
Abstract
The quest for an effective HIV-1 vaccine began as soon as the virus causing AIDS was identified. After several disappointing attempts, results of the Phase-III RV144 trial in Thailand were a beacon of hope for the field demonstrating correlation between protection and immunological markers. In order to optimize vaccine response, we underline results from yellow fever and hepatitis B vaccines, where protective responses were predicted by the pre-vaccination level of immune activation in healthy individuals. Such findings support the assessment and reduction of pre-vaccine immune activation in order to optimize vaccine response. Immune activation in healthy individuals can be influenced by age, presence of CMV infection, gut dysbiosis and microbial translocation. We speculate that the level of immune activation should therefore be assessed to better select participants in vaccine trials, and interventions to reduce inflammation should be used to increase protective HIV vaccine response.
Similar content being viewed by others

Background
Despite tremendous effort to develop a successful human immunodeficiency virus (HIV) vaccine, the quest for a safe and effective HIV vaccine seems to be remarkably long and winding. Most licensed vaccines against viral and bacterial infections demonstrate efficacy in inducing antibodies [1]. In addition to the induction of specific antibodies, vaccines also induce protective immunity by eliciting a combination of innate and acquired cellular responses [2]. However, specific immune responses following administration of licensed vaccines is not well-understood, but have recently been shown to be associated with pre-vaccine level of immune activation [3]. In contrast to the clearance of childhood viral infections like smallpox and measles, vaccine development against HIV remains challenging as the virus inexorably causes a chronic infection that cannot be naturally cleared by the host. Furthermore, there is little guidance in vaccine design when considering the determinants of host response, due to the absence of spontaneous clearance of HIV infection. Development of an HIV vaccine is even more difficult considering the unparalleled capacity of HIV to mutate its genome, allowing it to evade antibody recognition of its viral envelope [4], in addition to its ability to suppress the major conductor of the immune response, the helper CD4 T-cells [5]. The ideal HIV vaccine should provide protection against the broad genetic diversity of HIV, control CD4 T-cell activation and likely elicit long-term cytotoxic CD8 T-cell response to block both mucosal and parenteral transmission routes.
The humoral immune response is one of the first lines of defense against invading pathogens, which plays a crucial role in preventing infection. Vaccine efficacy can be predicted using a correlate of protection (CoP), which is a measurable immune biomarker that identifies immunity in the host. The antibody titre represents the most common CoP for licensed vaccines [6]. However, establishing immune determinants responsible for the protection remain difficult as several type of immune responses are intertwined [7]. Importantly, such immune responses may depend on type of vaccine, level of immune activation, micro-environment and host genetic factors. Very limited information exists on CoP in HIV as vaccine efficacy has been globally disappointing.
One of the most encouraging vaccine studies RV144, investigated the efficacy of a combined vaccine regimen in young adults in Thailand [8, 9]. The vaccination regimen consisted of ALVAC-HIV boosted with AIDSVAX® B/E, and demonstrated partial efficacy of 31% against HIV-1 infection. The level of protection was associated with the development of envelope specific antibodies [9, 10]. In addition, antibody-dependent cell-mediated cytotoxicity (ADCC), host genetics and virus diversity in HIV-1 were also correlates of protection. Such correlates are defined as vaccine-induced immune responses associated with the rate of HIV-1 infection among vaccine recipients. Measuring these forms of CoP can be labor intensive, costly, and not scalable, highlighting the need to identify surrogate markers that can simply and reliably predict the level of protection from HIV-1 infection.
Immune activation as newly identified correlate of protection: evidence from yellow fever and hepatitis B commercialized vaccines
Recent information generated from yellow fever and hepatitis B on CoP will be useful to further guide HIV vaccine development and implementation.
