subjects identified the following correlations: glucose correlated positively with insulin, HOMA-IR, plasma Lp-PLA2 activity (r = 0.454, P < 0.001), MDA, ox-LDL, Lp-PLA2 in PBMC (r = 0.511, P < 0.001), glucose AUC, C-peptide, HbA1c, and PBMC lysoPCs after adjusting for age, gender, BMI, smoking, and drinking. After adjusting for confounding variables, plasma Lp-PLA2 activity correlated positively with glucose, HOMA-IR, ox-LDL, Lp-PLA2 activity in PBMC (r = 0.516, P < 0.001), glucose AUC, PBMC palmitic amide, and PBMC oleamide. After adjusting for confounding variables, PBMC Lp-PLA2 activity correlated positively with glucose, HOMA-IR, plasma Lp-PLA2 activity, ox-LDL, PBMC L-leucine, PBMC oleamide, PBMC lysoPCs, and correlated negatively with LDL particle size (Fig. 2).

Correlation matrix for changes in biochemical characteristics and PBMC metabolites in total subjects. Supervised hierarchical clustering identifies the most important 10 metabolites and 9 biochemical characteristics. Correlations were obtained by deriving Spearman correlation coefficients. Red, positive correlation. Purple, negative correlation
Discussion
We identified four PBMC metabolites that had statistically significant differences after dietary intervention, including L-leucine, oleamide, lysoPC (16:0), and lysoPC (18:0); however, there were no significant differences in plasma metabolites after dietary intervention. These aspects of results were also shown in both subjects with prediabetes and T2D, respectively. PBMCs may be a useful tool for nutrigenomics and understanding the pathophysiology of chronic disease due to their active metabolism [14, 15]. These results identify PBMC metabolites as powerful metabolomics tools to detect diet-induced metabolic changes.
Improving glycemic control in the whole-grain group decreases ox-LDL, and reduces PBMC Lp-PLA2 activity and PBMC lysoPCs. A strong correlation between PBMC Lp-PLA2 activity and ox-LDL, but not LDL-cholesterol, is consistent with a previous report of a direct effect of ox-LDL on Lp-PLA2 expression in THP-1 monocytes [16]. Ox-LDL may upregulate PBMC Lp-PLA2 expression in smokers [17]. Ox-phospholipids in LDL particles are hydrolyzed by Lp-PLA2 at the sn-2 position to produce bioactive ox-FFAs and lysoPCs. Only 1 − 5 % of the total non-ox-LDL PC content is lysoPC; however, up to 40 − 50 % of LDL PC is converted to lysoPC during LDL oxidation [18]. This study identified strongly positive correlations among ox-LDL, PBMC Lp-PLA2 activity, PBMC lysoPCs, which may indicate that ox-LDL and PBMC Lp-PLA2 activity are major determinants of PBMC lysoPC levels.
A negative correlation between PBMC Lp-PLA2 activities, PBMC lysoPCs with LDL particle size is consistent with a previous report of Lp-PLA2 binding preference for small dense LDL [19]. This study and other work [6] reported positive correlations among glucose, PBMC Lp-PLA2 activity, and plasma Lp-PLA2 activity. A strongly positive correlation between PBMC and plasma Lp-PLA2 activities also is observed in healthy subjects [17, 20]. In a porcine diabetes model, PBMC Lp-PLA2 expression is upregulated in the presence of glycation end products [21]. Increases in circulating Lp-PLA2 activity and increased ox-LDL levels in hypercholesterolemic pigs are primarily due to plaque macrophages [22]. These results indicate that the primary sources of plasma Lp-PLA2 are plaque macrophages [22] and PBMC [17]. This could explain our observations of lower plasma Lp-PLA2 activity changes compared with that of PBMC Lp-PLA2 activity, and no significant differences in plasma metabolite changes between two groups.