Yellow fever vaccine
In a 2014 study, Muyanja and colleagues sought to understand the modulation of vaccine immune responses towards better guidance in vaccine development against difficult targets like HIV or malaria [11]. As such, they evaluated patient-specific immune responses to the commercialized yellow fever vaccine, YF-17D. By comparing healthy individuals from Lausanne (Switzerland) and Entebbe (Uganda), the authors showed that YF-17D induced B cell responses, as well as a substantial reduction in CD8 T-cells in age-matched individuals living in Entebbe [11]. Furthermore, the authors identified higher frequencies of exhausted and activated NK cells, more differentiated B and T-cell subsets, and an abundance of proinflammatory monocytes, demonstrating an activated immune microenvironment in the participants from Entebbe. Baseline B and CD8 T-cell activation levels, coupled with proinflammatory monocytes, negatively predicted titres of YF-17D-neutralizing antibody post vaccination. In addition, B cell and memory T-cell responses measured before vaccination in the Entebbe did not last as long as in the Lausanne participants. Such findings in healthy individuals indicate that immune activation prior to vaccination influences the response of one of the most protective vaccines ever commercialized, and illustrates the importance of considering immune modulation to optimize HIV vaccine response.
The importance of immune modulation in vaccine response is supported by a recent study that investigated the impact of infection history in a mouse model. Since laboratory-raised mice are not typically exposed to common infections acquired by human children, these mice may not be the best model to predict immune responses in humans. Blood immune responses were measured before and after YFV-17D vaccination in mice that were sequentially infected with herpes viruses, influenza and a helminth [12]. The authors uncovered significant differences in gene expression patterns between mock and co-infected samples, both before and after vaccination. They also found that co-infected mice had a reduced antibody response, but had similar neutralizing antibody titers after vaccination. As a consequence of this work, ongoing clinical trials are assessing the effect of an anthelminthic treatment before yellow fever vaccination to enhance protective vaccine response.
Hepatitis B vaccine
To further elucidate the factors influencing CoP, the Sekaly group studied vaccine response in elderly participants following hepatitis B vaccination [3]. After measuring gene expression profiles according to age, they assessed pre-vaccination predictors of response to HBV vaccine in distinct sets of older adults. A stronger HBV response was observed for those having heightened expression of genes that augment B-cell responses and higher memory B-cells. Conversely, higher levels of inflammatory response transcripts and increased frequencies of pro-inflammatory innate cells were associated with a lower vaccine response. Again, this work identified the contribution of immune activation as a key baseline determinant of HBV vaccine response, which may also be relevant for future HIV vaccine development.
Other host and environmental factors that may influence CoP are also intertwined with immune activation. Host factors include immune exhaustion, senescence, metabolic reprogramming and tissue inflammation, contributing to T-cell dysfunction and modulation of B-cell response to neo-antigens or vaccines. Among the environmental factors, gut microbial composition, gut mucosal barrier damage and cytomegalovirus (CMV) co-infection are the most studied and differ in Northern and Southern hemispheres [13].
Relevance of immune activation to improve HIV vaccine response
As noted, the encouraging results of the Thai RV144 vaccine trial provided insight into future HIV vaccine development [9]. The investigators identified that IgG for the V1V2 region of Env gp120 was associated with protection from infection. However, the protection did not persist and was temporally linked with antibodies mediating antibody-dependent-cellular cytotoxicity (ADCC). Importantly, mounting evidence indicates that antibody mechanisms beyond neutralization may contribute to protection, as Fc characteristics and ADCC have been identified as CoP against HIV acquisition conversely to the induction of neutralizing antibodies and/or cytotoxic T-cells [8]. Such findings suggest that combinations of broad antibodies targeting different binding sites on the Env protein could block immune escape. More recent findings from this trial support correlation between T follicular help (Tfh) function relevant for B-cell help and envelope-specific antibody development. Tfh responses generated by RV144 show a marked inter-individual variation indicating the presence of determinants that remain largely unknown [14].
Observations of inter-individual variation highlight a knowledge gap in the role of human pre-immune repertoires in driving vaccine response. One such repertoire is the gut microbiota, as it can elicit influences that are indirect and can act at a distance. These mechanisms may involve cross-reactivity between microbial and vaccine antigens shaping T-cell repertoires, and/or microbial products stimulating pattern recognition receptors that influence the type and intensity of vaccine responses. Preliminary data show that T helper cells cross react with HIV antigens which are identified in uninfected healthy individuals and are usually missed by standard intracellular assay for the measurement of Th1 cytokines [15]. Such cross-reactive responses may shape subsequent responses to HIV vaccine candidates in both magnitude and function [16]. This is further illustrated by the HVTN 055 trial which showed a lack of protection against HIV, owing to the unexpected induction of non-neutralizing gp41-reactive anti-HIV antibodies [17]. The antibodies elicited by the vaccine were related to a polyreactive antibody repertoire response from pre-existing B-cells that were cross-reactive with Escherichia coli likely originating from the gut. Understanding of the contribution of the microbiome and/or microbial translocation to both vaccine-induced immunity and non-specific immune activation represents a new and unforeseen challenge for the development of an optimal vaccine response.