Reduced Lp-PLA2 activity in plasma and PBMC in the whole-grain group could be a marker of metabolic changes induced by increased consumption of protein relative to carbohydrate. Diet composition is an important factor in inflammatory processes of blood cells [23]. A study of macronutrient composition determined that increasing dietary protein from 19 % energy intake to 30 % yielded immediate and persistent downregulation of immunological genes in PBMCs [9]. Replacing 5 % of energy from carbohydrates with energy from protein and measured a 2.2 nmol/min/mL reduction in Lp-PLA2 activity that was independent of other changes in lipid profiles [5]. Our study replaced 7 % of energy from carbohydrate with approximately 4 % energy from protein and 3 % energy from fat.
Whole grains, legumes, and vegetables contain many antioxidants, vitamins, minerals, and phytochemicals [24, 25]. Antioxidants slow the oxidation rate of reduced substrates [24, 26]. Soybean phytochemicals reduce lipid peroxidation in vivo and attenuate LDL oxidation [27]. Our observed changes in glucose and HOMA-IR strongly correlated with changes in MDA and ox-LDL in patients with prediabetes or T2D, consistent with a previous report [28]. We observed positive correlation between changes in HOMA-IR and PBMC L-leucine, but not plasma L-leucine. This may be due to a negligible effect of PBMC L-leucine on plasma L-leucine, or the 12-week dietary intervention may not be long enough to change plasma L-leucine. The whole-grain group also had greater reduction in PBMC oleamide, but not in plasma, compared with control group. We identified PBMC oleamide (VIP = 22.5845) as the most important metabolite for evaluating differences between two groups at the end of the study. Recently, Ha et al. [29] identified plasma oleamide as the most important metabolite for distinguishing nondiabetic from diabetic males. Therefore, positive correlations between changes in PBMC oleamide, Lp-PLA2, PBMC palmitic amide, and PBMC lysoPCs observed in our study could be partly due to dietary-induced effects on blood cell inflammatory processes [23].
This study detected many metabolic markers using UPLC-LTQ-Orbitrap MS, but most are currently unidentified. Endogenous biomolecule databases for use with LC-MS − based metabolomics research are still under construction [30]. Despite this limitation, UPLC-LTQ-Orbitrap MS metabolomics and multivariate data analysis identified greater reductions in PBMC L-leucine, PBMC oleamide, PBMC lysoPCs in the whole-grain group than control group; however, there were no significant differences in plasma metabolites between two groups.
Conclusion
This study demonstrates that replacing refined rice with whole grains and legumes induced greater differences in PBMC Lp-PLA2 activity and metabolites than in plasma metabolites in nonobese patients with prediabetes or newly-diagnosed T2D. Therefore, consumption of minimally refined grains, legumes, and vegetables should be recommended to control glucose metabolism and reduce cardiovascular risk factors in patients with IFG, IGT, or newly-diagnosed T2D.
Abbreviations
- Lp-PLA2 :
-
Lipoprotein-associated phospholipase A2
- T2D:
-
Type-2 diabetes
- PBMC:
-
Peripheral blood mononuclear cell
- IFG:
-
Impaired fasting glucose
- IGT:
-
Impaired glucose tolerance
- BMI:
-
Body mass index
- BP:
-
Blood pressure
- HDL:
-
High density lipoprotein
- LDL:
-
Low density lipoprotein
- FFA:
-
Free fatty acid
- HbA1c :
-
Hemoglobin A1c
- IR:
-
Insulin resistance
- HOMA:
-
Homeostasis-model assessment
- FBS:
-
Fetal bovine serum
- hs-CRP:
-
High-sensitivity C-reactive protein
- MDA:
-
Malondialdehyde
- TBARS:
-
Thiobarbituric acid-reactive substances
- Ox:
-
Oxidized
- MS:
-
Mass spectrometer
- PLS-DA:
-
Partial least-squares discriminant analysis
- WHR:
-
Waist-hip ratio
- AUC:
-
Area under the curve
- VIP:
-
Variable important in the projection
References
Statistics Korea: Causes of death statistics in 2013. Available from: http://kostat.go.kr (updated 23 Sep 2014).