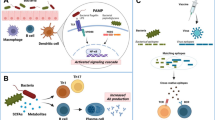
Gut-associated lymphoid tissue (GALT) represents a major site of immune response [18, 19]. This is due to the GALT that harbors a large fraction of activated CD4 T-cell populations, which are preferential targets for HIV replication. The interaction of GALT CD4-T cells with commensal/pathogenic microflora promotes cellular activation that favors immune activation. In the context of treated HIV-infection, the predictors of commercialized vaccine response are linked with the levels of T-cell activation and translocation of bacterial products such as LPS into the systemic circulation [20] (Fig. 1). LPS-triggered toll-like receptor (TLR) signaling induces development and activation in the B-cells in the germinal centres (GCs) thereby regulating GC formation and antibody production [21]. Such immune activation has also been characterized by the induction of B-cell attracting chemokine, CXCL13, which is recognized by the receptor CXCR5, and contributes to the follicular homing of B and T-cells. Unlike most healthy tissue, the normal intestinal lamina propria contains a large number of plasma cells containing IgA. Secretary IgA (sIgA) is almost completely dependent on the presence of microbiota which predominate the distal colon [22], which may influence the HIV vaccine specific IgA response [23] (Fig. 1).

Contribution of gut microbes and microbial translocation on immune response pertaining to HIV vaccine development
Gut microbiota, metabolites and vaccine response
Manipulation of microbiota composition and their metabolites via diet alteration or microbiota engraftment is under intense evaluation in cancer and autoimmune disorders and vaccinology. Learning from our gut endogenous original adjuvants and/or tolerogenic microbes and their metabolites will be critical in overcoming the HIV vaccine challenge [24]. We recently showed the contribution of dietary tryptophan (Trp), one of the essential amino acids mainly obtained from protein-rich foods, contributed to immune suppression by the production of an immunosuppressive catabolite in the context of HIV/HCV infections [25,26,27]. Other groups have identified a similar change in cancer, autism, and multiple sclerosis indicating the influence of kynurenine, a metabolite of Trp used in the production of niacin, as an immunosuppressor present in several chronic conditions [28, 29]. Of note, several Trp metabolites produced by bacteria in the gut are endogenous ligands for the transcription factor aryl hydrocarbon receptor (AhR) which has been recently shown to control B-cell fate decisions including suppression of class switching in vivo after influenza immunization [30]. AhR activation is linked to both diet and gut microbiota composition and leads to the local production of IL-22, a cytokine that plays an important role in maintaining mucosal immunity and integrity, mainly by innate lymphoid cells group 3 (ILC3) [31, 32]. Only specific subsets of bacteria, particularly Lactobacilli, an important member of Firmicutes, can metabolize dietary Trp into idole-3-aldehyde to modulate AhR/IL-22 axis, thus contributing to gut mucosal homeostasis [32, 33]. In addition, AhR activation occurring in macrophages and DCs further contributes to the local anti-inflammatory response.
Certain microbes like Taenia crassiceps, a parasite that causes a gut helminthic infection, have been shown to impair antibody response to pneumococcal vaccine in mice [34]. Similarly, Riner et al. showed that immunization against hepatitis B and tetanus toxoid in Kenyan adults having schistosomiasis, caused by members of the Schistosoma genus, resulted in a more rapid decline in antibody titres that can be prevented by a prior anti-helminthic treatment [35]. All these observations highlight the importance of gut microbial dysbiosis on distal immune response in physiology and disease [36].