The Expert Committee on the Diagnosis and Classification of Diabetes Mellitus. Report of the expert committee on the diagnosis and classification of diabetes mellitus. Diabetes Care. 1997;20:1183–97.
Alberti KG, Zimmet PZ. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet Med. 1998;15:539–53.
Nelson TL, Biggs ML, Kizer JR, Cushman M, Hokanson JE, Furberg CD, et al. Lipoprotein-associated phospholipase A2 (Lp-PLA2) and future risk of type 2 diabetes: results from the cardiovascular health study. J Clin Endocrinol Metab. 2012;97:1695–701.
Hatoum IJ, Nelson JJ, Cook NR, Hu FB, Rimm EB. Dietary, lifestyle, and clinical predictors of lipoprotein-associated phospholipase A2 activity in individuals without coronary artery disease. Am J Clin Nutr. 2010;91:786–93.
Kim M, Jeung SR, Jeong TS, Lee SH, Lee JH. Replacing with whole grains and legumes reduces Lp-PLA2 activities in plasma and PBMCs in patients with prediabetes or T2D. J Lipid Res. 2014;55:1762–71.
Dentan C, Lesnik P, Chapman MJ, Ninio E. Phagocytic activation induces formation of platelet-activating factor in human monocyte-derived macrophages and in macrophage-derived foam cells. Relevance to the inflammatory reaction in atherogenesis. Eur J Biochem. 1996;236:48–55.
Yang EH, McConnell JP, Lennon RJ, Barsness GW, Pumper G, Hartman SJ, et al. Lipoprotein-associated phospholipase A2 is an independent marker for coronary endothelial dysfunction in humans. Arterioscler Thromb Vasc Biol. 2006;26:106–11.
Brattbakk HR, Arbo I, Aagaard S, Lindseth I, de Soysa AK, Langaas M, et al. Balanced caloric macronutrient composition downregulates immunological gene expression in human blood cells-adipose tissue diverges. OMICS. 2013;17:41–52.
Christian JL, Greger JL. Energy sources and uses. In: Christian JL, Greger JL, editors. Nutrition for living. Redwood City: Benjamin/Cummings Publishing Co.; 1994. p. 242–66.
Butte NF, Caballero B. Energy needs: assessment and requirements. In: Shils ME, Shike M, Ross AC, Caballero B, Cousins RJ, editors. Modern nutrition in health and disease. Philadelphia: Lippincott Williams & Wilkins; 2006. p. 136–48.
Matthews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher DF, Turner RC. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. 1985;28:412–9.
Wilensky RL, Shi Y, Mohler 3rd ER, Hamamdzic D, Burgert ME, Li J, et al. Inhibition of lipoprotein-associated phospholipase A2 reduces complex coronary atherosclerotic plaque development. Nat Med. 2008;14:1059–66.
Crujeiras AB, Parra D, Milagro FI, Goyenechea E, Larrarte E, Margareto J, et al. Differential expression of oxidative stress and inflammation related genes in peripheral blood mononuclear cells in response to a low-calorie diet: a nutrigenomics study. OMICS. 2008;12:251–61.
Kussmann M, Raymond F, Affolter M. OMICS-driven biomarker discovery in nutrition and health. J Biotechnol. 2006;124:758–87.
Wang WY, Li J, Yang D, Xu W, Zha RP, Wang YP. OxLDL stimulated lipoprotein-associated phopholipase A2 expression in THP-1 monocytes via PI3K and p38 MAPK pathways. Cardiovasc Res. 2010;85:845–52.
Fratta Pasini A, Stranieri C, Pasini A, Vallerio P, Mozzini C, Solani E, et al. Lysophosphatidylcholine and carotid intima-media thickness in young smokers: a role for oxidized LDL-induced expression of PBMC lipoprotein-associated phospholiase A2? PLoS One. 2013;8, e83092.
Matsumono T, Konayashi T, Kamata K. Role of lysophosphatidyl-choline (LPC) in atherosclerosis. Curr Med Chem. 2007;14:3209–20.