CMV infection, aging and vaccine response
Human cytomegalovirus (CMV) establishes a latent infection that remains generally asymptomatic but can lead to serious illness in immune-suppressed individuals [37, 38]. The long-term control of CMV shapes the immune system contributing to a memory inflammation induced by CD8 T-cell specific response. Such CMV-associated low-grade inflammation also contributes to human aging and has been termed as “inflammaging” [39, 40]. CMV seropositivity has been shown to have a negative effect on influenza vaccine-specific antibody responses in the elderly as well as in the young [41]. CMV infection is prevalent in Africa and may contribute to a lower vaccine response observed in this population. Furthermore, CMV co-infection has been associated with faster disease progression and elevated CMV-specific IgG antibody levels in untreated HIV-infected individuals, further contributing to disease progression and immune activation [42]. For patients receiving long-term antiretroviral therapy (ART), CMV-co-infection remains an independent contributor to persistent CD8 T-Cell expansion and inflammation [43].
Conclusions
We are starting to appreciate the importance of pre-existing inflammation linked to gut microbial composition in vaccine response that should influence HIV vaccine development where the bulk of epidemic prevails in developing countries. The complex interplay between microbial composition and the gut epithelial barrier contributes to the state of systemic immune activation representing an emerging area to be considered to optimize HIV vaccine response. Modulation of chronic immune activation prior to vaccine administration may elicit persistent memory antibody response with CD8 T-cell cytotoxic function. Collaborative effort between microbiologists, immunologists epidemiologists and clinicians will be needed to foster HIV vaccine research.
References
Thakur A, Pedersen LE, Jungersen G. Immune markers and correlates of protection for vaccine induced immune responses. Vaccine. 2012;30(33):4907–20.
Pulendran B, Ahmed R. Immunological mechanisms of vaccination. Nat Immunol. 2011;12(6):509–17.
Fourati S, Cristescu R, Loboda A, Talla A, Filali A, Railkar R, Schaeffer AK, Favre D, Gagnon D, Peretz Y, et al. Pre-vaccination inflammation and B-cell signalling predict age-related hyporesponse to hepatitis B vaccination. Nat Commun. 2016;7:10369.
Lynch RM, Wong P, Tran L, O’Dell S, Nason MC, Li Y, Wu X, Mascola JR. HIV-1 fitness cost associated with escape from the VRC01 class of CD4 binding site neutralizing antibodies. J Virol. 2015;89(8):4201–13.
Routy JP, Mehraj V, Cao W. HIV immunotherapy comes of age: implications for prevention, treatment and cure. Expert Rev Clin Immunol. 2016;12(2):91–4.
Tomaras GD, Plotkin SA. Complex immune correlates of protection in HIV-1 vaccine efficacy trials. Immunol Rev. 2017;275(1):245–61.
Plotkin SA. Complex correlates of protection after vaccination. Clin Infect Dis. 2013;56(10):1458–65.
Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, et al. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N Engl J Med. 2012;366(14):1275–86.
Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N Engl J Med. 2009;361(23):2209–20.
Nicely NI, Wiehe K, Kepler TB, Jaeger FH, Dennison SM, Rerks-Ngarm S, Nitayaphan S, Pitisuttithum P, Kaewkungwal J, Robb ML, et al. Structural analysis of the unmutated ancestor of the HIV-1 envelope V2 region antibody CH58 isolated from an RV144 vaccine efficacy trial vaccinee. EBioMedicine. 2015;2(7):713–22.
Muyanja E, Ssemaganda A, Ngauv P, Cubas R, Perrin H, Srinivasan D, Canderan G, Lawson B, Kopycinski J, Graham AS, et al. Immune activation alters cellular and humoral responses to yellow fever 17D vaccine. J Clin Invest. 2014;124(7):3147–58.
Reese TA, Bi K, Kambal A, Filali-Mouhim A, Beura LK, Burger MC, Pulendran B, Sekaly RP, Jameson SC, Masopust D, et al. Sequential infection with common pathogens promotes human-like immune gene expression and altered vaccine response. Cell Host Microbe. 2016;19(5):713–9.
Qin L, Jing X, Qiu Z, Cao W, Jiao Y, Routy JP, Li T. Aging of immune system: immune signature from peripheral blood lymphocyte subsets in 1068 healthy adults. Aging (Albany NY). 2016;8(5):848–59.
Gottardo R, Bailer RT, Korber BT, Gnanakaran S, Phillips J, Shen X, Tomaras GD, Turk E, Imholte G, Eckler L, et al. Plasma IgG to linear epitopes in the V2 and V3 regions of HIV-1 gp120 correlate with a reduced risk of infection in the RV144 vaccine efficacy trial. PLoS ONE. 2013;8(9):e75665.
Ritchie AJ, Campion SL, Kopycinski J, Moodie Z, Wang ZM, Pandya K, Moore S, Liu MK, Brackenridge S, Kuldanek K, et al. Differences in HIV-specific T cell responses between HIV-exposed and -unexposed HIV-seronegative individuals. J Virol. 2011;85(7):3507–16.
Su LF, Kidd BA, Han A, Kotzin JJ, Davis MM. Virus-specific CD4(+) memory-phenotype T cells are abundant in unexposed adults. Immunity. 2013;38(2):373–83.
Williams WB, Liao HX, Moody MA, Kepler TB, Alam SM, Gao F, Wiehe K, Trama AM, Jones K, Zhang R, et al. HIV-1 VACCINES. Diversion of HIV-1 vaccine-induced immunity by gp41-microbiota cross-reactive antibodies. Science. 2015;349(6249):aab1253.
Goronzy JJ, Weyand CM. Successful and maladaptive T cell aging. Immunity. 2017;46(3):364–78.
Winstead CJ. Follicular helper T cell-mediated mucosal barrier maintenance. Immunol Lett. 2014;162(2 Pt A):39–47.
Paquin-Proulx D, Ching C, Vujkovic-Cvijin I, Fadrosh D, Loh L, Huang Y, Somsouk M, Lynch SV, Hunt PW, Nixon DF, et al. Bacteroides are associated with GALT iNKT cell function and reduction of microbial translocation in HIV-1 infection. Mucosal Immunol. 2017;10(1):69–78.
Hua Z, Hou B. TLR signaling in B-cell development and activation. Cell Mol Immunol. 2013;10(2):103–6.
Mowat AM, Agace WW. Regional specialization within the intestinal immune system. Nat Rev Immunol. 2014;14(10):667–85.
Nguyen QN, Himes JE, Martinez DR, Permar SR. The Impact of the gut microbiota on humoral immunity to pathogens and vaccination in early infancy. PLoS Pathog. 2016;12(12):e1005997.
Collins N, Belkaid Y. Do the microbiota influence vaccines and protective immunity to pathogens? Engaging our endogenous adjuvants. Cold Spring Harb Perspect Biol. 2017.
Jenabian MA, Patel M, Kema I, Kanagaratham C, Radzioch D, Thebault P, Lapointe R, Tremblay C, Gilmore N, Ancuta P, et al. Distinct tryptophan catabolism and Th17/Treg balance in HIV progressors and elite controllers. PLoS ONE. 2013;8(10):e78146.
Jenabian MA, Mehraj V, Costiniuk CT, Vyboh K, Kema I, Rollet K, Paulino Ramirez R, Klein MB, Routy JP. Influence of hepatitis C virus sustained virological response on immunosuppressive tryptophan catabolism in ART-treated HIV/HCV coinfected patients. J Acquir Immune Defic Syndr. 2016;71(3):254–62.
Mehraj V, Routy JP. Tryptophan catabolism in chronic viral infections: handling uninvited guests. Int J Tryptophan Res. 2015;8:41–8.
Mittal S. Targeting the broadly pathogenic kynurenine pathway. New York: Springer International Publishing; 2015.
Zitvogel L, Daillere R, Roberti MP, Routy B, Kroemer G. Anticancer effects of the microbiome and its products. Nat Rev Microbiol. 2017;15:465–78.
Vaidyanathan B, Chaudhry A, Yewdell WT, Angeletti D, Yen WF, Wheatley AK, Bradfield CA, McDermott AB, Yewdell JW, Rudensky AY, et al. The aryl hydrocarbon receptor controls cell-fate decisions in B cells. J Exp Med. 2017;214(1):197–208.