McNeill AM, Rosamond WD, Girman CJ, Golden SH, Schmidt MI, East HE, et al. The metabolic syndrome and 11-year risk of incident cardiovascular disease in the atherosclerosis risk in communities study. Diabetes Care. 2005;28:385–90.
Paik JK, Kim JY, Kim OY, Lee Y, Jeong TS, Sweeney G, et al. Circulating and PBMC Lp-PLA2 associate differently with oxidative stress and subclinical inflammation in nonobese women (menopausal status). PLoS One. 2012;7, e29675.
Shi Y, Zhang P, Zhang L, Osman H, Mohler 3rd ER, Macphee C, et al. Role of lipoprotein-associated phospholipase A2 in leukocyte activation and inflammatory responses. Atherosclerosis. 2007;191:54–62.
De Keyzer D, Karabina SA, Wei W, Geeraert B, Stengel D, Marsillach J, et al. Increased PAFAH and oxidized lipids are associated with inflammation and atherosclerosis in hypercholesterolemic pigs. Arterioscler Thrombo Vasc Biol. 2009;29:2041–6.
Munoz A, Costa M. Nutritionally mediated oxidative stress and inflammation. Oxid Med Cell Longev. 2013;2013:610950.
Belobrajdic DP, Bird AR. The potential role of phytochemicals in wholegrain cereals for the prevention of type-2 diabetes. Nutr J. 2013;12:62–74.
Liu S, Manson JE, Lee IM, Cole SR, Hennekens CH, Willett WC, et al. Fruit and vegetable intake and risk of cardiovascular disease: the Women’s Health Study. Am J Clin Nutr. 2000;72:922–8.
Jacobs Jr DR, Andersen LF, Blomhoff R. Whole-grain consumption is associated with a reduced risk of noncardiovascular, noncancer death attributed to inflammatory disease in the lowa Women’s Health Study. Am J Clin Nutr. 2007;85:1606–14.
Crujeiras AB, Parra D, Abete I, Martínez JA. A hypocaloric diet enriched in legumes specifically mitigates lipid peroxidation in obese subjects. Free Radic Res. 2007;41:498–506.
Chung HK, Chae JS, Hyun YJ, Paik JK, Kim JY, Jang Y, et al. Influence of adiponectin gene polymorphisms on adiponectin level and insulin resistance index in response to dietary intervention in overweight-obese patients with impaired fasting glucose or newly diagnosed type 2 diabetes. Diabetes Care. 2009;32:552–8.
Ha CY, Kim JY, Paik JK, Kim OY, Paik YH, Lee EJ, et al. The association of specific metabolites of lipid metabolism with markers of oxidative stress, inflammation and arterial stiffness in men with newly diagnosed type 2 diabetes. Clin Endocrinol. 2012;76:674–82.
Skoog T, Dichtl W, Boquist S, Skoglund-Andersson C, Karpe F, Tang R, et al. Plasma tumour necrosis factor-α and early carotid atherosclerosis in healthy middle-aged men. Eur Heart J. 2002;23:376–83.
Acknowledgments
This research was supported by the Bio-Synergy Research Projects (NRF-2012M3A9C4048762) of the National Research Foundation (NRF) funded by the Ministry of Science, ICT & Future Planning, Republic of Korea.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
All the authors were involved in the development of the study protocol and the experimental design. GS, MK and JHY collected samples and carried out experiments. MK performed data analysis. T-SJ, S-HL and JHL reviewed data and participated in general discussion. JHL provided the research funding and wrote the manuscript. All the authors read, commented on, and contributed to the submitted manuscript. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Kim, M., Song, G., Kang, M. et al. Replacing carbohydrate with protein and fat in prediabetes or type-2 diabetes: greater effect on metabolites in PBMC than plasma. Nutr Metab (Lond) 13, 3 (2016). https://doi.org/10.1186/s12986-016-0063-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12986-016-0063-4