Qiu J, Guo X, Chen ZM, He L, Sonnenberg GF, Artis D, Fu YX, Zhou L. Group 3 innate lymphoid cells inhibit T-cell-mediated intestinal inflammation through aryl hydrocarbon receptor signaling and regulation of microflora. Immunity. 2013;39(2):386–99.
Zelante T, Iannitti RG, Cunha C, De Luca A, Giovannini G, Pieraccini G, Zecchi R, D’Angelo C, Massi-Benedetti C, Fallarino F, et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity. 2013;39(2):372–85.
Rooks MG, Garrett WS. Gut microbiota, metabolites and host immunity. Nat Rev Immunol. 2016;16(6):341–52.
Apiwattanakul N, Thomas PG, Iverson AR, McCullers JA. Chronic helminth infections impair pneumococcal vaccine responses. Vaccine. 2014;32(42):5405–10.
Riner DK, Ndombi EM, Carter JM, Omondi A, Kittur N, Kavere E, Korir HK, Flaherty B, Karanja D, Colley DG. Schistosoma mansoni infection can jeopardize the duration of protective levels of antibody responses to immunizations against hepatitis B and tetanus toxoid. PLoS Negl Trop Dis. 2016;10(12):e0005180.
Schroeder BO, Backhed F. Signals from the gut microbiota to distant organs in physiology and disease. Nat Med. 2016;22(10):1079–89.
Friel TJ. Epidemiology, clinical manifestations, and treatment of cytomegalovirus infection in immunocompetent adults. In: Hirsch MS, Thorner AR, editors. UpToDate. Waltham: Ted. W. Post. UpToDate; 2017.
Vancikova Z, Dvorak P. Cytomegalovirus infection in immunocompetent and immunocompromised individuals—a review. Curr Drug Targets Immune Endocr Metabol Disord. 2001;1(2):179–87.
Fehervari Z. Inflammasomes in human aging. Nat Immunol. 2017;18(3):254.
Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci. 2014;69(Suppl 1):S4–9.
Frasca D, Blomberg BB. Aging, cytomegalovirus (CMV) and influenza vaccine responses. Hum Vaccin Immunother. 2016;12(3):682–90.
Patel EU, Gianella S, Newell K, Tobian AA, Kirkpatrick AR, Nalugoda F, Grabowski MK, Gray RH, Serwadda D, Quinn TC, et al. Elevated cytomegalovirus IgG antibody levels are associated with HIV-1 disease progression and immune activation. AIDS. 2017;31(6):807–13.
Freeman ML, Mudd JC, Shive CL, Younes SA, Panigrahi S, Sieg SF, Lee SA, Hunt PW, Calabrese LH, Gianella S, et al. CD8 T-cell expansion and inflammation linked to CMV coinfection in ART-treated HIV infection. Clin Infect Dis. 2016;62(3):392–6.
Authors’ contributions
JPR and VM participated in the drafting of the manuscript. Both authors read and approved the final manuscript.
Acknowledgements
The authors thank Allison Sherratt for critical review and editing and Angie Massicotte for administrative assistance.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Not applicable.
Consent for publication
The authors give their consent for publication.
Ethics approval and consent to participate
Not applicable.
Funding
This work was supported by the Fonds de la Recherche Québec-Santé (FRQ-S): Réseau SIDA/Maladies infectieuses and Thérapie cellulaire; the Canadian Institutes of Health Research (CIHR; Grant MOP 103230); the Vaccines & Immunotherapies Core of the CIHR Canadian HIV Trials Network (CTN; Grant CTN 257); the Canadian Foundation for AIDS Research (CANFAR; Grant 02-512); and the Canadian HIV Cure Enterprise Team Grant (HIG-133050) awarded by the CIHR in partnership with CANFAR. VM is supported by an FRQ-S Postdoctoral Fellowship Award. J-PR is the holder of the Louis Lowenstein Chair in Hematology and Oncology, McGill University.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Routy, JP., Mehraj, V. Potential contribution of gut microbiota and systemic inflammation on HIV vaccine effectiveness and vaccine design. AIDS Res Ther 14, 48 (2017). https://doi.org/10.1186/s12981-017-0164-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12981-017-0164-9